Nutrigenetic Contributions to Dyslipidemia: A Focus on Physiologically Relevant Pathways of Lipid and Lipoprotein Metabolism


1
Division of Nutritional Sciences, University of Illinois at Urbana-Champaign, Urbana-Champaign, IL 61801, USA
2
Department of Kinesiology and Community Health, University of Illinois at Urbana-Champaign, Urbana-Champaign, IL 61801, USA
3
Department of Human Development and Family Studies, Cooperative Extension, University of Illinois at Urbana-Champaign, Carle Illinois College of Medicine, Urbana-Champaign, IL 61801, USA
*
Author to whom correspondence should be addressed.
Nutrients 2018, 10(10), 1404; https://doi.org/10.3390/nu10101404
Submission received: 28 August 2018
/
Revised: 19 September 2018
/
Accepted: 27 September 2018
/
Published: 2 October 2018
(This article belongs to the Special Issue Diet, Lipid and Lipoprotein Metabolism and Human Health)
Abstract
:Cardiovascular disease (CVD) remains the number one cause of death worldwide, and dyslipidemia is a major predictor of CVD mortality. Elevated lipid concentrations are the result of multiple genetic and environmental factors. Over 150 genetic loci have been associated with blood lipid levels. However, not all variants are present in pathways relevant to the pathophysiology of dyslipidemia. The study of these physiologically relevant variants can provide mechanistic understanding of dyslipidemia and identify potential novel therapeutic targets. Additionally, dietary fatty acids have been evidenced to exert both positive and negative effects on lipid profiles. The metabolism of both dietary and endogenously synthesized lipids can be affected by individual genetic variation to produce elevated lipid concentrations. This review will explore the genetic, dietary, and nutrigenetic contributions to dyslipidemia.
1. Introduction
Elevated blood lipid concentrations, or dyslipidemia, currently affect 13% of the US population, and are strong predictors of cardiovascular disease (CVD) [1]. Dyslipidemia can be diagnosed by the presence of one or more of the following phenotypes: elevated concentrations of total cholesterol (TC), low-density lipoprotein cholesterol (LDL), and triglycerides (TG), or low concentrations of high-density lipoprotein cholesterol (HDL) [2]. Dyslipidemia is a complex disease that is the result of multiple biological and behavioral etiologies, such as genetic predisposition, metabolic capacity, dietary intake, and physical activity [3]. Understanding the interactions between these complex factors to produce phenotypes of dyslipidemia is crucial to identifying and implementing successful strategies to manage blood lipids. Among biological factors, the study of genetics is essential to improving scientific understanding of disease progression at its most basic level and understanding of the role of individual genetic variation in disease predisposition can lead to improvements in identification and prevention of disease in genetically at-risk individuals. Of the many behavioral contributors to dyslipidemia, diet offers one of the most efficacious behavioral approaches to disease prevention, and it is a crucial determinant of maintenance of health throughout the lifespan [4]. The role of dietary fat intake in exacerbation or amelioration of CVD risk has been a topic of debate in the field of nutrition. Dietary fatty acids are a heterogeneous group of nutrients, and their varying molecular properties, as well as the food matrix in which they reside, exert differential effects on blood lipids. The intake and metabolism of various fatty acids may be influenced by individual genetic variation. These biological and behavioral factors must be considered not as individual risk factors, but as interacting elements. The following review of the literature will present a summary of the recent work conducted to better elucidate the role of both biological (genetic) and behavioral (dietary) influences on dyslipidemia, and the interactions of these components in the clinical intervention setting.
2. Pathophysiology of Dyslipidemia
Elevated TG and decreased HDL concentrations are metabolic consequences of excess visceral adipose tissue and increase risk of atherosclerotic disease through various mechanisms. The role of elevated TG concentrations in CVD progression is not fully elucidated, but it has been postulated to be due to increased endothelial activation and inflammation [5]. Elevated TG concentrations are strongly associated with insulin resistance, CVD, and other indicators of metabolic dysfunction, due to excess adipose tissue mass [6,7]. A hypertriglyceridemic state promotes the exchange of TG from very-low density lipoprotein (VLDL) for cholesterol esters from LDL and HDL particles, creating small, lipid-poor particles. Small HDL particles are more susceptible to degradation, thus contributing to the low HDL concentrations observed in the presence of other dyslipidemias [8]. Elevated HDL concentrations are generally recognized as cardioprotective, as these lipoproteins serve to sequester excess cholesterol to the liver for excretion. Low concentrations of HDL are a diagnostic biomarker for both the Metabolic Syndrome (MetS) and CVD. The relationship between elevated HDL and metabolic disease has been challenged by results from clinical trials with HDL-raising agents, which did not lead to reduction in cardiovascular events compared to the control group [9]. However, due to the strong inverse relationship between HDL concentrations and CVD at the epidemiological level, it remains a key biomarker for assessing cardiometabolic health [10].
3. Genetic Contributions to Dyslipidemia
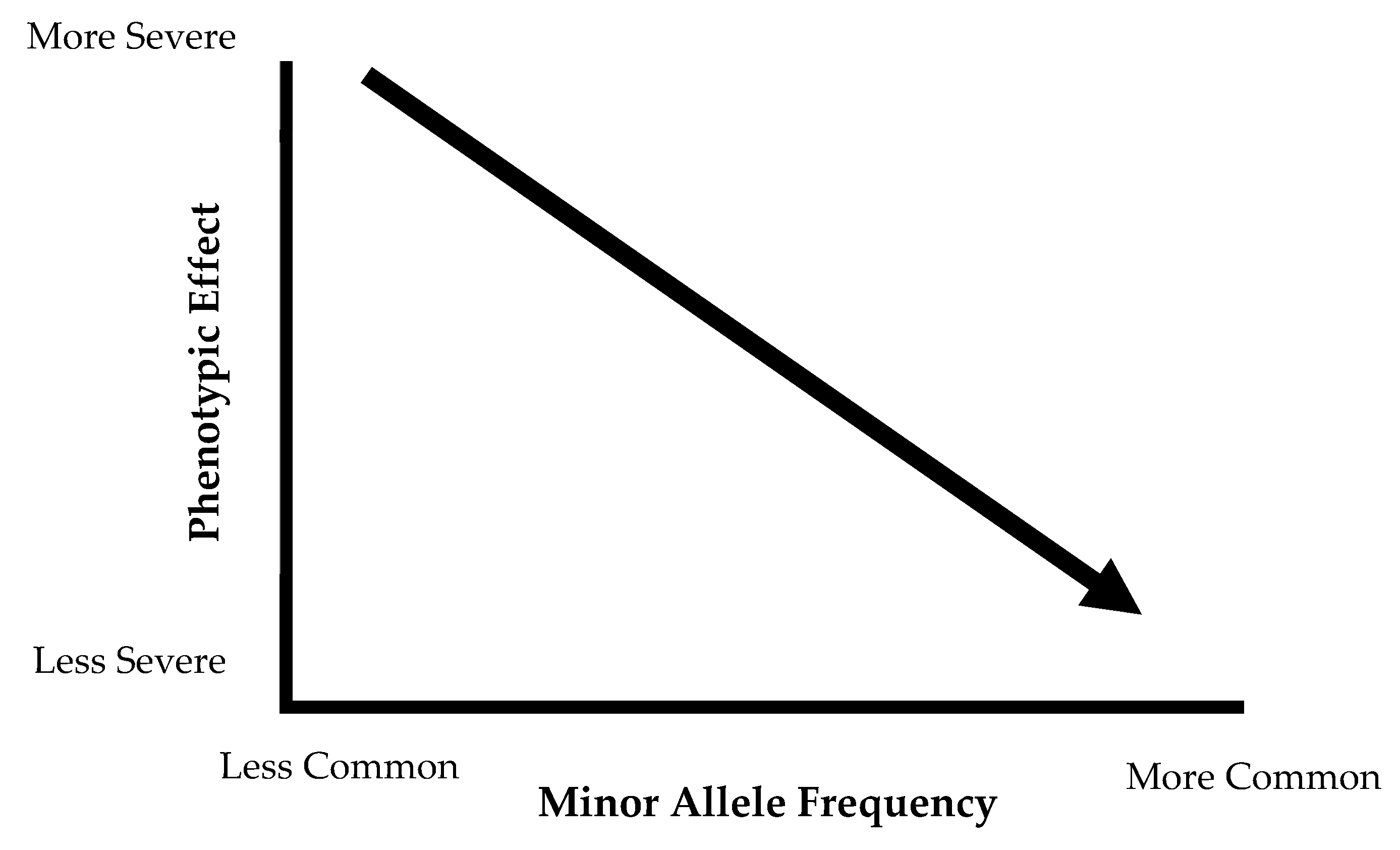
With the exception of rare genetic mutations, the majority of dyslipidemias are secondary to other metabolic abnormalities, including abdominal obesity [6]. When describing the complex genetic components of dyslipidemia, one can distinguish between monogenic and polygenic traits. Monogenic diseases are the result of a single mutation in one gene, resulting in a severe phenotype. Some classic examples of these monogenic conditions include Tangier disease (resulting in severely low HDL), LDL receptor deficiency (characterized by elevated LDL concentrations), familial chylomicronemia or lipoprotein lipase (LPL) deficiency (causes severe hypertriglyceridemia) and other familial hypercholesterolemias [11,12,13]. Table 1 presents summary of selected monogenic lipid disorders. These monogenic conditions produce a severe effect, but the frequency of these risk alleles in the population is considerably low. The common variants that produce smaller phenotypic effects contribute to the polygenic nature of obesity and dyslipidemia. These common variants, specifically the single nucleotide polymorphisms (SNPs), are present in at least 1% of the population, and the phenotypic effect of these SNPs individually is not likely to be observed. The differences in phenotypic effect and allele frequency between rare and common variances are represented in Figure 1. Monogenic conditions are represented on the left side of the graph, where the phenotypic effect is very severe, but the frequency is low. The common variants fall on the middle and right side of the graph, with variants exerting a small phenotypic effect that is not deleterious and are present in high frequency in the population. On their own, these variants cannot result in a pronounced phenotype, but the co-occurrence of many of these common variants may cumulatively increase genetic risk for these diseases. Dyslipidemia and atherosclerosis are complex phenotypes, and thus the genetic component of these disease is also the result of complex interactions between various metabolic pathways [14].
These polygenic, common variants associated with dyslipidemia have been identified through genome-wide association studies (GWAS). Over 150 loci have been specifically associated with blood lipid concentrations (total cholesterol (TC), TG, HDL, and LDL) in European populations [16,17]. Notably, several of the identified variants were in biologically and clinically relevant genes, such as angiopoietin-like proteins 3 and 4 (ANGPTL3/4), inhibitors of LPL, and HMGCR, which codes for 3-hydroxy-3-methylglutaryl-CoA reductase, a target for statin therapy and the rate-limiting enzyme in cholesterol synthesis. GWAS are a powerful and hypothesis-generating tool that can identify loci that are associated with phenotypes of dyslipidemia, research into the effects of the physiological relevance and implication of functional variants in dyslipidemia phenotypes will further increase the understanding of this complex disease.
