
Skin Microbiota, Circulating Metabolic Biomarker, Epilepsy: A Mendelian Randomization Analysis
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sources
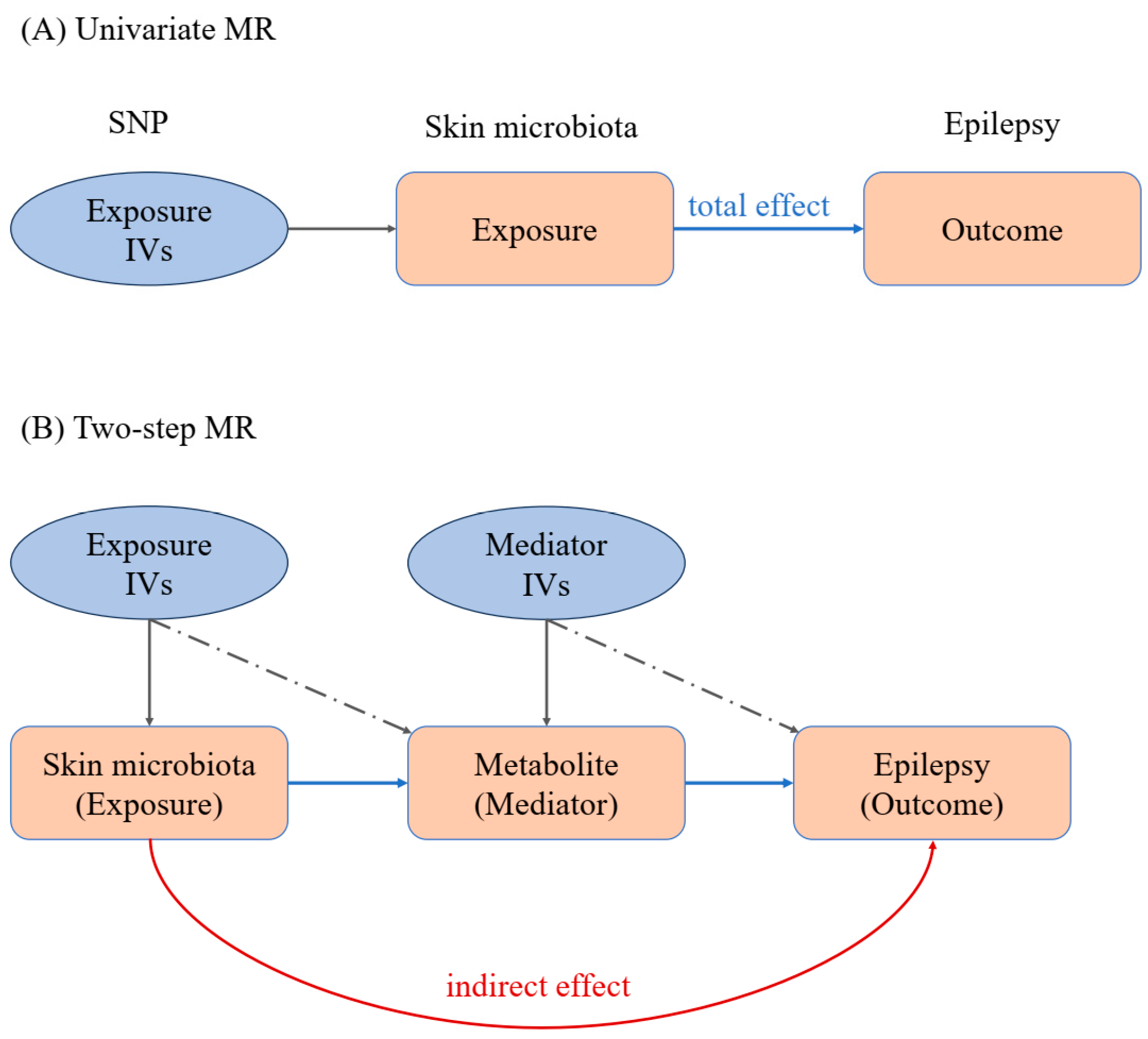
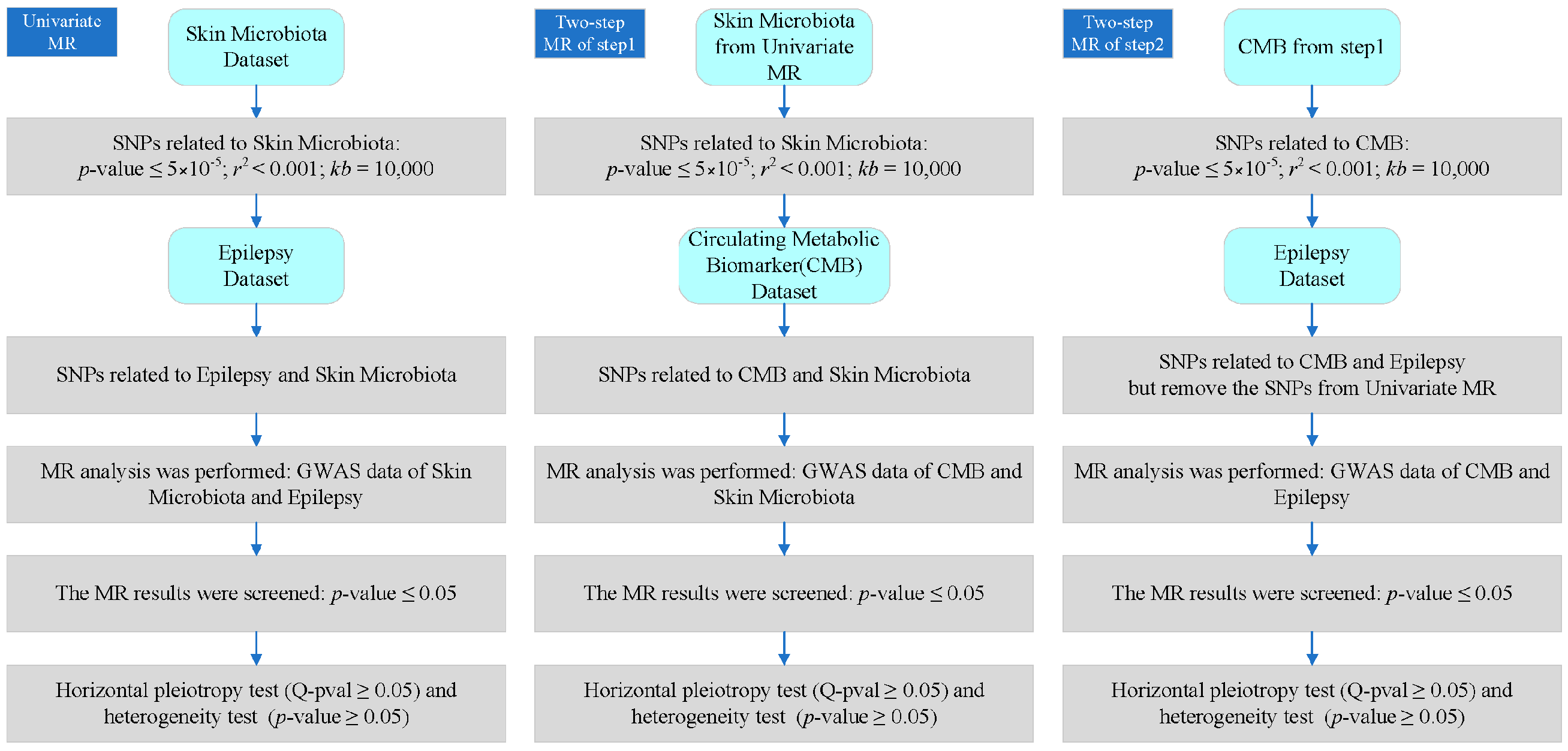
2.2. Univariate MR Analysis
- (1)
- Using a strict genome-wide significance threshold (p ≤ 5 × 10−5), SNPs were extracted from GWAS exposure data, establishing a strong association with exposure [24].
- (2)
- Remove SNPs with linkage disequilibrium (LD). When the distributions of two SNPs are correlated rather than independent, LD occurs, which violates the assumption of random classification and may bias the results. In this study, we used the “TwoSampleMR (version: 0.5.7)” package to screen SNPs with parameters r2 = 0.001 and kb = 10,000, which means that SNPs with r2 = 0.001 are removed within the 10,000 kb region surrounding the most significant SNP to eliminate potential LDs.
