Polyphenol Compound as a Transcription Factor Inhibitor

Department of Applied Chemistry, Dongduk Women’s University, Seoul 136-714, Korea
Nutrients 2015, 7(11), 8987-9004; https://doi.org/10.3390/nu7115445
Submission received: 15 July 2015
/
Revised: 12 October 2015
/
Accepted: 23 October 2015
/
Published: 30 October 2015
Abstract
:A target-based approach has been used to develop novel drugs in many therapeutic fields. In the final stage of intracellular signaling, transcription factor–DNA interactions are central to most biological processes and therefore represent a large and important class of targets for human therapeutics. Thus, we focused on the idea that the disruption of protein dimers and cognate DNA complexes could impair the transcriptional activation and cell transformation regulated by these proteins. Historically, natural products have been regarded as providing the primary leading compounds capable of modulating protein–protein or protein-DNA interactions. Although their mechanism of action is not fully defined, polyphenols including flavonoids were found to act mostly as site-directed small molecule inhibitors on signaling. There are many reports in the literature of screening initiatives suggesting improved drugs that can modulate the transcription factor interactions responsible for disease. In this review, we focus on polyphenol compound inhibitors against dimeric forms of transcription factor components of intracellular signaling pathways (for instance, c-jun/c-fos (Activator Protein-1; AP-1), c-myc/max, Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and β-catenin/T cell factor (Tcf)).
1. Introduction
Polyphenol compounds have been widely studied for their bioactive properties as antioxidants and for their broad range of roles in the prevention of cancers, cardiovascular diseases and neurodegenerative diseases. Polyphenol compounds are abundant in numerous groups of natural plant products (fruit, vegetables, grains and nuts) and can be a dietary food constituent. Polyphenols are divided into several classes, i.e., phenolic acids (hydroxybenzoic acids and hydroxycinnamic acids), flavonoids (flavonols, flavones, flavanols, flavanones, isoflavones, proanthocyanidins), stilbenes, and lignans [1,2,3].
The mechanism underlying how polyphenol compounds play roles in the prevention of cancers, cardiovascular diseases and neurodegenerative diseases may be related to their modulation of cell signaling pathways. Therefore, to explain the mechanisms behind the actions of polyphenols, a number of signaling targets have been studied.
In this review, we discuss in particular the disruption of multimeric forms of transcription factors such as c-jun/c-fos (Activator Protein-1; AP-1), c-myc/max, Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and β-catenin/T cell factor (Tcf) with small polyphenol compounds that block their protein-protein interactions or their interactions with DNA.
Cell proliferation, differentiation and transformation are regulated in response to cell surface stimuli in accordance with intracellular signals. In the final stage of regulation, transcription factors play an important role in the expression of genetic information. AP-1 is a heterodimeric transcription factor formed by the products of the fos and jun proto-oncogene families. The fos and the jun proteins have almost identical amino acid sequences that comprise their basic DNA binding sequence (B) and the adjacent leucine zipper region (Zip), by which the proteins dimerize with each other [4,5,6]. The AP-1 transcription factor recognizes and binds specifically to the DNA sequence 5′-TGAG/CTCA, known as the AP-1 site [7,8]. In many cases, activation of c-jun is involved in transmitting cancer-promoting signals. The c-jun gene may be permanently activated or overexpressed, which can lead to neoplastic transformation [9,10]. Jun is known to be over-expressed between 4- and 12-fold in 40% of human small-cell lung cancers and 20% of non-small cell lung cancers [9]. In addition, jun may be involved in leukemia [11]. According to some reported research, the elevated levels of c-jun and c-fos expression, as well as of AP-1-dependent target genes, are found in tumors derived from in vivo and in vitro transformation [12,13]. Disruption of fos and jun dimerization has been shown to impair the transcriptional activation and cell transformation regulated by these proteins [14,15].
Similarly, the c-myc proto-oncogene product is one of the essential transcription factors that induce cellular growth, proliferation, cell cycle entry and differentiation and is believed to be involved in the generation of many types of human malignancies, cell cycle progression and proliferation [16,17,18,19]. Biological activity of myc occurs upon hetero-dimerization with max, a small and ubiquitously expressed phosphoprotein [20,21,22]. The C-terminal domain of the c-myc and max proteins includes a basic domain/helix-loop-helix/leucine zipper (b/HLH/Z) motif that mediates binding each other through the HLH/Z region and specific DNA recognition of CACGTG E box motifs present in all target genes through the basic domain [20,23,24]. Myc is constitutively overexpressed in lymphoblastoid cells lines derived from individuals with the cancer-prone condition Bloom’s syndrome and there is evidence that myc de-regulation may be involved in the early stages of mammary carcinogenesis [25,26,27]. Myc is enhanced in many tumors, particularly small-cell-lung, breast and cervical carcinomas [25,26,27,28]. Especially, amplified c-myc oncogene was found in human stomach cancers and it has been suggested that c-myc mRNA overexpression might be crucial in the early development of primary lesions as well as in the formation of metastatic lesions of carcinomas of the stomach [29,30].
Additionally, functional activation of β-catenin/T-cell factor (Tcf) signaling has been implicated in human carcinogenesis. In cytoplasm, β-catenin contributes to cell-cell adhesion in cooperation with the cytoplasmic domain of E-cadherin, but β-catenin moves into the nucleus and possesses transcriptional activity in cooperation with the T-cell factor (Tcf)/lymphoid enhancer factor (Lef) transcription factor [31]. Activated β-catenin/Tcf signaling by the accumulation of β-catenin in the nucleus has been implicated in human carcinogenesis including colorectal cancer (CRC), melanoma, hepatocellular carcinoma, and gastric carcinoma [32,33,34]. One adenomatous polyposis coli (APC) mutation is observed in at least 60% of sporadic CRC cases and abnormalities in both APC alleles are shown in almost 30% of such cases [35]. Studies have reported the detection of APC mutations in 12 of 46 gastric cancers, with β-catenin nuclear localization occurring in both diffuse- and intestinal-type gastric cancers at a higher rate [36,37]. This means that the dysregulation of β-catenin plays a crucial role in some cancer cells. Thus, oncogenic transcription factors such as AP-1, myc-max and β-catenin/Tcf may present promising targets for cancer prevention.
NF-κB is also a protein complex transcription factor comprised of p50 and p65 or Rel. NF-κB is involved in cellular responses to stimuli such as oxidative stress and cytokines [38]. NF-kB is constitutively active in several cancer types and has been associated with the regulation of cell proliferation, cell survival, invasion, metastasis and inhibition of apoptosis [39,40]. It has been suggested that inhibition of NF-kB signaling suppresses tumor formation [41,42,43].
