The Production of Listeriolysin O and Subsequent Intracellular Infections by Listeria monocytogenes Are Regulated by Exogenous Short Chain Fatty Acid Mixtures

Abstract
:1. Introduction
2. Results
2.1. High but not Low Levels of SCFAs Reduced In Vitro Growth
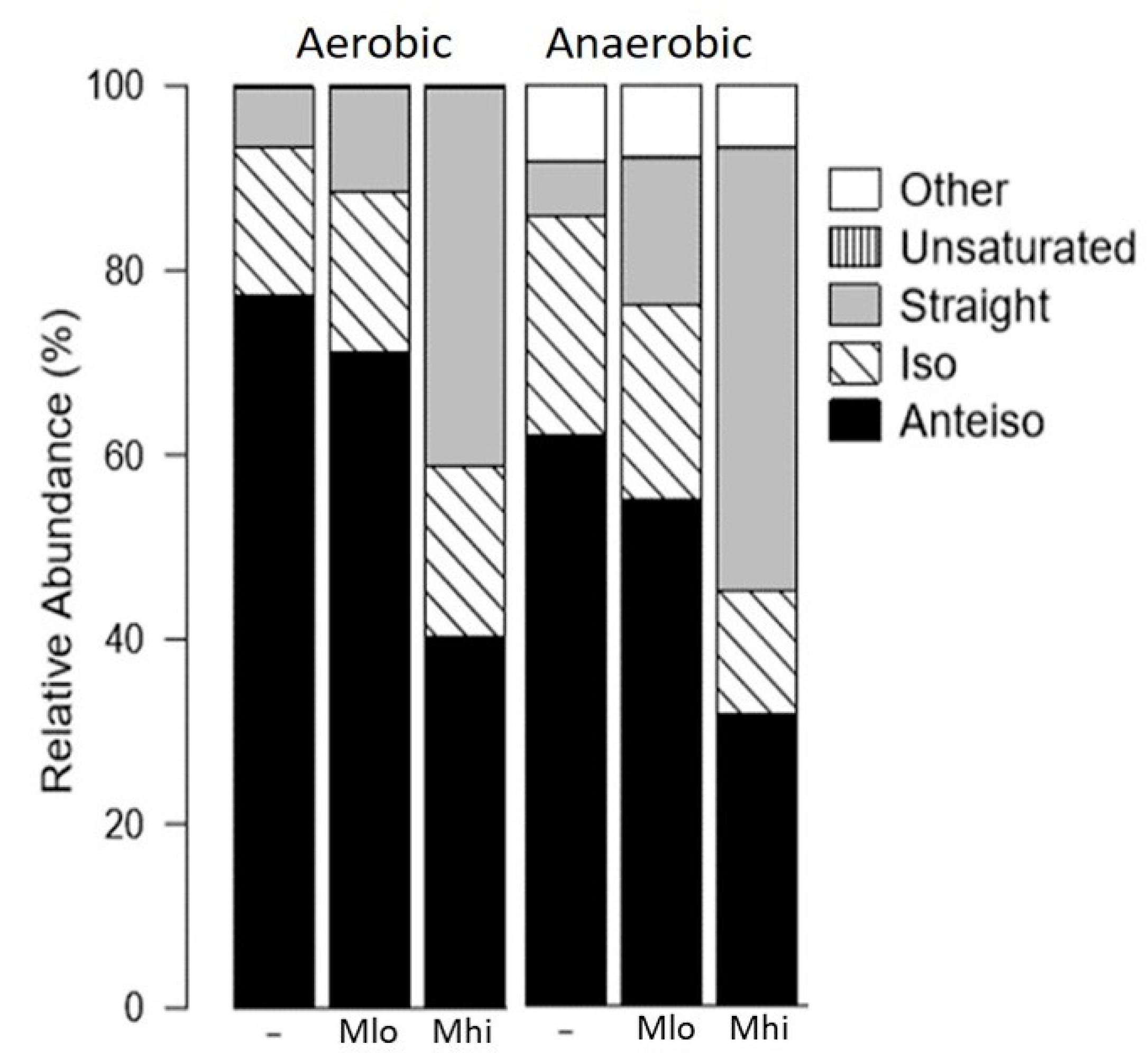
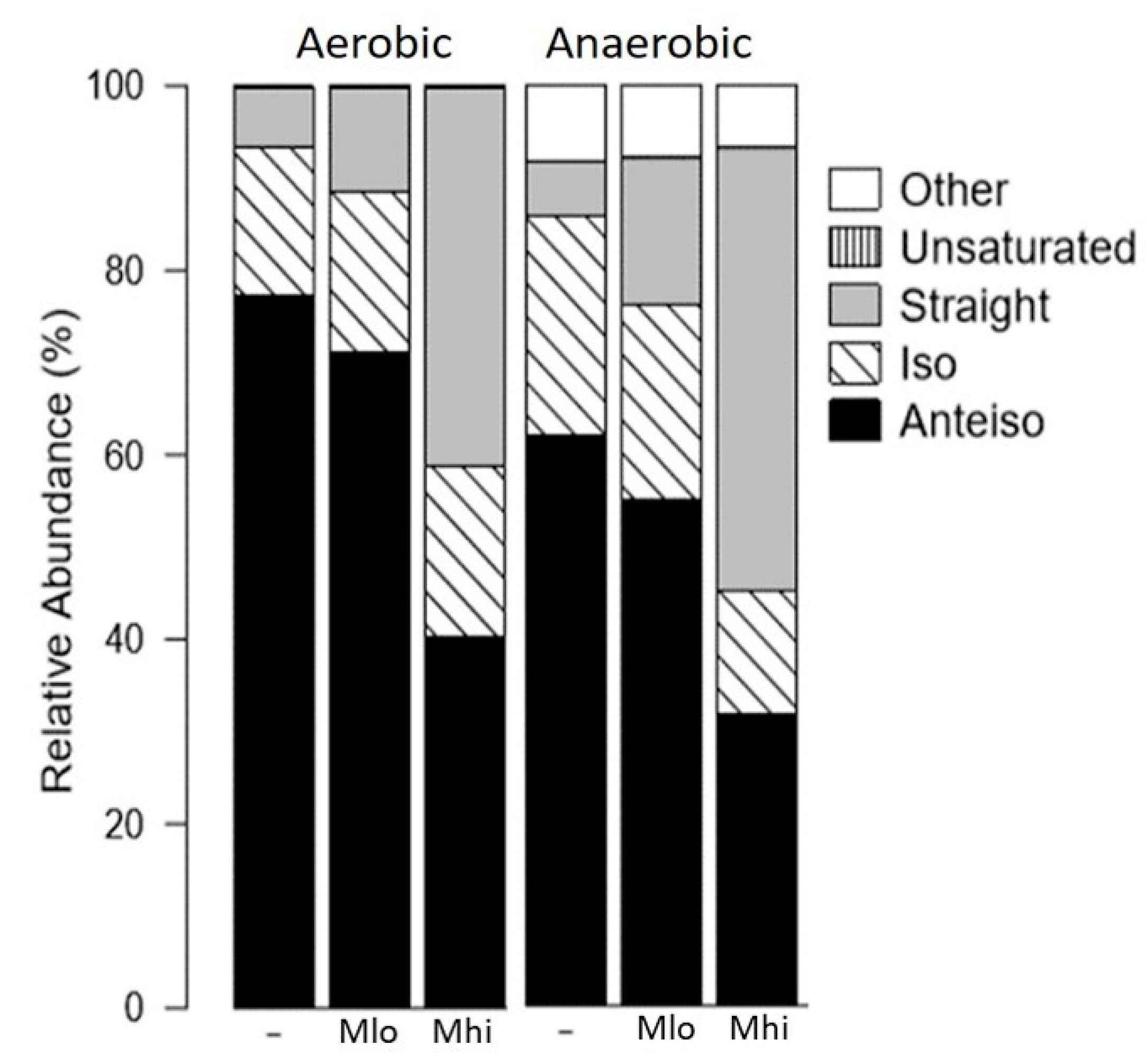
2.2. SCFA Supplementations Altered Membrane Fatty Acid Compositions
