1. Introduction
In adults, epigenetic changes in DNA can occur as a result of exposure to various environmental factors, including diet, tobacco, physical activity, stress, drugs, and carcinogens [
1,
2]. It is widely understood that epigenetic modifications in utero can occur as an adaptive response to suitably prepare the fetus for the expected postnatal environment, thereby giving them the best chance at survival [
3,
4]. Environmental factors during fetal development have been shown to influence variable DNA methylation patterns, which have been implicated in the determination of long-term health for the offspring [
5,
6], although causal relationships have not been established.
Adverse conditions in utero, such as smoking exposure during pregnancy, have been shown to influence epigenetic patterns in the offspring. A comprehensive review by Knopik et al. investigating maternal smoking during pregnancy and child epigenetic patterns identified that smoking during pregnancy was associated with alterations in placental and cord blood-derived DNA methylation patterns [
7]. Research has also shown that distinct markers of smoking in pregnancy persist in the methylome of offspring at five years of age [
8]. This analysis replicated findings from studies carried out in both younger and older age groups, which highlight the postnatal stability of DNA methylation changes that occur in utero. According to the Developmental Origins of Health and Disease (DOHaD) concept, both under- and over-nutrition during pregnancy may program adaptations of the fetus in order to survive in an environment that may be either lacking or be abundant in particular nutrients [
9]. Some of the most substantial research relating to long-lasting impacts of maternal diet during pregnancy on offspring in later life tends to focus on nutrient-depleted populations. This is clearly demonstrated by the severe energy deficiency experienced during the famine in the Dutch Hunger Winter [
10]. Research on this cohort has shown differential DNA methylation patterns in offspring aged 59 years whose mothers were exposed to the famine during their pregnancy [
11,
12].
Recently, research has also begun to focus on the impact of over-nutrition during pregnancy and other dietary factors that are found in developed countries. In relation to the maternal metabolic environment, exposure to adverse conditions, like gestational diabetes or maternal obesity, is increasing and is estimated to affect 10–20% of pregnancies [
13]. One study identified epigenome-wide changes in teenage children whose mothers had gestational diabetes during their pregnancy [
14]. Altered or increased glucose levels in utero may have an impact on the developing fetal methylome, and these changes may persist as the child ages. As demonstrated previously, a low glycemic index dietary intervention was associated with changes in neonatal methylation patterns at birth [
15]. A low glycemic index diet in pregnancy may provide a more stable glycemic environment for the fetus, which could result in alterations in the fetal methylome that persist during childhood. With maternal obesity in pregnancy and conditions like gestational diabetes being controlled using dietary measures, it is important to investigate the impact of dietary intake on offspring and their health later in life. Offspring exposed to maternal diabetes in utero display accelerated growth in body mass index (BMI), but only from age two onwards [
16]. This demonstrates the importance of longitudinal follow-up studies, particularly with rates of obesity in young children estimated at 8–13% in developed countries like Ireland, UK, and Australia [
17]. While our low glycemic index dietary intervention during pregnancy appeared to influence offspring methylation patterns at birth, research is limited as to whether these changes persist, or whether the child’s current environment becomes a more influential factor as they age.
Candidate gene studies have identified areas of the methylome that appear sensitive to modulation and were associated with childhood body composition. Research has shown that methylation patterns in candidate genes measured in cord blood were associated with body size in children at nine years of age [
18,
19]. These studies, however, did not investigate whether these epigenetic marks were maintained in the methylome of the children at this age, and genome-wide research is lacking, particularly in early childhood. Research carried out in Finland identified differences in salivary DNA methylation associated with body mass index (BMI) in 11-year-old females [
20]. They identified over 100 specific CpG sites (regions of DNA where a cytosine nucleotide is followed by a guanine nucleotide), with differential methylation between participants on the 10th and 90th centile for BMI. Genome-wide studies carried out in adults from multiple cohorts identified associations between DNA methylation and increased BMI [
21]. In order to gain a better understanding of the pathways involved, linking epigenetic patterns with child body composition and adiposity, similar research is required in young children.
We hypothesised that the maternal environment in pregnancy, including a low glycemic index dietary intervention, would have a lasting impact on DNA methylation patterns in five-year-old children. We also hypothesised that child growth velocity and body composition is associated with variation in DNA methylation at five years of age.