3.1. Focus on Physiological Relevance
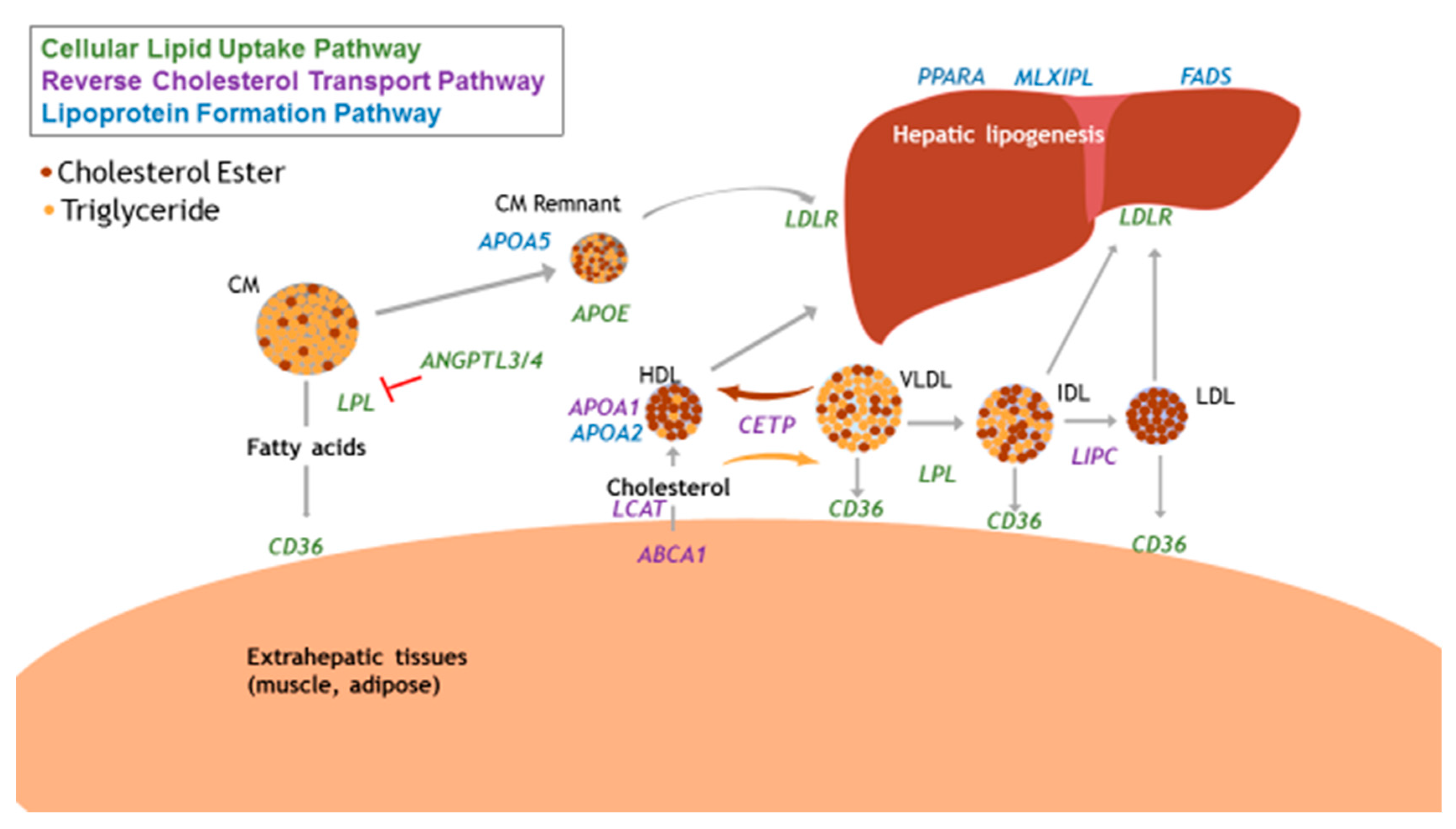
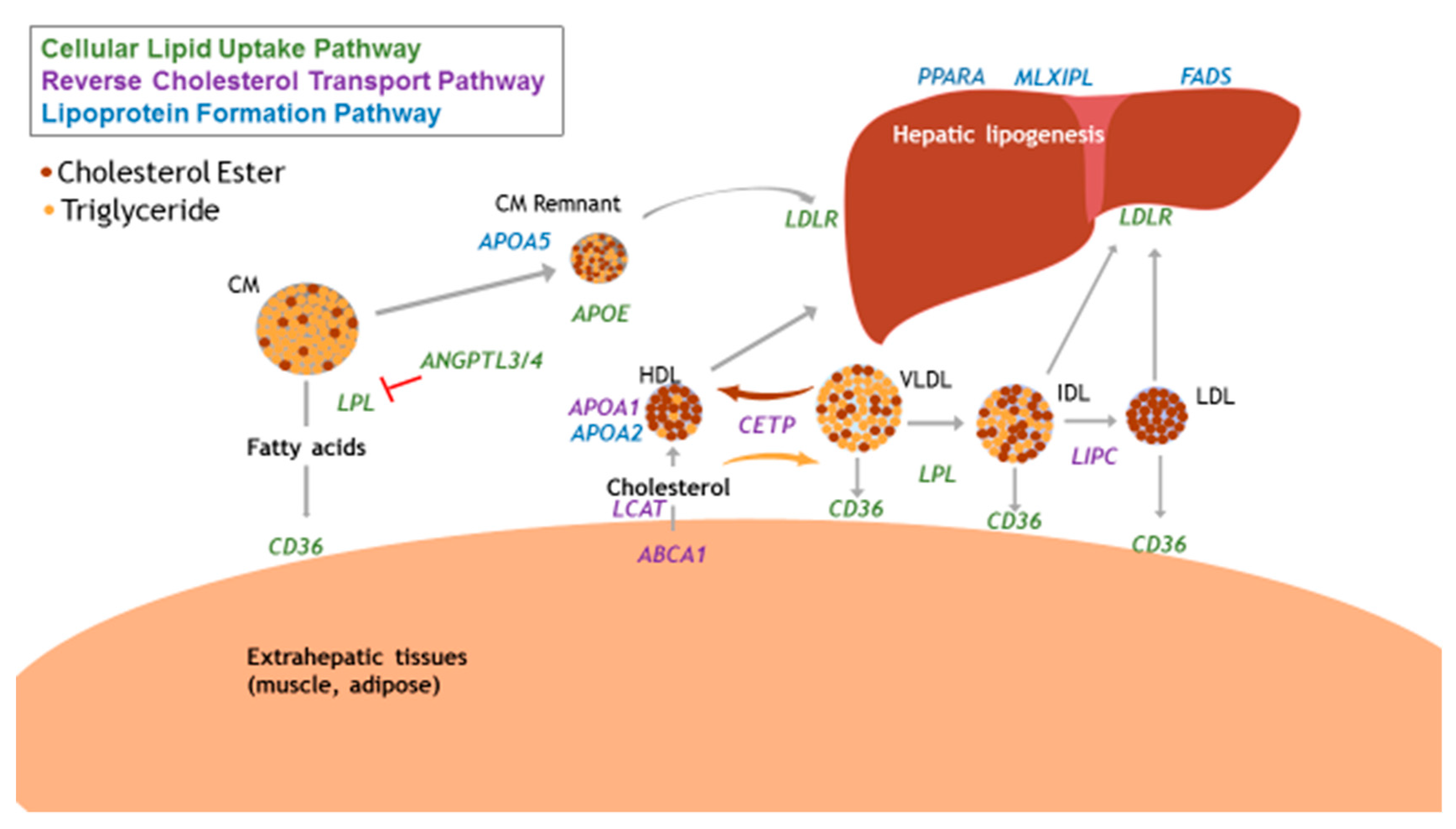
Not all associated common variants are in physiologically relevant pathways, and therefore cannot provide insight into the mechanisms by which nutrients interact with metabolic processes to produce phenotypes of dyslipidemia. In the pathophysiology and progression of atherogenic dyslipidemia, relevant pathways can include reverse cholesterol transport, cellular lipid uptake, and lipoprotein formation. The interactions of these pathways are depicted in Figure 2.
Reverse cholesterol transport (RCT) facilitates the return of excess cholesterol from peripheral tissues to the liver to be excreted from the body as bile [18]. Key proteins in this pathway include ATP-binding cassette subfamily A member 1 (ABCA1), cholesterol-ester transfer protein (CETP), apolipoprotein A1 (APOA1), hepatic lipase (HL, gene name: LIPC), and lecithin: cholesterol acyltransferase (LCAT), which serve to regulate concentrations of HDL and TG in circulation. Altered functionality of the RCT pathway can lead to decreased HDL concentrations, as fewer cholesteryl ester particles are accumulated within HDL particles [19]. SNPs in these genes have been previously associated with blood lipids in various populations. ABCA1 is essential in the efflux of cholesterol from peripheral tissues, and complete knockout of this protein results in Tangier disease. However, this gene contains several common polymorphisms that have been associated with HDL [20] and TG concentrations [21]. Mirmiran et al. recently described the gene-diet interactions of five CETP variants in observational and intervention studies [22]. These authors reported significant interactions between CETP genotype and dietary components, including alcohol and fat intake, to associate with blood lipid profiles. Interestingly, Nakamura et al. has reported evidence for the combined effects of multiple SNPs in the ABCA1 and CETP genes, suggesting a more significant genetic contribution to blood lipid concentrations when these variants are considered together, rather than on their own [23]. APOA1 is the predominant apolipoprotein on HDL particles and essential in RCT function and HDL formation. Variants in this gene have been associated with blood lipids in both European and Chinese populations [16,24]. HL is involved in the remodeling of HDL particles, and thus facilitates RCT [25]. Polymorphisms in the coding and promotor regions of LIPC have been identified through GWAS, and subsequently studied for associations with blood lipids in diverse populations [26,27]. SNPs in LIPC have also been implicated in affecting lipid response to weight loss interventions [28]. The effect of LIPC polymorphisms on blood lipids is more well-defined compared to other genes in this pathway, due to the extensive body of evidence conducted on this gene. LCAT is another protein involved in HDL maturation, as it is responsible for synthesis of cholesteryl ester in plasma. Due to its functional role in RCT, it is logical that the majority of candidate gene studies have focused on HDL as the phenotypic outcome of interest. Significant associations have been detected between LCAT polymorphisms and HDL in clinical populations, but there is not substantial evidence to definitively conclude that variants in this gene strongly impact blood lipid concentrations [29].