2. Symmetric Polyphenols
Curcumin, with a distinct symmetric polyphenol structure, is known to have diverse biological activities including antiinflammatory, antitumor, antioxidant, antifungal and antibacterial actions. Recently, a daily dose of curcumin was recommended for cancer patients [44]. In addition to curcumin, other types of symmetric polyphenols like dihydroguaiaretic acid (DHGA) and nordihydroguaiaretic acid (NDGA) blocked transcription factor and DNA binding (Table 1). Each bZIP domain of the jun and fos proteins used in electrophoresis mobility shift assay (EMSA) contain only a basic region and a leucine zipper region, not including dephosphorylation and phosphorylation sites activated by protein kinase C (PKC) and Jun N-terminal kinase (JNK) or Fyn-related kinase (FRK), respectively [45,46]. Thus, the possible inhibition sites of these inhibitors are limited to both basic regions and the leucine zippers of c-jun and c-fos proteins. To investigate whether functional group modification of the benzaldehyde moiety in the curcuminoid structure affects the inhibition of fos-jun and DNA complex formation, our group synthesized a variety of curcuminoids as shown in Table 2. Curcumin and some synthetic curcumin derivatives were found to be transcription factor inhibitors following EMSA experiments using in vitro expressed c-fos and c-jun proteins. The inhibitory actions of curcumin, curcuminoids, DHGA and NDGA on jun/AP-1 and DNA binding in cell nuclear extracts may be not via blocking the regulatory domain at c-jun sites that negatively regulate its DNA binding activity, but via direct interference with dimer binding to DNA. The inhibitors showed an inhibitory effect against the formation of the fos–jun–DNA complex in an in vitro assay with the IC50 values shown in Table 1 and Table 2. Curcumin and NDGA were revealed to have an IC50 value of around 0.3 mM in an in vitro assay, showing a more potent inhibitory effect on the formation of the fos–jun–DNA complex than DHGA. The change of the methoxy functional group of DHGA into a hydroxy group of NDGA caused a marked increase in the inhibitory activity. Remarkably, the synthetic curcumin derivative BJC005 showed the highest inhibitory activity out of all the synthetic curcuminoids tested. BJC005, with an IC50 of 5.4 μM, shows a much greater inhibitory effect than momordin, reported hitherto as the best fos-jun-DNA complex inhibitor [44,47]. However, some curcuminoids, such as CHC006 and BJC004, showed no inhibitory activity against fos-jun-DNA complex formation [44].
The myc and max proteins, containing basic regions and helix-loop-helix-zipper regions, were over-expressed in E. coli and used for the investigation into the inhibitors of myc-max-DNA complex formation. Curcuminoid 004, 005, and 008 demonstrated relatively selective inhibition of myc-max-DNA binding versus jun-fos-DNA complex formation. When assayed using the myc-max dimer protein expressed in E. coli, NDGA showed a weak inhibitory effect on the dimer and cognate DNA complex formation (~8 mM of IC50). This value represents about 50-fold specificity of NDGA to AP-1. However, the inhibitory effect of NDGA is more specific to β-catenin/Tcf than AP-1 or myc-max transcription factor [48,49]. This suggests that polyphenol compounds can be selective inhibitors against each transcription activator, therefore may be effective modulators of cell signaling and regulation.
Altogether, these results suggest that the substituted position of the benzaldehyde moiety and the polarity property of the substituted group play an important role in the inhibition. Regarding the inhibitory activity of curcuminoids on AP-1, p-position- and polar group-substituted curcuminoids have a tendency to inhibit the fos-jun-DNA complex formation more potently [44]. Considering that the insertion of a nitro group into the benzene ring at CHC 007, 008, 009, 011 and BJC005 contribute to efficient inhibitory activity, the substitution of polar groups in benzene rings is likely to confer a richly negative charge to the molecule itself, making them potent competitors to DNA for interaction with the protein.
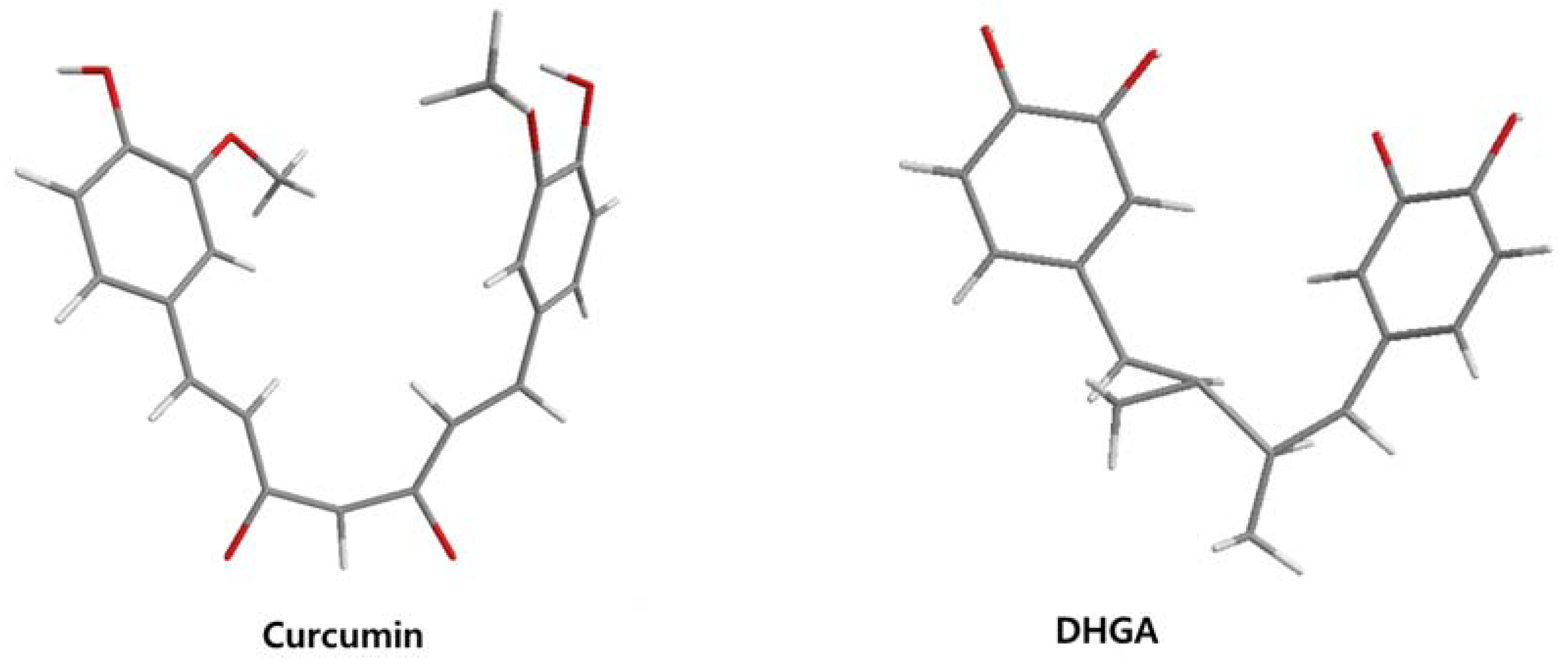
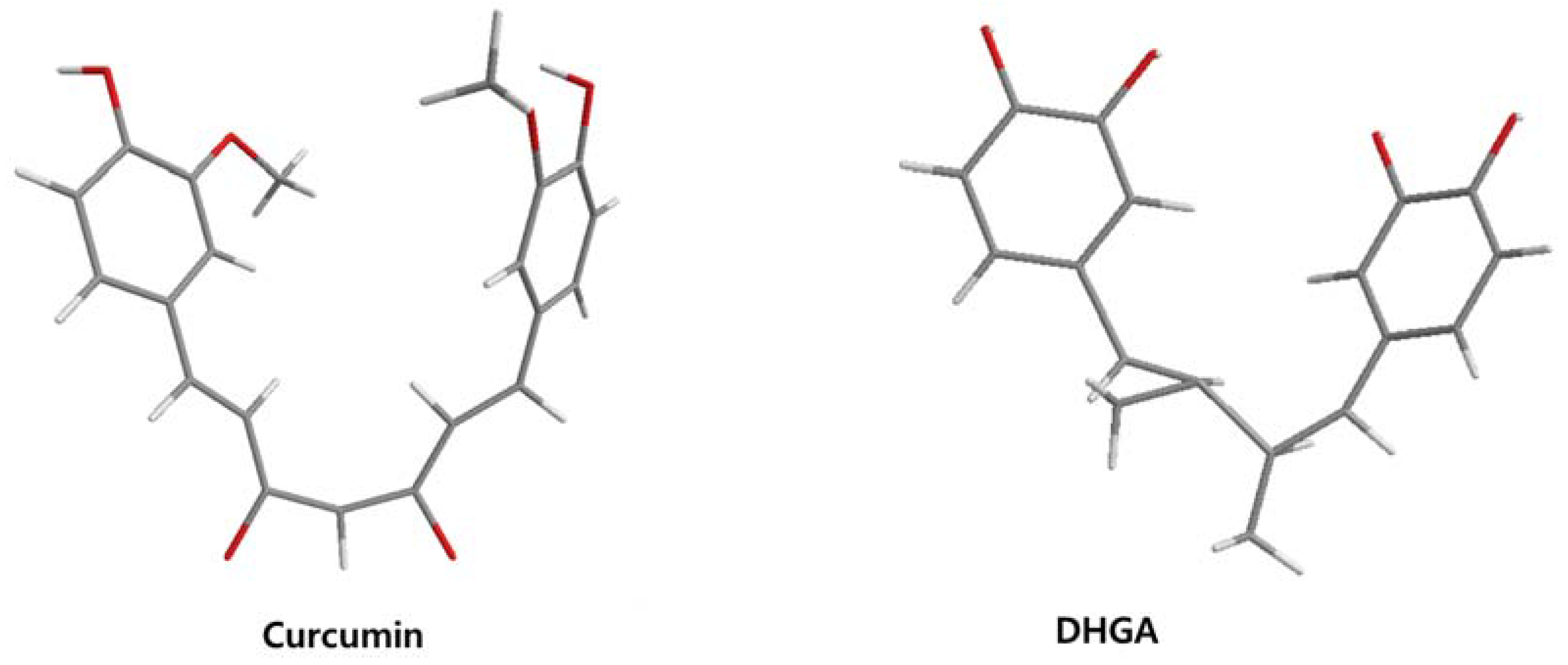