- (3)
- Screen out SNPs related to obsessive–compulsive disorder from the relevant GWAS data of the results. After initial SNP screening, calculate the total F-value of IVs to assess the strength of instrumental variables and mitigate potential bias caused by weak IVs. If the F value exceeds 10, it can be considered a strong instrumental variable.
2.3. Two-Step MR Analysis
2.4. Analysis of Heterogeneity
2.5. Analysis of Horizontal Pleiotropy
2.6. Analysis of Effect Value
2.7. Associated Genes Analysis
3. Results
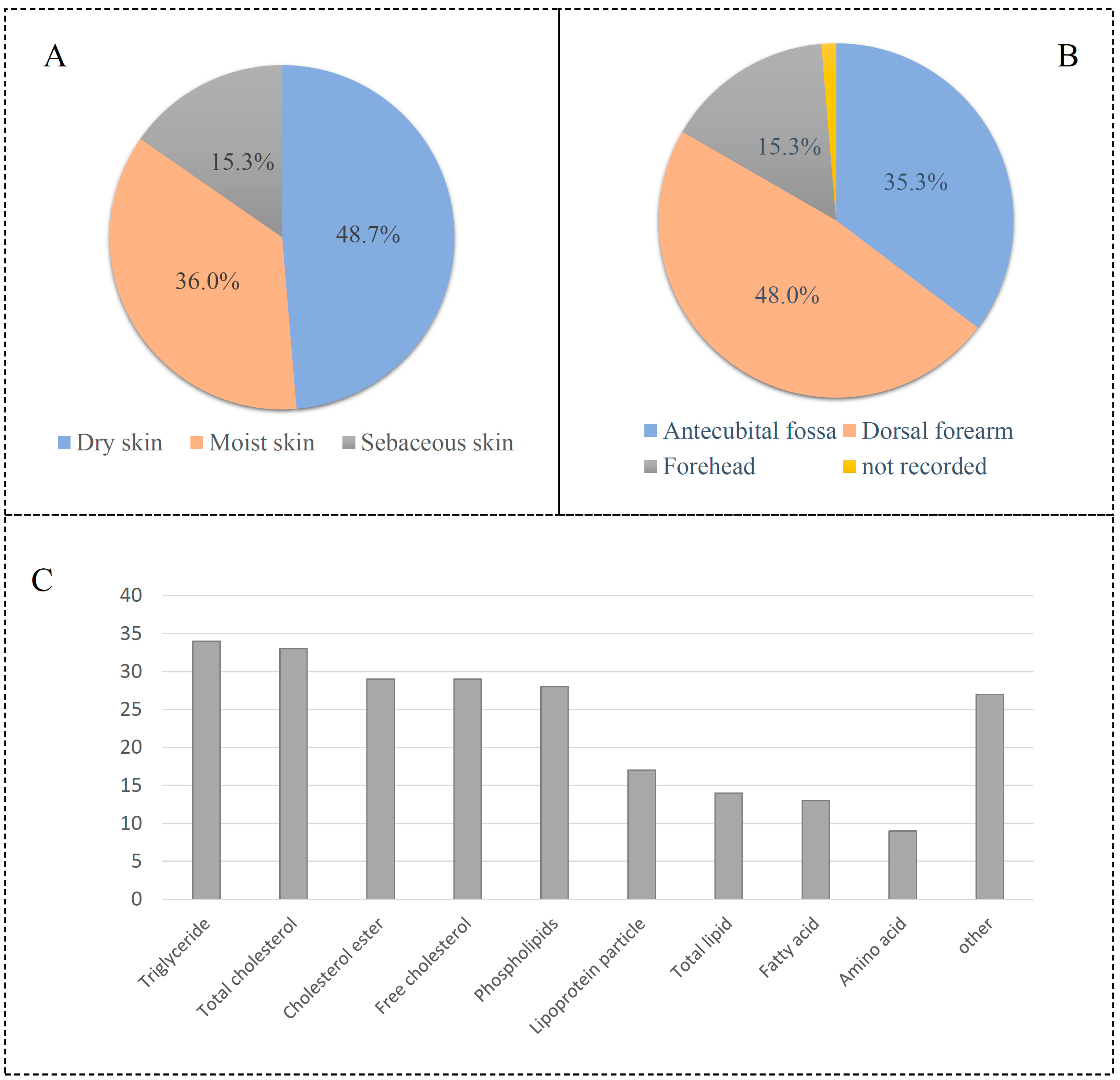
3.1. Data Classfication
3.2. Univariate MR Results
3.3. Two-Step MR Results
3.4. Results of Heterogeneity Analysis
3.5. Horizontal Pleiotropy
3.6. Effect Value
3.7. Associated Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Christensen, J.; Dreier, J.W.; Sun, Y.; Linehan, C.; Tomson, T.; Marson, A.; Forsgren, L.; Granbichler, C.A.; Trinka, E.; Illiescu, C. Estimates of epilepsy prevalence, psychiatric co-morbidity and cost. Seizure Eur. J. Epilepsy 2023, 107, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Sander, J.W. The global burden of epilepsy report: Implications for low-and middle-income countries. Epilepsy Behav. 2020, 105, 106949. [Google Scholar] [CrossRef] [PubMed]
- Falco-Walter, J.J.; Scheffer, I.E.; Fisher, R.S. The new definition and classification of seizures and epilepsy. Epilepsy Res. 2018, 139, 73–79. [Google Scholar] [CrossRef]
- Scheffer, I.E.; Berkovic, S.; Capovilla, G.; Connolly, M.B.; French, J.; Guilhoto, L.; Hirsch, E.; Jain, S.; Mathern, G.W.; Moshé, S.L. ILAE classification of the epilepsies: Position paper of the ILAE Commission for Classification and Terminology. Z. Epileptol. 2018, 31, 296–306. [Google Scholar] [CrossRef]
- Dichter, M.A. Emerging concepts in the pathogenesis of epilepsy and epileptogenesis. Arch. Neurol. 2009, 66, 443–447. [Google Scholar] [CrossRef]
- Porter, R.J.; Dhir, A.; Macdonald, R.L.; Rogawski, M.A. Chapter 39—Mechanisms of Action of Antiseizure Drugs. In Handbook of Clinical Neurology; Stefan, H., Theodore, W.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 108, pp. 663–681. [Google Scholar]
- Kwan, P.; Schachter, S.C.; Brodie, M.J. Drug-resistant epilepsy. N. Engl. J. Med. 2011, 365, 919–926. [Google Scholar] [CrossRef]
- Thakran, S.; Guin, D.; Singh, P.; Singh, P.; Kukal, S.; Rawat, C.; Yadav, S.; Kushwaha, S.S.; Srivastava, A.K.; Hasija, Y. Genetic landscape of common epilepsies: Advancing towards precision in treatment. Int. J. Mol. Sci. 2020, 21, 7784. [Google Scholar] [CrossRef]