Cellular lipid uptake refers to the movement of dietary or endogenously produced lipids and lipoproteins through circulation, peripheral tissues, and the liver [30,31]. Key proteins in this pathway include lipoprotein lipase (LPL), LDLR, ANGPTL3/4, and fatty acid translocase (cluster of differentiation 36, CD36). LPL is present on cellular membranes and is involved in the lipolysis of TG in lipoproteins to fatty acids. Several common variants in the LPL gene have been associated with blood lipids, including a gain-of-function mutation that is associated with TG concentrations in European, but not African, populations [32]. Several SNPs have also been associated with HDL concentrations and high-fat diet [33], indicating the importance of this protein in the metabolism of dietary and endogenous lipids. LDLR is expressed primarily in hepatocytes, and polymorphisms in this gene can affect protein functionality, splicing, or transcription. Associations between variants in LDLR and adverse blood lipid concentrations have been detected in GWAS [34], and one variant, rs688, has been studied in vitro to determine the mechanistic consequences of this polymorphism on altered protein functionality [35]. ANGPTL 3 and 4 inhibit LPL in cardiac and skeletal muscle and adipose tissue, preventing the lipolysis and removal of TG from circulation. ANGPTL3 expression also results in lower LDL production through increased clearance of ApoB-containing lipoproteins [36]. ANGPTL4 is induced in the fasting state, allowing for increased delivery of fatty acids to tissues other than adipose. Genetic associations between variants in ANGPTL4 and both LDL [16] and HDL concentrations [37] have been reported in European populations. The consequences of ANGPTL variants on dyslipidemia was recently summarized by Paththinige et al. [38]. CD36 is involved in the cellular uptake of both dietary and endogenous lipids, and variants in the CD36 gene were first associated with blood lipids by Ma et al. [39]. Mechanistically, CD36 is a logical target for gene-diet interaction studies, and polymorphisms have been associated with blood lipids in diverse populations with [40] and without the inclusion of dietary intake [41].
The endogenous synthesis and export of lipids and lipoproteins from the liver also has clinical relevance in dyslipidemia and obesity, as excess energy intake can upregulate these processes [42,43]. Common variants in these pathways can alter the functionality of encoded proteins, resulting in metabolic alterations and phenotypic traits such as dyslipidemia [44]. Genetic variants present in genes coding for apolipoproteins also have been evidenced to impact risk of dyslipidemia and atherosclerotic disease. The most classic example is APOE, coding for apolipoprotein E (APOE). APOE circulates on lipoproteins in both systemic circulation and the central nervous system. The isoforms of this gene affect the affinity of APOE to its binding protein, and the E4 genotype has been associated with increased CVD risk and elevated blood lipid levels, and has been summarized previously [45]. Genes coding for other apolipoproteins also contain common variants previously been associated with blood lipids and have functional relevance for dyslipidemia, such as APOA5 and APOA2. Variants in APOA5 have been evidenced to significantly impact TG concentrations and were recently summarized by Guardiola and Ribalta [46]. APOA2 is also associated with HDL, and variants in this gene have been evidenced to interact with dietary fat intake to affect inflammatory status among individuals with diabetes, although the mechanism remains to be elucidated [47,48]. Regarding endogenous lipogenesis, key proteins include fatty acid desaturase (FADS) and peroxisome-proliferator activator receptor alpha (PPARA). Genes in the FADS cluster (FADS1, FADS2, FADS3) code for proteins responsible for desaturation of dietary and endogenous lipids, and variants in these genes have been associated with circulating polyunsaturated fatty acid (PUFA) concentrations, as it is postulated that presence of certain polymorphisms results in decreased functionality of the enzymes [49,50,51]. Additionally, we and others have published on the associations between FADS SNPs and blood lipids [52,53]. PPARA regulates a host of lipid and glucose homeostatic processes in the liver, and as PUFAs are ligands for all PPAR isoforms, these genes are targets for gene-diet interaction studies [54]. Variants in PPARA have been evidenced to influence blood lipid concentrations in the context of a high-fat diet [55,56]. Regarding nutritional control of hepatic TG synthesis, max-like protein X (MLX) interacting protein like (MLXIPL) induces these pathways in a carbohydrate-dependent manner (an alias for MLXIPL is carbohydrate-response element binding protein, ChREBP). Variants in MLXIPL have been examined as a mechanism for elevated TG concentrations, and associations have been detected in a Chinese population [57]. However, there are other transcriptional regulators of lipid synthesis, such as sterol-regulatory element binding protein 1 (SREBP1) and upstream transcription factor (USF), that have not been extensively explored in genetic associations. The study of pathways involved in blood lipid concentrations is necessary to better understand of the biological aspects of dyslipidemia and potentially identify new targets in specific proteins or pathways to develop preventative and treatment therapies.
3.2. Differences in Minor Allele Frequency and Special Populations
The majority of genetic association studies have been conducted among individuals of European descent; as of 2011, only 4% of GWAS had been conducted in non-European populations [58]. Evidence from genetic association studies in non-European populations have concluded that findings from one study may not always apply to other populations. Examples of this include differences in minor allele frequency (MAF) across populations, differences in risk allele, and discovery of novel candidate loci. The study of genetic associations with dyslipidemia is necessary to better understand the biological reasons for increased disease prevalence among certain ethnic groups. The Mexican population has one of the highest prevalence of dyslipidemias, with low HDL and elevated TG concentrations affecting 61% and 32% of the adult population, respectively [59]. The following examples highlight some of the genetic studies in this population and the need for further study of the genetic and environmental basis for the disproportionate rates of dyslipidemia. The differences in MAF have been highlighted by the 1000 Genomes Project, and have informed databases such as the dbSNP database of National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/snp) [60]. One such example of major differences in MAF and CVD risk is rs1800588 (LIPC). This SNP has a global MAF of 0.39, but is as high as 0.50 in Mexican populations (1000 Genomes). The rs1800588 genotype has been associated with TG concentrations among Mexican adults [61]. The current MAF and risk allele definitions for Mexican populations that come from the 1000 Genomes Project is from 107 individuals living in Los Angeles, United States (US). This is not a large enough sample to generalize for the entire Mexican population in both the US and Mexico. Our group has published on a larger cohort study of almost 1000 individuals from Mexico, and have detected differences in minor and risk alleles for SNPs in the FADS cluster [53]. Furthermore, a recent GWAS conducted in Mexicans identified novel genetic loci to be associated with TG concentrations [62]. These variants had not been identified in previous studies, thus emphasizing the need for further investigation into the genetic effects of dyslipidemia in this understudied, at-risk group. The lack of diversity in genomic research is limiting the implementation of precision medicine and nutrition recommendations for people of diverse ethnicities [63].
The lack of reproducibility among genetic association studies is particularly problematic for diverse populations. These ethnic subgroups, such as Hispanic and African-Americans, have some of the highest prevalence of dyslipidemia and other chronic, non-communicable diseases in the US [64]. The results from genetic association studies conducted in populations of European ancestry may not translate to these diverse populations, delaying the benefit that these individuals might receive as medicine moves toward the direction of personalized, genotype-based recommendations.
4. Dietary Contributions to Dyslipidemia
Dietary intake has a crucial role in affecting metabolic health and disease risk. Dietary components that have been previously implicated in increasing blood lipid concentrations include alcohol, carbohydrates, and dietary fat. The role of alcohol [65] and carbohydrates [66] have been previously reviewed; therefore, the current review will focus on dietary fat. Currently, agencies such as the American Heart Association (AHA), the Department of Agriculture, and the Department of Health and Human Services to recommend the limiting of total and saturated fat (SFA) [2,67]. However, the recommendation of a low-fat diet for heart health was challenged when researchers from the Seven Countries Study observed a low prevalence of CVD in the Mediterranean region, despite the consumption of a diet containing a moderate amount of total fat, coming from olive oil and cold-water fish [68,69]. This led to several seminal clinical trials to explore the effects of the Mediterranean diet on CVD risk and mortality, such as the Lyon Heart Study and PREDIMED [70,71]. The promising results of these trials have led to further scientific exploration of differential effects of various types of dietary fatty acids in ameliorating or exacerbating CVD risk. The replacement of saturated for unsaturated fat in the diet has been evidenced to be lipid-lowering and protective against CVD [72,73].
Unsaturated fats, those present in high amounts in the Mediterranean diet described above, are classified into monounsaturated (MUFA) and polyunsaturated (PUFA) fatty acids. MUFAs are present in foods such as avocados, almonds and other nuts, and vegetable oils. MUFAs are cardioprotective in that they do not raise blood lipid concentrations, and are less susceptible to oxidation than are PUFAs, due to their lower degree of unsaturation [74]. MUFAs are also more effective than carbohydrates in reducing blood lipid concentrations when replaced for SFA in the diet [75]. The majority of studies examining MUFA intake and blood lipids have been conducted concurrently with consumption of a Mediterranean diet, which has been associated with lower TG concentrations in meta-analyses [76,77]. However, as there are additional dietary components present in a Mediterranean diet, such as fiber, micronutrients, phytochemicals, and PUFAs, it is difficult to elucidate the specific effects of MUFAs alone on blood lipids. The effects of PUFA intake, present in cold-water fish, walnuts, and corn oil, on blood lipids and CVD risk have been studied extensively [78]. Intake of n-3 and n-6 PUFAs have both been associated with decreased CVD risk, especially the n-3 series, as they have been evidenced to have anti-arrhythmic and potent TG-lowering effects [79]. N-3 PUFA supplementation is currently recommended by the AHA to prevent recurrence of myocardial infarction [80].
The mechanisms by which unsaturated fatty acids affect blood lipid profiles have been previously summarized [70,72,74,81]. In brief, PUFA can serve to upregulate mRNA and thus protein levels of LDL receptors, resulting in increased lipoprotein uptake to the liver [82]. PUFA also downregulate fatty acid synthase, a key step in de novo lipogenesis, and very-low density lipoprotein (VLDL) secretion from the liver [83]. MUFA has been associated with decreased apolipoprotein C-III, which is an activator of LPL [84]. Clinical studies have also shown the effects of MUFA intake on decreasing apoB-100 production, the primary apolipoprotein present on circulating VLDL [81].
The lipid and lipoprotein response to intake of these various fatty acids may not be consistent among different populations studied. The reasons for this variability can be due to age, sex, disease state, differences at the genetic level, or any combination of these factors. The role of individual genetic variation in determining differential phenotypes has become more clearly understood, as advances in genetic technology and large cohort studies have identified significant associations between dietary intake and genetic variants to produce differences in disease risk [85]. The field of nutrigenomics refers broadly to the study of the interactions between dietary intake and the genome [86]. These interactions can result in epigenetic modification of genes, transcriptional regulation, or alterations in protein functionality. A subset of nutrigenomics is nutrigenetics, which specifically examines the effect of individual genetic variants (i.e., SNPs) and dietary intake on phenotypic expression. The exploration of these interactions can direct the creation of personalized recommendations for consumption of certain dietary fatty acids for the maintenance of normal lipid profiles and achievement of a healthy weight.