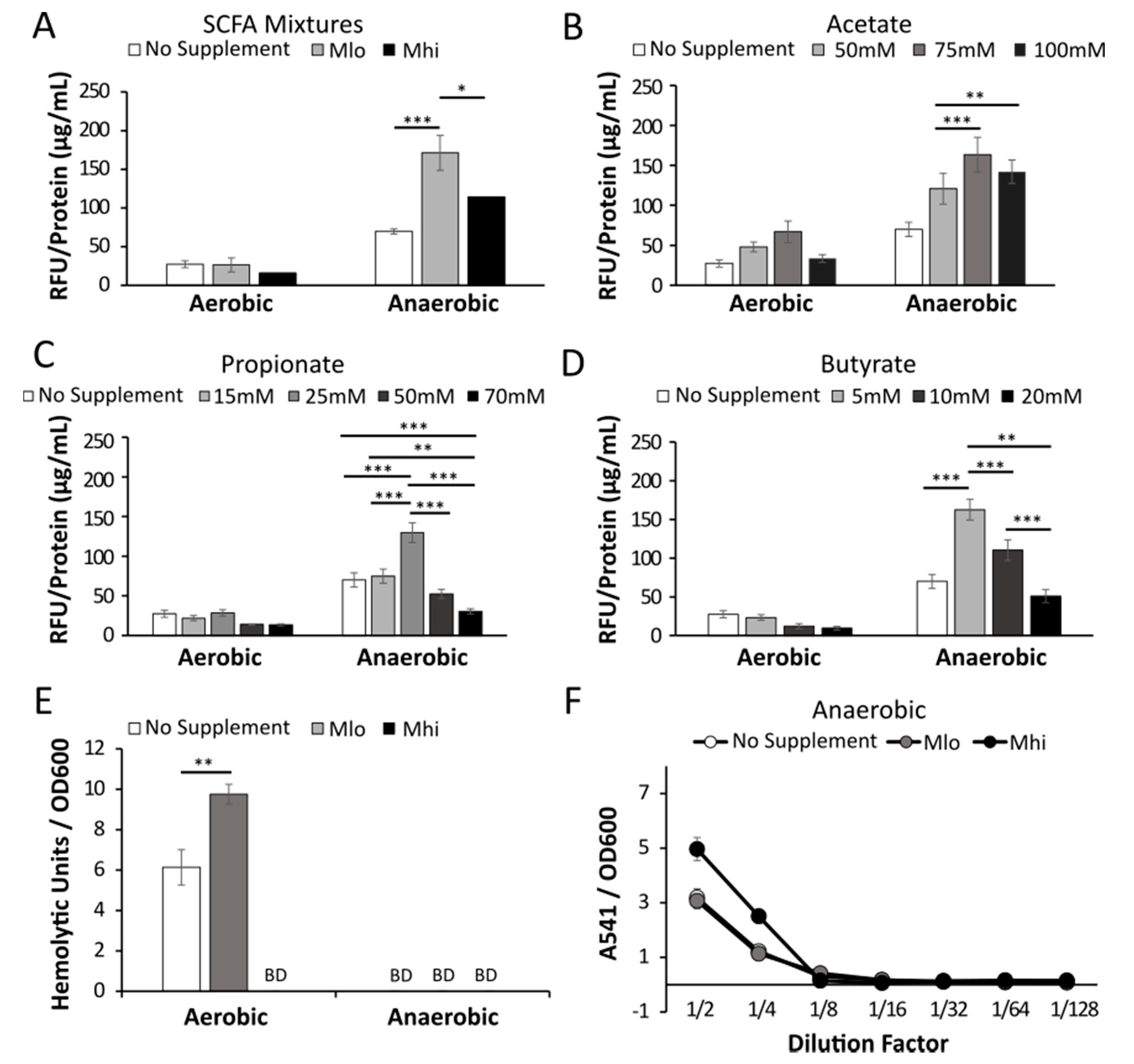
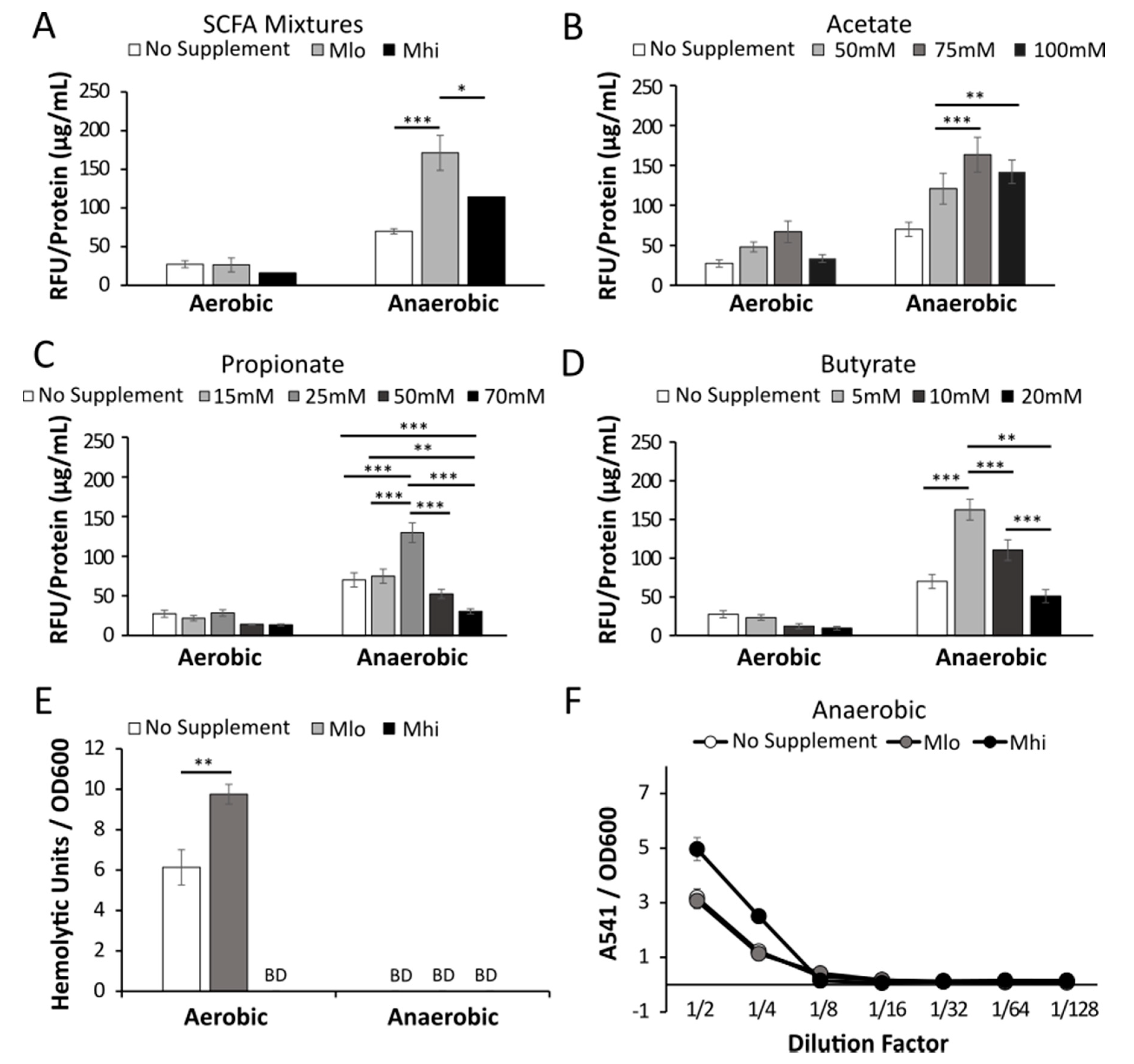
2.3. Anaerobic Transcription of Hly was Enhanced by SCFAs
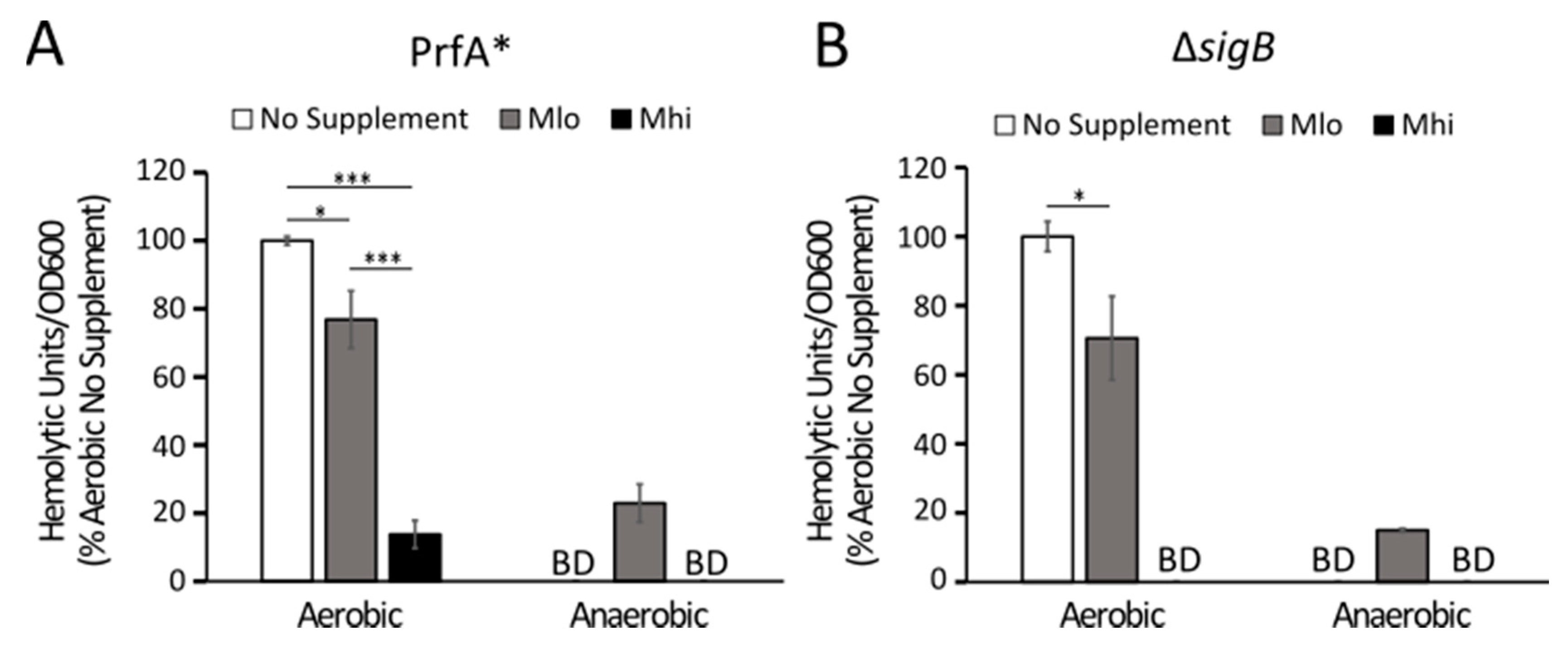
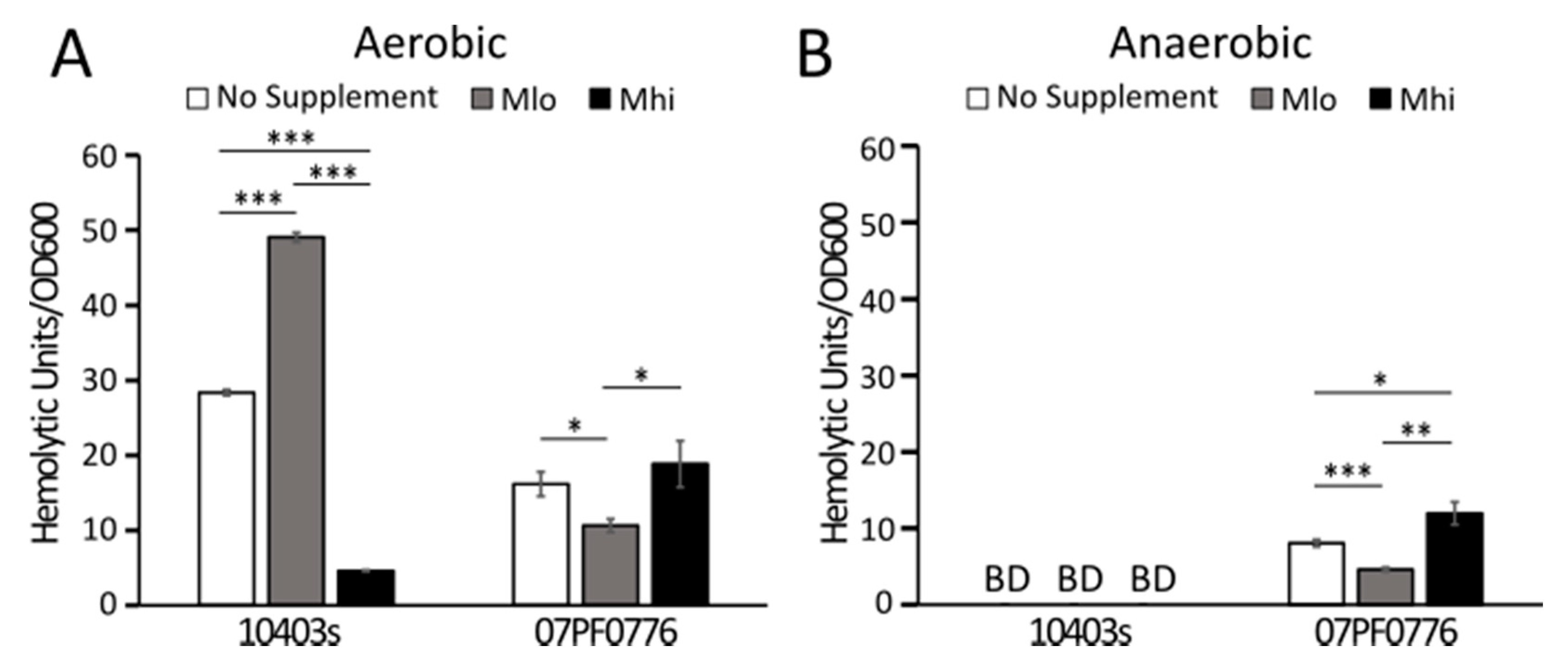
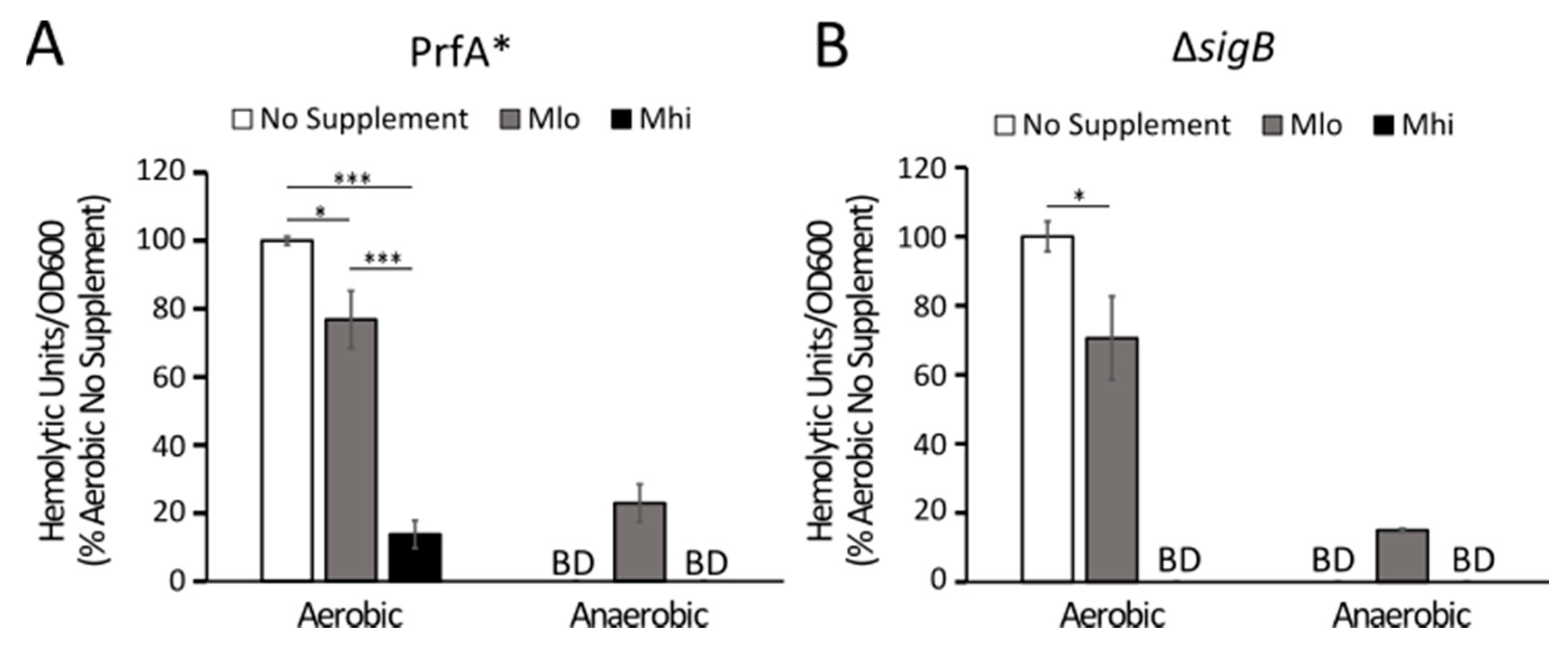
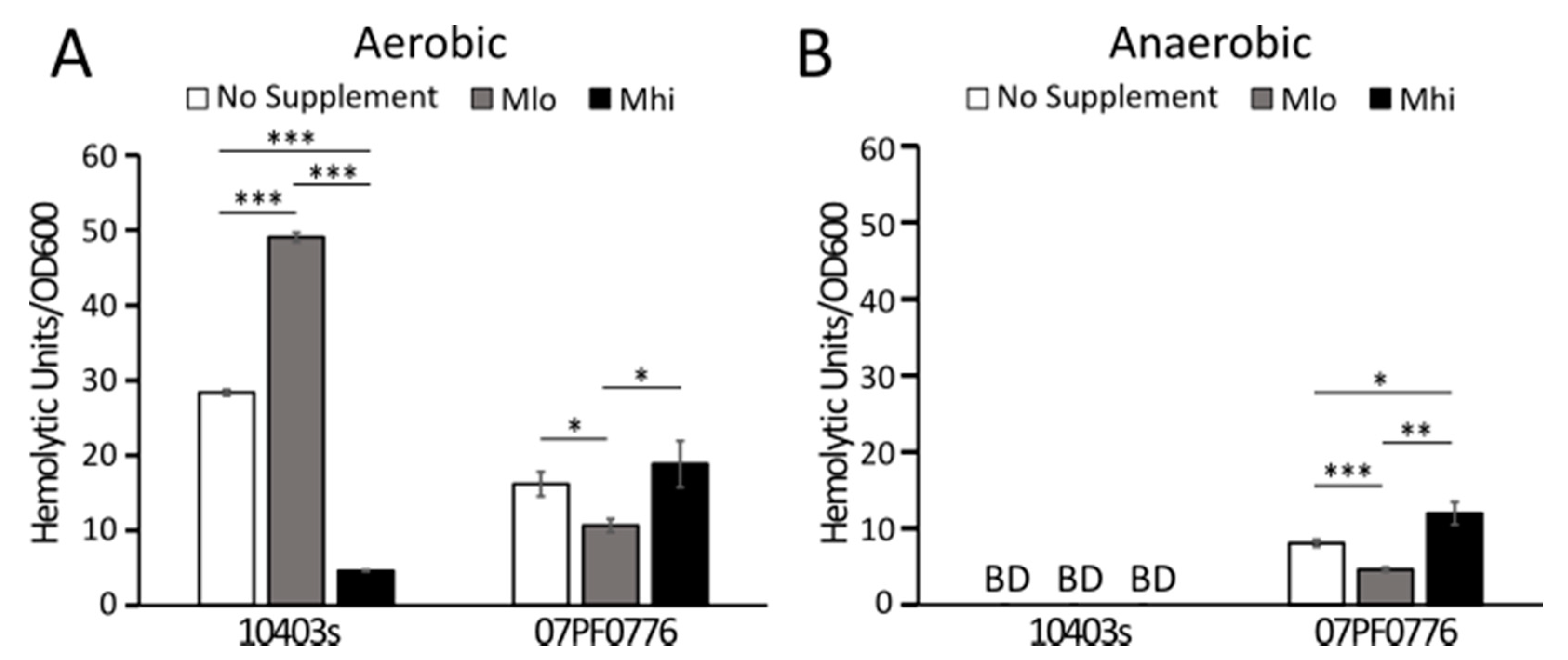
2.4. Production of LLO in Strain 07PF0776 Was Sensitive to Regulation by SCFAs
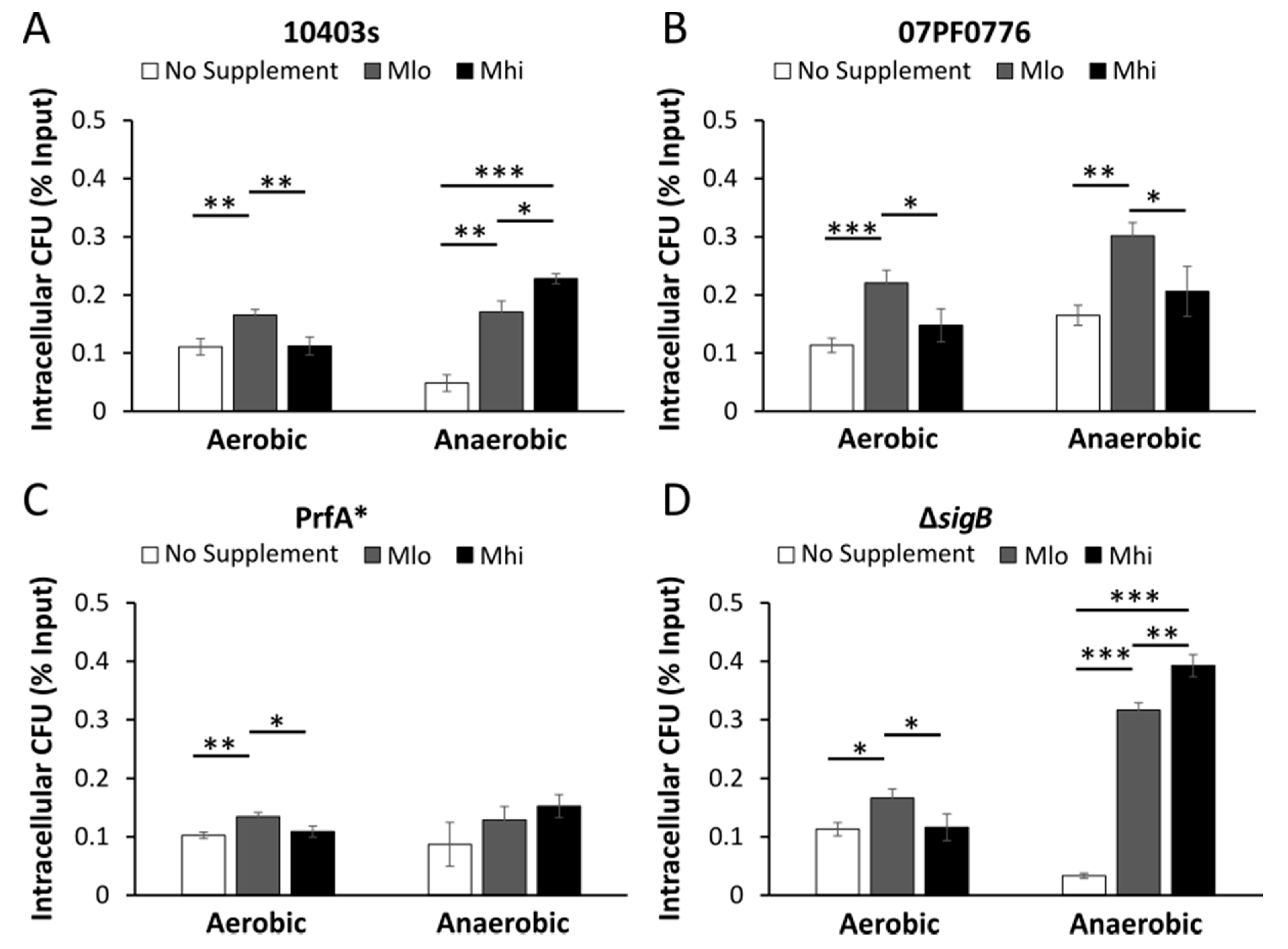
