
A Bxtlp Gene Affects the Pathogenicity of Bursaphelenchus xylophilus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Cloning of Bxtlp and Construction of Expression Vector
2.3. Bioinformatics Analysis of Bxtlp and BxTLP
2.4. Expression and Purification of Recombinant BxTLP
2.5. Synthesis of Bxtlp dsRNA
2.6. Effects of Bxtlp dsRNA on the Vitality of PWNs
2.7. Effects of Bxtlp dsRNA on ROS Levels, Lifespan, and Female–Male Ratio in PWNs
2.8. Effect of Recombinant BxTLP on PWN
2.9. Determination of Antioxidant Activity of Recombinant BxTLP
2.10. The Effect of dsRNA and Recombinant BxTLP on the Pathogenicity of PWN
2.11. Data Analysis
3. Results
3.1. Cloning and Sequence Analysis of Bxtlp in PWN
3.2. Expression and Purification of Recombinant BxTLP Protein
3.3. Determination of Antioxidant Activity of Recombinant BxTLP
3.4. Bxtlp dsRNA Can Effect PWN Vitality
3.5. Effects of Bxtlp dsRNA on ROS Content, Lifespan, and Female–Male Ratio of PWN
3.6. Effect of Recombinant BxTLP on PWN
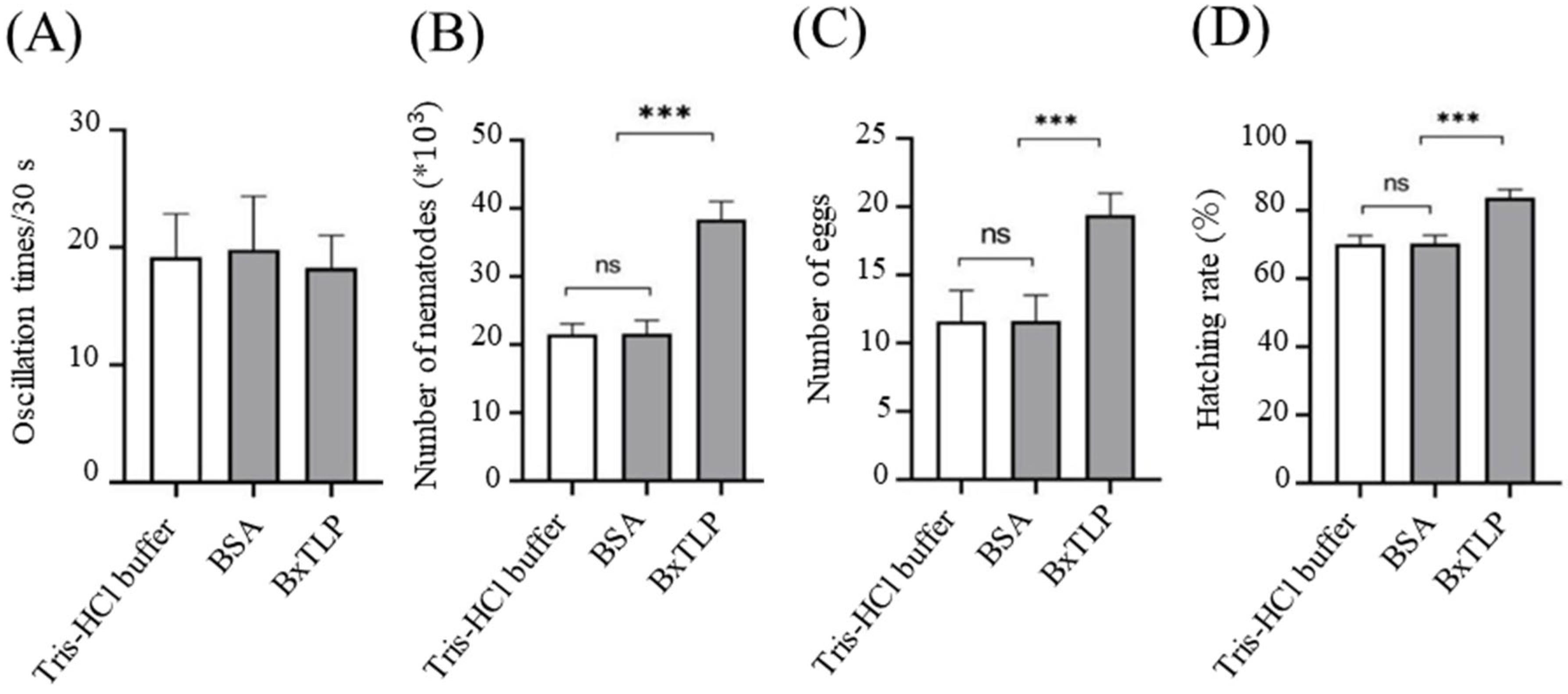
3.7. Bxtlp Affects the Pathogenicity of PWN
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rodrigues, J.M. Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Wang, L.S.; Zhang, T.T.; Pan, Z.S.; Lin, L.L.; Dong, G.Q.; Wang, M.; Li, R.G. The alcohol dehydrogenase with a broad range of substrate specificity regulates vitality and reproduction of the plant-parasitic nematode Bursaphelenchus xylophilus. Parasitology 2019, 146, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Suzuki, T.; Kawazu, K. Development and preventative effect against pine wilt disease of a novel liquid formulation of emamectin benzoate. Pest Manag. Sci. 2003, 59, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Lee, H.R.; Kim, D.S.; Kwon, J.H.; Huh, M.J.; Park, I.K. Emamectin benzoate 9.7% SL as a new formulation for a trunk-injections against pine wood nematode, Bursaphelenchus xylophilus. J. For. Res. 2020, 31, 1399–1403. [Google Scholar] [CrossRef]
- Gnanendra, S.; Sun, L.; Junhyun, J. Identification of Potential Nematicidal Compounds Against the Pine Wood Nematode, Bursaphelenchus xylophilus Through an In Silico Approach. Molecules 2018, 23, 1828. [Google Scholar] [CrossRef]
- Liu, M.I.; Hwang, B.O.; Jin, C.H.; Li, W.U.; Park, D.I.; Seo, S.A.; Kim, C.I. Screening, isolation and evaluation of a nematicidal compound from actinomycetes against the pine wood nematode. Pest Manag. Sci. 2019, 75, 1585–1593. [Google Scholar] [CrossRef]