5. Nutrient-Gene Interactions and Dyslipidemia
Nutrigenetics, the science of the effect of genetic variation on response to dietary intake, bridges the gap between biological (genetic) and behavioral (diet) factors contributing to complex diseases, and can offer explanation as to why researchers may observe differential effects among individuals fed identical diets. Knowledge gained from this field is promising, as it can lead to explanation of response variability in clinical trials with diverse populations, better identification of non-responders to various diets, and the development of personalized dietary strategies [87].
As dyslipidemia is the result of a combination of genetic and environmental factors, it is logical that these two elements be examined in conjunction with one another. As many genetic loci associated with dyslipidemia have already been identified, the next step is to identify gene-environment interactions that may exacerbate or ameliorate the effect of genetic variation on disease risk. The integration of environmental exposures, especially dietary intake, may be able to add to the understanding of the complex etiologies associated with dyslipidemia. Furthermore, a targeted approach through the selection of physiologically relevant genes involved in pathways of lipoprotein metabolism and atherosclerosis will yield important discovery into the biology behind how these proteins metabolize nutrients in the presence of genetic mutations [14].
Previous research conducted to further classify the effects of common variants and dietary fat intake on lipid profiles has provided initial evidence of the need for deeper understanding in this field. Current literature on gene-diet interactions of physiologically relevant genes and dietary fat intake is presented in Table 2. A classic example, from Ordovas et al. examined the interactive effects of a polymorphism in the promotor region (rs1800588) of the LIPC gene and total fat intake [88]. This polymorphism has been associated with decreased activity of hepatic lipase. Mutations in this enzyme can result in elevated TG. Results indicated that rs1800588 genotype was significantly associated with HDL concentrations, and this association was strengthened when dietary intake was also considered. The interaction between presence of the risk allele and fat intake greater than 30% of total calories was associated with increased HDL, suggesting that these individuals may benefit from a high fat diet, specifically one high in MUFAs [89]. A diet high in total fat, defined in a study by Sanchez-Moreno et al. as consumption greater than 98 grams per day (the study median), did not associate with significantly higher TG concentrations by APOA5 (rs662799) genotype, indicating there was no disadvantage to this mutation in individuals’ ability to metabolize a high-fat diet [90]. These studies indicate that a high-fat diet may be beneficial in maintaining desired blood lipid concentrations for individuals possessing the minor alleles of common variants.
The nutrigenetic interactions between n-3 PUFA intake and common variants in genes related to atherosclerosis are summarized by Merched and Chan [91]. PUFA intake has been evidenced to interact with a polymorphism in PPARA, a transcriptional regulator of lipid metabolism, to associate with lower TG concentrations among those consuming high PUFA intake [92]. Several variants in APOA5, coding for apolipoprotein AV, have been evidence to interacte with PUFA intake to associate with elevated TG concentrations in individuals possessing the risk allele [93]. However, the APOA5 gene has 14 known SNPs listed on the NCBI database alone, so it is quite possible that these variants may interact with one another, or other genetic or non-genetic factors, to affect response to dietary intervention.
As we recognize the need to translate basic science into clinical applications, individual genetic variants have been studied as predictors of the lipid response to dietary interventions among individuals with obesity [94,95]. Several studies have specifically examined the role of physiologically relevant variants on changes in lipid profiles, with promising results. Zhang et al., studied the role of the rs964184 variant (APOA5) on modifying changes in TC, HDL, and LDL after the POUNDS LOST (Preventing Overweight Using Novel Dietary Strategies) weight loss trial, in which participants were randomized to follow one of four diets of varying macronutrient composition [96]. However, the POUNDS LOST dietary conditions varied in total fat content, and the researchers did not account for the dietary fat composition (SFA, MUFA, PUFA) of the diets. The type and amount of dietary fat intake is evidenced to interact with genetic variants to influence blood lipid profiles [97,98,99,100].
The gene-diet interactions of cholesterol metabolism were reviewed by Abdullah et al., in 2015 [101]. These researchers focused their review on the outcomes of TC, LDL, and HDL concentrations and any dietary exposure. This review provides strong evidence for the role of variants in over 20 genes involved in cholesterol metabolism. Nuno et al. recently reviewed the literature on variants in genes involved in lipid metabolism and dietary intake on CVD risk [102]. Many of the studies reported in this review describe significant associations with variants in several of the genes mentioned, blood lipids, and dietary intake of carbohydrates and various fatty acids. One notable limitation in these reviews is the limited research on dietary patterns, rather than consideration of macronutrient intake alone. As nutrients are not consumed in isolation, the study of the interacting effects of different dietary components, as in a Mediterranean diet, with individual genetic variation on disease risk is a promising direction for future study. Additionally, as fatty acids are heterogeneous, there must be more research on the interacting effects between different classes of fatty acids (medium- to long-chain, degree of saturation, etc.) and the foods they are present in and genetic variation. A relevant example of this is dairy fat. Dairy consumption has been associated with protective benefits against T2DM, obesity, and other cardiometabolic biomarkers [103,104]. A recent longitudinal study of over 2000 adults concluded that circulating levels of the fatty acids present in dairy products (pentadecanoic, heptadecanoic, and trans-palmitoleic) were not associated with mortality, but were associated with lower risk of CVD mortality [105]. More research into the effects of various foods high in SFA on risk factors for metabolic diseases are warranted, as these fatty acids are not consumed in isolation, and even foods high in SFA contain some degree of unsaturated fat.
These nutrient-gene interactions only represent a portion of the functional variants that have been associated with blood lipids in GWAS. Nevertheless, additional research is necessary, especially in diverse or minority populations across the lifecycle, to further elucidate the mechanisms by which diet can interact with genetic variation. The ultimate goal of this field of research is the translation of these discoveries into personalized nutrition recommendations to treat and prevent disease [106].
6. Conclusions
The genetic, dietary, and nutrigenetic components described here highlight the strong relationships between biological and behavioral risk factors for dyslipidemia. The exploration of physiologically relevant variants and their interactions with dietary lipids is especially pertinent to the development of personalized dietary recommendations for management of dyslipidemia. As obesity, a major risk factor for the development of dyslipidemia, continues to increase in prevalence worldwide, effective strategies to achieve a healthy weight and manage lipid profiles are needed. Due to genetic variation, among other factors, not all individuals will respond uniformly to these strategies. Thus, the identification of factors that explain this variability in response will provide researchers and clinicians with the information to apply targeted treatment approaches to maximize benefits against dyslipidemia. Understanding the biological reasons behind why an individual may not respond is a key research priority to be addressed, as this can lead to implementation of individualized nutrition recommendations that can be implemented to prevent and treat dyslipidemia [107].
Author Contributions
Conceptualization, B.A.H.; Writing-Original Draft Preparation, B.A.H.; Writing-Review & Editing, N.A.K. and M.T.G.; Supervision, N.A.K. and M.T.G.; All authors approved the final manuscript.
Funding
B.A.H. and M.T.G. are supported by the Agriculture and Food Research Initiative Competitive Grant no. 2015-68001-23248 from the USDA National Institute of Food and Agriculture to Cooperative Extension and the Department of Human Development and Family Studies at the University of Illinois at Urbana-Champaign. Support for this work also comes from Hatch Projects #ILLU-971-368, #ILLU-793-327, and the Department of Kinesiology and Community Health at the University of Illinois at Urbana-Champaign.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carroll, M.D.; Fryer, C.D.; Nguyen, D.T. High Total and Low High-Density Lipoprotein Cholesterol in Adults: United States, 2015–2016; NCHS Data Brief, No 290; National Center for Health Statistics: Hyattsville, MD, USA, 2017.