- De Caro, C.; Iannone, L.F.; Citraro, R.; Striano, P.; De Sarro, G.; Constanti, A.; Cryan, J.F.; Russo, E. Can we ‘seize’the gut microbiota to treat epilepsy? Neurosci. Biobehav. Rev. 2019, 107, 750–764. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota–host interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 2016, 352, 565–569. [Google Scholar] [CrossRef]
- Fu, J.; Bonder, M.J.; Cenit, M.C.; Tigchelaar, E.F.; Maatman, A.; Dekens, J.A.; Brandsma, E.; Marczynska, J.; Imhann, F.; Weersma, R.K. The gut microbiome contributes to a substantial proportion of the variation in blood lipids. Circ. Res. 2015, 117, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.-F.; Oddos, T.; Stamatas, G. Deciphering the role of skin surface microbiome in skin health: An integrative multiomics approach reveals three distinct metabolite–microbe clusters. J. Investig. Dermatol. 2022, 142, 469–479.e5. [Google Scholar] [CrossRef]
- Rho, J.M.; Boison, D. The metabolic basis of epilepsy. Nat. Rev. Neurol. 2022, 18, 333–347. [Google Scholar] [CrossRef]
- Godoi, A.B.; do Canto, A.M.; Donatti, A.; Rosa, D.C.; Bruno, D.C.; Alvim, M.K.; Yasuda, C.L.; Martins, L.G.; Quintero, M.; Tasic, L. Circulating metabolites as biomarkers of disease in patients with mesial temporal lobe epilepsy. Metabolites 2022, 12, 446. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.; Du, D.; Chen, L. Metabolomics provides novel insights into epilepsy diagnosis and treatment: A review. Neurochem. Res. 2022, 47, 844–859. [Google Scholar] [CrossRef]
- Ji, D.; Chen, W.-Z.; Zhang, L.; Zhang, Z.-H.; Chen, L.-J. Gut microbiota, circulating cytokines and dementia: A Mendelian randomization study. J. Neuroinflamm. 2024, 21, 2. [Google Scholar] [CrossRef]
- Sanderson, E. Multivariable Mendelian randomization and mediation. Cold Spring Harb. Perspect. Med. 2021, 11, a038984. [Google Scholar] [CrossRef] [PubMed]
- Rogne, T.; Gill, D.; Liew, Z.; Shi, X.; Stensrud, V.H.; Nilsen, T.I.L.; Burgess, S. Mediating factors in the association of maternal educational level with pregnancy outcomes: A Mendelian Randomization Study. JAMA Netw. Open 2024, 7, e2351166. [Google Scholar] [CrossRef]
- Sollis, E.; Mosaku, A.; Abid, A.; Buniello, A.; Cerezo, M.; Gil, L.; Groza, T.; Güneş, O.; Hall, P.; Hayhurst, J. The NHGRI-EBI GWAS Catalog: Knowledgebase and deposition resource. Nucleic Acids Res. 2023, 51, D977–D985. [Google Scholar] [CrossRef]
- Kurki, M.I.; Karjalainen, J.; Palta, P.; Sipilä, T.P.; Kristiansson, K.; Donner, K.M.; Reeve, M.P.; Laivuori, H.; Aavikko, M.; Kaunisto, M.A. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature 2023, 613, 508–518. [Google Scholar] [CrossRef]
