
Secondary Metabolites and Biosynthetic Gene Clusters Analysis of Deep-Sea Hydrothermal Vent-Derived Streptomyces sp. SCSIO ZS0520

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
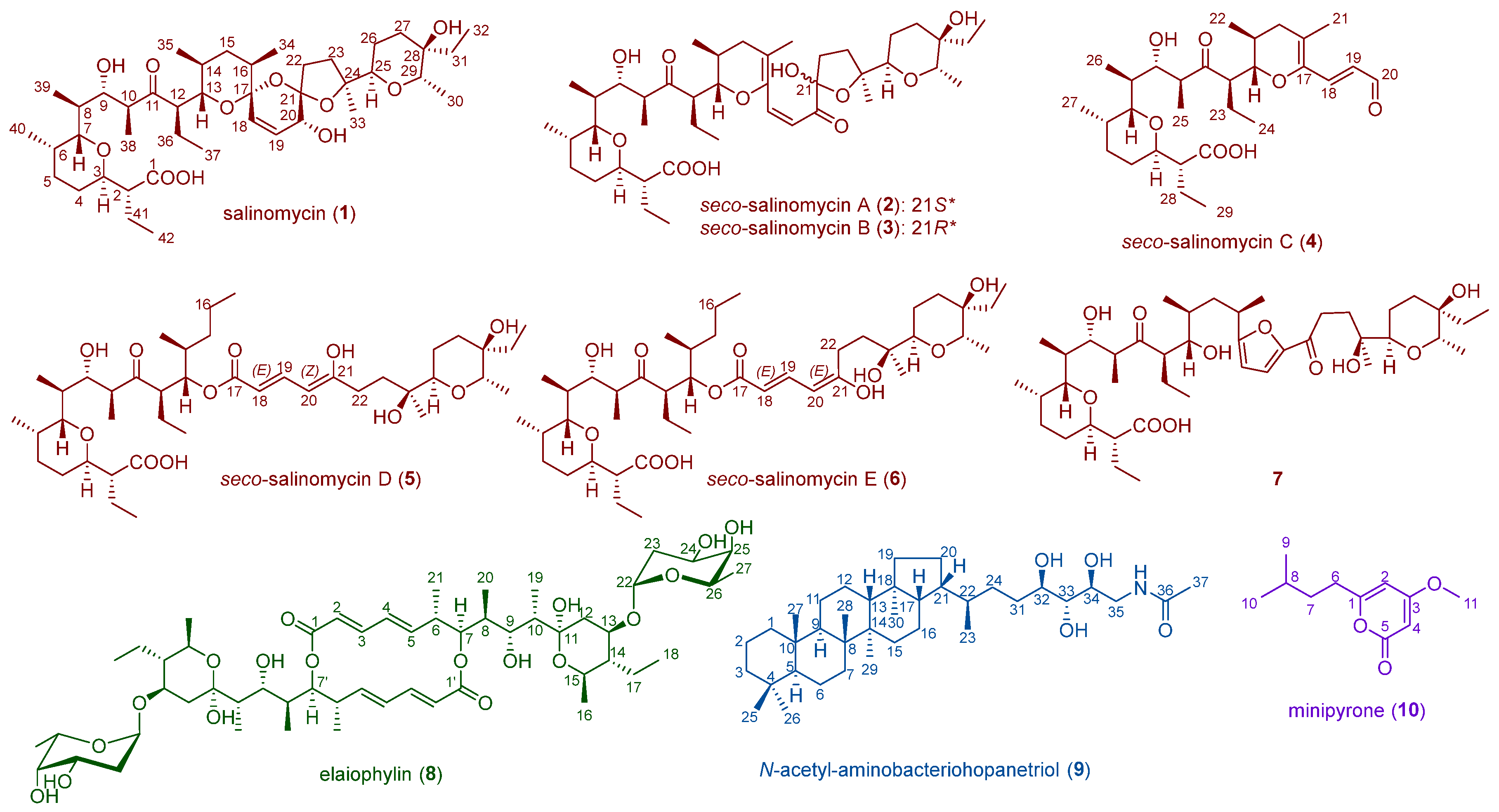
2.1. Metabolite Investigation of the Strain Streptomyces sp. SCSIO ZS0520
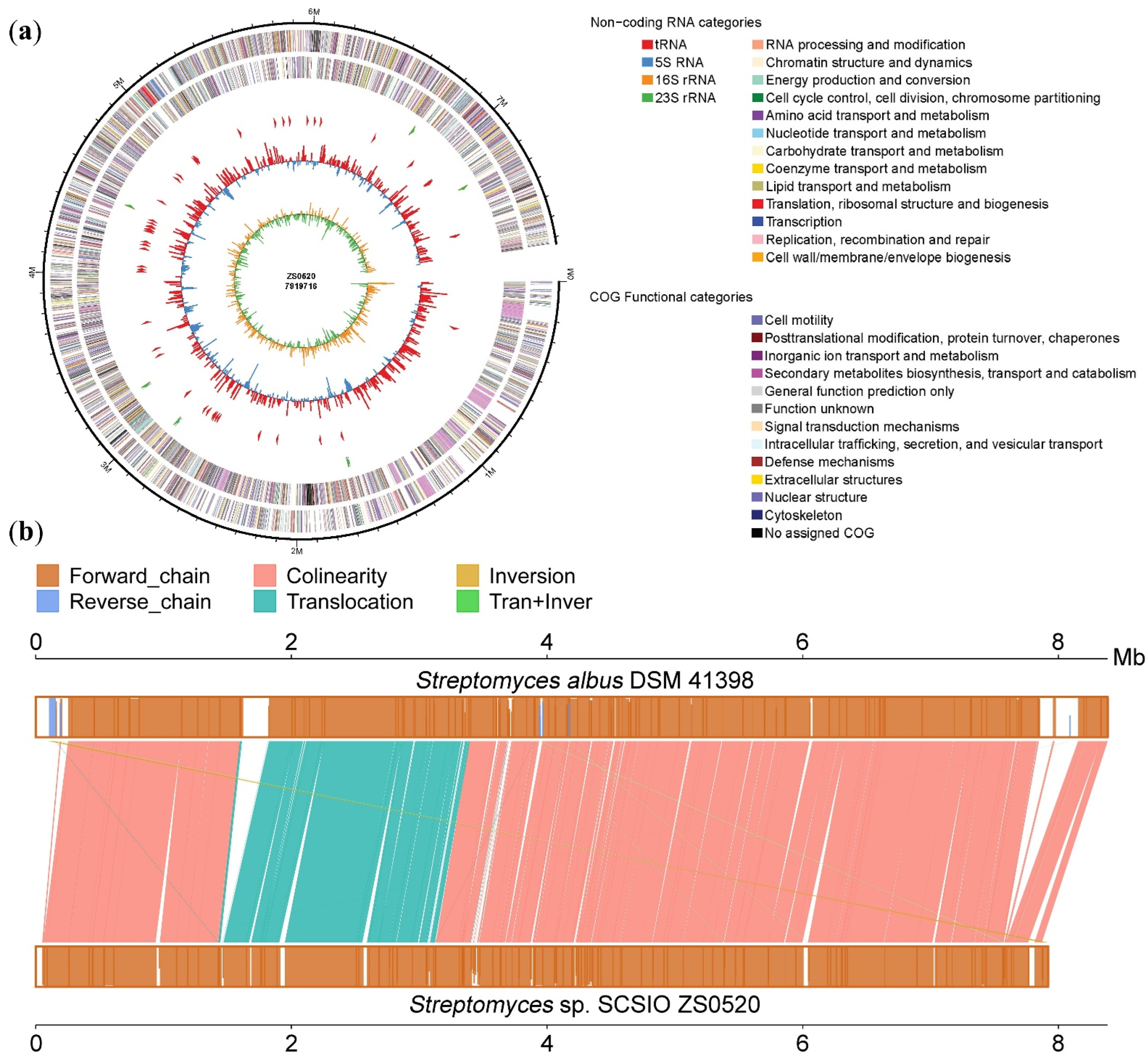
2.2. Genome Features and Annotations of Strain Streptomyces sp. SCSIO ZS0520
2.3. Biosynthesis and Isomeric Mechanism of Salinomycin
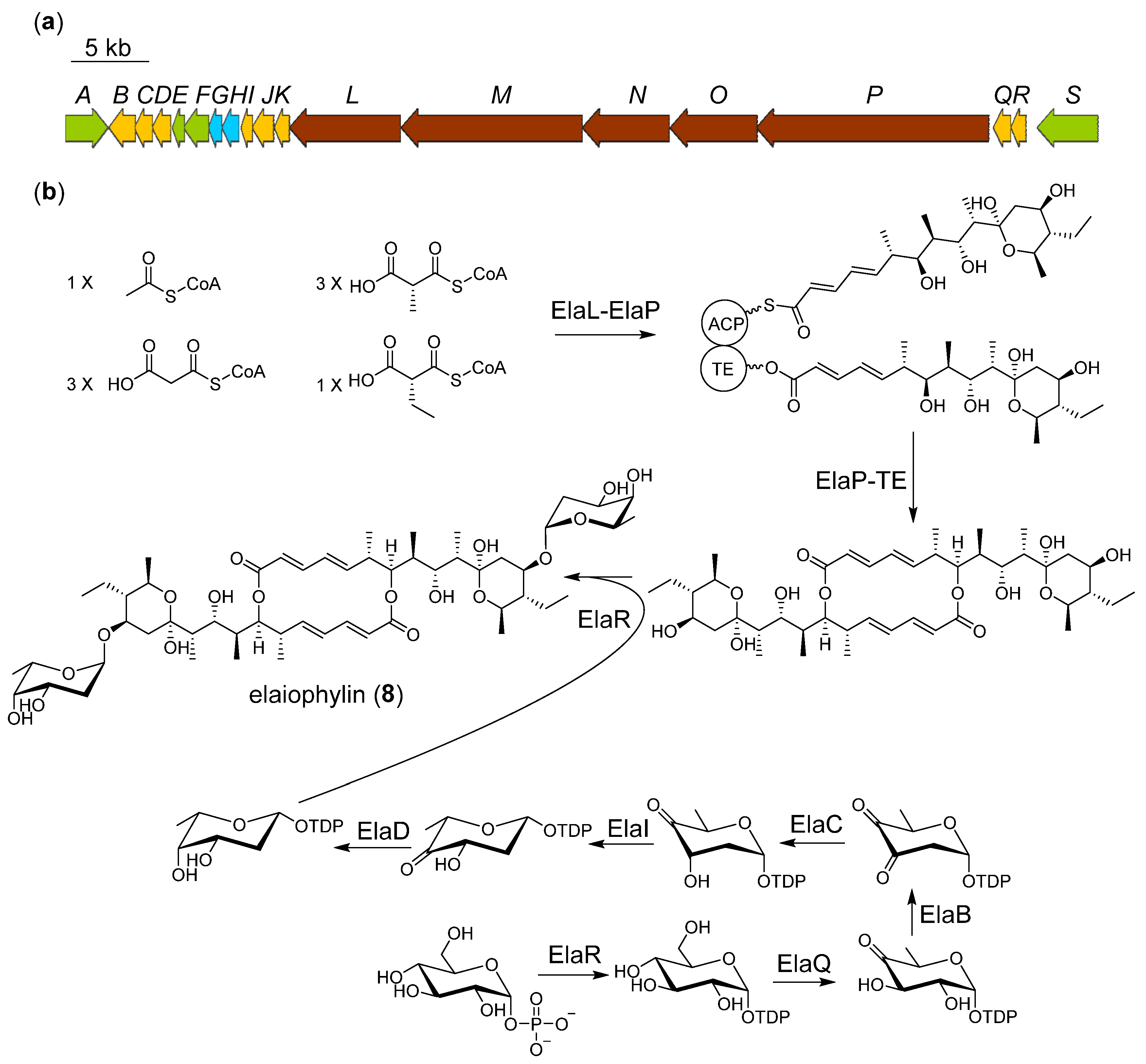
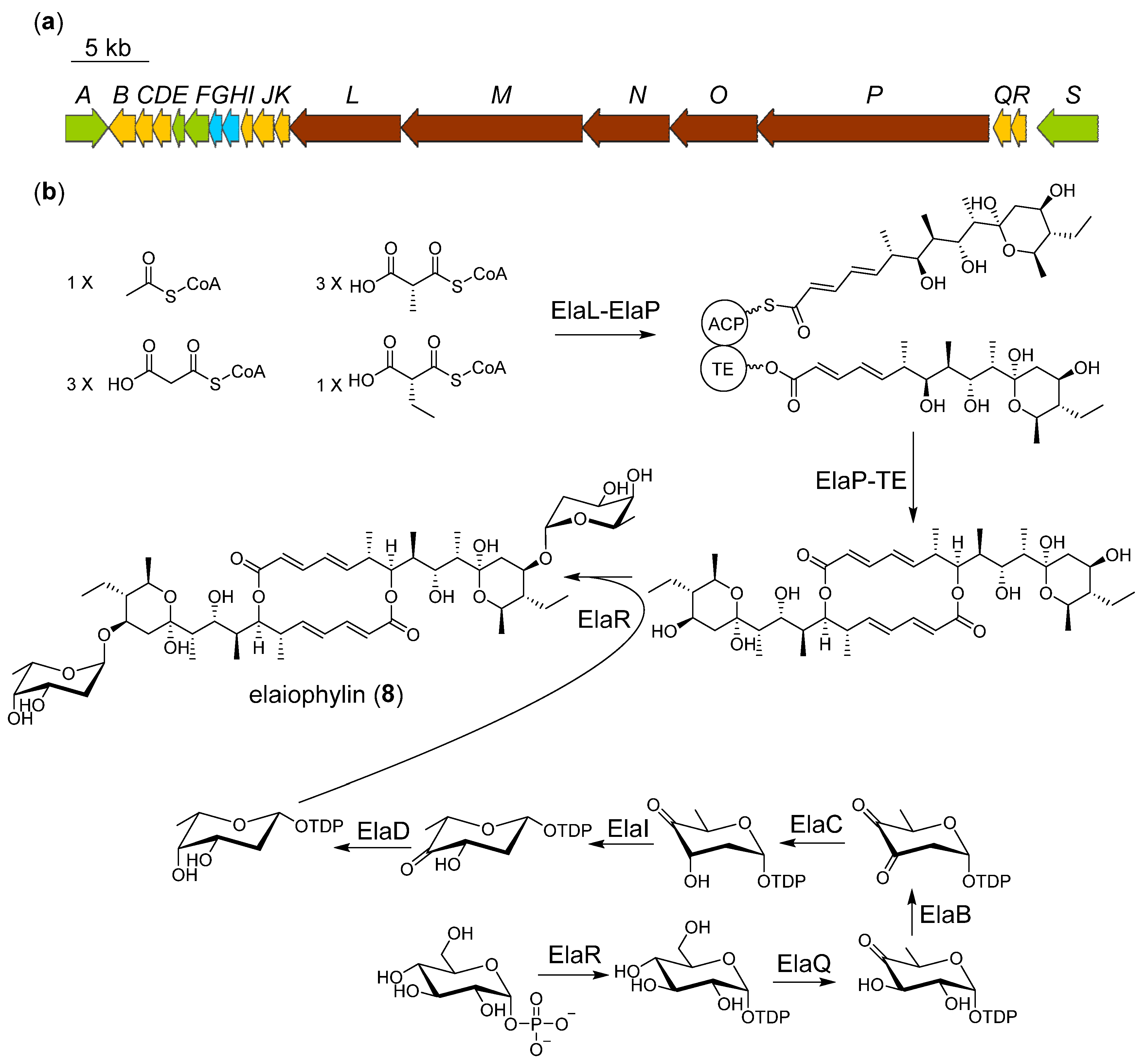
2.4. Proposed Biosynthetic Pathway of Elaiophilin
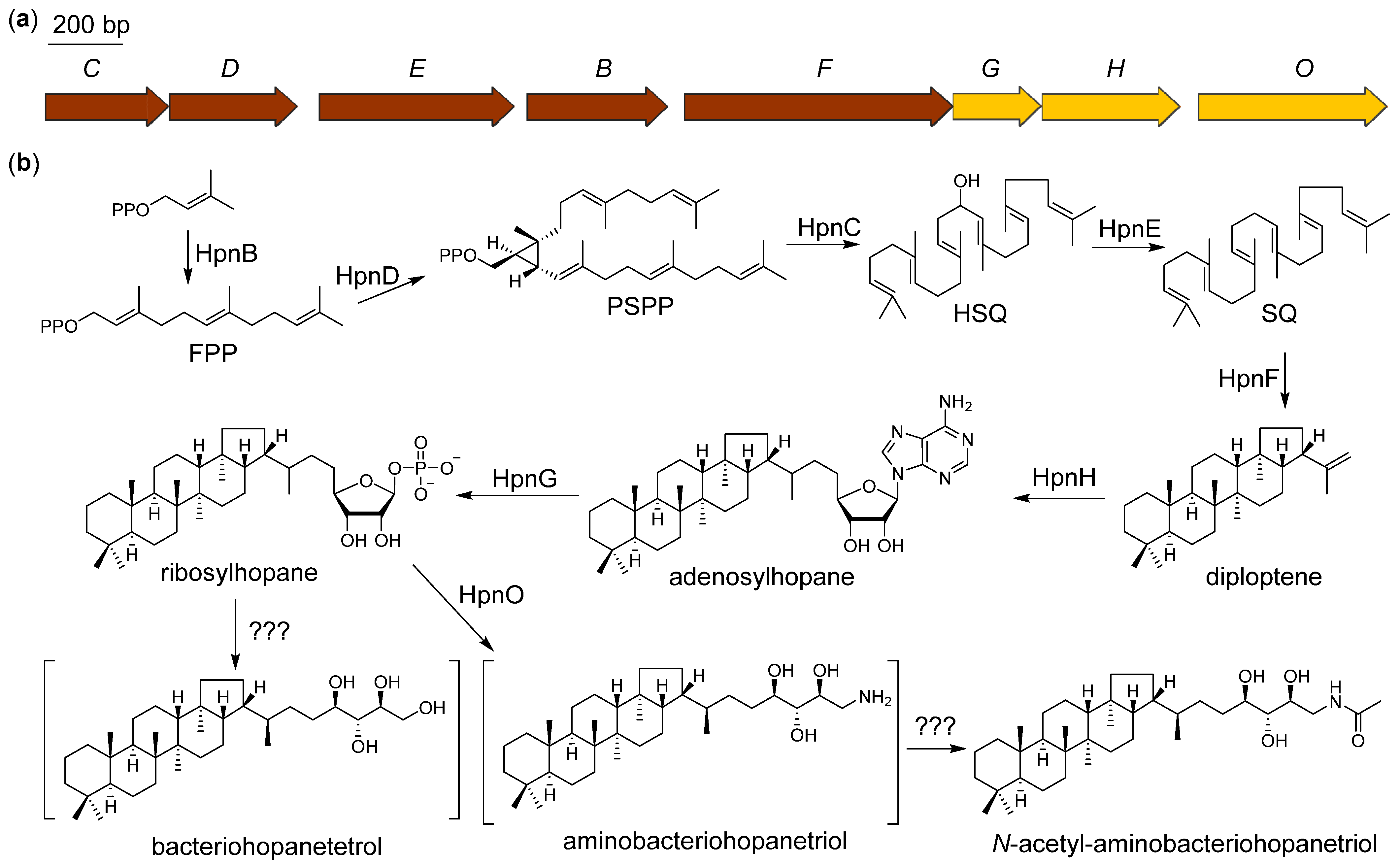
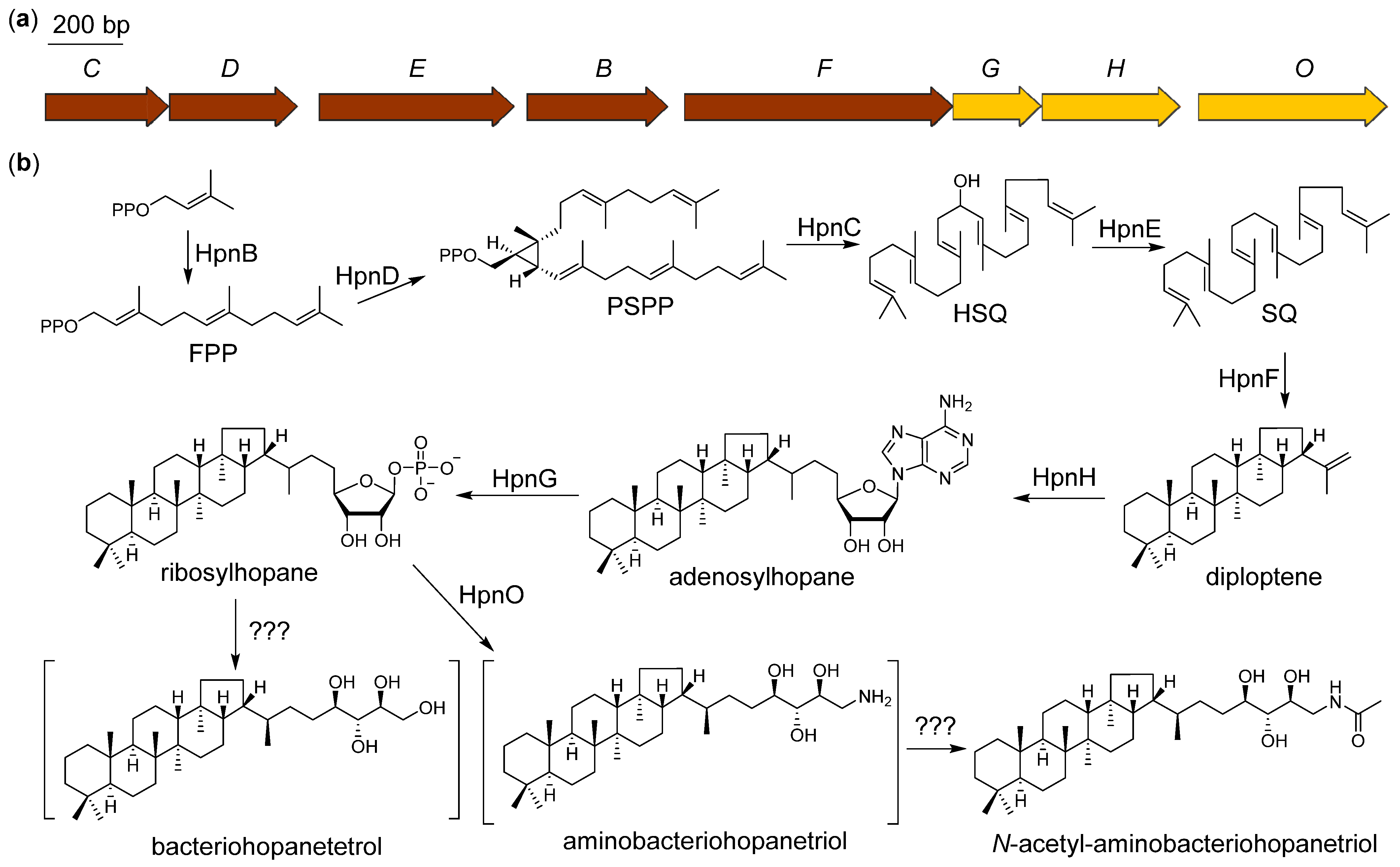
2.5. Proposed Biosynthetic Pathway of N-Acetyl-aminobacteriohopanetriol
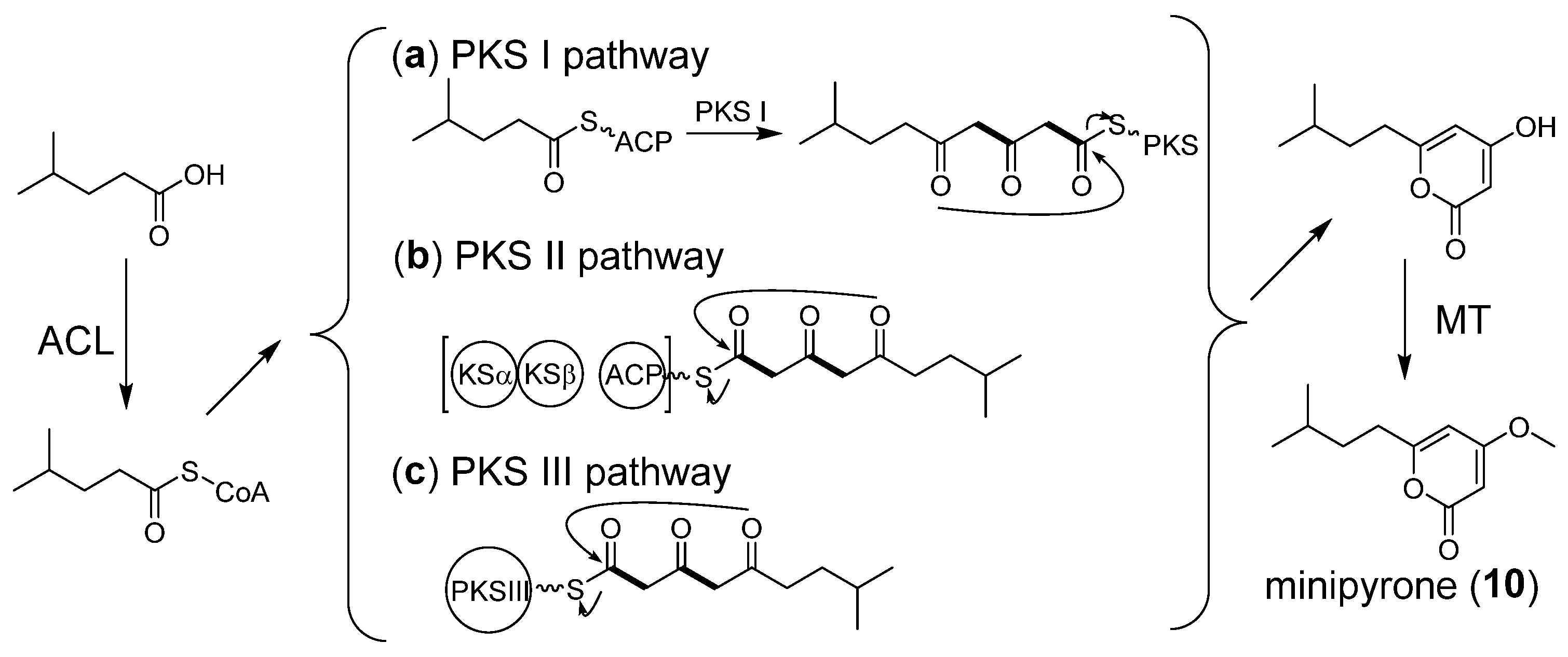
2.6. Proposed Biosynthetic Pathway of Minipyrone
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Extraction and Purification
3.3. Characterization of Compounds
3.4. Genome Sequencing and Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thornburg, C.C.; Zabriskie, T.M.; McPhail, K.L. Deep-sea hydrothermal vents: Potential hot spots for natural products discovery? J. Nat. Prod. 2010, 73, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, X.; Huang, Y.; Yuan, J.; Wei, X.; Ju, J. Discovery, structure correction, and biosynthesis of actinopyrones, cytotoxic polyketides from the deep-sea hydrothermal-vent-derived Streptomyces sp. SCSIO ZS0520. J. Nat. Prod. 2021, 85, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Shibuya, M.; Sugawara, H.; Kawaguchi, O.; Hirsoe, C. Salinomycin, a new polyether antibiotic. J. Antibiot. 1974, 27, 814–821. [Google Scholar] [CrossRef]
- Antoszczak, M. A comprehensive review of salinomycin derivatives as potent anticancer and anti-CSCs agents. Eur. J. Med. Chem. 2019, 166, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Riddell, F.G. Structure, conformation, and mechanism in the membrane transport of alkali metal ions by ionophoric antibiotics. Chirality 2002, 14, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Antoszczak, M.; Steverding, D.; Huczyński, A. Anti-parasitic activity of polyether ionophores. Eur. J. Med. Chem. 2019, 166, 32–47. [Google Scholar] [CrossRef] [Green Version]
- Sulik, M.; Stępień, K.; Stefańska, J.; Huczyński, A.; Antoszczak, M. Antibacterial activity of singly and doubly modified salinomycin derivatives. Bioorg. Med. Chem. Lett. 2020, 30, 127062. [Google Scholar] [CrossRef]
