Anti-Atherosclerotic Effect of a Polyphenol-Rich Ingredient, Oleactiv®, in a Hypercholesterolemia-Induced Golden Syrian Hamster Model

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Supplement
2.2. Design of the Study
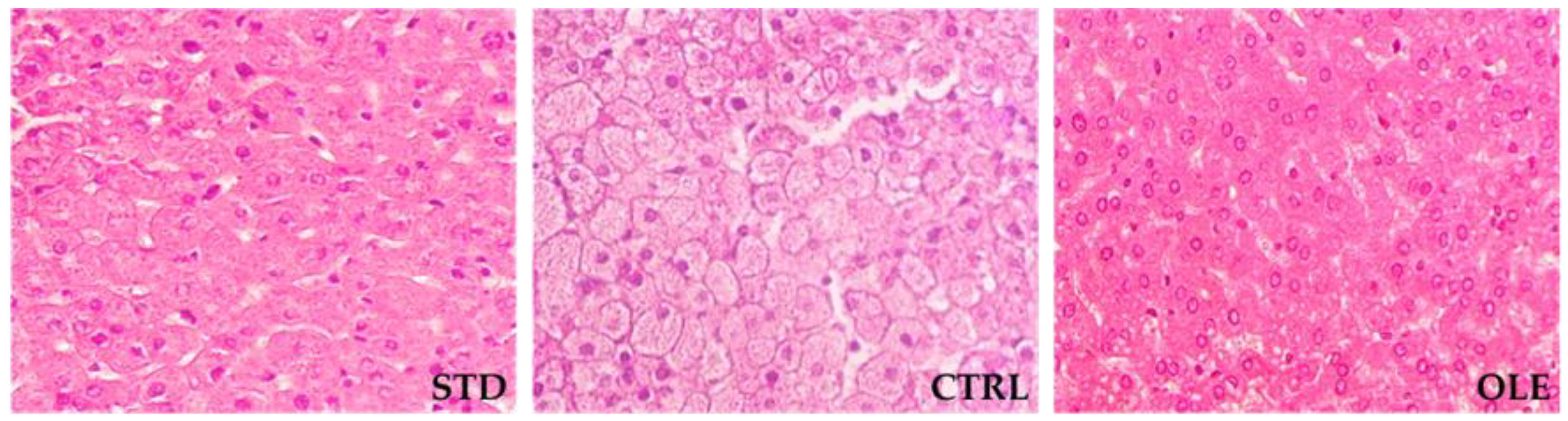
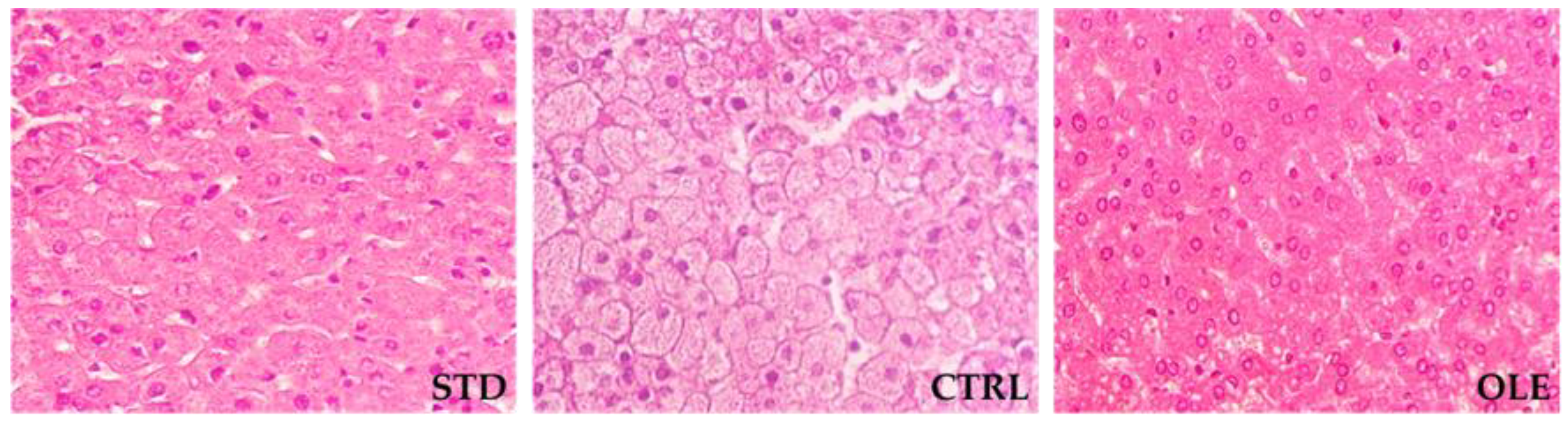
2.3. Evaluation of Hepatic Steatosis
2.4. Measurement of Lipid Profile
2.5. Evaluation of Early Atherosclerosis: Aortic Fatty Streak Area (AFSA)
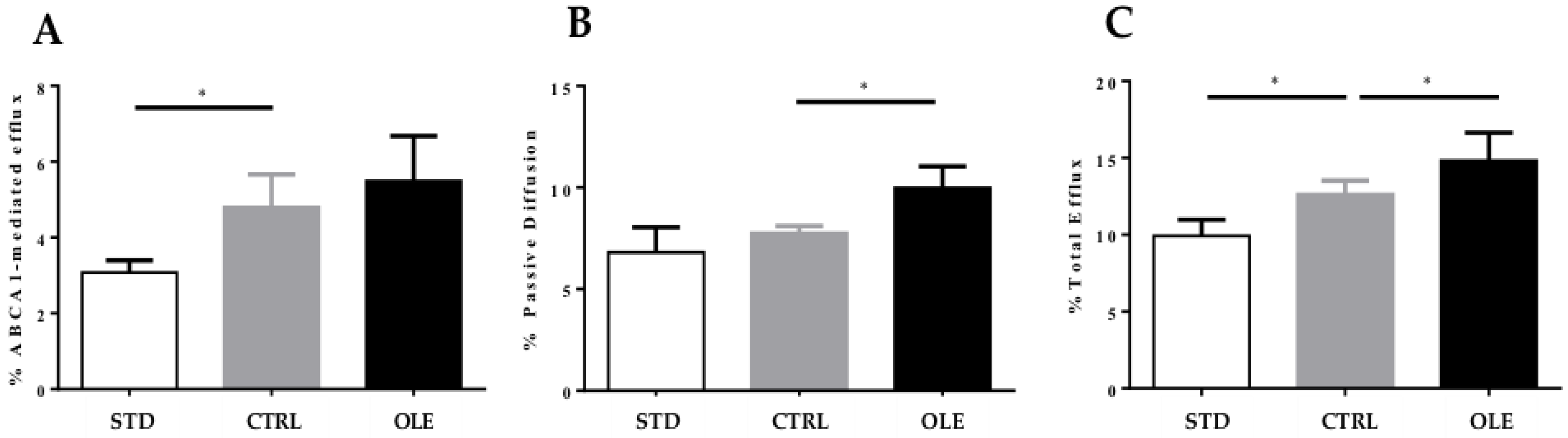
2.6. Evaluation of Cholesterol Efflux Capacity (CEC) of Sera
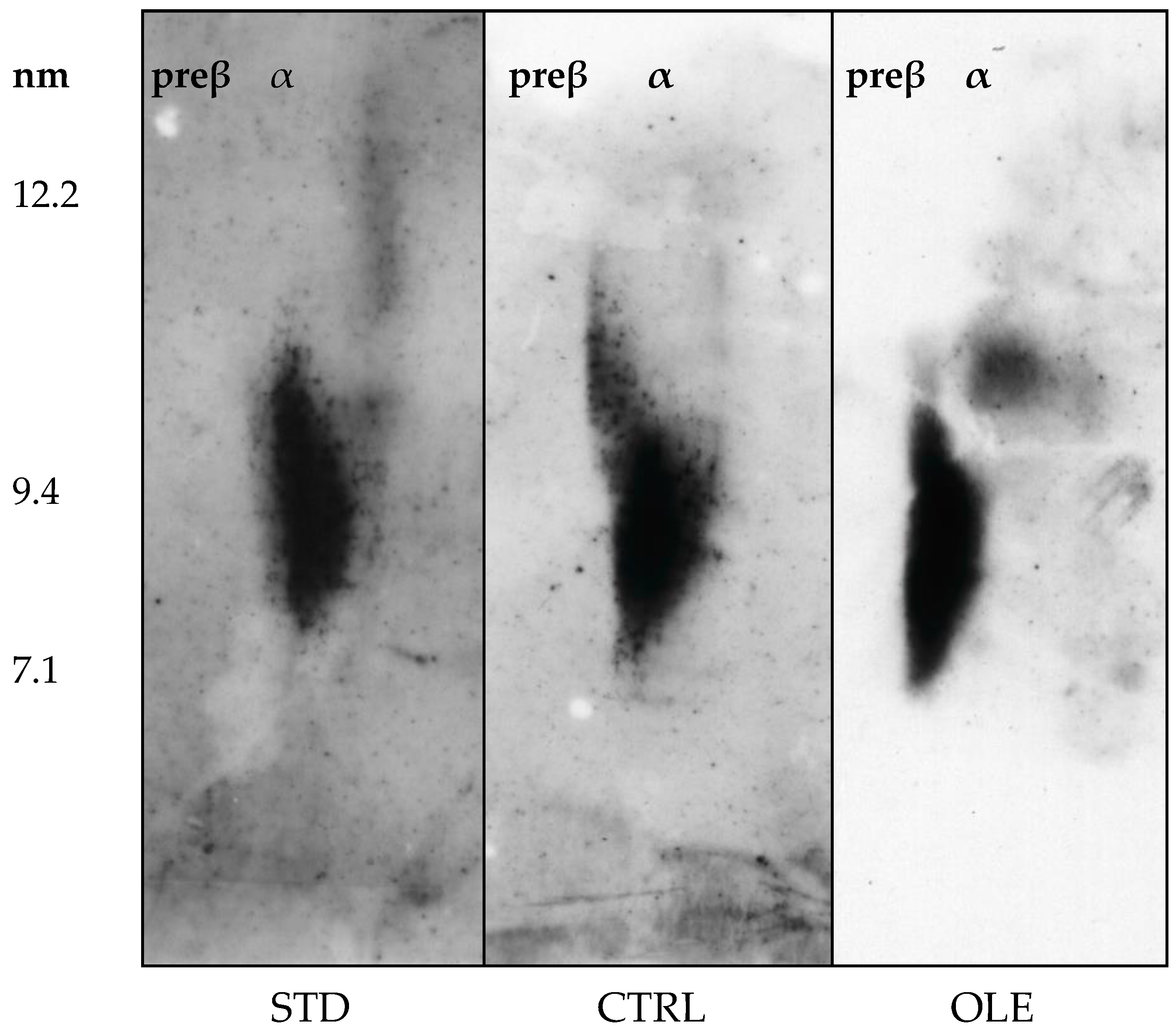
2.7. Characterization of HDL Particles
2.8. Statistical Analysis
3. Results
3.1. Characterization of the Polyphenolic Profile of the Supplement
3.2. Biometric Parameters and Food Intake
3.3. Hepatic Steatosis
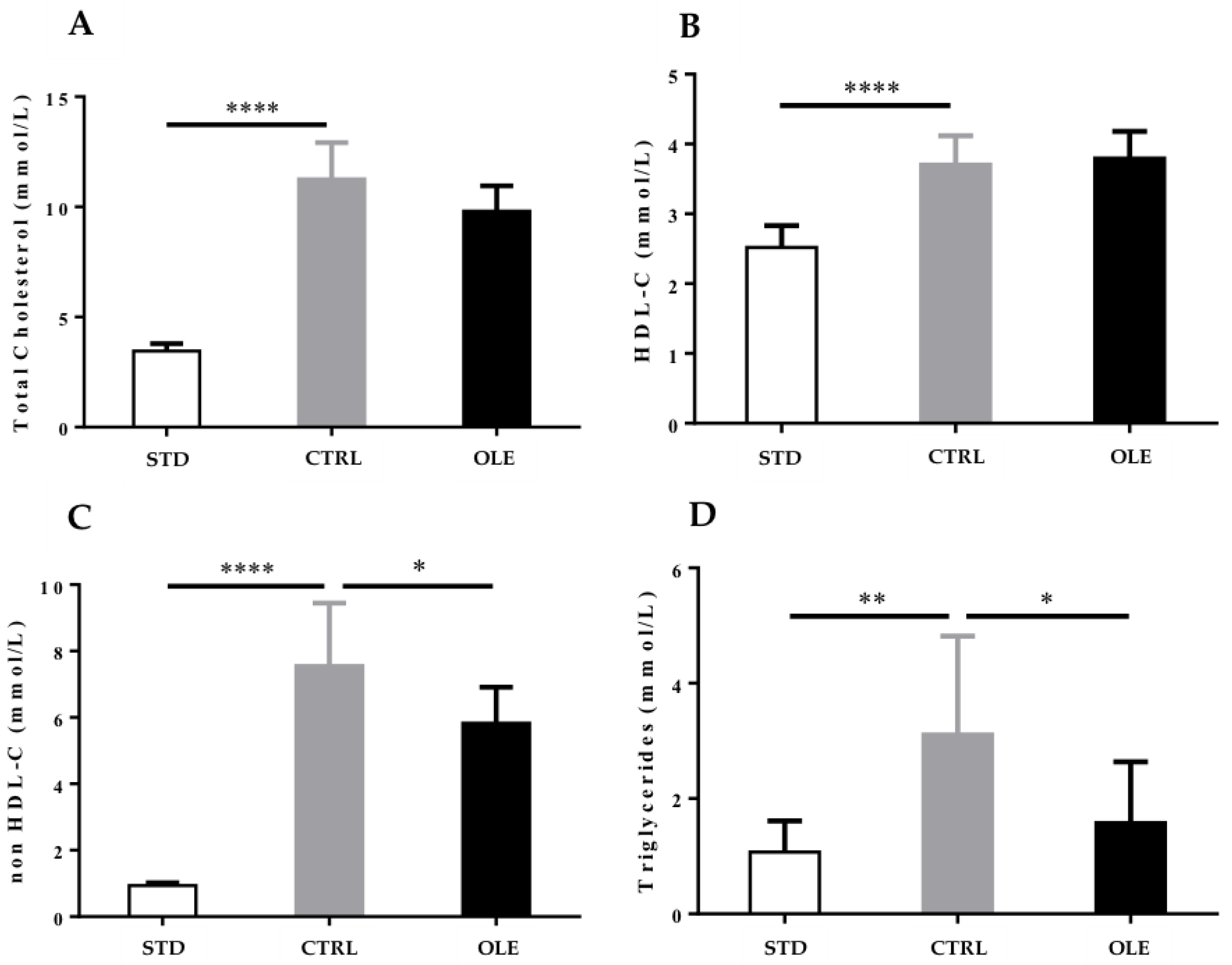
3.4. Plasma Lipid Profile
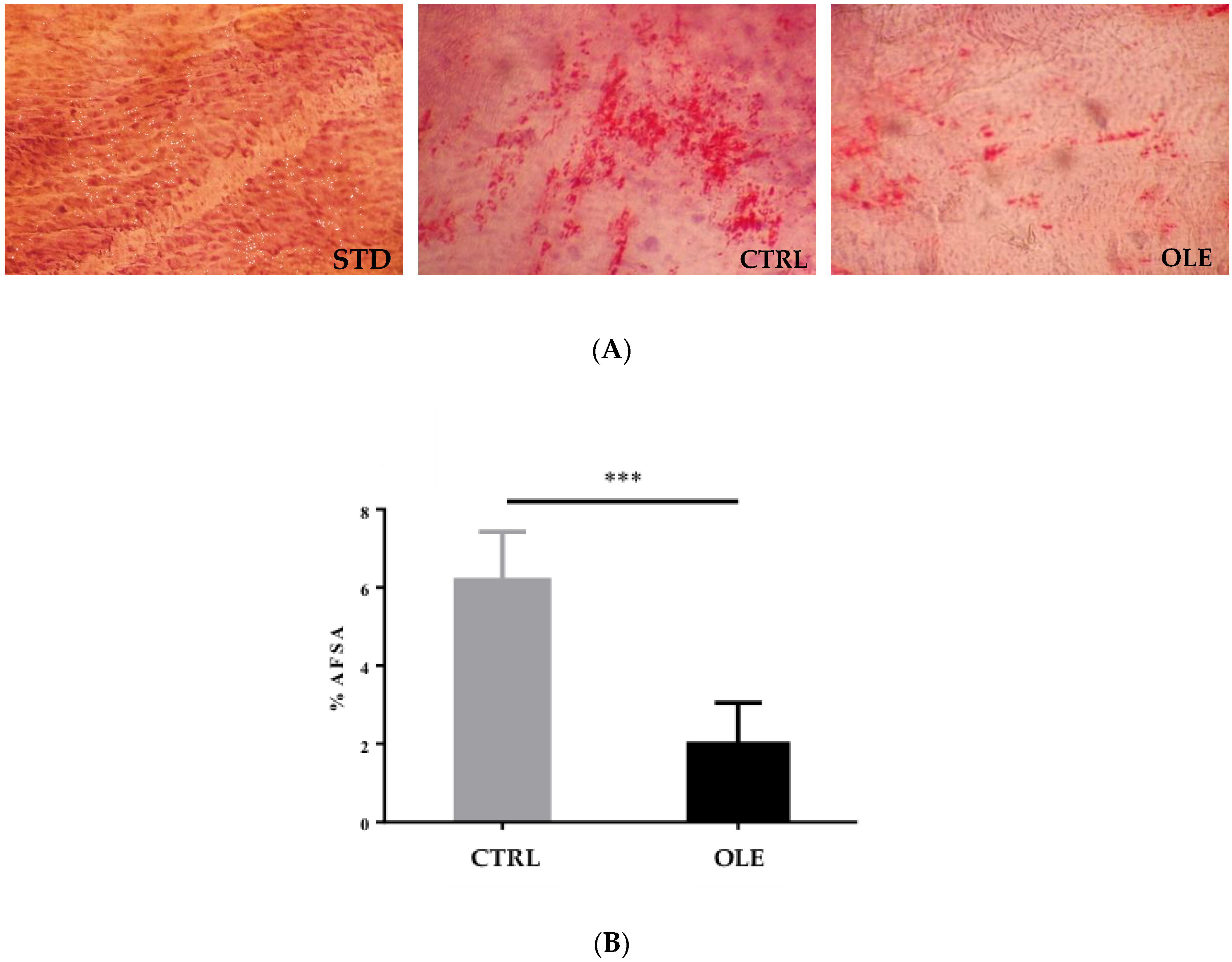
3.5. Aortic Fatty Streak Area (AFSA)
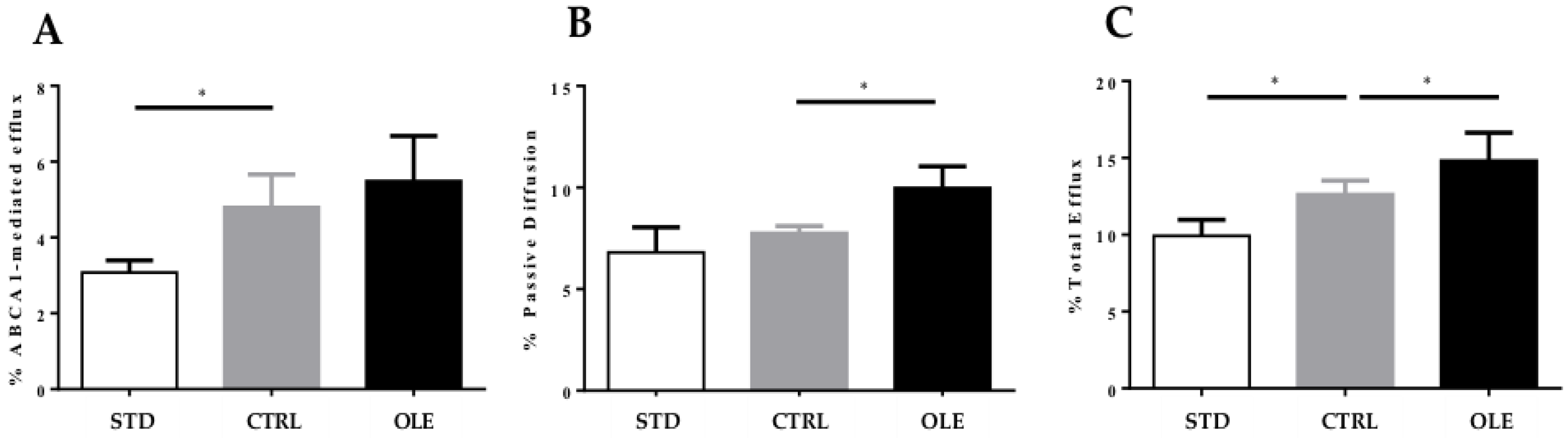
3.6. Cholesterol Efflux Capacity of Hamster Plasma
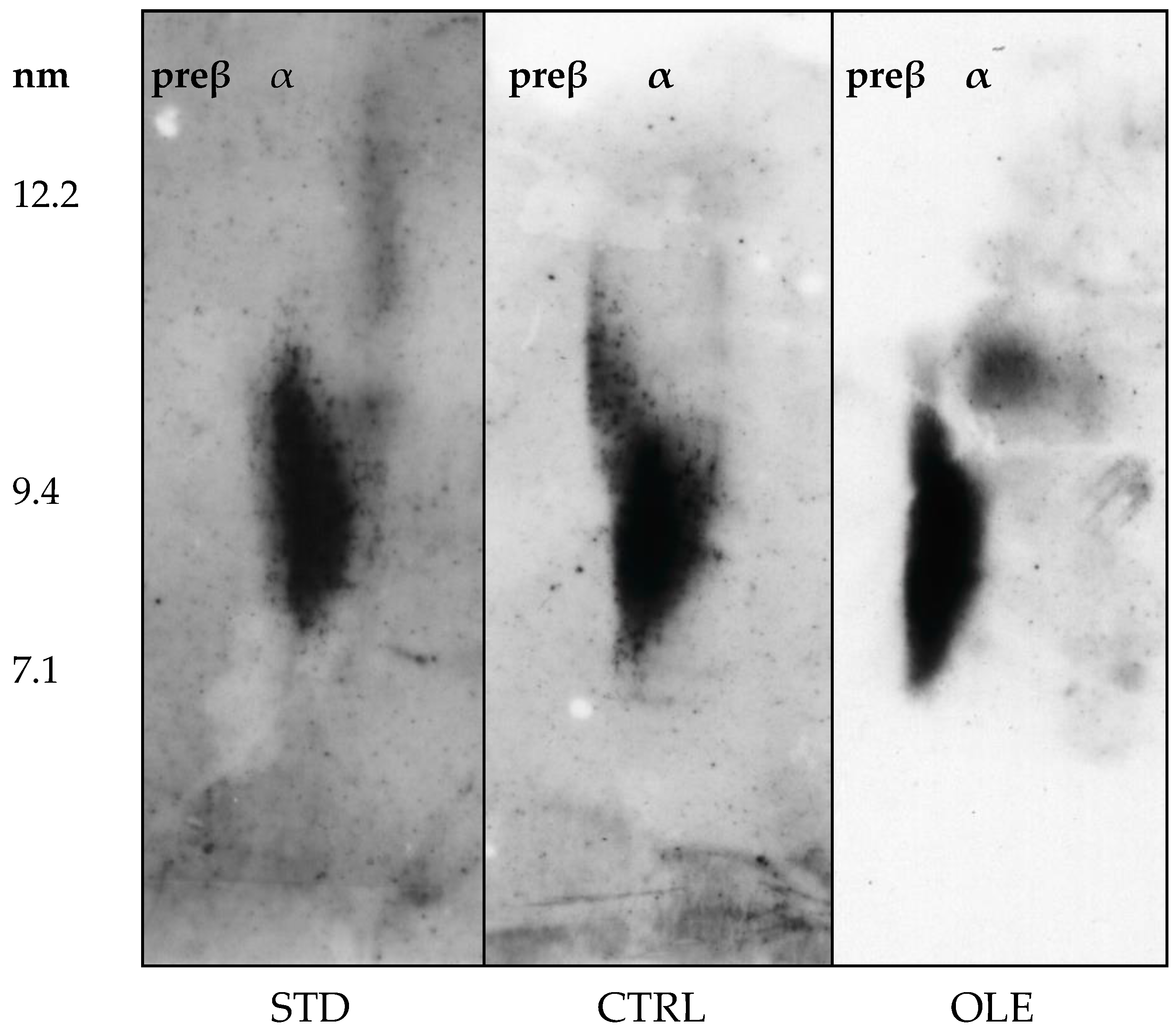
3.7. HDL Particle Characterization
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization (WHO). Available online: www.who.int/cardiovascular_diseases/en (accessed on 29 June 2018).[Green Version]
- Bentzon, J.F.; Otsuka, F.; Virmani, R.; Falk, E. Mechanisms of plaque formation and rupture. Circ. Res. 2014, 114, 1852–1866. [Google Scholar] [CrossRef] [PubMed]
- Khera, A.V.; Cuchel, M.; de la Llera-Moya, M.; Rodrigues, A.; Burke, M.F.; Jafri, K.; French, B.C.; Phillips, J.A.; Mucksavage, M.L.; Wilensky, R.L.; et al. Cholesterol efflux capacity, high-density lipoprotein function, and atherosclerosis. N. Engl. J. Med. 2011, 364, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Zhao, X.; Zhou, Q.; Zhang, Z. High-density lipoprotein cholesterol efflux capacity is inversely associated with cardiovascular risk: A systematic review and meta-analysis. Lipids Health Dis. 2017, 16, 212. [Google Scholar] [CrossRef] [PubMed]