- Karjalainen, M.K.; Karthikeyan, S.; Oliver-Williams, C.; Sliz, E.; Allara, E.; Fung, W.T.; Surendran, P.; Zhang, W.; Jousilahti, P.; Kristiansson, K. Genome-wide characterization of circulating metabolic biomarkers. Nature 2024, 628, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Deng, Y.; Pan, W. Combining the strengths of inverse-variance weighting and Egger regression in Mendelian randomization using a mixture of regressions model. PLoS Genet. 2021, 17, e1009922. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Lai, J.; Liu, Z.; Liu, W.; Zhang, Y. Specific blood metabolite associations with Gout: A Mendelian randomization study. Eur. J. Clin. Nutr. 2024, 8, 1–9. [Google Scholar] [CrossRef]
- Ong, J.S.; MacGregor, S. Implementing MR-PRESSO and GCTA-GSMR for pleiotropy assessment in Mendelian randomization studies from a practitioner’s perspective. Genet. Epidemiol. 2019, 43, 609–616. [Google Scholar] [CrossRef]
- Vabistsevits, M.; Davey Smith, G.; Sanderson, E.; Richardson, T.G.; Lloyd-Lewis, B.; Richmond, R.C. Deciphering how early life adiposity influences breast cancer risk using Mendelian randomization. Commun. Biol. 2022, 5, 337. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Taskesen, E.; Van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef]
- Shen, W.; Song, Z.; Zhong, X.; Huang, M.; Shen, D.; Gao, P.; Qian, X.; Wang, M.; He, X.; Wang, T. Sangerbox: A comprehensive, interaction-friendly clinical bioinformatics analysis platform. Imeta 2022, 1, e36. [Google Scholar] [CrossRef] [PubMed]
- Sumadewi, K.T.; Harkitasari, S.; Tjandra, D.C. Biomolecular mechanisms of epileptic seizures and epilepsy: A review. Acta Epileptol. 2023, 5, 28. [Google Scholar] [CrossRef]
- Ito, Y.; Amagai, M. Dissecting skin microbiota and microenvironment for the development of therapeutic strategies. Curr. Opin. Microbiol. 2023, 74, 102311. [Google Scholar] [CrossRef]
- Chen, H.-A.; Yang, C.-J.; Tsai, M.-S.; Liao, C.-H.; Lee, C.-H. Linezolid as salvage therapy for central nervous system infections due to methicillin-resistant Staphylococcus aureus at two medical centers in Taiwan. J. Microbiol. Immunol. Infect. 2020, 53, 909–915. [Google Scholar] [CrossRef]
- van der Spek, A.; Stewart, I.D.; Kühnel, B.; Pietzner, M.; Alshehri, T.; Gauß, F.; Hysi, P.G.; Mahmoudian Dehkordi, S.; Heinken, A.; Luik, A.I. Circulating metabolites modulated by diet are associated with depression. Mol. Psychiatry 2023, 28, 3874–3887. [Google Scholar] [CrossRef]
- Huang, W.; Wang, Z.; Zou, C.; Liu, Y.; Pan, Y.; Lu, J.; Zhou, K.; Jiao, F.; Zhong, S.; Jiang, G. Effects of metabolic factors in mediating the relationship between type 2 diabetes and depression in east Asian populations: A two-step, two-sample mendelian randomization study. J. Affect. Disord. 2023, 335, 120–128. [Google Scholar] [CrossRef]