- Eckel, R.H.; Jakicic, J.M.; Ard, J.D.; Hubbard, V.S.; de Jesus, J.M.; Lee, I.-M.; Lichtenstein, A.H.; Loria, C.M.; Millen, B.E.; Miller, N.H.; et al. 2013 AHA/ACC Guideline on Lifestyle Management to Reduce Cardiovascular Risk: A Report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. Circulation 2013, 129, S76–S99. [Google Scholar] [CrossRef] [PubMed]
- Bamba, V. Update on screening, etiology, and treatment of dyslipidemia in children. JCEM 2014, 99, 3093–3102. [Google Scholar] [CrossRef] [PubMed]
- Kussmann, M.; Stover, P.J. Nutrigenomics and Proteomics in Health and Disease: Towards a Systems-Level Understanding of Gene-Diet Interactions, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017; p. 1119098831. [Google Scholar]
- Tenenbaum, A.; Klempfner, R.; Fisman, E.Z. Hypertriglyceridemia: A too long unfairly neglected major cardiovascular risk factor. Cardiovasc. Diabetol. 2014, 13, 159. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Stone, N.J.; Ballantyne, C.; Bittner, V.; Criqui, M.H.; Ginsberg, H.N.; Goldberg, A.C.; Howard, W.J.; Jacobson, M.S.; Kris-Etherton, P.M.; et al. Triglycerides and cardiovascular disease: A scientific statement from the American Heart Association. Circulation 2011, 123, 2292–2333. [Google Scholar] [CrossRef] [PubMed]
- Wajchenberg, B.L. Subcutaneous and visceral adipose tissue: Their relation to the metabolic syndrome. Endocr. Rev. 2000, 21, 697–738. [Google Scholar] [CrossRef] [PubMed]
- Després, J.-P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.G.; Olsson, A.G.; Abt, M.; Ballantyne, C.M.; Barter, P.J.; Brumm, J.; Chaitman, B.R.; Holme, I.M.; Kallend, D.; Leiter, L.A. Effects of dalcetrapib in patients with a recent acute coronary syndrome. NEJM 2012, 367, 2089–2099. [Google Scholar] [CrossRef] [PubMed]
- Rader, D.J.; Hovingh, G.K. HDL and cardiovascular disease. Lancet 2014, 384, 618–625. [Google Scholar] [CrossRef]
- Marais, A.D. Familial hypercholesterolaemia. Clin. Biochem. Rev. 2004, 25, 49. [Google Scholar] [PubMed]
- Oliva, C.P.; Pisciotta, L.; Volti, G.L.; Sambataro, M.P.; Cantafora, A.; Bellocchio, A.; Catapano, A.; Tarugi, P.; Bertolini, S.; Calandra, S. Inherited apolipoprotein AV deficiency in severe hypertriglyceridemia. Arterioscl. Thromb. Vasc. Biol. 2005, 25, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Tolleshaug, H.; Hobgood, K.K.; Brown, M.S.; Goldstein, J.L. The LDL receptor locus in familial hypercholesterolemia: Multiple mutations disrupt transport and processing of a membrane receptor. Cell 1983, 32, 941–951. [Google Scholar] [CrossRef]
- Lusis, A.J.; Fogelman, A.M.; Fonarow, G.C. Genetic basis of atherosclerosis: Part I: New genes and pathways. Circulation 2004, 110, 1868–1873. [Google Scholar] [CrossRef] [PubMed]
- Dron, J.S.; Hegele, R.A. Genetics of lipid and lipoprotein disorders and traits. Curr. Genet. Med. Rep. 2016, 4, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Teslovich, T.M.; Musunuru, K.; Smith, A.V.; Edmondson, A.C.; Stylianou, I.M.; Koseki, M.; Pirruccello, J.P.; Ripatti, S.; Chasman, D.I.; Willer, C.J.; et al. Biological, clinical and population relevance of 95 loci for blood lipids. Nature 2010, 466, 707–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Lipids Genetics Consortium; Willer, C.J.; Schmidt, E.M.; Sengupta, S.; Peloso, G.M.; Gustafsson, S.; Kanoni, S.; Ganna, A.; Chen, J.; Buchkovich, M.L.; et al. Discovery and refinement of loci associated with lipid levels. Nat. Genet. 2013, 45, 1274. [Google Scholar] [CrossRef] [Green Version]
- Fielding, C.J.; Fielding, P.E. Molecular physiology of reverse cholesterol transport. J. Lipid Res. 1995, 36, 211–228. [Google Scholar] [PubMed]
- Tall, A.R. An overview of reverse cholesterol transport. Eur. Heart J. 1998, 19 (Suppl. A), A31–A35. [Google Scholar] [CrossRef]
- Clifford, A.J.; Rincon, G.; Owens, J.E.; Medrano, J.F.; Moshfegh, A.J.; Baer, D.J.; Novotny, J.A. Single nucleotide polymorphisms in CETP, SLC46A1, SLC19A1, CD36, BCMO1, APOA5, and ABCA1 are significant predictors of plasma HDL in healthy adults. Lipids Health Dis. 2013, 12, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Liu, Y.; Li, Y.; Zhang, H.; Yu, M.; Kanu, J.S.; Qiao, Y.; Tang, Y.; Zhen, Q.; Cheng, Y. Association of ATP-binding cassette transporter A1 gene polymorphisms with plasma lipid variability and coronary heart disease risk. Int. J. Clin. Exp. Path 2015, 8, 13441–13449. [Google Scholar]
- Mirmiran, P.; Esfandiar, Z.; Hosseini-Esfahani, F.; Koochakpoor, G.; Daneshpour, M.S.; Sedaghati-Khayat, B.; Azizi, F. Genetic variations of cholesteryl ester transfer protein and diet interactions in relation to lipid profiles and coronary heart disease: A systematic review. Nutr. Metab. 2017, 14, 77. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Niimura, H.; Kuwabara, K.; Takezaki, T.; Morita, E.; Wakai, K.; Hamajima, N.; Nishida, Y.; Turin, T.C.; Suzuki, S.; et al. Gene-Gene combination effect and interactions among ABCA1, APOA1, SR-B1, and CETP polymorphisms for serum high-density lipoprotein-cholesterol in the Japanese population. PLoS ONE 2013, 8, e82046. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Yin, R.-X.; Khounphinith, E.; Zhang, F.-H.; Yang, D.-Z.; Pan, S.-L. Association of the APOA1 rs964184 SNP and serum lipid traits in the Chinese Maonan and Han populations. Lipids Health Dis. 2018, 17, 105. [Google Scholar] [CrossRef] [PubMed]
- Deeb, S.S.; Zambon, A.; Carr, M.C.; Ayyobi, A.F.; Brunzell, J.D. Hepatic lipase and dyslipidemia: Interactions among genetic variants, obesity, gender, and diet. J. Lipid Res. 2003, 44, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Agirbasli, M.; Eren, F.; Agirbasli, D.; White, M.J.; Williams, S.M. Multi-Locus Candidate Gene Analyses of Lipid Levels in a Pediatric Turkish Cohort: Lessons Learned on LPL, CETP, LIPC, ABCA1, and SHBG. OMICS J. Integr. Biol. 2013, 17, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, D.; Ling, J.; Lu, W.; Zhang, S.; Zhu, Y.; Lai, M. Gender specific effect of LIPC C-514T polymorphism on obesity and relationship with plasma lipid levels in Chinese children. J. Cell. Mol. Med. 2015, 19, 2296–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Ng, S.S.; Bray, G.A.; Ryan, D.H.; Sacks, F.M.; Ning, G.; Qi, L. Dietary Fat Intake Modifies the Effect of a Common Variant in the LIPC Gene on Changes in Serum Lipid Concentrations during a Long-Term Weight-Loss Intervention Trial. J. Nutr. 2015, 145, 1289–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agirbasli, D.; Cirakoglu, B.; Eren, F.; Sumerkan, M.; Aksoy, S.; Aral, C.; Agirbasli, M. Effects of lecithin: Cholesterol acyltransferase genotypes, enzyme levels, and activity on high-density lipoprotein levels. J. Clin. Lipid 2011, 5, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, I.J.; Eckel, R.H.; Abumrad, N.A. Regulation of fatty acid uptake into tissues: Lipoprotein lipase and CD36-mediated pathways. J. Lipid Res. 2009, 50, S86–S90. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, R.L.; Febbraio, M. CD36, a scavenger receptor involved in immunity, metabolism, angiogenesis, and behavior. Sci. Signal. 2009, 2, re3. [Google Scholar] [CrossRef] [PubMed]
- Deo, R.C.; Reich, D.; Tandon, A.; Akylbekova, E.; Patterson, N.; Waliszewska, A.; Kathiresan, S.; Sarpong, D.; Taylor, H.A.; Wilson, J.G. Genetic Differences between the Determinants of Lipid Profile Phenotypes in African and European Americans: The Jackson Heart Study. PLoS Genet. 2009, 5, e1000342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayyappa, K.A.; Shatwan, I.; Bodhini, D.; Bramwell, L.R.; Ramya, K.; Sudha, V.; Anjana, R.M.; Lovegrove, J.A.; Mohan, V.; Radha, V.; et al. High fat diet modifies the association of lipoprotein lipase gene polymorphism with high density lipoprotein cholesterol in an Asian Indian population. Nutr. Metab. 2017, 14, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairoozy, R.H.; White, J.; Palmen, J.; Kalea, A.Z.; Humphries, S.E. Identification of the functional variant(s) that explain the low-density lipoprotein receptor (LDLR) GWAS SNP rs6511720 association with lower LDL-C and risk of CHD. PLoS ONE 2016, 11, e0167676. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Ihn, H.E.; Medina, M.W.; Krauss, R.M. A common polymorphism in the LDL receptor gene has multiple effects on LDL receptor function. Hum. Mol. Genet. 2013, 22, 1424–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gusarova, V.; Banfi, S.; Gromada, J.; Cohen, J.C.; Hobbs, H.H. Inactivation of ANGPTL3 reduces hepatic VLDL-triglyceride secretion. J. Lipid Res. 2015, 56, 1296–1307. [Google Scholar] [CrossRef] [PubMed]
- Legry, V.; Bokor, S.; Cottel, D.; Beghin, L.; Catasta, G.; Nagy, E.; Gonzalez-Gross, M.; Spinneker, A.; Stehle, P.; Molnar, D.; et al. Associations between common genetic polymorphisms in angiopoietin-like proteins 3 and 4 and lipid metabolism and adiposity in European adolescents and adults. JCEM 2009, 94, 5070–5077. [Google Scholar] [CrossRef] [PubMed]
- Paththinige, C.S.; Sirisena, N.D.; Dissanayake, V.H.W. Genetic determinants of inherited susceptibility to hypercholesterolemia—A comprehensive literature review. Lipids Health Dis. 2017, 16, 103. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Bacci, S.; Mlynarski, W.; Gottardo, L.; Soccio, T.; Menzaghi, C.; Iori, E.; Lager, R.A.; Shroff, A.R.; Gervino, E.V.; et al. A common haplotype at the CD36 locus is associated with high free fatty acid levels and increased cardiovascular risk in Caucasians. Hum. Mol. Genet. 2004, 13, 2197–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, Y.; Kim, Y. Oily Fish Consumption Modifies the Association between CD36 rs6969989 Polymorphism and Lipid Profiles in Korean Women. Prev. Nutr. Food Sci. 2016, 21, 202–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Arellano, L.E.; Salgado-Bernabe, A.B.; Guzman-Guzman, I.P.; Salgado-Goytia, L.; Munoz-Valle, J.F.; Parra-Rojas, I. CD36 haplotypes are associated with lipid profile in normal-weight subjects. Lipids Health Dis. 2013, 12, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellsworth, J.L.; Erickson, S.K.; Cooper, A.D. Very low and low density lipoprotein synthesis and secretion by the human hepatoma cell line Hep-G2: Effects of free fatty acid. J. Lipid Res. 1986, 27, 858–874. [Google Scholar] [PubMed]