The low energy conformations of curcumin and DHGA inhibitors have “V-shaped” two-fold symmetry as shown in Figure 1 [49]. Conformational studies of these inhibitors revealed that they share a common conformational feature. The two-fold symmetry of molecular conformation is also a characteristic of fos-jun or myc-max transcription activators that function as dimer proteins. As a result, a stable binding mode was proposed due to two potential interactions between the protein and the inhibitor. One is a hydrogen bond and/or an electrostatic interaction between the acidic hydroxyl (OH)-group in the phenol ring of the inhibitor and the amino-group of the basic amino acid, which is abundant in the DNA binding region of the dimeric forms of the transcription factor. The other is a hydrophobic interaction between the hydrocarbon chain of the inhibitor and the hydrocarbon chain of the amino acid in the fos-jun protein [49]. The inhibitors have differences in their molecular structures. DHGA and NDGA have the same hydrophobic carbon chain. DHGA has one phenolic OH and one methoxy group on each side while NDGA has two phenolic OH groups on each side. The methoxy group of DHGA cannot form hydrogen bonds. This might make DHGA a less potent inhibitor than NDGA. Although curcumin has one phenolic OH on each side, the conjugated double bond of the hydrophobic carbon chain may cause curcumin to be more hydrophobic than the other two inhibitors. It was also suggested that the rigid conformation of curcumin provides a better acidic OH and hydrocarbon orientation for interactions with the fos-jun protein [49]. Actually, the lowest interaction energy for each inhibitor, which was obtained using Monte Carlo docking simulations, showed a correlation with the IC50 value of each inhibitor, which was experimentally determined.

{kind=link}
{kind=link}
| Name | Structure | IC50 (mM) | ||
|---|---|---|---|---|
| AP-1 | Myc/Max | β-catenin | ||
| DHGA | 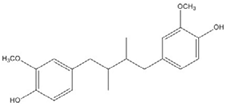 | 8.4 [45] | ||
| NDGA |  | 0.34 [45] | 8 [49] | 0.01 [48] |
| Curcumin | 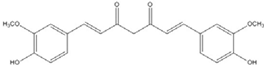 | 0.28 [45] | 0.6 [49] | 0.012 [46] |
| Asarinin |  | 0.3 [49] | ||
| Resveratrol | 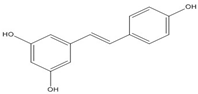 | 0.03 [50,51] | ||
| T-5224 |  | 2 [52] | ||
| Anthraquinone | 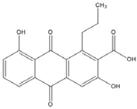 | 0.1 [53] | ||
DHGA: dihydroguaiaretic acid; NDGA: nordihydroguaiaretic acid; AP-1: Activator Protein-1.
Resveratrol and anthraquinone derivatives were also shown to inhibit the binding of AP-1 to the AP-1 binding site [50,51]. A new benzophenone derivative T-5224 was found to inhibit the DNA-binding activity of c-fos/c-jun, with IC50 values around 2 mM [52,53]. Although IC50 values around 2 mM is not very efficient, this compound exhibited a good specificity profile since the small-molecule did not affect the activities of other transcription factors including CCAAT-enhancer-binding proteins (C/EBP) and Activating Transcription Factor 2 (ATF-2) (bZIP domain) as well as MyoD (bHLH domain).
| Name | Structure | IC50 (mM) | ||
|---|---|---|---|---|
| Activator Protein-1 | Myc/Max | β-Catenin | ||
| Curcumin |  | 0.28 [44] | 0.6 [49] | 0.012 [54] |
| CHC001 | 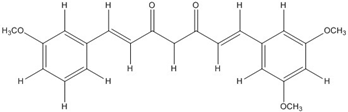 | 5.4 [44] | ||
| CHC002 |  | 6.7 [44] | ||
| CHC003 | 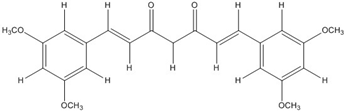 | 3.9 [44] | ||
| CHC004 | 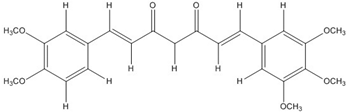 | 1.6 [44] | 0.082 [49] | |
| CHC005 | 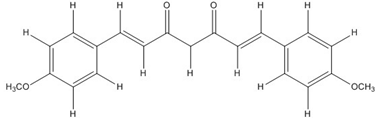 | 1.8 [44] | 0.4 [49] | |
| CHC006 | 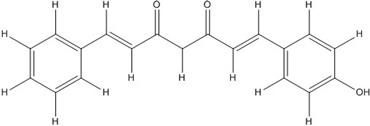 | No inhibition [44] | ||
| CHC007 |  | 0.38 [44] | 0.06 [54] | |
| CHC008 |  | 1.4 [44] | 0.16 [49] | |
| CHC009 |  | 0.64 [44] | ||
| CHC011 |  | 0.3 [44] | 0.43 [49] | |
| BJC004 |  | No inhibition [44] | ||
| BJC005 |  | 0.0054 [44] |
Curcumin and synthetic curcumin derivatives were also suggested to be β-catenin/Tcf signaling inhibitors [54,55,56]. In our previous reports, curcumin and some synthetic derivatives inhibited β-catenin/Tcf signaling in cell-based luciferase assays and their inhibitory mechanisms were found to be related to the reduced binding of nuclear β-catenin and Tcf-4 proteins [54]. 20 μM curcumin and 100 μM CHC007 inhibited β-catenin/Tcf signaling by 58%–63% and 70%–78%, respectively, compared with the control showing variations in a variety of cell lines [54].