- Liu, J.J.; Sturrock, R.; Ekramoddoullah, A.K.M. The superfamily of thaumatin-like proteins: Its origin, evolution, and expression towards biological function. Plant Cell Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef]
- Ho, V.S.M.; Wong, J.H.; Ng, T.B. A thaumatin-like antifungal protein from the emperor banana. Peptides 2007, 28, 760–766. [Google Scholar] [CrossRef]
- Wel, H.; Loeve, K. Isolation and characterization of thaumatin I and II, the sweet-tasting proteins from Thaumatococcus daniellii Benth. Eur. J. Biochem. 2010, 31, 221–225. [Google Scholar]
- Zhang, M.; Xu, J.H.; Liu, G.; Yang, X.P. Antifungal properties of a thaumatin-like protein from watermelon. Acta Physiol. Plant. 2018, 40, 186. [Google Scholar] [CrossRef]
- Garcia-Casado, C.; Collada, C.; Allona, I.; Soto, A.; Casado, R.; Rodriguez-Cerezo, E.; Gomez, L.; Aragoncillo, C. Characterization of an apoplastic basic thaumatin-like protein from recalcitrant chestnut seeds. Physiol. Plant. 2000, 110, 172–180. [Google Scholar] [CrossRef]
- Chu, K.T.; Ng, T.B. Isolation of a large thaumatin-like antifungal protein from seeds of the Kweilin chestnut Castanopsis chinensis. Biochem. Biophys. Res. Commun. 2003, 301, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Jayasankar, S.; Li, Z.J.; Gray, D.J. Constitutive expression of Vitis vinifera thaumatin-like protein after in vitro selection and its role in anthracnose resistance. Funct. Plant Biol. 2003, 30, 1105–1115. [Google Scholar] [CrossRef]
- Fierens, E.; Rombouts, S.; Gebruers, K.; Goesaert, H.; Brijs, K.; Beaugrand, J.; Volckaert, G.; Van Campenhout, S.; Proost, P.; Courtin, C.M.; et al. TLXI, a novel type of xylanase inhibitor from wheat (Triticum aestivum) belonging to the thaumatin family. Biochem. J. 2007, 403, 583–591. [Google Scholar] [CrossRef]
- Bormann, C.; Baier, D.; Hörr, I.; Raps, C.; Berger, J.; Jung, G.; Schwarz, H. Characterization of a novel, antifungal, chitin-binding protein from Streptomyces tendae Tu901 that interferes with growth polarity. J. Bacteriol. 1999, 181, 7421–7429. [Google Scholar] [CrossRef]
- Osmond, R.I.W.; Hrmova, M.; Fontaine, F.; Imberty, A.; Fincher, G.B. Binding interactions between barley thaumatin-like proteins and (1,3)-β-D-glucans: Kinetics, specificity, structural analysis and biological implications. Eur. J. Biochem. 2001, 268, 4190–4199. [Google Scholar] [CrossRef]
- Yu, X.M.; Griffith, M. Antifreeze proteins in winter rye leaves form oligomeric complexes. Plant Physiol. 1999, 119, 1361–1370. [Google Scholar] [CrossRef]