- Antoszczak, M.; Huczyński, A. Salinomycin and its derivatives—A new class of multiple-targeted “magic bullets”. Eur. J. Med. Chem. 2019, 176, 208–227. [Google Scholar] [CrossRef]
- Antoszczak, M. A medicinal chemistry perspective on salinomycin as a potent anticancer and anti-CSCs agent. Eur. J. Med. Chem. 2019, 164, 366–377. [Google Scholar] [CrossRef]
- Davis, A.L.; Harris, J.A.; Russell, C.A.L.; Wilkins, J.P.G. Investigations by HPLC-electrospray mass spectrometry and NMR spectroscopy into the isomerisation of salinomycin. Analyst 1999, 124, 251–256. [Google Scholar] [CrossRef]
- Wells, J.L.; Bordner, J.; Bowles, P.; McFarland, J.W. Novel degradation products from the treatment of salinomycin and narasin with formic acid. J. Med. Chem. 1988, 31, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Schlusener, M.P.; von Arb, M.A.; Bester, K. Elimination of macrolides, tiamulin, and salinomycin during manure storage. Arch. Environ. Contam. Toxicol. 2006, 51, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Pankiewicz, R. Spectroscopic and computational study of a new isomer of salinomycin. J. Mol. Struct. 2013, 1048, 464–470. [Google Scholar] [CrossRef]