Figure 1.
The lowest energy conformation of curcumin and DHGA.
3. Flavonoids
Many flavonoid target pathways related to intracellular signaling have been suggested and reviewed [57]. Although flavonoids were suggested as potential wnt/beta-catenin signaling modulators in cancer, all of the flavonoids did not show the same mechanism. Also, some compounds showed different mechanisms in different component targets. For instance, flavonoids like genistein, tangeretin, epigallocathechin gallate (EGCG), quercetin, apigenin and isoquercitrin were reported to modulate wnt/β-catenin signaling [57,58,59,60,61,62,63,64]. But these flavonoids do not all affect the same targets. Genistein seems to inhibit not only wnt/receptor binding but also Akt action, leading to β-catenin activation [54,65]. Moreover, genistein can inhibit the binding of β-catenin/Tcf complexes with consensus DNA [66]. Quercetin not only targets Akt and MAPK but also interferes with the binding of Tcf complexes to DNA [57,65,67,68]. Flavonoid compounds that disrupt the transcription factor complex are listed in Table 3.
Common flavonols including quercetin, kaempferol, and isorhamnetin as well as flavones such as baicalein and one of several known isoflavones, genistein, showed inhibitory effects in β-catenin activated cells [66,67]. The mechanism underlying the reduction of β-catenin/Tcf transcriptional activity by quercetin, kaempferol, isorhamnentin, baicalein and genistein was due to the decreased binding of β-catenin/Tcf complexes with consensus DNA. EMSA data showed that the binding of Tcf complexes to DNA was impaired by inhibitors. Conforming to these results, coimmunoprecipitation of β-catenin and Tcf-4 using Tcf-4 antibodies showed that the association of β-catenin with Tcf-4 was blocked by inhibitors. With genistein, the decreased distribution of β-catenin in the nucleus caused by decreased phosphorylation of Akt and GSK3b contributed as a partial mechanism for reduced β-catenin/Tcf transcriptional activity [66]. Thus, these results agree with the earlier reports by Jaiswal et al. and Sarkar et al. [54,65].
| Name | Structure | AP-1 | IC50 (mM) Myc/Max | β-catenin | NF-κB |
|---|---|---|---|---|---|
| Quercetin | 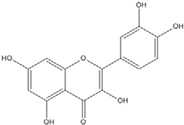 | 0.0076 [67] | 0.03 [69] | ||
| Genistein | 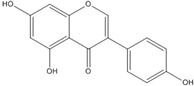 | 0.01 [66] | |||
| Kaempferol |  | 0.0029 [49] | 2 [49] | 0.0016 [66] | 0.003 [70] |
| Isorhamnetin | 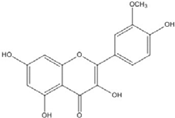 | 0.0062 [66] | |||
| Baicalein | 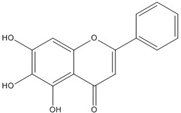 | 0.00026 [66] | |||
| Naringenin | 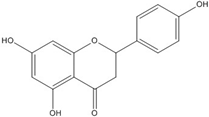 | No inhibition [71] | |||
| Epigallo catechingallate |  | No inhibition [49] | |||
| Sterptonigrin | 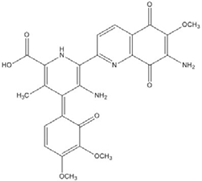 | 0.005 [72] | |||
| Tanshinone I | 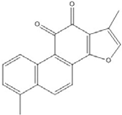 | 2.6 [49] | |||
| 7,8-dihydroxy flavanone | 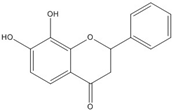 | 1.8 [49] | |||
| Chrysin | 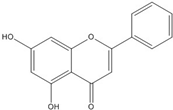 | 0.2 [53] | |||
| Apigenin | 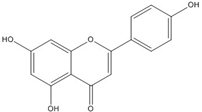 | 0.1 [53] | ~0.002 [53] | ||
| Luteolin | 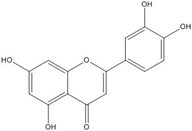 | 0.1 [53] | ~0.002 [53] | ||
| Flavanone |  | No inhibition [71] | >0.003 [53] |
AP-1: Activator Protein-1; NF-κB : Nuclear factor kappa-light-chain-enhancer of activated B cells.
Although flavonoids such as 7,8-dihydroxyflavanone showed inhibitory activity against jun-fos/DNA binding, the inhibitory action was not as effective as on β-catenin/Tcf, with a 1.8 mM IC50 value obtained from EMSA [49]. Although there are few reports on the inhibitory effect of flavonoids against AP-1 or other transcription factors, our unpublished data showed that kaempferol inhibited fos-jun-DNA binding and myc-max-DNA binding with IC50 values of 2.9 mM and 2 mM, respectively [49]. Also, tanshinone I and 7,8 dihydroxyflavanone showed inhibitory activities against fos-jun-DNA binding, with IC50 values of 2.6 and 1.8 mM, respectively [49]. All these data demonstrate that flavonoids show selective inhibitory activity against β-catenin/Tcf rather than against the AP-1 transcription factor. However, quercetin, kaempferol, apigenin, luteolin and chrysin were reported to inhibit the NF-kB transcription factor by interfering with complex formation [69,70]. Interestingly, flavanone and flavonoids that include a flavanone skeleton such as naringenin and EGCG showed no inhibitory activity on the direct binding of β-catenin/Tcf with DNA ([49,68,71].
Although genistein, kaempferol, isorhamnentin and baicalein possess a common phenylbenzopyrone structure (C6–C3–C6), they are mainly categorized into flavanols (kaempferol and isorhamnentin), flavone (baicalein), and isoflavone (genistein) according to the substitution state and direction of the phenyl ring. Genistein showed the least potent blocking activity in EMSA while baicalein showed the most [66]. Considering the relationship between the structure and β-catenin/Tcf inhibitory activity, the direction of the B phenyl ring at the 2-subtituted site (flavanol and flavone) appears to be more effective than at the 3-substituted (isoflavone) site. It also seems that hydroxyl group substitution at the 3-site (flavanol) reduces the potency of inhibition compared to without substitution (flavone).