- Hellerstein, M.K.; Schwarz, J.M.; Neese, R.A. Regulation of Hepatic De Novo Lipogenesis in Humans. Ann. Rev. Nutr. 1996, 16, 523–557. [Google Scholar] [CrossRef] [PubMed]
- Kathiresan, S.; Willer, C.J.; Peloso, G.M.; Demissie, S.; Musunuru, K.; Schadt, E.E.; Kaplan, L.; Bennett, D.; Li, Y.; Tanaka, T. Common variants at 30 loci contribute to polygenic dyslipidemia. Nat. Genet. 2009, 41, 56. [Google Scholar] [CrossRef] [PubMed]
- Raichlen, D.A.; Alexander, G.E. Exercise, APOE genotype, and the evolution of the human lifespan. Trends Neurosci. 2014, 37, 247–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guardiola, M.; Ribalta, J. Update on APOA5 Genetics: Toward a Better Understanding of Its Physiological Impact. Curr. Atheroscler. Rep. 2017, 19, 30. [Google Scholar] [CrossRef] [PubMed]
- Keramat, L.; Sadrzadeh-Yeganeh, H.; Sotoudeh, G.; Zamani, E.; Eshraghian, M.; Mansoori, A.; Koohdani, F. Apolipoprotein A2 -265 T>C polymorphism interacts with dietary fatty acids intake to modulate inflammation in type 2 diabetes mellitus patients. Nutrition 2017, 37, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Zamani, E.; Sadrzadeh-Yeganeh, H.; Sotoudeh, G.; Keramat, L.; Eshraghian, M.; Rafiee, M.; Koohdani, F. The interaction between ApoA2 -265T>C polymorphism and dietary fatty acids intake on oxidative stress in patients with type 2 diabetes mellitus. Eur. J. Nutr. 2017, 56, 1931–1938. [Google Scholar] [CrossRef] [PubMed]
- Bokor, S.; Dumont, J.; Spinneker, A.; Gonzalez-Gross, M.; Nova, E.; Widhalm, K.; Moschonis, G.; Stehle, P.; Amouyel, P.; De Henauw, S.; et al. Single nucleotide polymorphisms in the FADS gene cluster are associated with delta-5 and delta-6 desaturase activities estimated by serum fatty acid ratios. J. Lipid Res. 2010, 51, 2325–2333. [Google Scholar] [CrossRef] [PubMed]
- Mathias, R.A.; Sergeant, S.; Ruczinski, I.; Torgerson, D.G.; Hugenschmidt, C.E.; Kubala, M.; Vaidya, D.; Suktitipat, B.; Ziegler, J.T.; Ivester, P.; et al. The impact of FADS genetic variants on ω6 polyunsaturated fatty acid metabolism in African Americans. BMC Genet. 2011, 12, 50–50. [Google Scholar] [CrossRef] [PubMed]
- Merino, D.M.; Johnston, H.; Clarke, S.; Roke, K.; Nielsen, D.; Badawi, A.; El-Sohemy, A.; Ma, D.W.; Mutch, D.M. Polymorphisms in FADS1 and FADS2 alter desaturase activity in young Caucasian and Asian adults. Mol. Genet. Metab. 2011, 103, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Sone, Y.; Kido, T.; Ainuki, T.; Sonoda, M.; Ichi, I.; Kodama, S.; Sone, H.; Kondo, K.; Morita, Y.; Egawa, S.; et al. Genetic variants of the fatty acid desaturase gene cluster are associated with plasma LDL cholesterol levels in Japanese males. J. Nutr. Sci. Vitaminol. 2013, 59, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Vidal, I.; Voruganti, V.S.; Hannon, B.A.; Andrade, F.C.D.; Aradillas-Garcia, C.; Nakamura, M.T.; Teran-Garcia, M. Serum Lipid Concentrations and FADS Genetic Variants in Young Mexican College Students: The UP-AMIGOS Cohort Study. Lifestyle Genom. 2018, 1–9. [Google Scholar] [CrossRef]
- Yilmaz-Aydogan, H.; Kurnaz, O.; Kucukhuseyin, O.; Akadam-Teker, B.; Kurt, O.; Eronat, A.P.; Tekeli, A.; Bugra, Z.; Ozturk, O. Different effects of PPARA, PPARG and ApoE SNPs on serum lipids in patients with coronary heart disease based on the presence of diabetes. Gene 2013, 523, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Alsaleh, A.; Frost, G.S.; Griffin, B.A.; Lovegrove, J.A.; Jebb, S.A.; Sanders, T.A.; O’Dell, S.D. PPARgamma2 gene Pro12Ala and PPARalpha gene Leu162Val single nucleotide polymorphisms interact with dietary intake of fat in determination of plasma lipid concentrations. J. Nutrigenet. Nutrigenom. 2011, 4, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlSaleh, A.; Sanders, T.A.; O’Dell, S.D. Effect of interaction between PPARG, PPARA and ADIPOQ gene variants and dietary fatty acids on plasma lipid profile and adiponectin concentration in a large intervention study. Proc. Nutr. Soc. 2012, 71, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Aung, L.-H.-H.; Yin, R.-X.; Wu, J.-Z.; Wu, D.-F.; Wang, W.; Li, H. Association between the MLX interacting protein-like, BUD13 homolog and zinc finger protein 259 gene polymorphisms and serum lipid levels. Sci. Rep. 2014, 4, 5565. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, C.D.; Burchard, E.G.; De la Vega, F.M. Genomics for the world. Nature 2011, 475, 163–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Salinas, C.A.; Gomez-Perez, F.J.; Rull, J.; Villalpando, S.; Barquera, S.; Rojas, R. Prevalence of dyslipidemias in the Mexican National Health and Nutrition Survey 2006. Salud Publica Mexico 2010, 52 (Suppl. 1), S44–S53. [Google Scholar] [CrossRef]
- The Genomes Project Consortium; Auton, A.; Abecasis, G.R.; Altshuler, D.M.; Durbin, R.M.; Abecasis, G.R.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Donnelly, P.; et al. A global reference for human genetic variation. Nature 2015, 526, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posadas-Sanchez, R.; Ocampo-Arcos, W.A.; Lopez-Uribe, A.R.; Posadas-Romero, C.; Villarreal-Molina, T.; Leon, E.A.; Perez-Hernandez, N.; Rodriguez-Perez, J.M.; Cardoso-Saldana, G.; Medina-Urrutia, A.; et al. Hepatic lipase (LIPC) C-514T gene polymorphism is associated with cardiometabolic parameters and cardiovascular risk factors but not with fatty liver in Mexican population. Exp. Mol. Pathol. 2015, 98, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Weissglas-Volkov, D.; Aguilar-Salinas, C.A.; Nikkola, E.; Deere, K.A.; Cruz-Bautista, I.; Arellano-Campos, O.; Muñoz-Hernandez, L.L.; Gomez-Munguia, L.; Ordoñez-Sánchez, M.L.; Reddy, P.M.V.L.; et al. Genomic study in Mexicans identifies a new locus for triglycerides and refines European lipid loci. J. Med. Genet. 2013, 50, 298–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dainis, A.M.; Ashley, E.A. Cardiovascular Precision Medicine in the Genomics Era. JACC Basic Transl. Sci. 2018, 3, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; de Ferranti, S.; Despres, J.P.; Fullerton, H.J.; Howard, V.J.; et al. Heart disease and stroke statistics—2015 update: A report from the American Heart Association. Circulation 2015, 131, e29–e322. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, I. Alcohol and Dyslipidemia. In Alcohol, Nutrition, and Health Consequences; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 329–339. [Google Scholar]
- Feinman, R.D.; Volek, J.S. Carbohydrate restriction as the default treatment for type 2 diabetes and metabolic syndrome. Scand. Cardiovasc. J. 2008, 42, 256–263. [Google Scholar] [CrossRef] [PubMed]
- US Department of Health and Human Services. 2015–2020 Dietary Guidelines for Americans; USDA: Washington, DC, USA, 2015. [Google Scholar]
- Bach-Faig, A.; Berry, E.M.; Lairon, D.; Reguant, J.; Trichopoulou, A.; Dernini, S.; Medina, F.X.; Battino, M.; Belahsen, R.; Miranda, G.; et al. Mediterranean diet pyramid today: Science and cultural updates. Public Health Nutr. 2011, 14, 2274–2284. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.C.; Sacks, F.; Trichopoulou, A.; Drescher, G.; Ferro-Luzzi, A.; Helsing, E.; Trichopoulos, D. Mediterranean diet pyramid: A cultural model for healthy eating. Am. J. Clin. Nutr. 1995, 61, 1402S–1406S. [Google Scholar] [CrossRef] [PubMed]
- De Lorgeril, M.; Salen, P.; Martin, J.-L.; Monjaud, I.; Delaye, J.; Mamelle, N. Mediterranean diet, traditional risk factors, and the rate of cardiovascular complications after myocardial infarction: Final report of the Lyon Diet Heart Study. Circulation 1999, 99, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J. Primary prevention of cardiovascular disease with a Mediterranean diet. NEJM 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Hannon, B.A.; Thompson, S.V.; An, R.; Teran-Garcia, M. Clinical outcomes of dietary replacement of saturated fatty acids with unsaturated fat sources in adults with overweight and obesity: A systematic review and meta-analysis of randomized control trials. Ann. Nutr. Metab. 2017, 71, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.; Martin, N.; Abdelhamid, A.; Davey Smith, G. Reduction in saturated fat intake for cardiovascular disease. Cochrane Database Syst. Rev. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kris-Etherton, P.M. Monounsaturated fatty acids and risk of cardiovascular disease. Circulation 1999, 100, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Sacks, F.M.; Lichtenstein, A.H.; Wu, J.H.Y.; Appel, L.J.; Creager, M.A.; Kris-Etherton, P.M.; Miller, M.; Rimm, E.B.; Rudel, L.L.; Robinson, J.G.; et al. Dietary Fats and Cardiovascular Disease: A Presidential Advisory From the American Heart Association. Circulation 2017. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.; Bryan, J.; Hodgson, J.; Murphy, K. Definition of the Mediterranean diet: A literature review. Nutrients 2015, 7, 9139–9153. [Google Scholar] [CrossRef] [PubMed]
- Kastorini, C.M.; Milionis, H.J.; Esposito, K.; Giugliano, D.; Goudevenos, J.A.; Panagiotakos, D.B. The effect of Mediterranean diet on metabolic syndrome and its components: A meta-analysis of 50 studies and 534,906 individuals. J. Am. Coll. Cardiol. 2011, 57, 1299–1313. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, A.S.; Martin, N.; Bridges, C.; Brainard, J.S.; Wang, X.; Brown, T.J.; Hanson, S.; Jimoh, O.F.; Ajabnoor, S.M.; Deane, K.H.O.; et al. Polyunsaturated fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Databse Syst. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Erkkilä, A.; de Mello, V.D.F.; Risérus, U.; Laaksonen, D.E. Dietary fatty acids and cardiovascular disease: An epidemiological approach. Prog. Lipid Res. 2008, 47, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Siscovick, D.S.; Barringer, T.A.; Fretts, A.M.; Wu, J.H.Y.; Lichtenstein, A.H.; Costello, R.B.; Kris-Etherton, P.M.; Jacobson, T.A.; Engler, M.B.; Alger, H.M.; et al. Omega-3 polyunsaturated fatty acid (fish oil) supplementation and the prevention of clinical cardiovascular disease. Sci. Advis. Am. Heart Assoc. 2017, 135, e867–e884. [Google Scholar] [CrossRef]
- Lamarche, B.; Couture, P. Dietary fatty acids, dietary patterns, and lipoprotein metabolism. Curr. Opin. Lipidol. 2015, 26, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.L.; West, K.L. Mechanisms by which dietary fatty acids modulate plasma lipids. J. Nutr. 2005, 135, 2075–2078. [Google Scholar] [CrossRef] [PubMed]