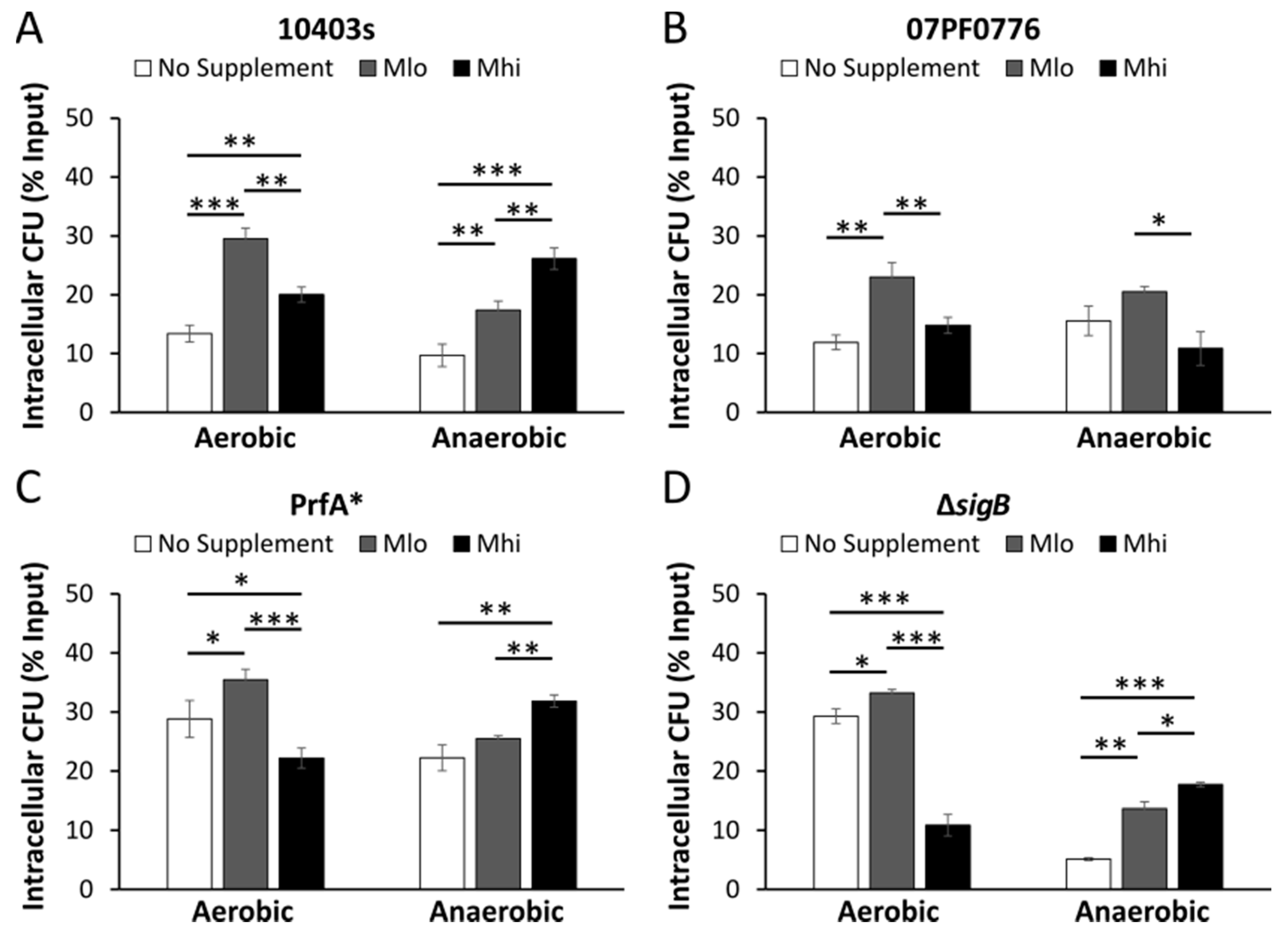
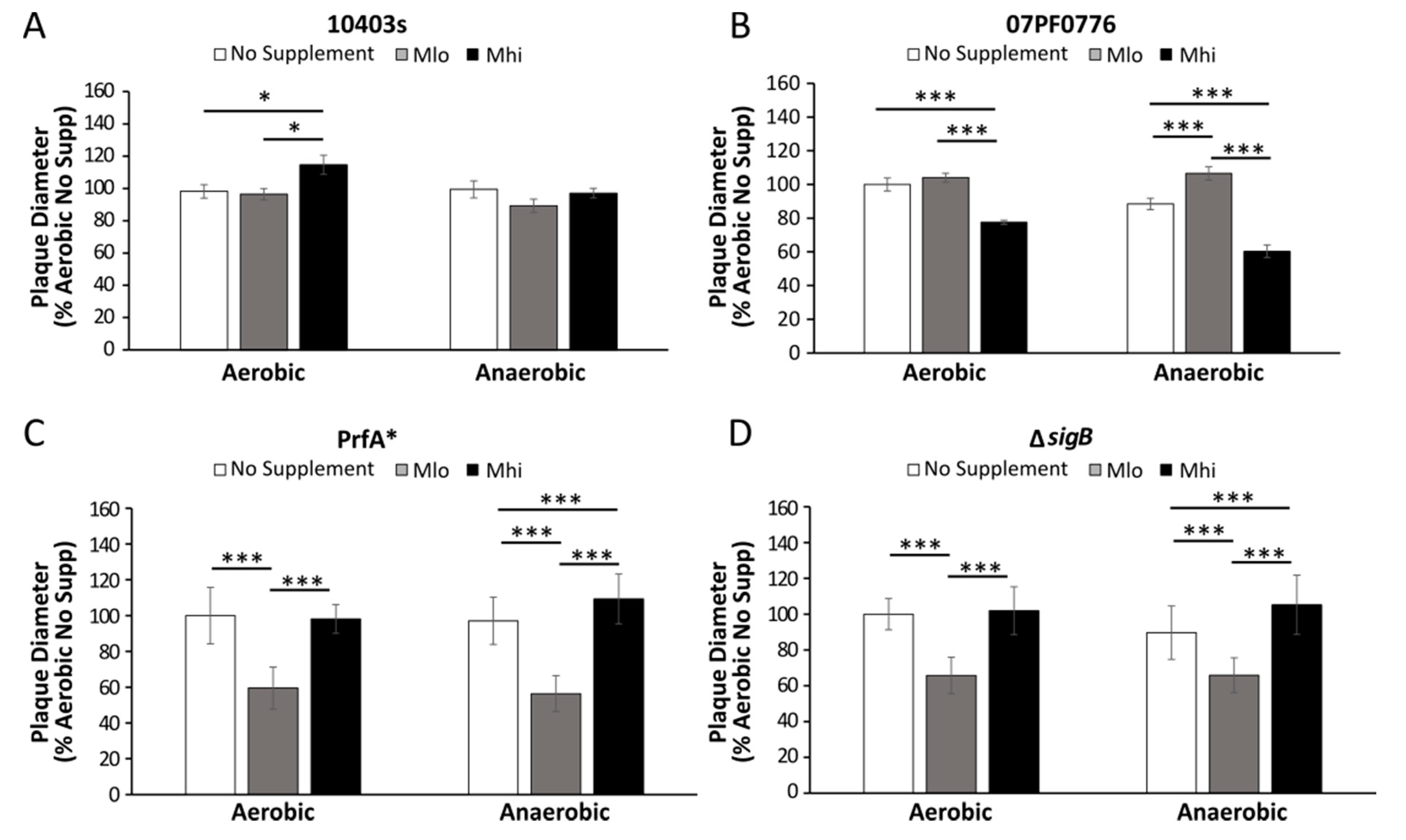
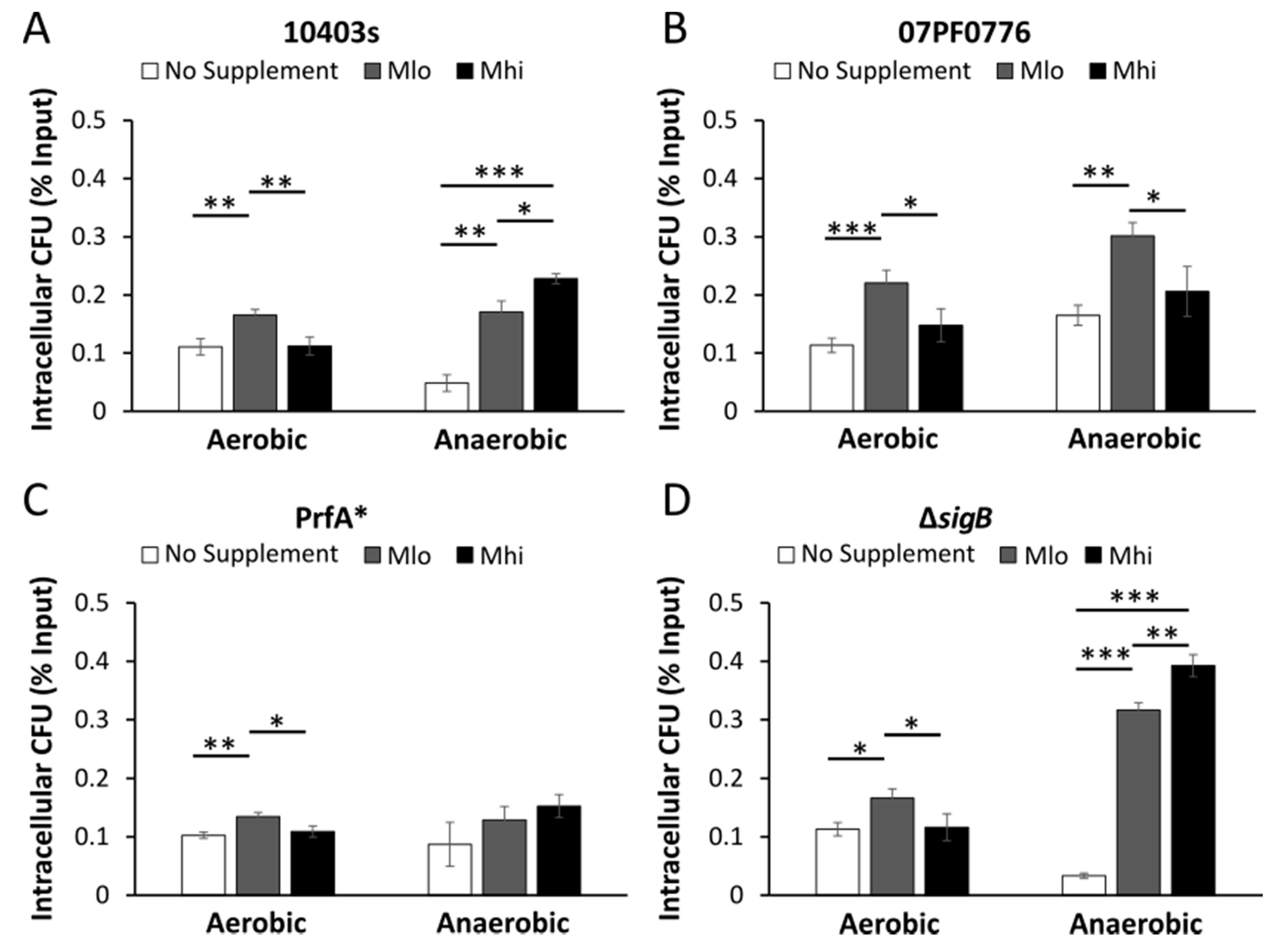
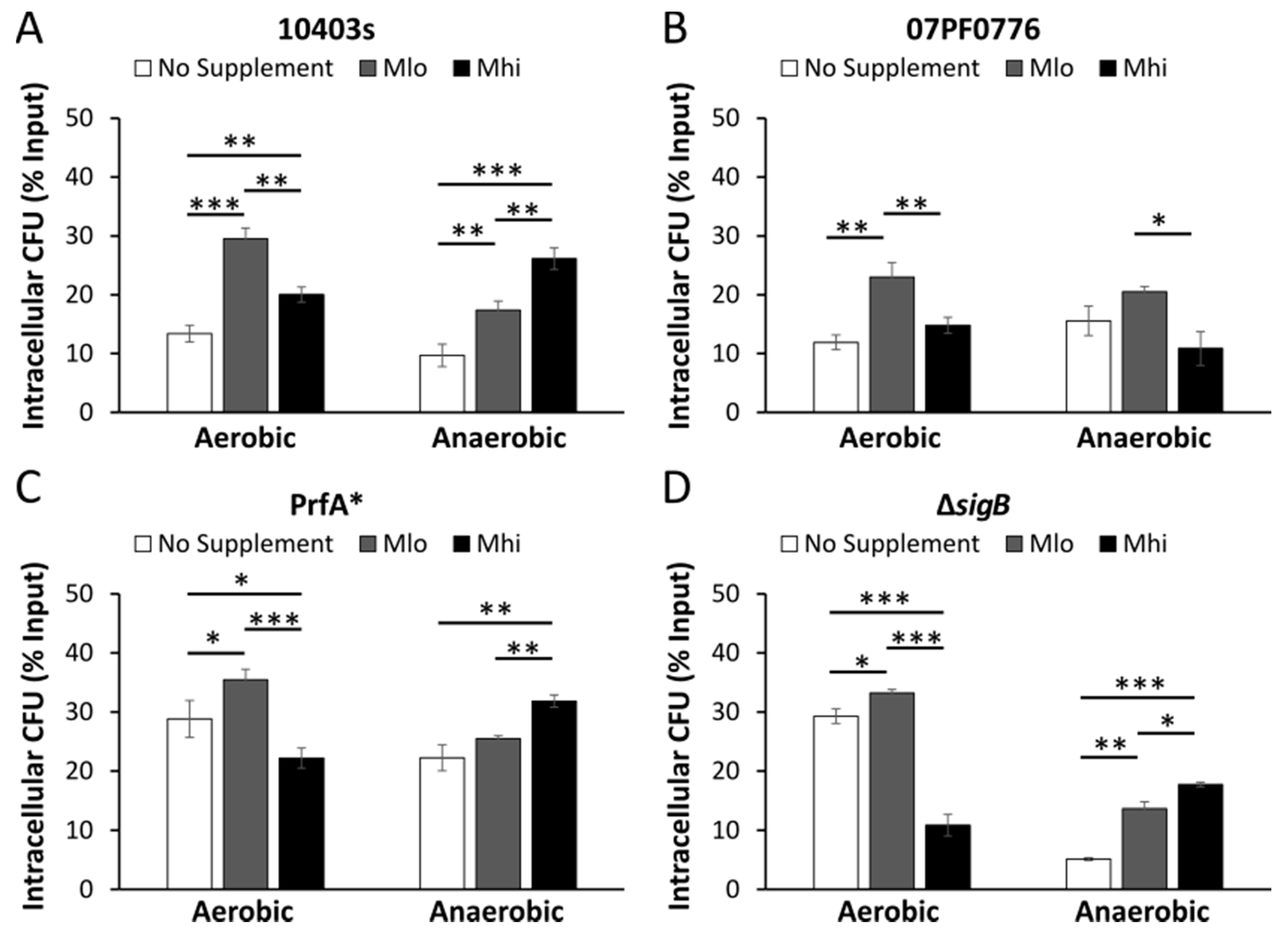
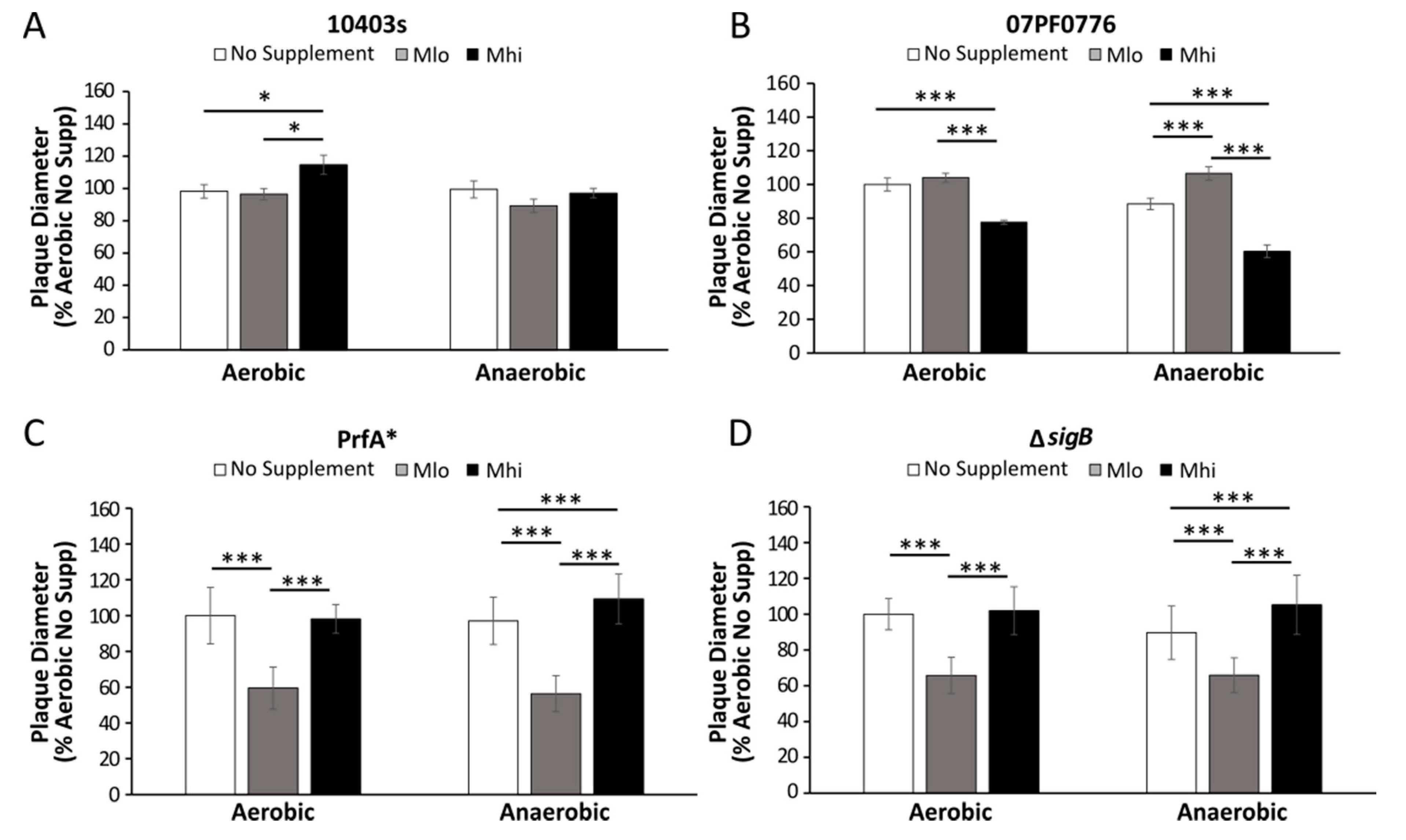
2.5. Prior SCFA Exposure Altered Subsequent Infection Outcomes in Tissue Culture Infections
3. Discussion
3.1. SCFA Metabolism and Fatty Acid Synthesis
3.2. Transcriptional and Post-Transcriptional Regulations of LLO Production by SCFAs
3.3. Strain-Dependent Variations in SCFA Responses
3.4. Impact of SCFA Exposure on Subsequent Cell Culture Infections
3.5. Potential Implications for Transmission
4. Materials and Methods
4.1. Strains
4.2. Growth Conditions
4.3. Culture Supplementation
4.4. GUS Reporter Assay
4.5. Hemolytic Assay
4.6. FAME Analysis
4.7. Tissue Culture Infection
4.8. Fibroblast Plaque Assay
4.9. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scallan, E. Foodborne Illness Acquired in the United States—Major Pathogens. Emerg. Infect. Dis. J. 2011, 17, 7. [Google Scholar] [CrossRef]
- USDA Economic Research Service—Cost Estimates of Foodborne Illnesses. Available online: http://www.ers.usda.gov/data-products/cost-estimates-of-foodborne-illnesses.aspx (accessed on 28 March 2015).