- Gui, M.; Zhang, M.-x.; Wu, W.-h.; Sun, P. Natural occurrence, bioactivity and biosynthesis of elaiophylin analogues. Mar. Drugs. 2019, 24, 3840. [Google Scholar] [CrossRef] [Green Version]
- Kolouchová, I.; Timkina, E.; Maťátková, O.; Kyselová, L.; Řezanka, T. Analysis of bacteriohopanoids from thermophilic bacteria by liquid chromatography–Mass spectrometry. Microorganisms 2021, 9, 2062. [Google Scholar] [CrossRef]
- Caron, B.; Mark, A.E.; Poger, D. Some like it hot: The effect of sterols and hopanoids on lipid ordering at high temperature. J. Phys. Chem. Lett. 2014, 5, 3953–3957. [Google Scholar] [CrossRef]
- Welander, P.V.; Hunter, R.C.; Zhang, L.; Sessions, A.L.; Summons, R.E.; Newman, D.K. Hopanoids play a role in membrane integrity and pH homeostasis in Rhodopseudomonas palustris TIE-1. J. Bacteriol. 2009, 191, 6145–6156. [Google Scholar] [CrossRef] [Green Version]
- Schäberle, T.F. Biosynthesis of α-pyrones. Beilstein. J. Org. Chem. 2016, 12, 571–588. [Google Scholar] [CrossRef]
- Zhou, Y.; Prediger, P.; Dias, L.C.; Murphy, A.C.; Leadlay, P.F. Macrodiolide formation by the thioesterase of a modular polyketide synthase. Angew. Chem. Int. Ed. 2015, 54, 5232–5235. [Google Scholar] [CrossRef] [Green Version]