- Parkhi, V.; Kumar, V.; Sunilkumar, G.; Campbell, L.M.; Singh, N.K.; Rathore, K.S. Expression of apoplastically secreted tobacco osmotin in cotton confers drought tolerance. Mol. Breed. 2009, 23, 625–639. [Google Scholar] [CrossRef]
- Husaini, A.M.; Abdin, M.Z. Development of transgenic strawberry (Fragaria × ananassa Duch.) plants tolerant to salt stress. Plant Sci. 2008, 174, 446–455. [Google Scholar] [CrossRef]
- Kitajima, S.; Sato, F. Plant pathogenesis-related proteins: Molecular mechanisms of gene expression and protein function. J. Biochem. 1999, 125, 1–8. [Google Scholar] [CrossRef]
- Brandazza, A.; Angeli, S.; Tegoni, M.; Cambillau, C.; Pelosi, P. Plant stress proteins of the thaumatin-like family discovered in animals. Febs Lett. 2004, 572, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.Q.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Chi-Ham, C.L.; Clark, K.L.; Bennett, A.B. The intellectual property landscape for gene suppression technologies in plants. Nat. Biotechnol. 2010, 28, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.H.; Jian, H.; Xu, J.M.; Guo, Y.D.; Liu, Q.A. RNAi technology extends its reach: Engineering plant resistance against harmful eukaryotes. Afr. J. Biotechnol. 2010, 9, 7573–7582. [Google Scholar]
- Wang, M.; Wang, D.D.; Zhang, X.; Wang, X.; Liu, W.C.; Hou, X.M.; Huang, X.Y.; Xie, B.Y.; Cheng, X.Y. Double-stranded RNA-mediated interference of dumpy genes in Bursaphelenchus xylophilus by feeding on filamentous fungal transformants. Int. J. Parasitol. 2016, 46, 351–360. [Google Scholar] [CrossRef]
- Cheng, X.Y.; Dai, S.M.; Xiao, L.; Xie, B.Y. Influence of cellulase gene knockdown by dsRNA interference on the development and reproduction of the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2010, 12, 225–233. [Google Scholar] [CrossRef]
- Zhao, Q.; Hu, L.J.; Wu, X.Q.; Wang, Y.C. A key effector, BxSapB2, plays a role in the pathogenicity of the pine wood nematode Bursaphelenchus xylophilus. For. Pathol. 2020, 50, e12600. [Google Scholar] [CrossRef]
- Viglierchio, D.R.; Schmitt, R.V. On the methodology of nematode extraction from field samples: Baermann funnel modifications. J. Nematol. 1983, 15, 438–444. [Google Scholar]
- Guo, Q.Q.; Du, G.C.; Qi, H.T.; Zhang, Y.A.; Yue, T.Q.; Wang, J.C.; Li, R.G. A nematicidal tannin from Punica granatum L. rind and its physiological effect on pine wood nematode (Bursaphelenchus xylophilus). Pestic. Biochem. Physiol. 2017, 135, 64–68. [Google Scholar] [CrossRef]
- Liu, B.-J.; Hu, J.-F.; Liu, Z.-Y.; Xu, L.; Lu, Q.; Li, Y.-X.; Zhang, X.-Y. Behavioural features of Bursaphelenchus xylophilus in the matingprocess. Nematology 2014, 16, 895–902. [Google Scholar] [CrossRef]