- Schnupf, P.; Portnoy, D.A. Listeriolysin O: A phagosome-specific lysin. Microbes Infect. 2007, 9, 1176–1187. [Google Scholar] [CrossRef]
- Myers, E.R.; Dallmier, A.W.; Martin, S.E. Sodium chloride, potassium chloride, and virulence in Listeria monocytogenes. Appl. Environ. Microbiol. 1993, 59, 2082–2086. [Google Scholar] [CrossRef] [Green Version]
- Datta, A.R.; Kothary, M.H. Effects of glucose, growth temperature, and pH on listeriolysin O production in Listeria monocytogenes. Appl. Environ. Microbiol. 1993, 59, 3495–3497. [Google Scholar] [CrossRef] [Green Version]
- Buncic, S.; Avery, S.M.; Rogers, A.R. Listeriolysin O production and pathogenicity of non-growing Listeria monocytogenes stored at refrigeration temperature. Int. J. Food Microbiol. 1996, 31, 133–147. [Google Scholar] [CrossRef]
- Wallace, N.; Newton, E.; Abrams, E.; Zani, A.; Sun, Y. Metabolic determinants in Listeria monocytogenes anaerobic listeriolysin O production. Arch. Microbiol. 2017, 199, 827–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Wilkinson, B.J.; Standiford, T.J.; Akinbi, H.T.; O’Riordan, M.X.D. Fatty acids regulate stress resistance and virulence factor production for Listeria monocytogenes. J. Bacteriol. 2012, 194, 5274–5284. [Google Scholar] [CrossRef] [Green Version]
- Rinehart, E.; Newton, E.; Marasco, M.A.; Beemiller, K.; Zani, A.; Muratore, M.K.; Weis, J.; Steinbicker, N.; Wallace, N.; Sun, Y. Listeria monocytogenes Response to Propionate Is Differentially Modulated by Anaerobicity. Pathog. Basel Switz. 2018, 7, 60. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, N.; Pluznick, J.L. From microbe to man: The role of microbial short chain fatty acid metabolites in host cell biology. Am. J. Physiol. Cell Physiol. 2014, 307, C979–C985. [Google Scholar] [CrossRef] [Green Version]
- McNeil, N.I. The contribution of the large intestine to energy supplies in man. Am. J. Clin. Nutr. 1984, 39, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.J.; Arusi, E.; Vinik, A.I.; Johnson, D.A. The Role and Influence of Gut Microbiota in Pathogenesis and Management of Obesity and Metabolic Syndrome. Front. Endocrinol. 2014, 5, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzierzewicz, Z.; Orchel, A.; Weglarz, L.; Latocha, M.; Wilczok, T. Changes in the cellular behaviour of human colonic cell line Caco-2 in response to butyrate treatment. Acta Biochim. Pol. 2002, 49, 211–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatayama, H.; Iwashita, J.; Kuwajima, A.; Abe, T. The short chain fatty acid, butyrate, stimulates MUC2 mucin production in the human colon cancer cell line, LS174T. Biochem. Biophys. Res. Commun. 2007, 356, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of butyrate on intestinal barrier function in a Caco-2 cell monolayer model of intestinal barrier. Pediatr. Res. 2007, 61, 37–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, P.R.; Rosella, O.; Wilson, A.J.; Mariadason, J.M.; Rickard, K.; Byron, K.; Barkla, D.H. Colonic epithelial cell activation and the paradoxical effects of butyrate. Carcinogenesis 1999, 20, 539–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boren, J.; Lee, W.-N.P.; Bassilian, S.; Centelles, J.J.; Lim, S.; Ahmed, S.; Boros, L.G.; Cascante, M. The stable isotope-based dynamic metabolic profile of butyrate-induced HT29 cell differentiation. J. Biol. Chem. 2003, 278, 28395–28402. [Google Scholar] [CrossRef] [Green Version]
- Vinolo, M.A.R.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of Inflammation by Short Chain Fatty Acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Vinolo, M.A.R.; Rodrigues, H.G.; Hatanaka, E.; Sato, F.T.; Sampaio, S.C.; Curi, R. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils. J. Nutr. Biochem. 2011, 22, 849–855. [Google Scholar] [CrossRef]
- Liu, T.; Li, J.; Liu, Y.; Xiao, N.; Suo, H.; Xie, K.; Yang, C.; Wu, C. Short-chain fatty acids suppress lipopolysaccharide-induced production of nitric oxide and proinflammatory cytokines through inhibition of NF-κB pathway in RAW264.7 cells. Inflammation 2012, 35, 1676–1684. [Google Scholar] [CrossRef]
- Singh, N.; Thangaraju, M.; Prasad, P.D.; Martin, P.M.; Lambert, N.A.; Boettger, T.; Offermanns, S.; Ganapathy, V. Blockade of dendritic cell development by bacterial fermentation products butyrate and propionate through a transporter (Slc5a8)-dependent inhibition of histone deacetylases. J. Biol. Chem. 2010, 285, 27601–27608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neish, A.S. Microbes in gastrointestinal health and disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annous, B.A.; Becker, L.A.; Bayles, D.O.; Labeda, D.P.; Wilkinson, B.J. Critical role of anteiso-C-15:0 fatty acid in the growth of Listeria monocytogenes at low temperatures. Appl. Environ. Microbiol. 1997, 63, 3887–3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giotis, E.S.; McDowell, D.A.; Blair, I.S.; Wilkinson, B.J. Role of Branched-Chain Fatty Acids in pH Stress Tolerance in Listeria monocytogenes. Appl. Environ. Microbiol. 2007, 73, 997–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneda, T. Iso- and anteiso-fatty acids in bacteria: Biosynthesis, function, and taxonomic significance. Microbiol. Rev. 1991, 55, 288–302. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; O’Riordan, M.X.D. Branched-chain fatty acids promote Listeria monocytogenes intracellular infection and virulence. Infect. Immun. 2010, 78, 4667–4673. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Bayles, D.O.; Xiong, A.; Jayaswal, R.K.; Wilkinson, B.J. Precursor and temperature modulation of fatty acid composition and growth of Listeria monocytogenes cold-sensitive mutants with transposon-interrupted branched-chain α-keto acid dehydrogenase. Microbiology 2005, 151, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Lawhon, S.; Maurer, R.; Suyemoto, M.; Altier, C. Intestinal short-chain fatty acids alter Salmonella typhimurium invasion gene expression and virulence through BarA/SirA RID G-5147-2011. Mol. Microbiol. 2002, 46, 1451–1464. [Google Scholar] [CrossRef]
- Kazmierczak, M.J.; Wiedmann, M.; Boor, K.J. Contributions of Listeria monocytogenes σB and PrfA to expression of virulence and stress response genes during extra- and intracellular growth. Microbiology 2006, 152, 1827–1838. [Google Scholar] [CrossRef] [Green Version]
- McMullen, P.D.; Gillaspy, A.F.; Gipson, J.; Bobo, L.D.; Skiest, D.J.; Freitag, N.E. Genome sequence of Listeria monocytogenes 07PF0776, a cardiotropic serovar 4b strain. J. Bacteriol. 2012, 194, 3552. [Google Scholar] [CrossRef] [Green Version]
- Sirobhushanam, S.; Galva, C.; Sen, S.; Wilkinson, B.J.; Gatto, C. Broad substrate specificity of phosphotransbutyrylase from Listeria monocytogenes: A potential participant in an alternative pathway for provision of acyl CoA precursors for fatty acid biosynthesis. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2016, 1861, 1102–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suutari, M.; Laakso, S. Microbial fatty acids and thermal adaptation. Crit. Rev. Microbiol. 1994, 20, 285–328. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Zhang, Y.-M.; Zhu, K.; Subramanian, C.; Li, Z.; Jayaswal, R.K.; Gatto, C.; Rock, C.O.; Wilkinson, B.J. FabH selectivity for anteiso branched-chain fatty acid precursors in low-temperature adaptation in Listeria monocytogenes. FEMS Microbiol. Lett. 2009, 301, 188–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastronicolis, S.K.; German, J.B.; Megoulas, N.; Petrou, E.; Foka, P.; Smith, G.M. Influence of cold shock on the fatty-acid composition of different lipid classes of the food-borne pathogen Listeria monocytogenes. Food Microbiol. 1998, 15, 299–306. [Google Scholar] [CrossRef]