4. Discussion
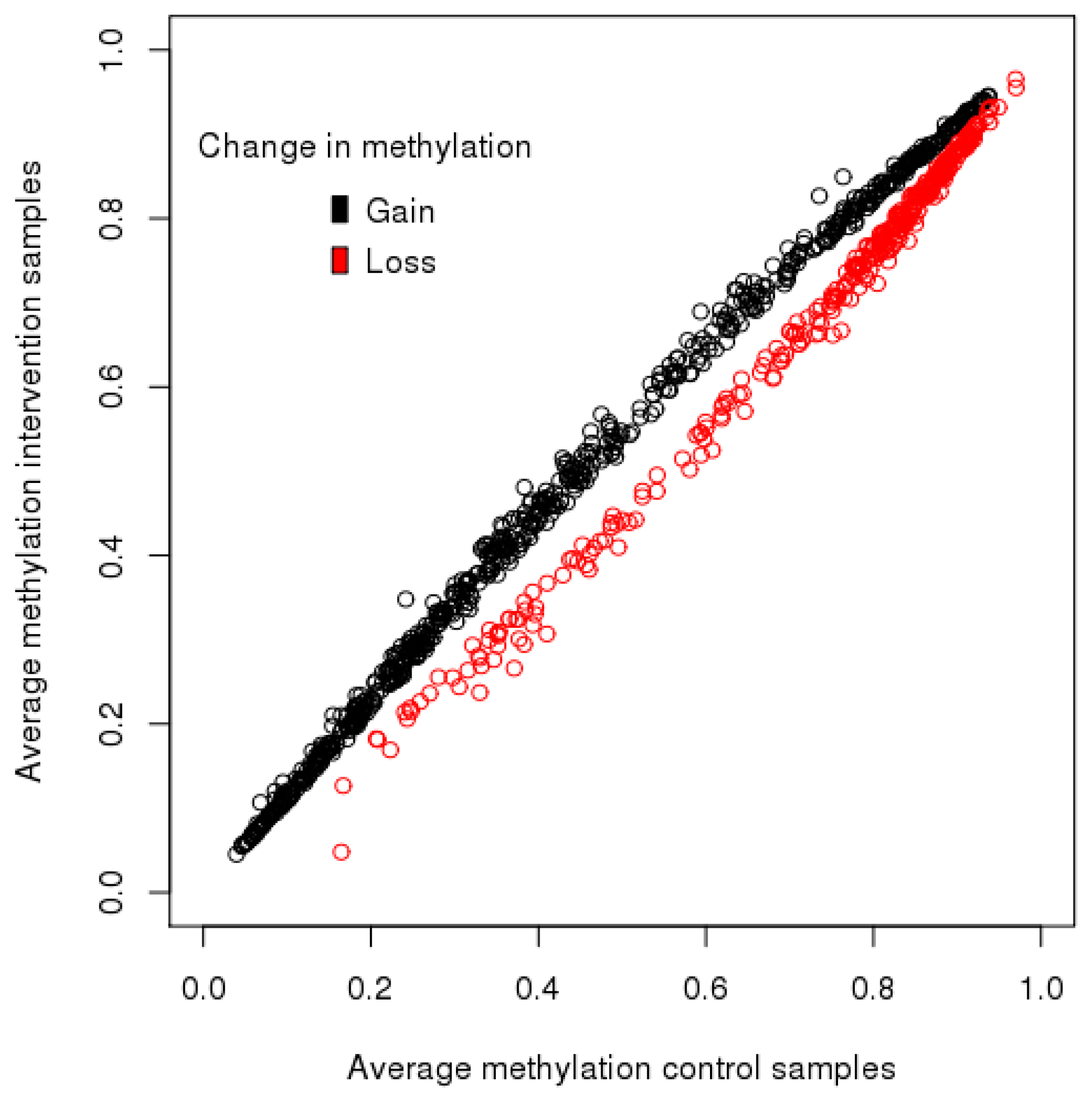
We found no association of maternal factors during pregnancy with offspring DNA methylation status at five years at age. Child weight at birth and gestational age were not associated with methylation status, and limited evidence for a lasting impact of the intervention was identified at this age. Body composition at five years was not associated with methylation status, however, change in weight centile from birth to five years of age was associated with variation in the sample set, which suggests that growth patterns may be linked with the methylome in childhood.
The lack of associations between maternal factors, particularly maternal weight or BMI, and child methylation patterns at five years of age suggests that these factors may not have a lasting impact on the methylome. Previous research has found some differences in methylation patterns in offspring at birth based on maternal BMI during pregnancy [
33], and these changes may persist as the child ages, impacting later body composition. However, more recent research carried out an erudite investigation into maternal adiposity during pregnancy and concluded that maternal BMI in pregnancy and subsequent offspring adiposity is explained by genetic transmission of BMI-associated gene variants [
34], as opposed to pervasive changes in DNA methylation. Longitudinal analysis of DNA methylation associated with birth weight and gestational age was also carried out from birth to adolescence, in which a lack of persistence of DNA methylation differences beyond early childhood was observed [
35].