- Santos-Gallego, C.G.; Giannarelli, C.; Badimón, J.J. Experimental models for the investigation of high-density lipoprotein-mediated cholesterol efflux. Curr. Atheroscler. Rep. 2011, 13, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, P.M.; Heinecke, J.W. Cholesterol efflux capacity, macrophage reverse cholesterol transport and cardioprotective HDL. Curr. Opin. Lipidol. 2015, 26, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Knoops, K.T.; de Groot, L.C.; Kromhout, D.; Perrin, A.E.; Moreiras-Valera, O.; Menotti, A.; van Staveren, W.A. Mediterranean diet, lifestyle factors, and 10-year mortality in elderly European men and women: The HALE project. JAMA 2004, 292, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary prevention of cardiovascular disease with a Mediterranean diet. N. Engl. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Tressera-Rimbau, A.; Rimm, E.B.; Medina-Remón, A.; Martínez-González, M.A.; López-Sabater, M.C.; Covas, M.I.; Corella, D.; Salas-Salvadó, J.; Gómez-Gracia, E.; Lapetra, J.; et al. Polyphenol intake and mortality risk: A re-analysis of the PREDIMED trial. BMC Med. 2014, 12, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanotti, I.; Dall’Asta, M.; Mena, P.; Mele, L.; Bruni, R.; Ray, S.; Del Rio, D. Atheroprotective effects of (poly)phenols: A focus on cell cholesterol metabolism. Food Funct. 2015, 6, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Patrick, L.; Uzick, M. Cardiovascular disease: C-reactive protein and the inflammatory disease paradigm: HMG-CoA reductase inhibitors, alpha-tocopherol, red yeast rice, and olive oil polyphenols. A review of the literature. Altern. Med. Rev. 2001, 6, 248–271. [Google Scholar] [PubMed]