- Zhu, N.J.; Bai, L.Q.; Schütz, S.; Liu, B.J.; Liu, Z.Y.; Zhang, X.Y.; Yu, H.S.; Hu, J.F. Observation and Quantification of Mating Behavior in the Pinewood Nematode, Bursaphelenchus xylophilus. JoVE-J. Vis. Exp. 2016, 118, e54842. [Google Scholar]
- Tang, J.; Ma, R.Q.; Zhu, N.J.; Guo, K.; Guo, Y.Q.; Bai, L.Q.; Yu, H.S.; Hu, J.F.; Zhang, X.Y. Bxy-fuca encoding α-L-fucosidase plays crucial roles in development and reproduction of the pathogenic pinewood nematode, Bursaphelenchus xylophilus. Pest Manag. Sci. 2020, 76, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Shinya, R.; Takeuchi, Y.; Miura, N.; Kuroda, K.; Ueda, M.; Futai, K. Surface coat proteins of the pine wood nematode, Bursaphelenchus xylophilus: Profiles of stage- and isolate-specific characters. Nematology 2009, 11, 429–438. [Google Scholar]
- Wang, M.; Du, G.C.; Fang, J.N.; Wang, L.S.; Guo, Q.Q.; Zhang, T.T.; Li, R.G. UGT440A1 Is Associated With Motility, Reproduction, and Pathogenicity of the Plant-Parasitic Nematode Bursaphelenchus xylophilus. Front. Plant Sci. 2022, 13, 862594. [Google Scholar] [CrossRef]
- Zhang, X.L.; Huang, X.L.; Li, J.; Mei, M.; Zeng, W.Q.; Lu, X.J. Evaluation of the RNA extraction methods in different Ginkgo biloba L. tissues. Biologia 2021, 76, 2393–2402. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, S.N.; Lu, F.; Guo, K.; Huang, L.L.; Su, X.; Chen, Y. Nematotoxicity of a Cyt-like protein toxin from Conidiobolus obscurus (Entomophthoromycotina) on the pine wood nematode Bursaphelenchus xylophilus. Pest Manag. Sci. 2021, 77, 686–692. [Google Scholar] [CrossRef]
- Xu, X.L.; Wu, X.Q.; Ye, J.R.; Huang, L. Molecular Characterization and Functional Analysis of Three Pathogenesis-Related Cytochrome P450 Genes from Bursaphelenchus xylophilus (Tylenchida: Aphelenchoidoidea). Int. J. Mol. Sci. 2015, 16, 5216–5234. [Google Scholar] [CrossRef]
- Huang, L.; Wang, P.; Tian, M.Q.; Zhu, L.H.; Ye, J.R. Major sperm protein BxMSP10 is required for reproduction and egg hatching in Bursaphelenchus xylophilus. Exp. Parasitol. 2019, 197, 51–56. [Google Scholar] [CrossRef]
- Park, J.E.; Lee, K.Y.; Lee, S.J.; Oh, W.S.; Jeong, P.Y.; Woo, T.; Kim, C.B.; Paik, Y.K.; Koo, H.S. The efficiency of RNA interference in Bursaphelenchus xylophilus. Mol. Cells 2008, 26, 81–86. [Google Scholar] [CrossRef]
- Li, X.C.; Gao, Y.X.; Li, F.; Liang, A.F.; Xu, Z.M.; Bai, Y.; Mai, W.Q.; Han, L.; Chen, D.F. Maclurin protects against hydroxyl radical-induced damages to mesenchymal stem cells: Antioxidant evaluation and mechanistic insight. Chem.-Biol. Interact. 2014, 219, 221–228. [Google Scholar] [CrossRef]