- Püttmann, M.; Ade, N.; Hof, H. Dependence of fatty acid composition of Listeria spp. on growth temperature. Res. Microbiol. 1993, 144, 279–283. [Google Scholar]
- Juneja, V.K.; Davidson, P.M. Influence of Temperature on the Fatty Acid Profile of Listeria Monocytogenes. J. Rapid Methods Autom. Microbiol. 1993, 2, 73–81. [Google Scholar] [CrossRef]
- Nichols, D.S.; Presser, K.A.; Olley, J.; Ross, T.; McMeekin, T.A. Variation of Branched-Chain Fatty Acids Marks the Normal Physiological Range for Growth in Listeria monocytogenes. Appl. Environ. Microbiol. 2002, 68, 2809–2813. [Google Scholar] [CrossRef] [Green Version]
- Saunders, L.P.; Sen, S.; Wilkinson, B.J.; Gatto, C. Insights into the Mechanism of Homeoviscous Adaptation to Low Temperature in Branched-Chain Fatty Acid-Containing Bacteria through Modeling FabH Kinetics from the Foodborne Pathogen Listeria monocytogenes. Front. Microbiol. 2016, 7, 1386. [Google Scholar] [CrossRef] [Green Version]
- Dubois-Brissonnet, F.; Trotier, E.; Briandet, R. The Biofilm Lifestyle Involves an Increase in Bacterial Membrane Saturated Fatty Acids. Front. Microbiol. 2016, 7, 1673. [Google Scholar] [CrossRef] [Green Version]
- Kader, A.A.; Zagory, D.; Kerbel, E.L. Modified atmosphere packaging of fruits and vegetables. Crit. Rev. Food Sci. Nutr. 1989, 28, 1–30. [Google Scholar] [CrossRef]
- Bohne, J.; Sokolovic, Z.; Goebel, W. Transcriptional regulation of prfA and PrfA-regulated virulence genes in Listeria monocytogenes. Mol. Microbiol. 1994, 11, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- de las Heras, A.; Cain, R.J.; Bielecka, M.K.; Vázquez-Boland, J.A. Regulation of Listeria virulence: PrfA master and commander. Curr. Opin. Microbiol. 2011, 14, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Freitag, N.E.; Rong, L.; Portnoy, D.A. Regulation of the prfA transcriptional activator of Listeria monocytogenes: Multiple promoter elements contribute to intracellular growth and cell-to-cell spread. Infect. Immun. 1993, 61, 2537–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scortti, M.; Monzó, H.J.; Lacharme-Lora, L.; Lewis, D.A.; Vázquez-Boland, J.A. The PrfA virulence regulon. Microbes Infect. Inst. Pasteur 2007, 9, 1196–1207. [Google Scholar] [CrossRef]
- Hecker, M.; Pané-Farré, J.; Uwe, V. SigB-Dependent General Stress Response in Bacillus subtilis and Related Gram-Positive Bacteria. Annu. Rev. Microbiol. 2007, 61, 215–236. [Google Scholar] [CrossRef]
- Nadon, C.A.; Bowen, B.M.; Wiedmann, M.; Boor, K.J. Sigma B contributes to PrfA-mediated virulence in Listeria monocytogenes. Infect. Immun. 2002, 70, 3948–3952. [Google Scholar] [CrossRef] [Green Version]
- Ollinger, J.; Bowen, B.; Wiedmann, M.; Boor, K.J.; Bergholz, T.M. Listeria monocytogenes σB Modulates PrfA-Mediated Virulence Factor Expression. Infect. Immun. 2009, 77, 2113–2124. [Google Scholar] [CrossRef] [Green Version]
- Schwab, U.; Bowen, B.; Nadon, C.; Wiedmann, M.; Boor, K.J. The Listeria monocytogenes prfAP2 promoter is regulated by sigma B in a growth phase dependent manner. FEMS Microbiol. Lett. 2005, 245, 329–336. [Google Scholar] [CrossRef]
- Rauch, M.; Luo, Q.; Müller-Altrock, S.; Goebel, W. SigB-Dependent In Vitro Transcription of prfA and Some Newly Identified Genes of Listeria monocytogenes Whose Expression Is Affected by PrfA In Vivo. J. Bacteriol. 2005, 187, 800–804. [Google Scholar] [CrossRef] [Green Version]
- Bruno, J.C.; Freitag, N.E. Constitutive Activation of PrfA Tilts the Balance of Listeria monocytogenes Fitness Towards Life within the Host versus Environmental Survival. PLoS ONE 2010, 5, e15138. [Google Scholar] [CrossRef]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- McLauchlin, J.; Mitchell, R.T.; Smerdon, W.J.; Jewell, K. Listeria monocytogenes and listeriosis: A review of hazard characterisation for use in microbiological risk assessment of foods. Int. J. Food Microbiol. 2004, 92, 15–33. [Google Scholar] [CrossRef]
- Jacquet, C.; Gouin, E.; Jeannel, D.; Cossart, P.; Rocourt, J. Expression of ActA, Ami, InlB, and Listeriolysin O in Listeria monocytogenes of Human and Food Origin. Appl. Environ. Microbiol. 2002, 68, 616–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabouret, M.; De Rycke, J.; Audurier, A.; Poutrel, B. Pathogenicity of Listeria monocytogenes isolates in immunocompromised mice in relation to listeriolysin production. J. Med. Microbiol. 1991, 34, 13–18. [Google Scholar] [CrossRef]
- Erdenlig, S.; Ainsworth, A.J.; Austin, F.W. Pathogenicity and Production of Virulence Factors by Listeria monocytogenes Isolates from Channel Catfish. J. Food Prot. 2000, 63, 613–619. [Google Scholar] [CrossRef]
- Ripio, M.-T.; Domínguez-Bernal, G.; Suárez, M.; Brehm, K.; Berche, P.; Vázquez-Boland, J.-A. Transcriptional activation of virulence genes in wild-type strains of Listeria monocytogenes in response to a change in the extracellular medium composition. Res. Microbiol. 1996, 147, 371–384. [Google Scholar] [CrossRef]
- Lemes-Marques, E.G.; Yano, T. Influence of environmental conditions on the expression of virulence factors by Listeria monocytogenes and their use in species identification. FEMS Microbiol. Lett. 2004, 239, 63–70. [Google Scholar] [CrossRef]
- Alonzo, F.; Bobo, L.D.; Skiest, D.J.; Freitag, N.E. Evidence for subpopulations of Listeria monocytogenes with enhanced invasion of cardiac cells. J. Med. Microbiol. 2011, 60, 423–434. [Google Scholar] [CrossRef]
- Hasebe, R.; Nakao, R.; Ohnuma, A.; Yamasaki, T.; Sawa, H.; Takai, S.; Horiuchi, M. Listeria monocytogenes serotype 4b strains replicate in monocytes/macrophages more than the other serotypes. J. Vet. Med. Sci. 2017, 79, 962–969. [Google Scholar] [CrossRef] [Green Version]
- Menudier, A.; Bosiraud, C.; Nicolas, J.-A. Virulence of Listeria monocytogenes Serovars and Listeria spp. in Experimental Infection of Mice. J. Food Prot. 1991, 54, 917–921. [Google Scholar] [CrossRef]
- Sun, Y.; O’Riordan, M.X.D. Regulation of bacterial pathogenesis by intestinal short-chain Fatty acids. Adv. Appl. Microbiol. 2013, 85, 93–118. [Google Scholar] [PubMed] [Green Version]
- Burkholder, K.M.; Kim, K.-P.; Mishra, K.K.; Medina, S.; Hahm, B.-K.; Kim, H.; Bhunia, A.K. Expression of LAP, a SecA2-dependent secretory protein, is induced under anaerobic environment. Microbes Infect. 2009, 11, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.B.; Roldgaard, B.B.; Christensen, B.B.; Licht, T.R. Oxygen restriction increases the infective potential of Listeria monocytogenes in vitro in Caco-2 cells and in vivo in guinea pigs. BMC Microbiol. 2007, 7, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gesbert, G.; Ramond, E.; Tros, F.; Dairou, J.; Frapy, E.; Barel, M.; Charbit, A. Importance of Branched-Chain Amino Acid Utilization in Francisella Intracellular Adaptation. Infect. Immun. 2015, 83, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CFR—Code of Federal Regulations Title 21. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=184.1081 (accessed on 16 June 2019).
- Affairs, O. of R. CPG Sec. 562.100 Acetic Acid—Use in Foods—Labeling of Foods in Which Used. Available online: http://www.fda.gov/regulatory-information/search-fda-guidance-documents/cpg-sec-562100-acetic-acid-use-foods-labeling-foods-which-used (accessed on 20 December 2019).
- CFR—Code of Federal Regulations Title 21. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/cfrsearch.cfm?fr=582.60 (accessed on 16 June 2019).