- Millar, C.L.; Duclos, Q.; Blesso, C.N. Effects of dietary flavonoids on reverse cholesterol transport, HDL metabolism, and HDL function. Adv. Nutr. 2017, 8, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Weingand, K.W.; Daggy, B.P. Quantification of high-density-lipoprotein cholesterol in plasma from hamsters by differential precipitation. Clin. Chem. 1990, 36, 575. [Google Scholar] [PubMed]
- Auger, C.; Teissedre, P.L.; Gérain, P.; Lequeux, N.; Bornet, A.; Serisier, S.; Besançon, P.; Caporiccio, B.; Cristol, J.P.; Rouanet, J.M. Dietary wine phenolics catechin, quercetin, and resveratrol efficiently protect hypercholesterolemic hamsters against aortic fatty streak accumulation. J. Agric. Food Chem. 2005, 53, 2015–2021. [Google Scholar] [CrossRef] [PubMed]
- Bortnick, A.E.; Rothblat, G.H.; Stoudt, G.; Hoppe, K.L.; Royer, L.J.; McNeish, J.; Francone, O.L. The correlation of ATP-binding cassette 1 mRNA levels with cholesterol efflux from various cell lines. J. Biol. Chem. 2000, 275, 28634–28640. [Google Scholar] [CrossRef] [PubMed]
- Zanotti, I.; Favari, E.; Bernini, F. Cellular cholesterol efflux pathways: Impact on intracellular lipid trafficking and methodological considerations. Curr. Pharm. Biotechnol. 2012, 13, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Pedrelli, M.; Davoodpour, P.; Degirolamo, C.; Gomaraschi, M.; Graham, M.; Ossoli, A.; Larsson, L.; Calabresi, L.; Gustafsson, J.Å.; Steffensen, K.R.; et al. Hepatic ACAT2 knock down increases ABCA1 and modifies HDL metabolism in mice. PLoS ONE. 2014, 9, e93552. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.B.; Mengi, S.A.; Xu, Y.J.; Arneja, A.S.; Dhalla, N.S. Pathogenesis of atherosclerosis: A multifactorial process. Exp. Clin. Cardiol. 2002, 7, 40–53. [Google Scholar] [PubMed]