- Li, X.J.; Cui, S.Y. DPPH Radical Scavenging Mechanism of Ascorbic Acid. Food Sci. 2011, 32, 86–90. [Google Scholar]
- Qiu, X.W.; Wu, X.Q.; Huang, L.; Ye, J.R. Influence of Bxpel1 Gene Silencing by dsRNA Interference on the Development and Pathogenicity of the Pine Wood Nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2016, 17, 125. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C. Comparative Study of 1,1-Diphenyl-2-picryl-hydrazyl Radical (DPPH•) Scavenging Capacity of the Antioxidant Xanthones Family. Chemistryselect 2018, 3, 13081–13086. [Google Scholar] [CrossRef]
- Wang, B.W.; Hao, X.; Xu, J.Y.; Wang, B.Y.; Ma, W.; Liu, X.F.; Ma, L. Cytochrome P450 metabolism mediates low-temperature resistance in pinewood nematode. FEBS Open Bio 2020, 10, 1171–1179. [Google Scholar] [CrossRef]
- Liu, S.S.; Wang, L.S.; Li, R.G.; Chen, M.Y.; Deng, W.J.; Wang, C.; Du, G.C.; Guo, Q.Q. Cloning of sft-4 and its influence on vitality and virulence of pine wood nematode, Bursaphelenchus xylophilus. J. For. Res. 2024, 35, 43. [Google Scholar] [CrossRef]
- Kuwabara, C.; Takezawa, D.; Shimada, T.; Hamada, T.; Fujikawa, S.; Arakawa, K. Abscisic acid- and cold-induced thaumatin-like protein in winter wheat has an antifungal activity against snow mould, Microdochium nivale. Physiol. Plant. 2002, 115, 101–110. [Google Scholar] [CrossRef]
- Kehrer, J.P. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Liochev, S.I.; Fridovich, I. The Haber-Weiss cycle—70 years later: An alternative view. Redox Rep. 2002, 7, 55–57. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Guo, Q.; Huang, Z.; Feng, W.; Zhang, Y.; Zhao, W.; Li, R.; Du, G. A Bxtlp Gene Affects the Pathogenicity of Bursaphelenchus xylophilus. Forests 2025, 16, 1122. https://doi.org/10.3390/f16071122
Liu S, Guo Q, Huang Z, Feng W, Zhang Y, Zhao W, Li R, Du G. A Bxtlp Gene Affects the Pathogenicity of Bursaphelenchus xylophilus. Forests. 2025; 16(7):1122. https://doi.org/10.3390/f16071122
Chicago/Turabian StyleLiu, Shuisong, Qunqun Guo, Ziyun Huang, Wentao Feng, Yingying Zhang, Wenying Zhao, Ronggui Li, and Guicai Du. 2025. "A Bxtlp Gene Affects the Pathogenicity of Bursaphelenchus xylophilus" Forests 16, no. 7: 1122. https://doi.org/10.3390/f16071122
APA StyleLiu, S., Guo, Q., Huang, Z., Feng, W., Zhang, Y., Zhao, W., Li, R., & Du, G. (2025). A Bxtlp Gene Affects the Pathogenicity of Bursaphelenchus xylophilus. Forests, 16(7), 1122. https://doi.org/10.3390/f16071122