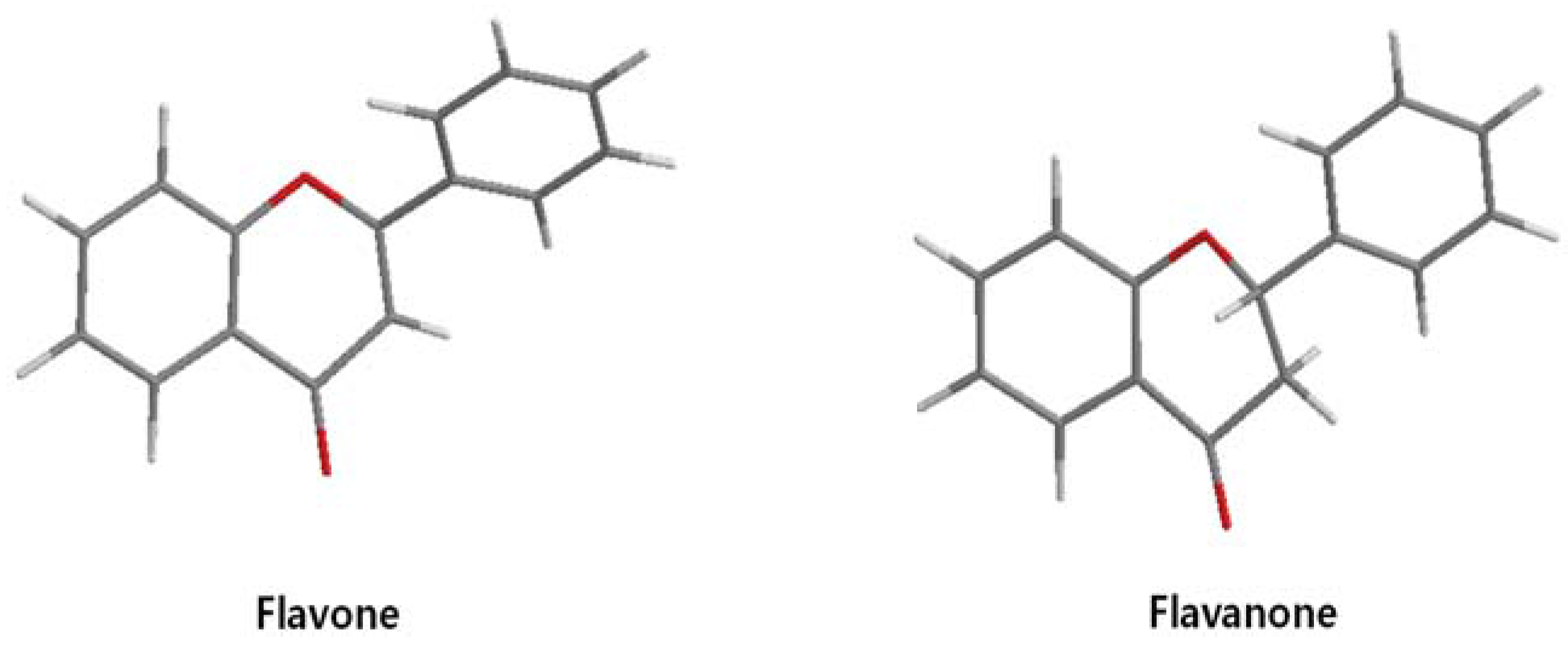
On the other hand, flavanone and flavonoids that include the same skeleton such as naringenin did not show inhibitory activity against β-catenin/Tcf and DNA binding, in discord with other types of flavonoids [68,71]. From the three dimensional structure, the 2-substituted B ring of flavanone seems to be puckered from the plane, with a different shape from the other flavone skeletons containing conjugated double bonds of C3 rings as shown in Figure 2. This puckered structure may be unfit to block β-catenin/Tcf and DNA complex formation.
Figure 2.
Flavone and flavanone.

4. Non Flavonoid Polyphenol β-catenin Inhibitors
Because wnt/β-catenin signaling is involved in a broad range of biological systems, small molecular inhibitors of wnt/β-catenin signaling have received attention as targets for therapeutic interventions. A variety of inhibitors against other components in wnt/β-catenin signaling were recently reviewed [73]. Out of several small molecules, polyphenol compounds that disrupt the β-catenin/Tcf-4 complex are listed in Table 4.
| Compound | Structure | β-Catenin IC50 (μM) |
|---|---|---|
| PKF118-744 |  | 2.4 [73,74,75,76] |
| CGP049090 |  | 8.7 [73,74,75,76] |
| PKF118-310 | 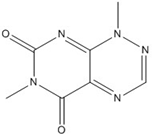 | 0.8 [73,74,75,76] |
| ZTM00990 |  | 0.64 [73,74,75,76] |
| BC21 |  | Not Determined [73,77] |
| Ethacrynic acid | 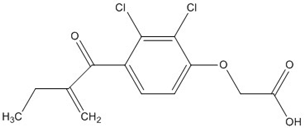 | ~70 [73,78,79] |
| Ethacrynic acid derivative | 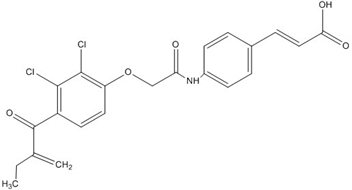 | ~5 [73,79] |
| PKF115-584 | 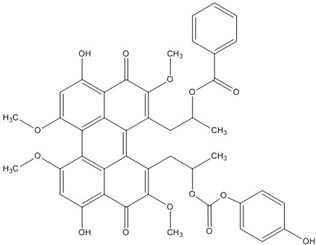 | 3.2 [73,74,75,76] |
| PNU-74654 |  | Not Determined [73,74,75,76] |
5. Conclusions
A useful starting point for investigations into target-specific drug design is to identify effective targets that elicit responses in physiological states and to compile an abundant candidate library of effective leading compounds. Polyphenol compounds are abundant and ideally include more comprehensive structural variance, with effects on intracellular signaling induced by the altered responsiveness (for instance, inhibition against tumor necrosis factors (TNF)-induced, transforming growth factor (TGF)-induced, α-melanocyte-stimulating hormone (α-MSH)-induced, oxidative stress-induced and UV-induced signaling).
A general concern is that many of the targets for intracellular signaling may play vital roles in other signaling cascades. Moreover, a critical issue that should be addressed is whether targeting of the dimeric forms of transcription factors is sufficiently specific. For instance, AP-1, myc-max, NF-κB and β-catenin/Tcf transcription factors play specific roles in a variety of signaling pathways. Therefore, broad inhibition of the transcription factors above is predicted to cause toxicity; this is why specificity will be an important determinant of the success of transcription factor inhibiting drugs. Thus, whether the inhibition of AP-1, myc-max, NF-κB and β-catenin/Tcf can be discriminated by chemical inhibitors remains to be explored. Theoretically, DNA can form very large interfaces with proteins and the buried surfaces of the protein–nucleic acid interfaces have a broad range of sizes [80]. Usually, protein–protein recognition sites are formed by protein surfaces with good shape and electrostatic complementarity [81]. Protein–protein interfaces are frequently hydrophobic and bury a large portion of the nonpolar surface area [82]. However, in protein-DNA interaction, the DNA binding surface of the protein harbors basic amino acids because of the negative charge of DNA. It has been reported that standard sizes for the interface range from 1200–2000 A˚ [83]. The interface between transcription factor and DNA is enough for multi-facial binding of polyphenol compounds showing approximate sizes from 7 to 10 A˚.
This review discusses polyphenol compounds with roles in direct inhibition against transcription factor complexes such as AP-1, myc-max, NF-κB and β-catenin/Tcf and suggests structure-activity relationships for the development of small-molecule drug candidates that modulate this clinically relevant signaling pathway. The structural diversity of curcumin derivative compounds or flavonoids may provide specificity for recognizing transcription factors. Polyphenol groups have structural variability and abundance in nature and show distinctive inhibitory activities against various transcription factors. Altogether, polyphenol transcription factor inhibitors have significant value as clinical drug candidates. However, another aspect that has to be considered is that some polyphenol compounds that reach cells and tissues are chemically different from the original dietary form because of structural modifications due to the conjugation process that takes place in the small intestine and, mostly, in the liver [84]. As animal trials are performed in future studies, they will provide extremely valuable information and overcome some of the difficulties associated with bioavailability in humans as clinical drug candidates.