- Neunlist, S.; Rohmer, M. A convenient route to an acetylenic C35 hopanoid and the absolute configuration of the side-chain of aminobacteriohopanetriol. Chem. Commun. 1988, 12, 830–832. [Google Scholar] [CrossRef]
- Yurkovich, M.E.; Tyrakis, P.A.; Hong, H.; Sun, Y.; Samborskyy, M.; Kamiya, K.; Leadlay, P.F. A late-stage intermediate in salinomycin biosynthesis is revealed by specific mutation in the biosynthetic gene cluster. ChemBioChem 2012, 13, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Wang, H.; Kang, Q.; Liu, J.; Bai, L. Cloning and characterization of the polyether salinomycin biosynthesis gene cluster of Streptomyces albus XM211. Appl. Environ. Microb. 2012, 78, 994–1003. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Kudo, F.; Rohmer, M.; Eguchi, T. Characterization of radical SAM adenosylhopane synthase, HpnH, which catalyzes the 5′-deoxyadenosyl radical addition to diploptene in the biosynthesis of C35 bacteriohopanepolyols. Angew. Chem. Int. Ed. 2020, 59, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Tian, Y.; Liu, X.; Wang, W.; Li, Y.; Tan, H.; Zhang, J. Reconstitution of a mini-gene cluster combined with ribosome engineering led to effective enhancement of salinomycin production in Streptomyces albus. Microb. Biotechnol. 2021, 14, 2356–2368. [Google Scholar] [CrossRef] [PubMed]