Despite the low glycemic index diet being associated with variation in DNA methylation in newborns in this cohort [
15], there was no association of the intervention group with DNA methylation profiles at five years of age. Interestingly, however, maternal glycemic index in trimester 1 of pregnancy was significantly associated with some variation in the child DNA methylation profile. Studies have previously shown a lasting impact of a low glycemic index diet in offspring, with associations identified between dietary glycemic index in pregnancy and markers of metabolic syndrome in the offspring at 20 years of age [
36]. It may be that more persistent changes in DNA methylation are triggered when the fetus is exposed to the stimuli in the earliest stage of development at the beginning of pregnancy. The Dutch Hunger Winter study has shown that the timing of the exposure is critical, with evidence suggesting that early gestation (before 10 weeks) is uniquely sensitive to pervasive changes in offspring DNA methylation [
12]. So while dietary glycemic index in trimester 1 was associated with a small amount of variation in DNA methylation patterns of the offspring at five years of age, our low glycemic index intervention took place at approximately the thirteenth week of gestation, which may have been too late in fetal development to have a long-lasting impact on the methylome in later life. The Dutch Hunger Winter study concluded that early gestation, but not mid or late gestation, was critical for detectable changes in adult DNA methylation [
12]. Although associations with the dietary intervention weren’t statistically significant, exploratory pathway analysis into the top 1000 differentially methylated CpG sites between the control and intervention groups revealed some intriguing findings. The top cluster was related to insulin functioning, a pathway plausibly linked to the low glycemic index intervention. This could potentially explain the mechanism behind previous research, where a low glycemic index diet in pregnancy was reported to influence insulin functioning in the adult offspring [
36]. AMPK signaling, the third pathway identified, has also been shown to have a role in metabolism pathways influencing glucose uptake [
37]. Additionally, an Australian pilot randomized control trial observed that a low glycemic index diet during pregnancy influenced offspring arterial wall thickness in early childhood [
38]. The second pathway we identified related to the intervention was Notch signaling, which has been shown to play a role in cardiovascular health, particularly in pediatric cardiac disease [
39]. These findings may aid our understanding of the long-term impacts of a low glycemic index dietary intervention during pregnancy.
There were no associations of child weight or adiposity with methylation status at five years of age in this cohort. While previous studies have found associations with methylation and child adiposity, these tend to be gene-specific, targeted studies with methylation status at a younger age predicting body composition in later life [
18,
19,
40]. Our results suggest that change in weight and growth patterns rather than body composition at one time point in offspring may be guided by DNA methylation, particularly in early life, with our findings of weight centiles and BMI at six months of age being associated with some variation in methylation status in a subset of the sample group. There is a very powerful link between DNA methylation and aging, suggesting a plausible mechanistic role for variable DNA methylation patterns in controlling body composition trajectories as the child ages. This is consistent with the DOHaD hypothesis, which proposes that prenatal exposures can alter developmental trajectories [
41]. The DOHaD hypothesis highlights the link between the peri-conceptual period, fetal development, and early infancy with the subsequent development of metabolic disorders in later life [
42]. With DNA methylation patterns at birth being associated with child adiposity in later life [
18,
43], research is limited as to whether these patterns persist and are measurable in later life. These patterns of methylation may resolve or change as the child ages, and further research is required to determine this.