References
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [PubMed]
- El Gharras, H. Polyphenols: Food sources, properties and applications—A review. Int. J. Food Sci. Tech. 2009, 44, 2512–2518. [Google Scholar] [CrossRef]
- Curran, T.; Franza, B.R., Jr. Fos and Jun: The AP-1 connection. Cell 1988, 55, 395–397. [Google Scholar] [CrossRef]
- Johnson, P.F.; McKnight, S.L. Eukaryotic transcriptional regulatory proteins. Annu. Rev. Biochem. 1989, 58, 799–839. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.J.; Tjian, R. Transcriptional regulation in mammalian cells by sequence-specific DNA binding proteins. Science 1989, 245, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Mitchell, P.; Tjian, R. Purified transcription factor AP-1 interacts with TPA-inducible enhancer elements. Cell 1987, 49, 741–752. [Google Scholar] [CrossRef]
- Angel, P.; Imagawa, M.; Chiu, R.; Stein, B.; Imbra, R.J.; Rahmsdorf, H.J.; Jonat, C.; Herrlich, P.; Karin, M. Phorbol ester-inducible genes contain a common cis element recognized by a TPA-modulated trans-acting factor. Cell 1987, 49, 729–739. [Google Scholar] [CrossRef]
- Schutte, J.; Nau, M.; Birrer, M.; Thomas, F.; Gazdar, A.; Minna, J. Constitutive expression of multiple mRNA forms of the c-jun oncogene in human lung cancer cell lines. Proc. Am. Assoc. Cancer Res. 1988, 1808, 455–462. [Google Scholar]
- Bossy-Wetzel, E.; Bravo, R.; Hanahan, D. Transcription factors junB and c-jun are selectively up-regulated and functionally implicated in fibrosarcoma development. Genes Dev. 1992, 6, 2340–2351. [Google Scholar] [CrossRef] [PubMed]
- Mattei, M.G.; Simon-Chazottes, D.; Hirai, S.; Ryseck, R.P.; Galcheva-Gargova, Z.; Guénet, J.L.; Mattei, J.F.; Bravo, R.; Yaniv, M. Chromosomal localization of the three members of the jun proto-oncogene family in mouse and man. Oncogene 1990, 5, 151–156. [Google Scholar] [PubMed]
- Schutte, J.; Minna, J.D.; Birrer, M.J. Deregulated expression of human c-jun transforms primary rat embryo cells in cooperation with an activated c-Ha-ras gene and transforms Rat-1a cells as a single gene. Proc. Natl. Acad. Sci. USA 1989, 86, 2257–2261. [Google Scholar] [CrossRef] [PubMed]
- Van der Burg, B.; van Selm-Miltenburg, A.J.P.; de Laat, S.W.; van Zoolen, F.J.J. Direct effects of estrogen on c-fos and c-myc protooncogene expression and cellular proliferation in human mammary breast cancer cells. Mol. Cell. Endocrinol. 1989, 64, 223–228. [Google Scholar] [CrossRef]
- Baichwal, V.R.; Tjian, R. Control of c-Jun activity by interaction of a cell-specific inhibitor with regulatory domain delta: Differences between v- and c-Jun. Cell 1990, 63, 815–825. [Google Scholar] [CrossRef]
- Persson, A.E.; Pontén, I.; Cotgreave, I.; Jernström, B. Inhibitory effects on the DNA binding of AP-1 transcription factor to an AP-1 binding site modified by benzo[α]pyrene 7,8-dihydrodiol 9,10-epoxide diastereomers. Carcinogenesis 1996, 17, 1963–1969. [Google Scholar] [CrossRef] [PubMed]
- Dalla-Favera, R.; Wong-Staal, F.; Gallo, R.C. Onc gene amplification in promyelocytic leukaemia cell line HL-60 and primary leukaemic cells of the same patient. Nature 1982, 299, 61–63. [Google Scholar] [CrossRef] [PubMed]
- Magrath, I. The pathogenesis of Burkitt’s lymphoma. Adv. Cancer Res. 1990, 55, 133–270. [Google Scholar] [PubMed]
- Payne, G.S.; Bishop, J.M.; Varmus, H.E. Multiple arrangements of viral DNA and an activated host oncogene in bursal lymphomas. Nature 1982, 295, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Neil, J.C.; Forrest, D.; Doggett, D.L.; Mullins, J.I. The role of feline leukaemia virus in naturally occurring leukaemias. Cancer Surviv. 1987, 6, 117–137. [Google Scholar]
- Eilers, M.; Schirm, S.; Bishop, J.M. The MYC protein activates transcription of the alpha-prothymosin gene. EMBO J. 1991, 10, 133–141. [Google Scholar] [PubMed]
- Penn, L.J.; Laufer, E.M.; Land, H. C-MYC: Evidence for multiple regulatory functions. Semin. Cancer Biol. 1990, 1, 69–80. [Google Scholar] [PubMed]
- Land, H.; Parada, L.F.; Weinberg, R.A. Tumorigenic conversion of primary embryo fibroblasts requires at least two cooperating oncogenes. Nature 1983, 304, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Coppola, J.A.; Cole, M.D. Constitutive c-myc oncogene expression blocks mouse erythroleukaemia cell differentiation but not commitment. Nature 1986, 320, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.H.; Wold, B.J. c-myc inhibition of MyoD and myogenin-initiated myogenic differentiation. Mol. Cell. Biol. 1991, 11, 2842–2851. [Google Scholar] [CrossRef] [PubMed]
- Little, C.D.; Nau, M.M.; Carney, D.N.; Gazdar, A.F.; Minna, J.D. Amplification and expression of the c-myc oncogene in human lung cancer cell lines. Nature 1983, 306, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Escot, C.; Theillet, C.; Lidereau, R.; Spyratos, F.; Champeme, M.H.; Gest, J.; Callahan, R. Genetic alteration of the c-myc protooncogene (MYC) in human primary breast carcinomas. Proc. Natl. Acad. Sci. USA 1986, 83, 4834–4838. [Google Scholar] [CrossRef] [PubMed]
- Pinion, S.B.; Kennedy, J.H.; Miller, R.W.; MacLean, A.B. Oncogene expression in cervical intraepithelial neoplasia and invasive cancer of cervix. Lancet 1991, 337, 819–820. [Google Scholar] [CrossRef]
- Kim, Y.S.; Yoo, H.S.; Lee, K.T.; Goh, S.H.; Jung, J.S.; Oh, S.W.; Baba, M.; Yasuda, T.; Matsubara, K.; Nagai, H.; et al. Detection of genetic alterations in the human gastric cancer cell lines by two-dimensional analysis of genomic DNA. Int. J. Oncol. 2000, 17, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Morgenbesser, S.D.; DePinho, R.A. Use of transgenic mice to study myc family gene function in normal mammalian development and in cancer. Semin. Cancer Biol. 1994, 5, 21–36. [Google Scholar] [PubMed]
- Pelengaris, S.; Rudolph, B.; Littlewood, T. Action of Myc in vivo—Proliferation and apoptosis. Curr. Opin. Genet. Dev. 2000, 10, 100–105. [Google Scholar] [CrossRef]
- Bullions, L.C.; Levine, A.J. The role of beta-catenin in cell adhesion, signal transduction, and cancer. Curr. Opin. Oncol. 1998, 10, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Morin, P.J.; Sparks, A.B.; Korinek, V.; Barker, N.; Clevers, H.; Vogelstein, B.; Kinzler, K.W. Activation of b-catenin-Tcf signaling in colon cancer by mutations in β-catenin or APC. Science 1997, 275, 1787–1790. [Google Scholar] [CrossRef] [PubMed]