- Lusis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Rohatgi, A.; Khera, A.; Berry, J.D.; Givens, E.G.; Ayers, C.R.; Wedin, K.E.; Neeland, I.J.; Yuhanna, I.S.; Rader, D.R.; de Lemos, J.A.; et al. HDL cholesterol efflux capacity and incident cardiovascular events. N. Engl. J. Med. 2014, 371, 2383–2393. [Google Scholar] [CrossRef] [PubMed]
- Saleheen, D.; Scott, R.; Javad, S.; Zhao, W.; Rodrigues, A.; Picataggi, A.; Lukmanova, D.; Mucksavage, M.L.; Luben, R.; Billheimer, J.; et al. Association of HDL cholesterol efflux capacity with incident coronary heart disease events: A prospective case-control study. Lancet Diabetes Endocrinol. 2015, 3, 507–513. [Google Scholar] [CrossRef]
- Ishikawa, T.; Ayaori, M.; Uto-Kondo, H.; Nakajima, T.; Mutoh, M.; Ikewaki, K. High-density lipoprotein cholesterol efllux capacity as a relevant predictor of atherosclerotic coronary disease. Atheroscleosis 2015, 242, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Du, X.M.; Kim, M.J.; Hou, L.; Le Goff, W.; Chapman, M.J.; Van Eck, M.; Curtiss, L.K.; Burnett, J.R.; Cartland, S.P.; Quinn, C.M.; et al. HDL particle size is a critical determinant of ABCA1-mediated macrophage cellular cholesterol export. Circ. Res. 2015, 116, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Favari, E.; Chroni, A.; Tietge, U.J.; Zanotti, I.; Escolà-Gil, J.C.; Bernini, F. Cholesterol efllux and reverse cholesterol transport. Handb. Exp. Pharmacol. 2015, 224, 181–206. [Google Scholar] [PubMed]
- Berrougui, H.; Ikhlef, S.; Khalil, A. Extra virgin olive oil polyphenols promote cholesterol efflux and improve HDL functionality. Evid. Based Complement. Altern. Med. 2015, 2015, 208062. [Google Scholar] [CrossRef] [PubMed]