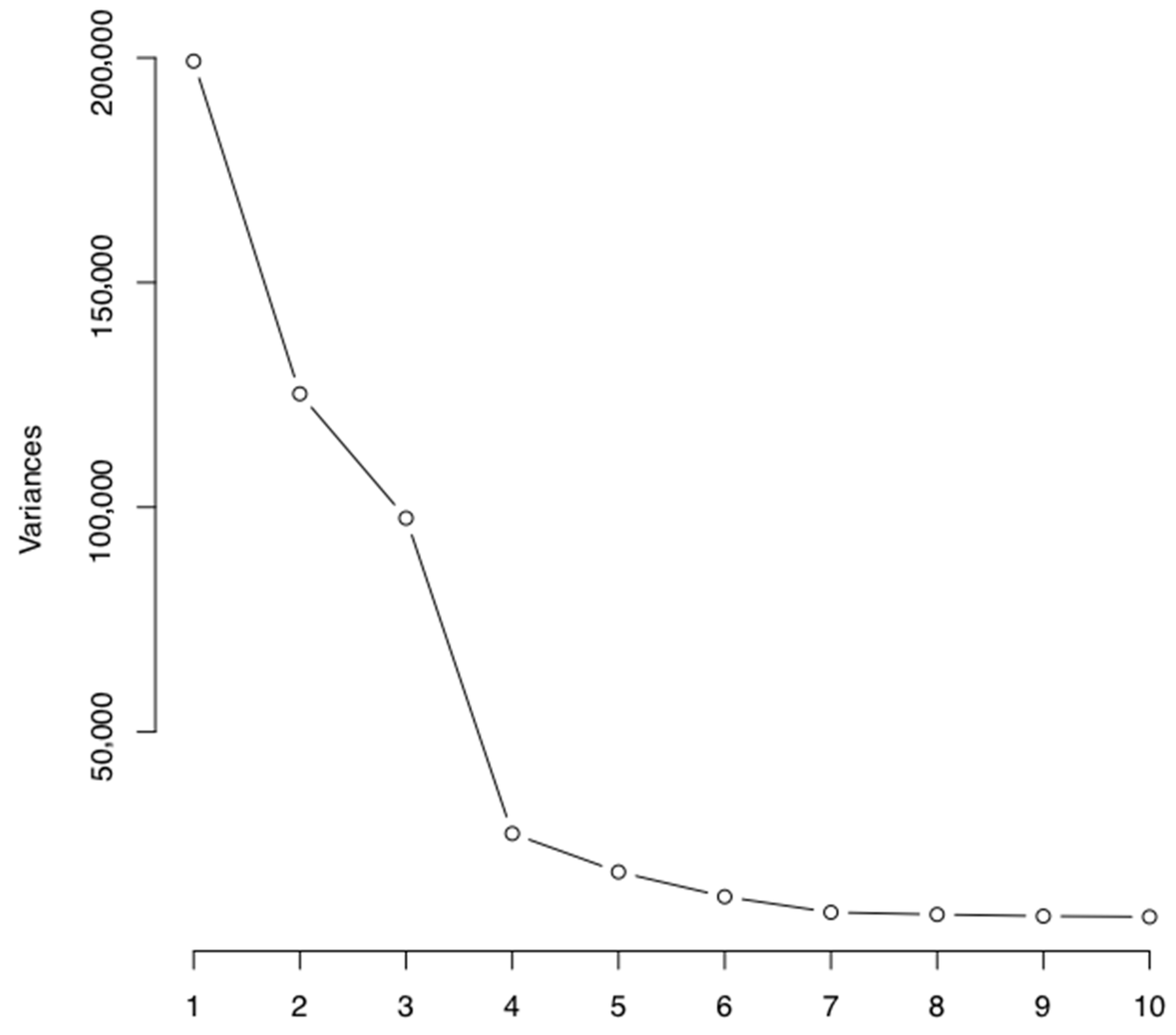
This study has many strengths, including the utilization of an extensive DNA methylation array, the Illumina Infinum MethylationEPIC BeadChip Array, which allowed the investigation of 780,501 CpG sites in the salivary DNA. We also had extensive comprehensive information on the participants, with each of the body measurements being collected by trained research staff, as opposed to relying on self-reported measures. We had detailed information on the participants at numerous time points, both phenotypic data along with dietary data. This allowed us to investigate the timings of associations and how this may change. This analysis was not without limitations, however. While sufficient for exploratory analysis, with the current sample size, statistical power may be limited to detect more subtle alterations in the methylome, particularly in relation to the biological impacts of the dietary intervention five years postpartum. The DNA methylation profiles were derived from saliva samples, and methylation patterns can vary based on tissue type. Other DNA sources, such as adipose tissue or blood, may be more strongly related to child body composition. However, research on adolescent females has previously shown an association between salivary DNA methylation and body size [
20]. There also are various other epigenetic mechanisms, like histone modification, which may have a role here or may be more responsive to environmental stimuli. In addition, more sensitive measures of body composition, such as air displacement plethysmography or dual-energy X-ray absorptiometry, may be more suitable methods to detect subtle differences. This represents further avenues of investigation.
These findings may serve to elucidate the factors that influence long-lasting DNA methylation patterns and the mechanisms behind which environment factors impact child body composition. With current research recognizing the vital role that epigenetics has in programming for later health [
42], this research expands on this area with longitudinal findings related to children. We found that in this cohort, factors in pregnancy did not have a large or lasting impact on overall variation in DNA methylation status of offspring at five years of age. We also found that methylation patterns were not associated with current child body composition. These findings may be important in guiding future research in this area. Extensive longitudinal research is required, along with suitably powered randomized controlled trials, to investigate potentially time-sensitive long-term impacts of environmental factors during pregnancy on child DNA methylation patterns and those in later life.

 ,
, 

{kind=link}
{kind=link}