- Luhavaya, H.; Dias, M.V.B.; Williams, S.R.; Hong, H.; de Oliveira, L.G.; Leadlay, P.F. Enzymology of pyran ring a formation in salinomycin biosynthesis. Angew. Chem. Int. Ed. 2015, 127, 13826–13829. [Google Scholar] [CrossRef] [Green Version]
- Chiu, H.-T.; Weng, C.-P.; Lin, Y.-C.; Chen, K.-H. Target-specific identification and characterization of the putative gene cluster for brasilinolide biosynthesis revealing the mechanistic insights and combinatorial synthetic utility of 2-deoxy-l-fucose biosynthetic enzymes. Org. Biomol. Chem. 2016, 14, 1988–2006. [Google Scholar] [CrossRef]
- Capon, R.J. Extracting value: Mechanistic insights into the formation of natural product artifacts—Case studies in marine natural products. Nat. Prod. Rep. 2020, 37, 55–79. [Google Scholar] [CrossRef]
- Epstein, W.W.; Rilling, H.C. Studies on the mechanism of squalene biosynthesis: The structure of presqualene pyrophosphate. J. Biol. Chem. 1970, 245, 4597–4605. [Google Scholar] [CrossRef]
- Pan, J.-J.; Solbiati, J.O.; Ramamoorthy, G.; Hillerich, B.S.; Seidel, R.D.; Cronan, J.E.; Almo, S.C.; Poulter, C.D. Biosynthesis of squalene from farnesyl diphosphate in bacteria: Three steps catalyzed by three enzymes. ACS Cent. Sci. 2015, 1, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Welander, P.V.; Doughty, D.M.; Wu, C.-H.; Mehay, S.; Summons, R.E.; Newman, D.K. Identification and characterization of Rhodopseudomonas palustris TIE-1 hopanoid biosynthesis mutants. Geobiology 2012, 10, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Neunlist, S.; Holst, O.; Rohmer, M. Prokaryotic triterpenoids. Eur. J. Biochem. 1985, 147, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Torres, J.P.; Ammon, M.A.; Marett, L.; Teichert, R.W.; Reilly, C.A.; Kwan, J.C.; Hughen, R.W.; Flores, M.; Tianero, M.D.; et al. A bacterial source for mollusk pyrone polyketides. Chem. Biol. 2013, 20, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]

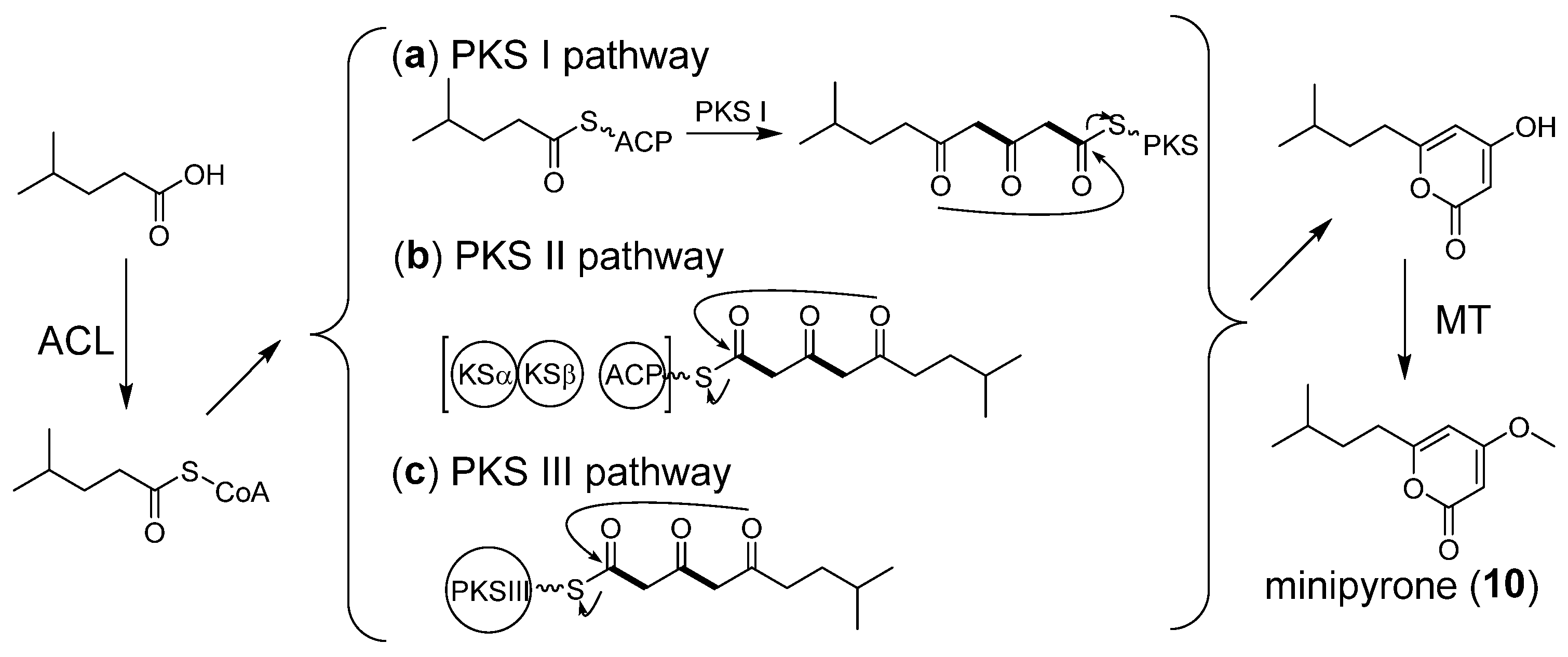

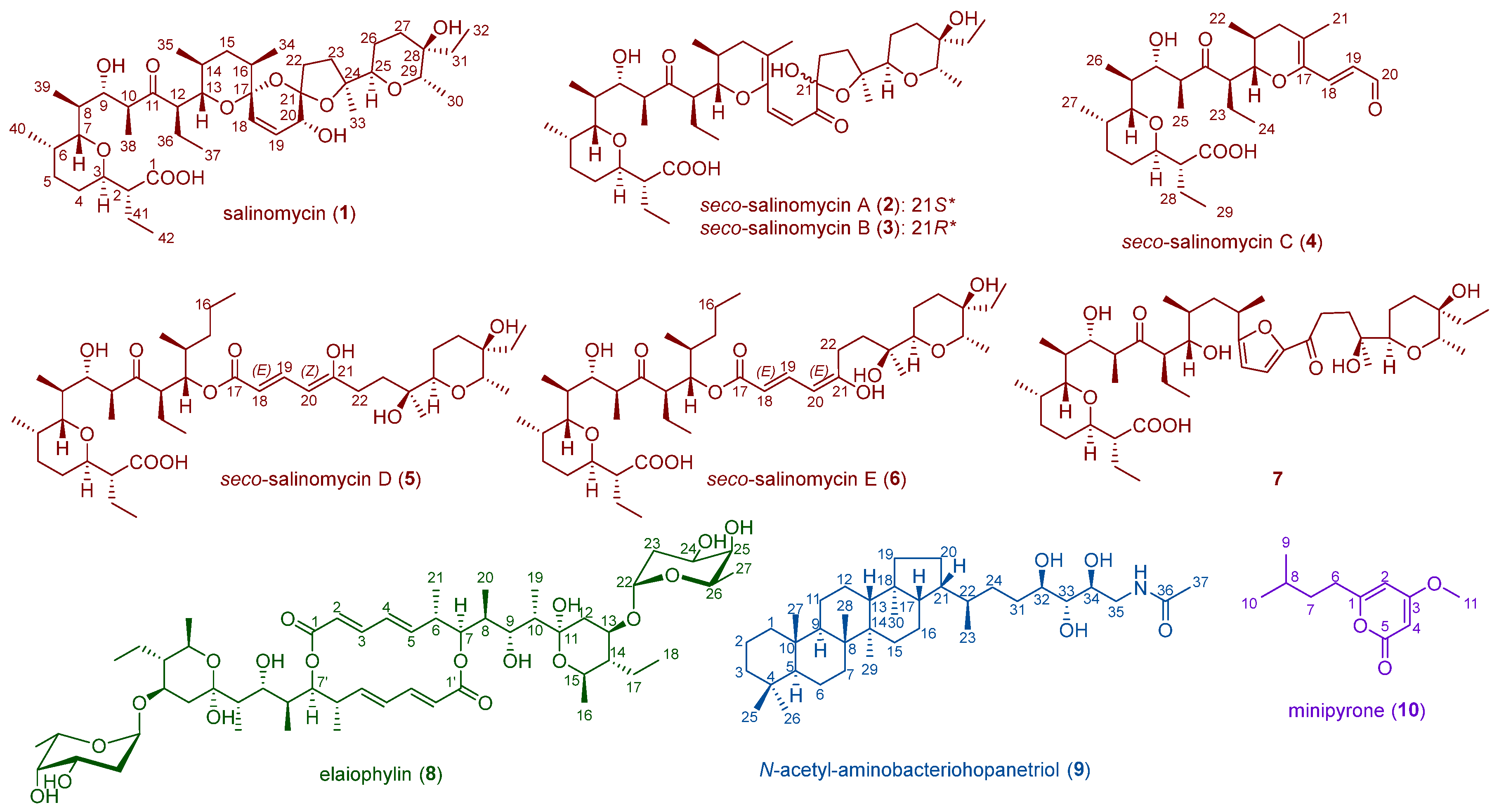{kind=link}
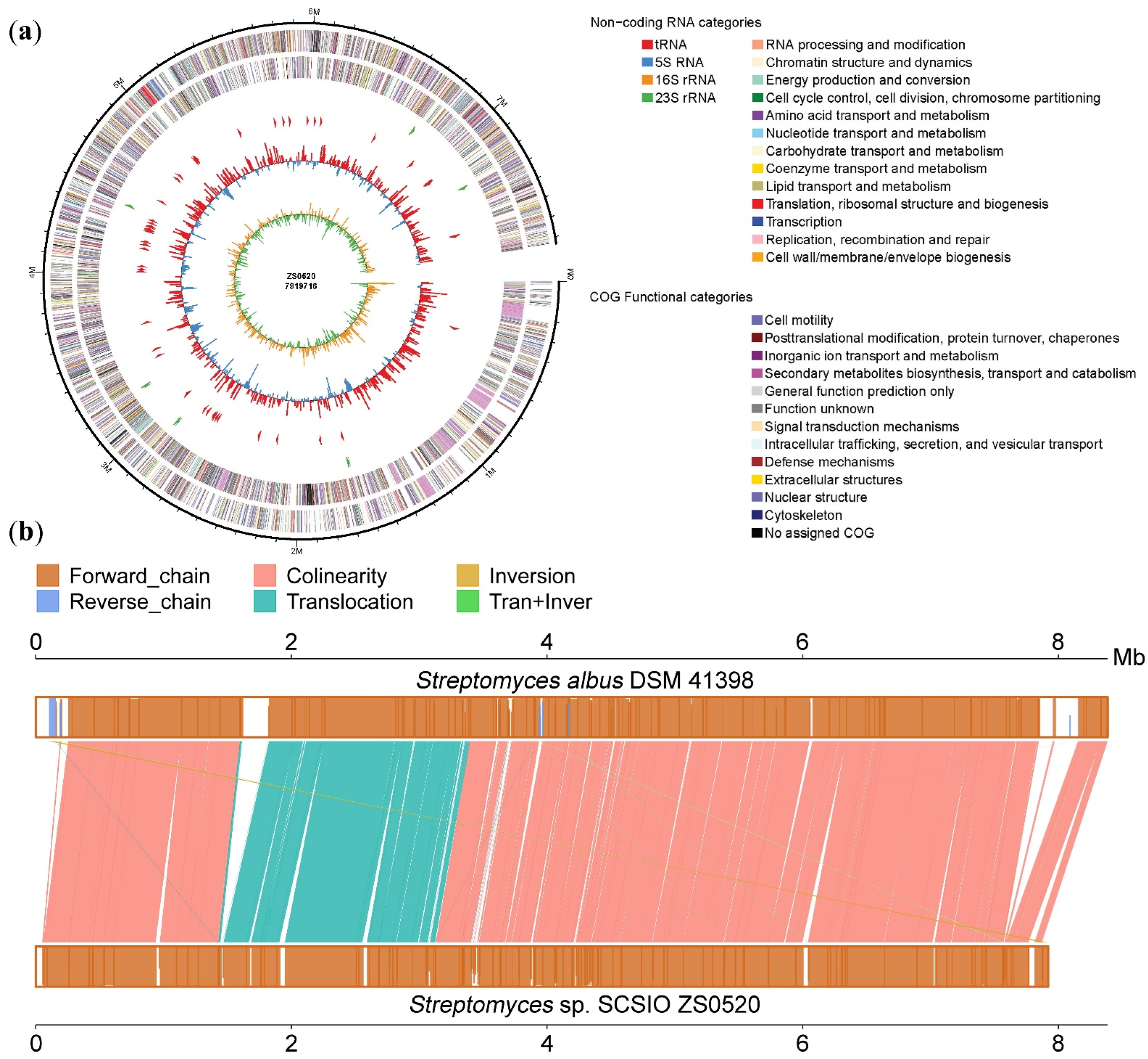{kind=link}
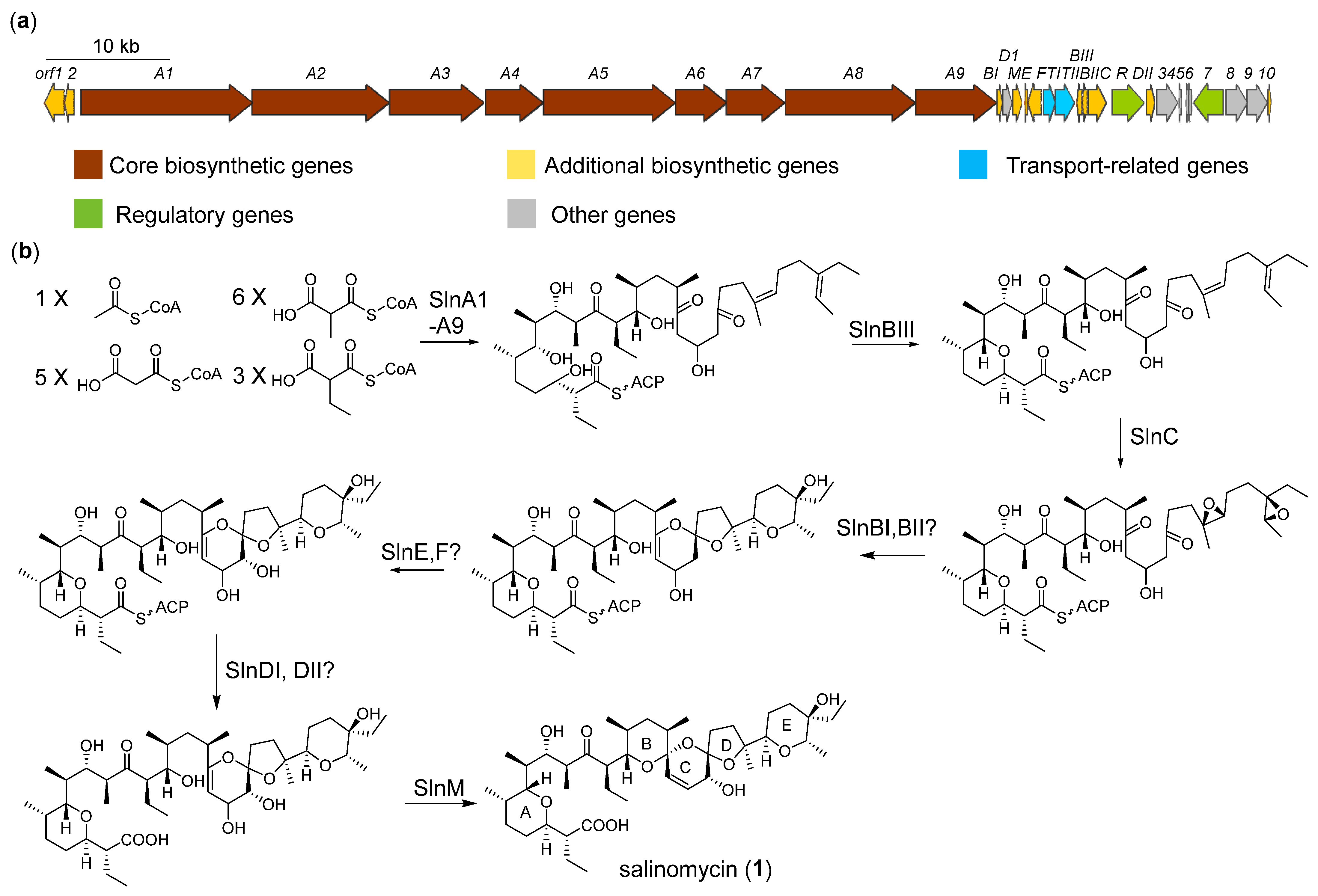{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | Value |
|---|---|
| Genome topology | linear |
| Assembly size (bp) | 7,919,716 |
| G + C content (%) | 72.85 |
| Coding DNA sequences | 5956 |