- Teran-Garcia, M.; Rufo, C.; Nakamura, M.T.; Osborne, T.F.; Clarke, S.D. NF-Y involvement in the polyunsaturated fat inhibition of fatty acid synthase gene transcription. Biochem. Piophys. Res. Commun. 2002, 290, 1295–1299. [Google Scholar] [CrossRef] [PubMed]
- Brousseau, M.E.; Ordovas, J.M.; Osada, J.; Fasulo, J.; Robins, S.J.; Nicolosi, R.J.; Schaefer, E.J. Dietary monounsaturated and polyunsaturated fatty acids are comparable in their effects on hepatic apolipoprotein mRNA abundance and liver lipid concentrations when substituted for saturated fatty acids in cynomolgus monkeys. J. Nutr. 1995, 125, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, O.; Milagro, F.I.; Allayee, H.; Chmurzynska, A.; Choi, M.S.; Curi, R.; De Caterina, R.; Ferguson, L.R.; Goni, L.; Kang, J.X.; et al. Guide for current nutrigenetic, nutrigenomic, and nutriepigenetic approaches for precision nutrition involving the prevention and management of chronic diseases associated with Oobesity. Lifestyle Genom. 2017, 10, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Ordovas, J.M.; Ferguson, L.R.; Tai, E.S.; Mathers, J.C. Personalised nutrition and health. BMJ 2018, 361. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; El-Sohemy, A.; Cahill, L.; Ferguson, L.R.; French, T.-A.C.; Tai, E.S.; Milner, J.; Koh, W.-P.; Xie, L.; Zucker, M.; et al. Nutrigenetics and nutrigenomics: Viewpoints on the current status and applications in nutrition research and practice. J. Nutrigenet. Nutrigenom. 2011, 4, 69–89. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Sparks, D.L. Hepatic lipase, high density lipoproteins, and hypertriglyceridemia. Am. J. Pathol. 2011, 178, 1429–1433. [Google Scholar] [CrossRef] [PubMed]
- Ordovas, J.M.; Corella, D.; Demissie, S.; Cupples, L.A.; Couture, P.; Coltell, O.; Wilson, P.W.F.; Schaefer, E.J.; Tucker, K.L. Dietary fat intake determines the effect of a common polymorphism in the hepatic lipase gene promoter on high-density lipoprotein metabolism: Evidence of a strong dose effect in this gene-nutrient interaction in the Framingham study. Circulation 2002, 106, 2315–2321. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moreno, C.; Ordovás, J.M.; Smith, C.E.; Baraza, J.C.; Lee, Y.-C.; Garaulet, M. APOA5 gene variation interacts with dietary fat intake to modulate obesity and circulating triglycerides in a Mediterranean population. J. Nutr. 2011, 141, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Merched, A.J.; Chan, L. Nutrigenetics and nutrigenomics of atherosclerosis. Curr. Atheroscler. Rep. 2013, 15, 328–328. [Google Scholar] [CrossRef] [PubMed]
- Tai, E.S.; Corella, D.; Demissie, S.; Cupples, L.A.; Coltell, O.; Schaefer, E.J.; Tucker, K.L.; Ordovas, J.M. Polyunsaturated fatty acids interact with the PPARA-L162V polymorphism to affect plasma triglyceride and apolipoprotein C-III concentrations in the Framingham Heart Study. J. Nutr. 2005, 135, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.Q.; Corella, D.; Demissie, S.; Cupples, L.A.; Adiconis, X.; Zhu, Y.; Parnell, L.D.; Tucker, K.L.; Ordovas, J.M. Dietary intake of n-6 fatty acids modulates effect of apolipoprotein A5 gene on plasma fasting triglycerides, remnant lipoprotein concentrations, and lipoprotein particle size: The Framingham Heart Study. Circulation 2006, 113, 2062–2070. [Google Scholar] [CrossRef] [PubMed]
- Garaulet, M.; Smith, C.E.; Hernandez-Gonzalez, T.; Lee, Y.C.; Ordovas, J.M. PPARgamma Pro12Ala interacts with fat intake for obesity and weight loss in a behavioural treatment based on the Mediterranean diet. Mol. Nutr. Food Res. 2011, 55, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qi, Q.; Zhang, C.; Smith, S.R.; Hu, F.B.; Sacks, F.M.; Bray, G.A.; Qi, L. FTO Genotype and 2-Year Change in Body Composition and Fat Distribution in Response to Weight-Loss Diets: The POUNDS LOST Trial. Diabetes 2012, 61, 3005–3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Qi, Q.; Bray, G.A.; Hu, F.B.; Sacks, F.M.; Qi, L. APOA5 genotype modulates 2-y changes in lipid profile in response to weight-loss diet intervention: The Pounds Lost Trial. Am. J. Clin. Nutr. 2012, 96, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Corella, D.; Peloso, G.; Arnett, D.K.; Demissie, S.; Cupples, L.A.; Tucker, K.; Lai, C.Q.; Parnell, L.D.; Coltell, O.; Lee, Y.C.; et al. APOA2, dietary fat, and body mass index: Replication of a gene-diet interaction in 3 independent populations. Arch. Intern. Med. 2009, 169, 1897–1906. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Tucker, K.L.; Smith, C.E.; Lee, Y.C.; Huang, T.; Richardson, K.; Parnell, L.D.; Lai, C.Q.; Young, K.L.; Justice, A.E.; et al. Lipoprotein lipase variants interact with polyunsaturated fatty acids for obesity traits in women: Replication in two populations. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1323–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robitaille, J.; Brouillette, C.; Lemieux, S.; Perusse, L.; Gaudet, D.; Vohl, M.C. Plasma concentrations of apolipoprotein B are modulated by a gene–diet interaction effect between the LFABP T94A polymorphism and dietary fat intake in French-Canadian men. Mol. Genet. Metab. 2004, 82, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.E.; Arnett, D.K.; Corella, D.; Tsai, M.Y.; Lai, C.Q.; Parnell, L.D.; Lee, Y.C.; Ordovas, J.M. Perilipin polymorphism interacts with saturated fat and carbohydrates to modulate insulin resistance. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 449–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, M.M.; Jones, P.J.; Eck, P.K. Nutrigenetics of cholesterol metabolism: Observational and dietary intervention studies in the postgenomic era. Nutr. Rev. 2015, 73, 523–543. [Google Scholar] [CrossRef] [PubMed]
- Nuno, N.B.; Heuberger, R. Nutrigenetic associations with cardiovascular disease. Rev. Cardiovasc. Med. 2014, 15, 217–225. [Google Scholar] [PubMed]
- de Goede, J.; Geleijnse, J.M.; Ding, E.L.; Soedamah-Muthu, S.S. Effect of cheese consumption on blood lipids: A systematic review and meta-analysis of randomized controlled trials. Nutr. Rev. 2015, 73, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Lordan, R.; Tsoupras, A.; Mitra, B.; Zabetakis, I. Dairy fats and cardiovascular disease: Do we really need to be concerned? Foods 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Otto, M.C.; Lemaitre, R.N.; Song, X.; King, I.B.; Siscovick, D.S.; Mozaffarian, D. Serial measures of circulating biomarkers of dairy fat and total and cause-specific mortality in older adults: The Cardiovascular Health Study. Am. J. Clin. Nutr. 2018, 108, 476–484. [Google Scholar] [CrossRef]
- De Toro-Martín, J.; Arsenault, B.; Després, J.-P.; Vohl, M.-C. Precision Nutrition: A Review of Personalized Nutritional Approaches for the Prevention and Management of Metabolic Syndrome. Nutrients 2017, 9, 913. [Google Scholar] [CrossRef] [PubMed]
- Ohlhorst, S.D.; Russell, R.; Bier, D.; Klurfeld, D.M.; Li, Z.; Mein, J.R.; Milner, J.; Ross, A.C.; Stover, P.; Konopka, E. Nutrition research to affect food and a healthy lifespan. Adv. Nutr. 2013, 4, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudkowska, I.; Dewailly, E.; Hegele, R.A.; Boiteau, V.; Dube-Linteau, A.; Abdous, B.; Giguere, Y.; Chateau-Degat, M.L.; Vohl, M.C. Gene-diet interactions on plasma lipid levels in the Inuit population. Br. J. Nutr. 2013, 109, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Jacobo-Albavera, L.; Posadas-Romero, C.; Vargas-Alarcón, G.; Romero-Hidalgo, S.; Posadas-Sánchez, R.; González-Salazar, M.D.C.; Carnevale, A.; Canizales-Quinteros, S.; Medina-Urrutia, A.; Antúnez-Argüelles, E.; et al. Dietary fat and carbohydrate modulate the effect of the ATP-binding cassette A1 (ABCA1) R230C variant on metabolic risk parameters in premenopausal women from the Genetics of Atherosclerotic Disease (GEA) Study. Nutr. Metab. 2015, 12, 45. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, J.A.; Steffen, L.M.; Ballantyne, C.M.; Boerwinkle, E.; Folsom, A.R. Associations between HDL-cholesterol and polymorphisms in hepatic lipase and lipoprotein lipase genes are modified by dietary fat intake in African American and White adults. Atherosclerosis 2007, 194, e131–e140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Feskens, E.J.; Dolle, M.E.; Imholz, S.; Verschuren, W.M.; Muller, M.; Boer, J.M. Dietary n-3 and n-6 polyunsaturated fatty acid intake interacts with FADS1 genetic variation to affect total and HDL-cholesterol concentrations in the Doetinchem Cohort Study. Am. J. Clin. Nutr. 2010, 92, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Dumont, J.; Huybrechts, I.; Spinneker, A.; Gottrand, F.; Grammatikaki, E.; Bevilacqua, N.; Vyncke, K.; Widhalm, K.; Kafatos, A.; Molnar, D.; et al. FADS1 genetic variability interacts with dietary alpha-linolenic acid intake to affect serum non-HDL-cholesterol concentrations in European adolescents. J. Nutr. 2011, 141, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Azorin, C.; Sorli, J.V.; Estruch, R.; Asensio, E.M.; Coltell, O.; Gonzalez, J.I.; Martinez-Gonzalez, M.A.; Ros, E.; Salas-Salvado, J.; Fito, M.; et al. Amino acid change in the carbohydrate response element binding protein is associated with lower triglycerides and myocardial infarction incidence depending on level of adherence to the Mediterranean diet in the PREDIMED trial. Circ. Cardiovasc. Genet. 2014, 7, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Volcik, K.A.; Nettleton, J.A.; Ballantyne, C.M.; Boerwinkle, E. Peroxisome proliferator–activated receptor α genetic variation interacts with n–6 and long-chain n–3 fatty acid intake to affect total cholesterol and LDL-cholesterol concentrations in the Atherosclerosis Risk in Communities Study. Am. J. Clin. Nutr. 2008, 87, 1926–1931. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Graphic representation of phenotypic effects of rare versus common variants. Rare variants, such as monogenic disorders, fall on the left of the graph. Common variants with a less severe phenotypic effects are on the right.
Figure 1.
Graphic representation of phenotypic effects of rare versus common variants. Rare variants, such as monogenic disorders, fall on the left of the graph. Common variants with a less severe phenotypic effects are on the right.