- Fujie, H.; Moriya, K.; Shintani, Y.; Tsutsumi, T.; Takayama, T.; Makuuchi, M.; Kimura, S.; Koike, K. Frequent β-catenin aberration in human hepatocellular carcinoma. Hepatol. Res. 2001, 20, 39–51. [Google Scholar] [CrossRef]
- Woo, D.K.; Kim, H.S.; Lee, H.S.; Kang, Y.H.; Yang, H.K.; Kim, W.H. Altered expression and mutation of β-catenin gene in gastric carcinomas and cell lines. Int. J. Cancer 2001, 95, 108–113. [Google Scholar] [CrossRef]
- Powell, S.M.; Zilz, N.; Beazer-Barclay, Y.; Bryan, T.M.; Hamilton, S.R.; Thibodeau, S.N.; Vogelstein, B.; Kinzler, K.W. APC mutations occur early during colorectal tumorigenesis. Nature 1992, 359, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuru, S.; Yanagisawa, A.; Ichii, S.; Tahara, E.; Kato, Y.; Nakamura, Y.; Horii, A. Somatic mutation of the APC gene in gastric cancer: Frequent mutations in very well differentiated adenocarcinoma and signet-ring cell carcinoma. Hum. Mol. Genet. 1992, 1, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Clements, W.M.; Wang, J.; Sarnaik, A.; Kim, O.J.; MacDonald, J.; Fenoglio-Preiser, C.; Groden, J.; Lowy, A.M. β-catenin mutation is a frequent cause of Wnt pathway activation in gastric cancer. Cancer Res. 2002, 62, 3503–3506. [Google Scholar] [PubMed]
- Tian, B.; Brasier, A.R. Identification of a nuclear factor κB-dependent gene network. Recent Prog. Horm. Res. 2003, 58, 95–130. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.W. NF-κB in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Mees, C.; Nemunaitis, J.; Senzer, N. Transcription factors: Their potential as targets for an individualized therapeutic approach to cancer. Cancer Gene Ther. 2009, 16, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.Q.; Abbruzzese, J.L.; Lin, E.; Wang, L.; Zheng, L.; Xie, K. NF-κB activity blockade impairs the angiogenic potential of human pancreatic cancer cells. Int. J. Cancer 2004, 108, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Brickman, J.M.; Adam, M.; Ptashne, M. Interactions between an HMG-1 protein and members of the Rel family. Proc. Natl. Acad. Sci. USA 1999, 96, 10679–10683. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J. Cellular senescence as a tumor suppressor mechanism. Trends Cell Biol. 2001, 11, S27–S31. [Google Scholar] [CrossRef]
- Hahm, E.R.; Cheon, G.; Lee, J.; Kim, B.; Park, C.; Yang, C.H. New and known symmetrical curcumin derivatives inhibit the formation of Fos-Jun-DNA complex. Cancer Lett. 2002, 184, 89–96. [Google Scholar] [CrossRef]
- Park, S.; Lee, D.K.; Yang, C.H. Inhibition of fos-jun-DNA complex formation by dihydroguaiaretic acid and in vitro cytotoxic effects on cancer cells. Cancer Lett. 1998, 127, 23–28. [Google Scholar] [CrossRef]
- Hahm, E.R.; Gho, Y.S.; Park, S.; Park, C.; Kim, K.W.; Yang, C.H. Synthetic curcumin analogs inhibit activator protein-1 transcription and tumor-induced angiogenesis. Biochem. Biophys. Res. Commun. 2004, 321, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Kim, B.; Lee, S.G.; Gwon, H.J.; Moon, E.Y.; Hwang, H.S.; Seong, S.K.; Lee, M.; Lim, M.J.; Sung, H.J.; et al. Momordins inhibit both AP-1 function and cell proliferation. Anticancer Res. 1998, 18, 119–124. [Google Scholar] [PubMed]
- Park, S.; Lee, J. Inhibitory effect of nordihydroguaiaretic acid on β-catenin/Tcf signalling in β-catenin-activated cells. Cell Biochem. Funct. 2011, 29, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Park, S. Studies on the Inhibitory Mechanism of the Dimeric Forms of Transcription Activators. Ph.D. Thesis, Seoul National University, Seoul, Korea, 2002. [Google Scholar]
- Yu, R.; Hebbar, V.; Kim, D.W.; Mandlekar, S.; Pezzuto, J.M.; Kong, A.N. Resveratrol inhibits phorbol ester and UV-induced activator protein 1 activation by interfering with mitogen-activated protein kinase pathways. Mol. Pharmacol. 2001, 60, 217–224. [Google Scholar] [PubMed]
- Goto, M.; Masegi, M.; Yamauchi, T.; Chiba, K.; Kuboi, Y.; Harada, K.; Naruse, N. K1115 A, a new anthraquinone derivative that inhibits the binding of activator protein-1 (AP-1) to its recognition sites. I. Biological. activities. J. Antibiot. 1998, 51, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, Y.; Morimoto, K.; Yamamoto, T.; Chaki, H.; Hashiramoto, A.; Narita, H.; Hirono, S.; Shiozawa, S. Treatment of arthritis with a selective inhibitor of c-Fos/activator protein-1. Nat. Biotechnol. 2008, 26, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Yap, J.L.; Chauhan, J.; Jung, K.Y.; Chen, L.; Prochownik, E.V.; Fletcher, S. Small-molecule inhibitors of dimeric transcription factors: Antagonism of protein–protein and protein–DNA interactions. Med. Chem. Commun. 2012, 3, 541–551. [Google Scholar] [CrossRef]
- Park, C.H.; Hahm, E.R.; Park, S.; Kim, H.K.; Yang, C.H. The inhibitory mechanism of curcumin and its derivative against beta-catenin/Tcf signaling. FEBS Lett. 2005, 579, 2965–2971. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.S.; Marlow, B.P.; Gupta, N.; Narayan, S. Beta-catenin-mediated transactivation and cell-cell adhesion pathways are important in curcumin (diferuylmethane)-induced growth arrest and apoptosis in colon cancer cells. Oncogene 2002, 21, 8414–8427. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, N.N.; Carothers, A.M.; Grunberger, D.; Bilinski, R.T.; Churchill, M.R.; Martucci, C.; Newmark, H.L.; Bertagnolli, M.M. Plant phenolics decrease intestinal tumors in an animal model of familial adenomatous polyposis. Carcinogenesis 2000, 21, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Amado, N.G.; Fonseca, B.F.; Cerqueira, D.M.; Neto, V.M.; Abreu, J.G. Flavonoids: Potential Wnt/beta-catenin signaling modulators in cancer. Life Sci. 2011, 89, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Song, D.H.; Sussman, D.J.; Seldin, D.C. Endogenous protein kinase CK2 participates in Wnt signaling in mammary epithelial cells. J. Biol. Chem. 2000, 275, 23790–23797. [Google Scholar] [CrossRef] [PubMed]