| tRNA genes | 65 |
| rRNA genes | 18 |
| Secondary metabolite gene clusters | 34 |
| GenBank accession | CP092644 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Chen, Y.; Li, Y.; Song, Y.; Ma, J.; Ju, J. Secondary Metabolites and Biosynthetic Gene Clusters Analysis of Deep-Sea Hydrothermal Vent-Derived Streptomyces sp. SCSIO ZS0520. Mar. Drugs 2022, 20, 393. https://doi.org/10.3390/md20060393
Zhang H, Chen Y, Li Y, Song Y, Ma J, Ju J. Secondary Metabolites and Biosynthetic Gene Clusters Analysis of Deep-Sea Hydrothermal Vent-Derived Streptomyces sp. SCSIO ZS0520. Marine Drugs. 2022; 20(6):393. https://doi.org/10.3390/md20060393
Chicago/Turabian StyleZhang, Huaran, Yingying Chen, Yanqing Li, Yongxiang Song, Junying Ma, and Jianhua Ju. 2022. "Secondary Metabolites and Biosynthetic Gene Clusters Analysis of Deep-Sea Hydrothermal Vent-Derived Streptomyces sp. SCSIO ZS0520" Marine Drugs 20, no. 6: 393. https://doi.org/10.3390/md20060393
APA StyleZhang, H., Chen, Y., Li, Y., Song, Y., Ma, J., & Ju, J. (2022). Secondary Metabolites and Biosynthetic Gene Clusters Analysis of Deep-Sea Hydrothermal Vent-Derived Streptomyces sp. SCSIO ZS0520. Marine Drugs, 20(6), 393. https://doi.org/10.3390/md20060393