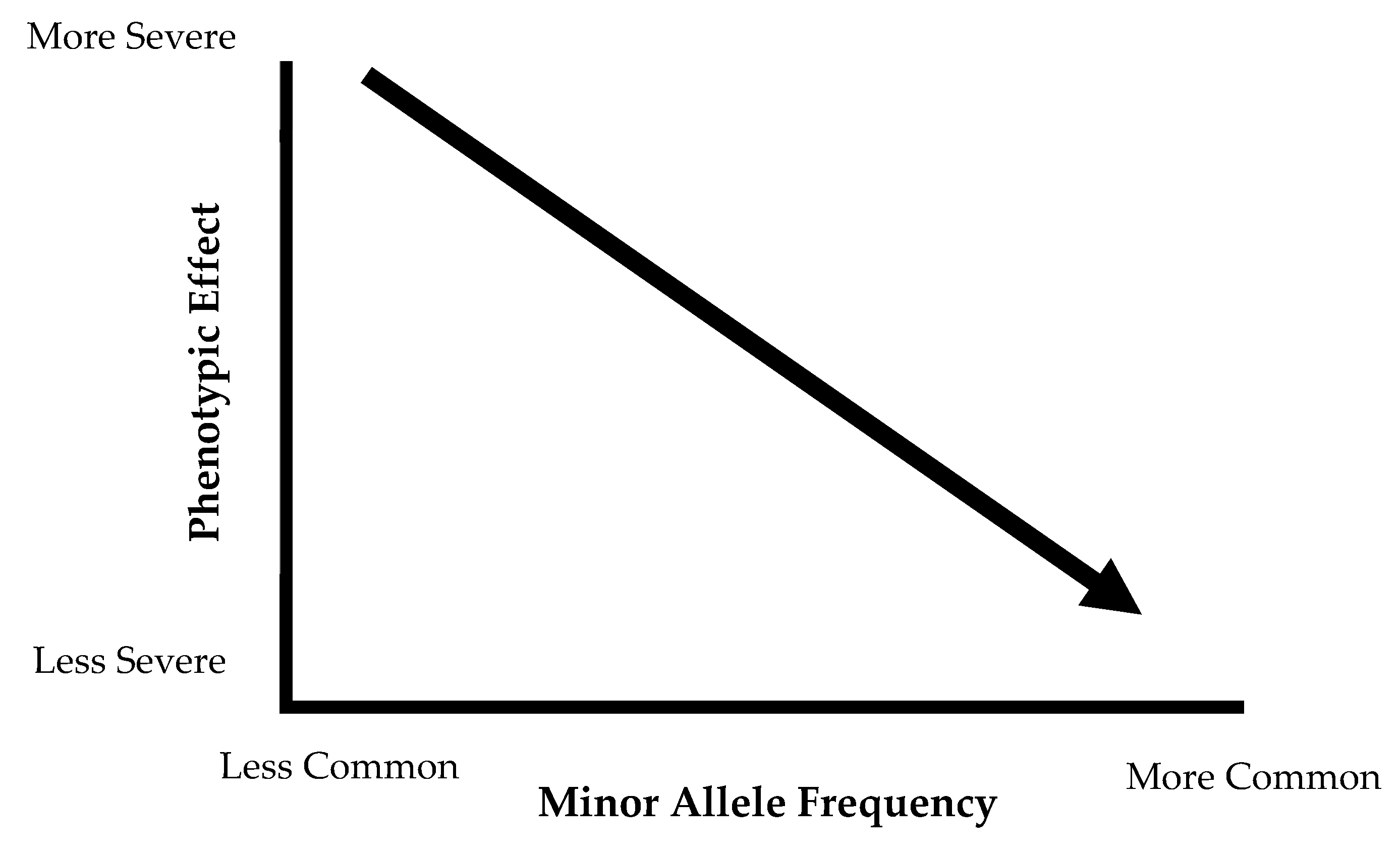
Figure 2.
Physiologically relevant genes of lipid and lipoprotein metabolism pathways. CM, chylomicron; HDL, high-density lipoprotein; IDL, intermediate density lipoprotein; LDL, low-density lipoprotein; VLDL, very-low density lipoprotein; ABCA1, ATP-binding cassette transported subfamily A member 1; ANGPTL3/4, angiopoietin-like proteins 3 & 4; APOA1, apolipoprotein AI; APOA2, apolipoprotein AII; APOAV, apolipoprotein AV; APOE, apolipoprotein E; CD36, cluster of differentiation 36 (fatty acid translocase); CETP, cholesterol esterase transfer protein; FADS, fatty acid desaturase cluster; MLXIPL, MLX interacting protein like; LCAT, lecithin, cholesterol acyltransferase; LDLR, LDL receptor; LIPC, hepatic lipase; LPL, lipoprotein lipase; PPARA, peroxisome-proliferator activator receptor alpha.
Figure 2.
Physiologically relevant genes of lipid and lipoprotein metabolism pathways. CM, chylomicron; HDL, high-density lipoprotein; IDL, intermediate density lipoprotein; LDL, low-density lipoprotein; VLDL, very-low density lipoprotein; ABCA1, ATP-binding cassette transported subfamily A member 1; ANGPTL3/4, angiopoietin-like proteins 3 & 4; APOA1, apolipoprotein AI; APOA2, apolipoprotein AII; APOAV, apolipoprotein AV; APOE, apolipoprotein E; CD36, cluster of differentiation 36 (fatty acid translocase); CETP, cholesterol esterase transfer protein; FADS, fatty acid desaturase cluster; MLXIPL, MLX interacting protein like; LCAT, lecithin, cholesterol acyltransferase; LDLR, LDL receptor; LIPC, hepatic lipase; LPL, lipoprotein lipase; PPARA, peroxisome-proliferator activator receptor alpha.

{kind=link}
{kind=link}
Table 1.
Monogenic disorders affect blood lipid concentrations (not an extensive list).
| Phenotype | Disorder | Gene Affected | Prevalence |
|---|---|---|---|
| High LDL | Hyperlipoproteinemia Type 2A | LDLR | 0.2% |
| Autosomal Dominant Hypercholesterolemia | PCSK9, APOE | 0.5% | |
| Low HDL | Tangier Disease | ABCA1 | <100 cases reported worldwide |
| Familial LCAT deficiency | LCAT | 70 reported cases | |
| High TG | Familial Chylomicronemia | LPL, APOC2 | <0.0001 |
| Severe Hypertriglyceridemia | APOA5, LMF1 | <0.5% |
Table adapted from Dron and Hegele [15]. LDL, low-density lipoprotein cholesterol; HDL, high-density lipoprotein cholesterol; TG, triglycerides; LDL-R, LDL receptor; PCSK9, proprotein convertase subtilisin/kexin type 9; APOE, apolipoprotein E; ABCA1, adenosine triphosphate (ATP) binding cassette subfamily A member 1; LCAT, lecithin-cholesterol acyltransferase; LPL, lipoprotein lipase; APOC2, apolipoprotein C2; APOA5, apolipoprotein A5; LMF1, lipase maturation factor 1.
Table 2.
Summary of gene-diet interactions between physiologically relevant variables of lipid and lipoprotein metabolism and dietary fat intake associated with blood lipids.
Table 2.
Summary of gene-diet interactions between physiologically relevant variables of lipid and lipoprotein metabolism and dietary fat intake associated with blood lipids.
| Gene | Locus | Protein Function | Previous Nutrient-Gene Interaction with Blood Lipids | SNP | Function of Variant | Risk Allele | MAF Global |
|---|---|---|---|---|---|---|---|
| Reverse Cholesterol Transport Pathway | |||||||
| CETP | 16q13 | Facilitates the exchange of cholesterol esters for TG between lipoproteins in circulation | Total fat and TG [98]; total fat and TG [108] | rs5882 | Missense variant | G | 0.37 |
| ABCA1 | 9q31.1 | HDL-C bound protein that transports intracellular cholesterol onto HDL-C | Total fat and HDL [109] | rs9282541 | Missense variant | T | 0.01 |
| SFA and TG [108] | rs2230806 | Missense variant | T | 0.32 | |||
| LIPC | 15q21.3 | Hepatic triglyceride lipase, also involved in lipoprotein uptake | SFA and HDL, TG [98]; total fat and HDL [110] | rs1800588 | Intron variant in promotor region, associated with lowered LIPC activity | T | 0.29 |
| APOA1 | 11q23.3 | Predominant apolipoprotein on HDL; activator of LCAT | SFA, total fat, and TC [108] | rs670 | Upstream intronic variant | T | 0.18 |
| Total fat and HDL [108] | rs5070 | Intron variant | G | 0.44 | |||
| Cellular Lipid Uptake Pathway | |||||||
| APOE | 19q13.32 | Present on TG-rich lipoproteins (chylomicrons, VLDL) | Total fat, SFA, and HDL [98] | rs405509 | Upstream variant in promoter region | T | 0.47 |
| CD36 | 7q21.11 | Scavenger receptor, binds to oxidized LDL and LCFA. | Oily fish (n-3 PUFA) and HDL [40] | rs6969989 | Intron variant | G | 0.33 |
| LPL | 8p21.3 | Hydrolyzes TG to allow fatty acids from lipoproteins into circulation | Total fat and HDL [33,110] | rs328 | Nonsense variant | G | 0.10 |
| Lipid/Lipoprotein Formation Pathway | |||||||
| APOA5 | 11q23.3 | Present on HDL particles, stimulates LPL, major determinant of plasma TG concentrations | Total fat and TC, LDL, HDL [96]; | rs964184 | 3’ untranslated region (UTR) variant | G | 0.22 |
| Total fat and TG [90] | rs662799 | Upstream variant in promoter region | G | 0.16 | |||
| FADS Complex | 11q12-13.1 | Desaturation of long-chain fatty acids | n-3, n-6 PUFAs and HDL [111]; alpha-linolenic acid and non-HDL cholesterol [112] | rs174546 | 3’ UTR variant | T | 0.28 |
| MLXIPL | 7q11.23 | Activates carbohydrate-responsive element binding protein and promotes hepatic TG synthesis | Mediterranean diet and TG [113] | rs3812316 | Missense variant | G | 0.11 |
| PPARA | 22q13.31 | Nuclear receptor in liver, ligand for PUFAs | n-3 PUFA and TC, LDL | rs6008259 | Non-coding transcript variant | A | 0.32 |
| n-6 PUFA and TC, LDL [114] | rs3892755 | Non-coding transcript variant | A | 0.09 | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hannon, B.A.; Khan, N.A.; Teran-Garcia, M. Nutrigenetic Contributions to Dyslipidemia: A Focus on Physiologically Relevant Pathways of Lipid and Lipoprotein Metabolism. Nutrients 2018, 10, 1404. https://doi.org/10.3390/nu10101404
AMA Style
Hannon BA, Khan NA, Teran-Garcia M. Nutrigenetic Contributions to Dyslipidemia: A Focus on Physiologically Relevant Pathways of Lipid and Lipoprotein Metabolism. Nutrients. 2018; 10(10):1404. https://doi.org/10.3390/nu10101404
Chicago/Turabian StyleHannon, Bridget A., Naiman A. Khan, and Margarita Teran-Garcia. 2018. "Nutrigenetic Contributions to Dyslipidemia: A Focus on Physiologically Relevant Pathways of Lipid and Lipoprotein Metabolism" Nutrients 10, no. 10: 1404. https://doi.org/10.3390/nu10101404
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.