- Rifici, V.A.; Schneider, S.H.; Khachadurian, A.K. Lipoprotein oxidation mediated by J774 muirine macrophages is inhibited by individual red wine polyphenols but not by ethanol. J. Nutr. 2002, 132, 2532–2537. [Google Scholar] [CrossRef] [PubMed]
- Herrera, L.V.; Lou-Bonafonte, J.M.; Arnal, C.; Navarro, M.A.; Osada, J. Transcriptomics and the Mediterranean Diet: A systematic review. Nutrients 2017, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- Lou-Bonafonte, J.M.; Gabas-Rivera, C.; Navarro, M.A.; Osada, J. PON1 and Mediterranean Diet. Nutrients 2015, 7, 4062–4092. [Google Scholar] [CrossRef] [PubMed]
- Acin, S.; Navarro, M.A.; Carnicer, R.; Arbones-Mainar, J.M.; Guzman, M.A.; Arnal, C.; Beltran, G.; Uceda, M.; Maeda, N.; Osada, J. Dietary cholesterol suppresses the ability of olive oil to delay the development of atherosclerotic lesions in apolipoprotein E knockout mice. Atherosclerosis 2005, 182, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Moreno, D.A.; Illic, N.; Poulev, A.; Brasaemle, D.L.; Fried, S.K.; Raskin, I. Inhibitory effects of grape seed extract on lipases. Nutrition 2003, 19, 876–879. [Google Scholar] [CrossRef]
- Meng, S.; Cao, J.; Feng, Q.; Peng, J.; Hu, Y. Roles of chlorogenic acid on regulating glucose and lipids metabolism: A review. Evid. Based Complement. Altern. Med. 2013, 2013, 801457. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Jung, U.J.; Lee, M.K.; Cho, S.J.; Jung, H.K.; Hong, J.H.; Park, Y.B.; Kim, S.R.; Shim, S.; Jung, J.; et al. Modulation of lipid metabolism by polyphenol-rich grape skin extract improves liver steatosis and adiposity in high fat fed mice. Mol. Nutr. Food Res. 2013, 57, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, R. Inhibition of cholesterol biosynthesis in HepG2 cells by artichoke extracts is reinforced by glucosidase pretreatment. Phytother. Res. 2002, 16, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z.; Mirmiran, P.; Azizi, F. Dietary polyphenols as potential nutraceuticals in management of diabetes: A review. J. Diabetes Metab. Disord. 2013, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Dawson, H.D.; Schoene, N.W.; Polansky, M.M.; Anderson, R.A. Cinnamon polyphenols regulate multiple metabolic pathways involved in insulin signaling and intestinal lipoprotein metabolism of small intestinal enterocytes. Nutrition 2012, 28, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Nekohashi, M.; Ogawa, M.; Ogihara, T.; Nakazawa, K.; Kato, H.; Misaka, T.; Abe, K.; Kobayashi, S. Luteolin and quercetin affect the cholesterol absorption mediated by epithelial cholesterol transporter niemann-pick c1-like 1 in caco-2 cells and rats. PLoS ONE 2014, 9, e97901. [Google Scholar] [CrossRef] [PubMed]
- Pons, Z.; Margalef, M.; Bravo, F.I.; Arola-Arnal, A.; Muguerza, B. Chronic administration of grape-seed polyphenols attenuates the development of hypertension and improves other cardiometabolic risk factors associated with the metabolic syndrome in cafeteria diet-fed rats. Br. J. Nutr. 2017, 117, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Lv, O.; Wang, L.; Li, J.; Ma, Q.; Zhao, W. Effects of pomegranate peel polyphenols on lipid accumulation and cholesterol metabolic transformation in L-02 human hepatic cells via the PPARγ-ABCA1/CYP7A1 pathway. Food Funct. 2016, 7, 4976–4983. [Google Scholar] [CrossRef] [PubMed]