- Landesman-Bollag, E.; Song, D.H.; Romieu-Mourez, R.; Sussman, D.J.; Cardiff, R.D.; Sonenshein, G.E.; Seldin, D.C. Protein kinase CK2: Signaling and tumorigenesis in the mammary gland. Mol. Cell. Biochem. 2001, 227, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Zhang, X.; Rieger-Christ, K.M.; Summerhayes, I.C.; Wazer, D.E.; Paulson, K.E.; Yee, A.S. Suppression of Wnt signaling by the green tea compound (−)−epigallocatechin 3-gallate (EGCG) in invasive breast cancer cells. Requirement of the transcriptional repressor HBP1. J. Biol. Chem. 2006, 281, 10865–10875. [Google Scholar] [CrossRef] [PubMed]
- Dashwood, W.M.; Orner, G.A.; Dashwood, R.H. Inhibition of beta-catenin/Tcf activity by white tea, green tea, and epigallocatechin-3-gallate (EGCG): Minor contribution of H2O2 at physiologically relevant EGCG concentrations. Biochem. Biophys. Res. Commun. 2002, 296, 584–588. [Google Scholar] [CrossRef]
- Pahlke, G.; Ngiewih, Y.; Kern, M.; Jakobs, S.; Marko, D.; Eisenbrand, G. Impact of quercetin and EGCG on key elements of the Wnt pathway in human colon carcinoma cells. J. Agric. Food Chem. 2006, 54, 7075–7082. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Xu, Z.; Hung, M.S.; Lin, Y.C.; Wang, T.; Gong, M.; Zhi, X.; Jablon, D.M.; You, L. Promoter demethylation of WIF-1 by epigallocatechin-3-gallate in lung cancer cells. Anticancer Res. 2009, 29, 2025–2030. [Google Scholar] [PubMed]
- Mount, J.G.; Muzylak, M.; Allen, S.; Althnaian, T.; McGonnell, I.M.; Price, J.S. Evidence that the canonical Wnt signalling pathway regulates deer antler regeneration. Dev. Dyn. 2006, 235, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, F.H.; Li, Y.; Wang, Z.; Kong, D. Cellular signaling perturbation by natural products. Cell Signal. 2009, 21, 1541–1547. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Choi, J. Inhibition of beta-catenin/Tcf signaling by flavonoids. J. Cell. Biochem. 2010, 110, 1376–1385. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Chang, J.Y.; Hahm, E.R.; Park, S.; Kim, H.K.; Yang, C.H. Quercetin, a potent inhibitor against beta-catenin/Tcf signaling in SW480 colon cancer cells. Biochem. Biophys. Res. Commun. 2005, 328, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Hahm, E.R.; Lee, J.H.; Jung, K.C.; Yang, C.H. Inhibition of beta-catenin-mediated transactivation by flavanone in AGS gastric cancer cells. Biochem. Biophys. Res. Commun. 2005, 331, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.D.; Choi, C.H.; Bark, H.; Son, H.Y.; Park, H.H.; Lee, S.; Park, J.W.; Park, E.K.; Shin, H.I.; Kim, S.H.; et al. Quercetin inhibits expression of inflammatory cytokines through attenuation of NF-κB and p38 MAPK in HMC-1 human mast cell line. Inflamm. Res. 2007, 56, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Chow, M.P.; Huang, W.C.; Lin, Y.C.; Chang, Y.J. Flavonoids inhibit tumor necrosis factor-alpha-induced up-regulation of intercellular adhesion molecule-1 (ICAM-1) in respiratory epithelial cells through activator protein-1 and nuclear factor-κB: Structure-activity relationships. Mol. Pharmacol. 2004, 66, 683–693. [Google Scholar] [PubMed]
- Lee, J.H.; Park, C.H.; Jung, K.C.; Rhee, H.S.; Yang, C.H. Negative regulation of β-catenin/Tcf signaling by naringenin in AGS gastric cancer cell. Biochem. Biophys. Res. Commun. 2005, 335, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Chun, S. Streptonigrin inhibits β-Catenin/Tcf signaling and shows cytotoxicity in β-catenin-activated cells. Biochem. Biophys. Acta 2011, 1810, 1340–1345. [Google Scholar] [CrossRef] [PubMed]
- Voronkov, A.; Krauss, S. Wnt/β-catenin signaling and small molecule inhibitors. Curr. Pharm. Des. 2013, 19, 634–664. [Google Scholar] [CrossRef] [PubMed]
- Lepourcelet, M.; Chen, Y.N.; France, D.S.; Lubecka, B.; Matulewicz, L.; Maniakowski, Z.; Polaniak, R.; Birkner, E.; Rzeszowska-Wolny, J. Small-molecule antagonists of the oncogenic Tcf/β-catenin protein complex. Cancer Lett. 2004, 5, 91–102. [Google Scholar] [CrossRef]
- Barker, N.; Clevers, H. Mining the Wnt pathway for cancer therapeutics. Nat. Rev. Drug. Discov. 2006, 5, 997–1014. [Google Scholar] [CrossRef] [PubMed]
- Trosset, J.Y.; Dalvit, C.; Knapp, S.; Fasolini, M.; Veronesi, M.; Mantegani, S.; Gianellini, L.M.; Catana, C.; Sundström, M.; Stouten, P.F.; et al. Inhibition of protein-protein interactions: The discovery of druglike β-catenin inhibitors by combining virtual and biophysical screening. Proteins 2006, 64, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Han, X.; Yan, M.; Xu, Y.; Duggineni, S.; Lin, N.; Luo, G.; Li, Y.M.; Han, X.; Huang, Z.; et al. Structure-based discovery of a novel inhibitor targeting the β-catenin/Tcf4 interaction. Biochemistry 2012, 51, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Liu, J.X.; Endo, T.; Zhou, H.; Yao, S.; Willert, K.; Schmidt-Wolf, I.G.; Kipps, T.J.; Carson, D.A. Ethacrynic acid exhibits selective toxicity to chronic lymphocytic leukemia cells by inhibition of the Wnt/beta-catenin pathway. PLoS ONE 2009, 4, e8294. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Lu, D.; Yao, S.; Wu, C.C.; Liu, J.X.; Carson, D.A.; Cottam, H.B. Amide derivatives of ethacrynic acid: Synthesis and evaluation as antagonists of Wnt/β-catenin signaling and CLL cell survival. Bioorganic Med. Chem. Lett. 2009, 19, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Moreira, I.S.; Fernandes, P.A.; Ramos, M.J. Hot spots—A review of the protein-protein interface determinant amino-acid residues. Proteins 2007, 68, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Young, L.; Jernigan, R.L.; Covell, D.G. A role for surface hydrophobicity in protein-protein recognition. Protein Sci. 1994, 3, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Horton, N.; Lewis, M. Calculation of the free energy of association for protein complexes. Protein Sci. 1992, 1, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Janin, J.; Bahadur, R.P.; Chakrabarti, P. Protein-protein interaction and quaternary structure. Q. Rev. Biophys. 2008, 41, 133–180. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the Polyphenols: Status and Controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, S. Polyphenol Compound as a Transcription Factor Inhibitor. Nutrients 2015, 7, 8987-9004. https://doi.org/10.3390/nu7115445
AMA Style
Park S. Polyphenol Compound as a Transcription Factor Inhibitor. Nutrients. 2015; 7(11):8987-9004. https://doi.org/10.3390/nu7115445
Chicago/Turabian StylePark, Seyeon. 2015. "Polyphenol Compound as a Transcription Factor Inhibitor" Nutrients 7, no. 11: 8987-9004. https://doi.org/10.3390/nu7115445