- Sarlo, G.L.; Holton, K.F. Brain concentrations of glutamate and GABA in human epilepsy: A review. Seizure 2021, 91, 213–227. [Google Scholar] [CrossRef]
- Holden, K.; Hartman, A.L. d-Leucine: Evaluation in an epilepsy model. Epilepsy Behav. 2018, 78, 202–209. [Google Scholar] [CrossRef]
- Neves, G.S.; Lunardi, M.S.; Lin, K.; Rieger, D.K.; Ribeiro, L.C.; Moreira, J.D. Ketogenic diet, seizure control, and cardiometabolic risk in adult patients with pharmacoresistant epilepsy: A review. Nutr. Rev. 2021, 79, 931–944. [Google Scholar] [CrossRef]
- Islam, M.A.; Alam, F.; Cavestro, C.; Calcii, C.; Sasongko, T.H.; Levy, R.A.; Gan, S.H. Antiphospholipid antibodies in epilepsy: A systematic review and meta-analysis. Autoimmun. Rev. 2018, 17, 755–767. [Google Scholar] [CrossRef]
- Chen, Z.-P.; Wang, S.; Zhao, X.; Fang, W.; Wang, Z.; Ye, H.; Wang, M.-J.; Ke, L.; Huang, T.; Lv, P. Lipid-accumulated reactive astrocytes promote disease progression in epilepsy. Nat. Neurosci. 2023, 26, 542–554. [Google Scholar] [CrossRef]
- Liu, S.; He, Z.; Shi, W.; Li, J. The association between APOE gene polymorphisms and the risk, characteristics, and prognosis of epilepsy: A systematic review and meta-analysis. Epilepsy Behav. 2024, 160, 110070. [Google Scholar] [CrossRef]
- Zhou, K.; Yang, H.; Chen, R.; Wang, W.; Qu, Z. Causal relationship among obesity and body fat distribution and epilepsy subtypes. Front. Neurol. 2022, 13, 984824. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Skin Microbiota | Skin Type | Skin Location | Outcome | p-Value | OR | 95% CI |
|---|---|---|---|---|---|---|
| asv008 | sebaceous skin | forehead | epilepsy | 0.0114 | 0.9899 | 0.982–0.998 |
| asv022 | moist skin | antecubital fossa | epilepsy | 0.0083 | 0.9886 | 0.980–0.997 |
| asv039 | moist skin | antecubital fossa | epilepsy | 0.0081 | 1.0101 | 1.002–1.018 |
| asv042 | moist skin | antecubital fossa | epilepsy | 0.042 | 0.9924 | 0.985–0.999 |
| bacillales | dry skin | dorsal forearm | epilepsy | 0.0195 | 1.0119 | 1.002–1.022 |
| clostridiales | dry skin | dorsal forearm | epilepsy | 0.0428 | 1.0102 | 1.000–1.020 |
| micrococcaceae | dry skin | dorsal forearm | epilepsy | 0.0397 | 1.0111 | 1.000–1.021 |
| neisseriaceae | sebaceous skin | forehead | epilepsy | 0.0291 | 1.0133 | 1.001–1.025 |
| staphylococcus | dry skin | dorsal forearm | epilepsy | 0.0287 | 1.0106 | 1.001–1.020 |
| Step | Exposure | Detail | Outcome | p-Value | OR | 95% CI |
|---|---|---|---|---|---|---|
| Step 1 | asv008 | sebaceous skin in forehead | GCST90302078 | 0.0294 | 1.0003 | 1.0000–1.0051 |
| asv022 | moist skin in antecubital fossa | GCST90301994 | 0.0011 | 0.9960 | 0.9936–0.9984 | |