- Ben Salem, M.; Affes, H.; Ksouda, K.; Dhouibi, R.; Sahnoun, Z.; Hammami, S.; Zeghal, K.M. Pharmacological studies of artichoke leaf extract and their health benefits. Plant Foods Hum. Nutr. 2015, 70, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Auger, C.; Caporiccio, B.; Landrault, N.; Teissedre, P.L.; Laurent, C.; Cros, G.; Besançon, P.; Rouanet, J.M. Red wine phenolic compounds reduce plasma lipids and apolipoprotein B and prevent early aortic atherosclerosis in hypercholesterolemic golden Syrian hamsters (Mesocricetus auratus). J. Nutr. 2002, 132, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Y.; Zhang, H.; Ren, L.; Li, Q.; Li, N. Animal models for the atherosclerosis research: A review. Protein Cell 2011, 2, 189–201. [Google Scholar]
- Dillard, A.; Matthan, N.R.; Lichtenstein, A.H. Use of hamster as a model to study diet-induced atherosclerosis. Nutr. Metab. 2010, 7, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Standard Diet | High-Fat Diet | |||
|---|---|---|---|---|
| Ingredients | % | kcal/kg Diet | % | kcal/kg Diet |
| Casein | 23.60 | 944 | 20.00 | 800 |
| L-Methionin | 0.35 | 14 | 3.0 | 12 |
| Corn starch | 30.00 | 1200 | 39.30 | 1572 |
| Maltodextrin | 3.0 | 120 | 5.3 | 212 |
| Sucrose | 29.05 | 1162 | 15.40 | 616 |
| Cellulose | 5.0 | 0 | 5.0 | 0 |
| Vegetables oil (corn/soy, 1:1) | 4.5 | 405 | 0.0 | 0 |
| Hydrogenated coconut oil | 0.0 | 0 | 10.00 | 900 |
| Cholesterol | 0.0 | 0 | 0.2 | 18 |
| Minerals mix | 3.5 | 0 | 3.5 | 0 |
| Vitamins mix | 1.0 | 40 | 1.0 | 40 |
| Total | 100 | 3885 | 100 | 4170 |
| Compounds | Rt (min) | Λ Max (nm) | Content (g/100 g) |
|---|---|---|---|
| Gallic acid | 5.4 | 272 | 0.14 ± 0.11 |
| Hydroxytyrosol | 12.3 | 280 | 2.93 ± 0.35 |
| Procyanidin B1 | 16.6 | 278 | 0.59 ± 0.09 |
| Procyanidin-like | 17.3 | 280 | 0.38 ± 0.06 |
| Catechin | 19.0 | 280 | 1.39 ± 0.39 |
| Chlorogenic acid | 24.4 | 300; 326 | 0.80 ± 0.07 |