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.B.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Hippe, B.; Zwielehner, J.; Liszt, K.; Lassl, C.; Unger, F.; Haslberger, A.G. Quantification of butyryl CoA:acetate CoA-transferase genes reveals different butyrate production capacity in individuals according to diet and age. FEMS Microbiol. Lett. 2011, 316, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Mathers, J.C.; Kennard, J.; James, O.F. Gastrointestinal responses to oats consumption in young adult and elderly rats: Digestion, large bowel fermentation and crypt cell proliferation rates. Br. J. Nutr. 1993, 70, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Cuervo, A.; Salazar, N.; Ruas-Madiedo, P.; Gueimonde, M.; González, S. Fiber from a regular diet is directly associated with fecal short-chain fatty acid concentrations in the elderly. Nutr. Res. N. Y. N 2013, 33, 811–816. [Google Scholar] [CrossRef]
- Arboleya, S.; Watkins, C.; Stanton, C.; Ross, R.P. Gut Bifidobacteria Populations in Human Health and Aging. Front. Microbiol. 2016, 7, 1204. [Google Scholar] [CrossRef] [Green Version]
- Coconnier, M.H.; Lorrot, M.; Barbat, A.; Laboisse, C.; Servin, A.L. Listeriolysin O-induced stimulation of mucin exocytosis in polarized intestinal mucin-secreting cells: Evidence for toxin recognition of membrane-associated lipids and subsequent toxin internalization through caveolae. Cell. Microbiol. 2000, 2, 487–504. [Google Scholar] [CrossRef]
- Liévin-Le Moal, V.; Servin, A.L.; Coconnier-Polter, M.-H. The increase in mucin exocytosis and the upregulation of MUC genes encoding for membrane-bound mucins induced by the thiol-activated exotoxin listeriolysin O is a host cell defence response that inhibits the cell-entry of Listeria monocytogenes. Cell. Microbiol. 2005, 7, 1035–1048. [Google Scholar]
- Coconnier, M.H.; Dlissi, E.; Robard, M.; Laboisse, C.L.; Gaillard, J.L.; Servin, A.L. Listeria monocytogenes stimulates mucus exocytosis in cultured human polarized mucosecreting intestinal cells through action of listeriolysin O. Infect. Immun. 1998, 66, 3673–3681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liévin-Le Moal, V.; Huet, G.; Aubert, J.-P.; Bara, J.; Forgue-Lafitte, M.-E.; Servin, A.L.; Coconnier, M.-H. Activation of mucin exocytosis and upregulation of MUC genes in polarized human intestinal mucin-secreting cells by the thiol-activated exotoxin listeriolysin O. Cell. Microbiol. 2002, 4, 515–529. [Google Scholar]
- Vadia, S.; Arnett, E.; Haghighat, A.-C.; Wilson-Kubalek, E.M.; Tweten, R.K.; Seveau, S. The pore-forming toxin listeriolysin O mediates a novel entry pathway of L. monocytogenes into human hepatocytes. PLoS Pathog. 2011, 7, e1002356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadia, S.; Seveau, S. Fluxes of Ca2+ and K+ are required for the listeriolysin O-dependent internalization pathway of Listeria monocytogenes. Infect. Immun. 2014, 82, 1084–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, J.G.T.; Vadia, S.; Pathak-Sharma, S.; McLaughlin, E.; Zhang, X.; Swanson, J.; Seveau, S. Host cell perforation by listeriolysin O (LLO) activates a Ca2+-dependent cPKC/Rac1/Arp2/3 signaling pathway that promotes Listeria monocytogenes internalization independently of membrane resealing. Mol. Biol. Cell 2018, 29, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Phelps, C.C.; Vadia, S.; Arnett, E.; Tan, Y.; Zhang, X.; Pathak-Sharma, S.; Gavrilin, M.A.; Seveau, S. Relative Roles of Listeriolysin O, InlA, and InlB in Listeria monocytogenes Uptake by Host Cells. Infect. Immun. 2018, 86, e00555-18. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.; O’Byrne, C.P.; Boor, K.J. Role of sigma(B) in heat, ethanol, acid, and oxidative stress resistance and during carbon starvation in Listeria monocytogenes. Appl. Environ. Microbiol. 2001, 67, 4454–4457. [Google Scholar] [CrossRef] [Green Version]
- Behari, J.; Youngman, P. Regulation of hly expression in Listeria monocytogenes by carbon sources and pH occurs through separate mechanisms mediated by PrfA. Infect. Immun. 1998, 66, 3635–3642. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, G.T.; Gibson, G.R.; Cummings, J.H. Comparison of fermentation reactions in different regions of the human colon. J. Appl. Bacteriol. 1992, 72, 57–64. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SCFA | OD | pH | Doubling Time (Minutes) | |||
|---|---|---|---|---|---|---|
| WT | WT | WT | PrfA* | ΔsigB | ||
| Aerobic | No | 0.96 ± 0.02 | 5.30 ± 0.02 | 74.73 ± 4.01 | 67.02 ± 1.49 | 74.02 ± 2.21 |
| Mhi | 0.90 ± 0.06 | 5.75 ± 0.01* | 77.69 ± 4.11 | 70.01 ± 2.88 | 75.12 ± 3.34 | |
| Mlo | 0.60 ± 0.01 * | 6.16 ± 0.04 * | 95.88 ± 5.18 * | 96.00 ± 12.60 * | 91.41 ± 4.93 ** | |
| Anaerobic | No | 0.39 ± 0.02 | 4.78 ± 0.03 | 73.99 ± 2.03 | 80.39 ± 3.58 | 80.54 ± 2.28 |
| Mhi | 0.41 ± 0.01 | 5.20 ± 0.03 * | 77.69 ± 3.51 | 83.17 ± 11.07 | 83.84 ± 2.43 | |
| Mlo | 0.32 ± 0.03* | 5.79 ± 0.07 * | 108.87 ± 3.35 *** | 119.61 ± 15.34 *** | 126.59 ± 8.51 *** | |
| SCFA | Buffered BHI | ||||||
|---|---|---|---|---|---|---|---|
| 6.0 | 7.0 | 8.0 | |||||
| OD | pH | OD | pH | OD | pH | ||
| Aerobic | no | 0.87 ± 0.01 | 5.26 ± 0.03 | 0.74 ± 0.04 | 6.65 ± 0.02 | 0.38 ± 0.08 | 7.31 ± 0.01 |
| Mlo | 0.52 ± 0.07 ** | 5.67 ± 0.07 ** | 0.70 ± 0.03 | 6.73 ± 0.04 | 0.33 ± 0.07 | 7.32 ± 0.01 | |
| Mhi | NG | 6.09 ± 0.0 *** | 0.57 ± 0.0 ** | 6.72 ± 0.0 ** | 0.26 ± 0.05 ** | 7.36 ± 0.01 ** | |
| Anaerobic | no | 0.19 ± 0.02 | 5.66 ± 0.02 | 0.40 ± 0.03 | 6.67 ± 0.01 | 0.11 ± 0.03 | 7.84 ± 0.02 |
| Mlo | 0.11 ± 0.03 | 5.78 ± 0.03 * | 0.34 ± 0.03 | 6.67 ± 0.04 | 0.23 ± 0.02 | 7.90 ± 0.12 | |
| Mhi | NG | 6.12 ± 0.03 *** | 0.20 ± 0.2 ** | 6.68 ± 0.06 | 0.15 ± 0.01 ** | 7.55 ± 0.06 ** | |
| Aerobic BHI | |||
|---|---|---|---|
| None | Mlo | Mhi | |
| 13 anteiso | 0.06 | 0.07 | 0.07 |
| 15 anteiso | 40.90 | 40.11 | 29.03 |
| 17 anteiso | 36.26 | 30.82 | 11.31 |
| 19 anteiso | 0.17 | 0.12 | 0 |
| Anteiso Total | 77.39 | 71.11 | 40.41 |
| 13 iso | 0.06 | 0 | 0.09 |
| 15 iso | 9.01 | 9.28 | 9.79 |
| 17 iso | 3.93 | 3.58 | 2.31 |
| Odd Iso Total | 12.99 | 12.86 | 12.18 |
| 14 iso | 0.48 | 0.83 | 2.11 |
| 16 iso | 2.57 | 3.62 | 4.11 |
| 18 iso | 0 | 0 | 0 |
| Even Iso Total | 3.04 | 4.45 | 6.22 |
| Branched Total | 93.42 | 88.41 | 58.8 |
| 11 straight | 0 | 0 | 0.16 |
| 12 straight | 0.11 | 0.17 | 0.65 |
| 13 straight | 0 | 0.07 | 1.78 |
| 14 straight | 0.30 | 1.18 | 5.45 |
| 16 straight | 0 | 1.4 | 13.17 |
| 17 straight | 4.54 | 7.21 | 15.67 |
| 18 straight | 0 | 0.24 | 1.35 |
| 20 straight | 1.72 | 1.25 | 2.87 |
| Straight Total | 6.655 | 11.51 | 41.085 |
| 16:1 w7c | 0 | 0 | 0 |
| 18.1 w9c | 0.08 | 0.08 | 0.11 |
| Unsaturated Total | 0.08 | 0.08 | 0.11 |
| Branched to Straight Ratio | 14.04 | 7.68 | 1.43 |
| Culture OD | 0.685 ± 0.10 | 0.74 ± 0.02 | 0.480 ± 0.01 |
| Anaerobic BHI | |||
|---|---|---|---|
| No Supp | Mlo | Mhi | |
| 13 anteiso | 0 | 0 | 0 |
| 15 anteiso | 36.85 | 33.86 | 23.68 |
| 17 anteiso | 24.86 | 21.06 | 8.12 |
| 19 anteiso | 0.17 | 0.13 | 0 |
| Anteiso Total | 61.88 | 55.05 | 31.80 |
| 13 iso | 0 | 0 | 0 |
| 15 iso | 13.22 | 10.89 | 7.38 |
| 17 iso | 5.25 | 4.32 | 1.91 |
| Odd Iso Total | 18.47 | 15.21 | 9.29 |
| 14 iso | 1.27 | 1.41 | 1.53 |
| 16 iso | 4.3 | 4.42 | 2.57 |
| 18 iso | 0 | 0 | 0 |
| Even Iso Total | 5.57 | 5.83 | 4.10 |
| Branched Total | 85.93 | 76.10 | 45.20 |
| 11 straight | 0 | 0 | 0.1 |
| 12 straight | 0.19 | 0.27 | 0.35 |
| 13 straight | 0 | 0.12 | 2.33 |
| 14 straight | 0.56 | 2.23 | 5.08 |
| 15 Straight | 0 | 2.815 | 25.39 |
| 16 straight | 4.25 | 9.14 | 11.34 |
| 17 straight | 0 | 0.41 | 2.11 |
| 18 straight | 0.84 | 1.07 | 1.39 |
| 20 straight | 0 | 0 | 0 |
| Straight Total | 5.85 | 16.07 | 48.11 |
| 16:1 w7c | 0 | 0 | 0 |
| 18.1 w9c | 0 | 0 | 0 |
| Unsaturated Total | 0 | 0 | 0 |
| 15:0 ISO aldehyde | 0.29 | 0.2 | 0.1 |
| unknown 13.493 | 0 | 0 | 0 |
| 14:0 2OH | 0 | 0 | 0 |
| 14:0 DMA | 0 | 0 | 0 |
| 15:0 iso DMA | 1.04 | 0.71 | 0.31 |
| 15:0 DMA | 0 | 0.21 | 1.89 |
| 16:0 aldehyde | 0 | 0.19 | 0.30 |
| UN 16.107 | 1.08 | 0.98 | 0.50 |
| 16:0 DMA | 0.44 | 1.08 | 1.39 |
| un 17.103 | 0.73 | 0.62 | 0.33 |
| 17:0 anteiso DMA | 4.66 | 3.84 | 1.90 |
| Other Total | 8.24 | 7.83 | 6.72 |
| Branched to Straight Ratio | 14.67 | 4.73 | 0.94 |
| Culture OD | 0.524 ± 0.01 | 0.485 ± 0.00 | 0.301 ± 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinehart, E.; Chapman, J.; Sun, Y. The Production of Listeriolysin O and Subsequent Intracellular Infections by Listeria monocytogenes Are Regulated by Exogenous Short Chain Fatty Acid Mixtures. Toxins 2020, 12, 218. https://doi.org/10.3390/toxins12040218
Rinehart E, Chapman J, Sun Y. The Production of Listeriolysin O and Subsequent Intracellular Infections by Listeria monocytogenes Are Regulated by Exogenous Short Chain Fatty Acid Mixtures. Toxins. 2020; 12(4):218. https://doi.org/10.3390/toxins12040218
Chicago/Turabian StyleRinehart, Erica, Julia Chapman, and Yvonne Sun. 2020. "The Production of Listeriolysin O and Subsequent Intracellular Infections by Listeria monocytogenes Are Regulated by Exogenous Short Chain Fatty Acid Mixtures" Toxins 12, no. 4: 218. https://doi.org/10.3390/toxins12040218