| bacillales | dry skin in dorsal forearm | GCST90302151 | 0.0316 | 0.9932 | 0.9932–0.9996 | |
| clostridiales | dry skin in dorsal forearm | GCST90302078 | 0.0381 | 0.9969 | 0.9939–0.9998 | |
| micrococcaceae | dry skin in dorsal forearm | GCST90301965 | 0.0291 | 0.9963 | 0.9930–0.9996 | |
| Step 2 | GCST90301965 | Glutamine level | epilepsy | 0.0372 | 1.0655 | 1.0038–1.1311 |
| GCST90301994 | Leucine level | epilepsy | 0.0487 | 1.0675 | 1.0004–1.1391 | |
| GCST90302078 | Ratio of saturated fatty acid | epilepsy | 0.0309 | 0.9017 | 0.8209–0.9905 | |
| GCST90302151 | Phospholipids in medium LDL | epilepsy | 0.0302 | 0.9308 | 0.8723–0.9932 |
| Step | Exposure | Outcome | Method | Q Value | Q-df | Q-pval |
|---|---|---|---|---|---|---|
| Step 1 | asv008 | GCST90302078 | IVW | 38.9 | 49 | 0.850 |
| asv022 | GCST90301994 | IVW | 36.1 | 41 | 0.690 | |
| bacillales | GCST90302151 | IVW | 56.5 | 47 | 0.162 | |
| clostridiales | GCST90302078 | IVW | 60.3 | 52 | 0.200 | |
| micrococcaceae | GCST90301965 | IVW | 57.6 | 51 | 0.243 | |
| asv008 | GCST90302078 | MR Egger | 37.1 | 48 | 0.874 | |
| asv022 | GCST90301994 | MR Egger | 35.5 | 40 | 0.674 | |
| bacillales | GCST90302151 | MR Egger | 56.5 | 46 | 0.139 | |
| clostridiales | GCST90302078 | MR Egger | 59.1 | 51 | 0.203 | |
| micrococcaceae | GCST90301965 | MR Egger | 57.6 | 50 | 0.214 | |
| Step 2 | GCST90301994 | epilepsy | IVW | 204.6 | 189 | 0.207 |
| GCST90302151 | epilepsy | IVW | 226.2 | 207 | 0.171 | |
| GCST90302078 | epilepsy | IVW | 149.0 | 151 | 0.530 | |
| GCST90302078 | epilepsy | IVW | 149.0 | 151 | 0.530 | |
| GCST90301965 | epilepsy | IVW | 246.9 | 224 | 0.141 | |
| GCST90301994 | epilepsy | MR Egger | 204.1 | 188 | 0.200 | |
| GCST90302151 | epilepsy | MR Egger | 226.1 | 206 | 0.160 | |
| GCST90302078 | epilepsy | MR Egger | 149.0 | 150 | 0.507 | |
| GCST90302078 | epilepsy | MR Egger | 149.0 | 150 | 0.507 | |
| GCST90301965 | epilepsy | MR Egger | 246.8 | 223 | 0.131 |
| Step | Exposure | Outcome | Intercept | SE | p-Value |
|---|---|---|---|---|---|
| Step 1 | asv022 | GCST90301994 | 0.0026 | 0.0035 | 0.4511 |
| bacillales | GCST90302151 | 0.0001 | 0.0037 | 0.9716 | |
| asv008 | GCST90302078 | −0.0041 | 0.0031 | 0.1856 | |
| clostridiales | GCST90302078 | −0.0036 | 0.0035 | 0.3179 | |
| micrococcaceae | GCST90301965 | 0.0001 | 0.0034 | 0.9798 | |
| Step 2 | GCST90301994 | epilepsy | 0.0015 | 0.0021 | 0.4842 |
| GCST90302151 | epilepsy | −0.0006 | 0.0023 | 0.7865 | |
| GCST90302078 | epilepsy | 0.0003 | 0.0032 | 0.9161 | |
| GCST90301965 | epilepsy | −0.0005 | 0.0020 | 0.8182 |
| Exposure | Mediator | Outcome | Total Effect | Indirect Effect | Direct Effect | Mediator Percentage |
|---|---|---|---|---|---|---|