| Cryptochlorogenic acid | 27.5 | 300; 326 | 0.54 ± 0.04 |
| Epicatechin | 33.9 | 278 | 1.01 ± 0.41 |
| Flavanol-like | 36.9 | 278 | 0.73 ± 0.13 |
| Cynarin | 43.5 | 302; 318 | 0.45 ± 0.07 |
| Cynarosid | 87.6 | 348 | 0.99 ± 0.19 |
| Oleuropein | 102.4 | 281 | 5.02 ± 0.18 |
| Oleuropein-like | 107.4 | 281 | 1.13 ± 0.06 |
| STD Group | CTRL Group | OLE Group | |
|---|---|---|---|
| Body Weight (g) | 99.95 ± 15.11 | 128.73 ± 7.09 **** | 125.01 ± 5.40 (ns) |
| Liver Weight (g) | 3.74 ± 0.67 | 7.34 ± 0.85 **** | 7.22 ± 0.76 (ns) |
| % Liver/Weight | 3.74 ± 0.32 | 5.70 ± 0.57 **** | 5.77 ± 0.54 (ns) |
| Food intake (g/day) | 4.57 ± 0.59 | 6.04 ± 1.07 **** | 6.14 ± 1.26 (ns) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romain, C.; Piemontese, A.; Battista, S.; Bernini, F.; Ossoli, A.; Strazzella, A.; Gaillet, S.; Rouanet, J.-M.; Cases, J.; Zanotti, I. Anti-Atherosclerotic Effect of a Polyphenol-Rich Ingredient, Oleactiv®, in a Hypercholesterolemia-Induced Golden Syrian Hamster Model. Nutrients 2018, 10, 1511. https://doi.org/10.3390/nu10101511
Romain C, Piemontese A, Battista S, Bernini F, Ossoli A, Strazzella A, Gaillet S, Rouanet J-M, Cases J, Zanotti I. Anti-Atherosclerotic Effect of a Polyphenol-Rich Ingredient, Oleactiv®, in a Hypercholesterolemia-Induced Golden Syrian Hamster Model. Nutrients. 2018; 10(10):1511. https://doi.org/10.3390/nu10101511
Chicago/Turabian StyleRomain, Cindy, Antonio Piemontese, Simone Battista, Franco Bernini, Alice Ossoli, Arianna Strazzella, Sylvie Gaillet, Jean-Max Rouanet, Julien Cases, and Ilaria Zanotti. 2018. "Anti-Atherosclerotic Effect of a Polyphenol-Rich Ingredient, Oleactiv®, in a Hypercholesterolemia-Induced Golden Syrian Hamster Model" Nutrients 10, no. 10: 1511. https://doi.org/10.3390/nu10101511