| asv022 | GCST90301994 | epilepsy | −1.149% | −0.026% | −1.123% | 2.26% |
| bacillales | GCST90302151 | epilepsy | 1.183% | 0.025% | 1.158% | 2.15% |
| asv008 | GCST90302078 | epilepsy | −1.013% | −0.001% | −1.012% | 0.08% |
| clostridiales | GCST90302078 | epilepsy | 1.015% | −0.001% | 1.016% | −0.08% |
| micrococcaceae | GCST90301965 | epilepsy | 1.106% | −0.023% | 1.129% | −2.10% |
| Gene | GCST90301965 | GCST90301994 | GCST90302078 | GCST90302151 | Number |
|---|---|---|---|---|---|
| MLXIPL | rs13246993 | rs80189144 | rs6460047 | rs17145738 | 4 |
| AC109829.1 | rs1260326 | rs1260326 | NA | rs1260326 | 3 |
| EIF2B4 | rs1260326 | rs1260326 | NA | rs1260326 | 3 |
| GCKR | rs1260326 | rs1260326 | NA | rs1260326 | 3 |
| PPM1G | rs1260326 | rs1260326 | NA | rs1260326 | 3 |
| SNX17 | rs1260326 | rs1260326 | NA | rs1260326 | 3 |
| ZNF513 | rs1260326 | rs1260326 | NA | rs1260326 | 3 |
| APOC1 | NA | rs390082 | rs429358 | rs7412 | 3 |
| APOE | NA | rs390082 | rs429358 | rs7412 | 3 |
| TOMM40 | NA | rs390082 | rs429358 | rs7412 | 3 |
| ARNTL2 | rs61915900 | rs71534286 | NA | rs71534286 | 3 |
| PPFIBP1 | rs74969426 | rs74969426 | NA | rs74969426 | 3 |
| PTHLH | rs71534286 | rs71534286 | NA | rs71534286 | 3 |
| BAZ1B | rs13246993 | rs80189144 | NA | rs17145738 | 3 |
| BCL7B | rs13246993 | rs80189144 | NA | rs17145738 | 3 |
| FZD9 | rs13246993 | rs80189144 | NA | rs17145738 | 3 |
| TBL2 | rs13246993 | rs80189144 | NA | rs17145738 | 3 |
| BUD13 | NA | rs964184 | rs964184 | rs144018203 | 3 |
| ZNF259 | NA | rs964184 | rs964184 | rs964184 | 3 |
| APOA5 | NA | rs964184 | rs964184 | rs144018203 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, T.; Wu, S.; Sun, J.; Mu, M.; Yang, Z. Skin Microbiota, Circulating Metabolic Biomarker, Epilepsy: A Mendelian Randomization Analysis. Curr. Issues Mol. Biol. 2024, 46, 13937-13950. https://doi.org/10.3390/cimb46120833
Wang Y, Liu T, Wu S, Sun J, Mu M, Yang Z. Skin Microbiota, Circulating Metabolic Biomarker, Epilepsy: A Mendelian Randomization Analysis. Current Issues in Molecular Biology. 2024; 46(12):13937-13950. https://doi.org/10.3390/cimb46120833
Chicago/Turabian StyleWang, Yongheng, Tianyi Liu, Shangying Wu, Jiawei Sun, Mingdao Mu, and Zhiyuan Yang. 2024. "Skin Microbiota, Circulating Metabolic Biomarker, Epilepsy: A Mendelian Randomization Analysis" Current Issues in Molecular Biology 46, no. 12: 13937-13950. https://doi.org/10.3390/cimb46120833
APA StyleWang, Y., Liu, T., Wu, S., Sun, J., Mu, M., & Yang, Z. (2024). Skin Microbiota, Circulating Metabolic Biomarker, Epilepsy: A Mendelian Randomization Analysis. Current Issues in Molecular Biology, 46(12), 13937-13950. https://doi.org/10.3390/cimb46120833