
Contribution of Lipid Oxidation and Ferroptosis to Radiotherapy Efficacy
by
 , ,
, ,
Ashley N. Pearson
1 ,
,
Joseph Carmicheal
2,
Long Jiang
3,
Yu Leo Lei
4,5,6 and
Michael D. Green
3,6,7,8,9,* 1
Program in Biomedical Sciences, University of Michigan Medical School, Ann Arbor, MI 48109, USA
2
College of Medicine, University of Nebraska Medical Center, Omaha, NE 68198, USA
3
Department of Radiation Oncology, University of Michigan School of Medicine, Ann Arbor, MI 48109, USA
4
Department of Periodontics and Oral Medicine, University of Michigan, Ann Arbor, MI 48109, USA
5
Department of Otolaryngology, University of Michigan, Ann Arbor, MI 48109, USA
6
Rogel Cancer Center, University of Michigan, Ann Arbor, MI 48109, USA
7
Department of Microbiology and Immunology, University of Michigan School of Medicine, Ann Arbor, MI 48109, USA
8
Graduate Program in Immunology, University of Michigan School of Medicine, Ann Arbor, MI 48109, USA
9
Veterans Affairs Ann Arbor Healthcare System, Ann Arbor, MI 48105, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(22), 12603; https://doi.org/10.3390/ijms222212603
Submission received: 21 October 2021
/
Revised: 16 November 2021
/
Accepted: 18 November 2021
/
Published: 22 November 2021
(This article belongs to the Special Issue Radiation Biology and Molecular Radiation Oncology)
Abstract
:Radiotherapy promotes tumor cell death and senescence through the induction of oxidative damage. Recent work has highlighted the importance of lipid peroxidation for radiotherapy efficacy. Excessive lipid peroxidation can promote ferroptosis, a regulated form of cell death. In this review, we address the evidence supporting a role of ferroptosis in response to radiotherapy and discuss the molecular regulators that underlie this interaction. Finally, we postulate on the clinical implications for the intersection of ferroptosis and radiotherapy.
1. Introduction
Radiotherapy is used in the curative and palliative management of more than half of all cancer patients [1]. Unfortunately, resistance to radiotherapy limits its therapeutic efficacy. Radiotherapy induces multiple forms of regulated and unregulated cell death [2]. Without an improved understanding of the molecular mechanisms behind radiotherapy induced cell death, the design of new therapeutic strategies which augment radiotherapy efficacy is limited. Recently, it has been found that radiotherapy generated lipid oxidation and ferroptosis [3,4,5]. In this review, we will discuss the connections between lipid oxidation, ferroptosis, and radiotherapy, as well as elaborate on the clinical implications of ferroptosis modulators as radiosensitizers.
2. Molecular Regulators of Ferroptosis
Lipids are a critical buffer to reactive oxygen species (ROS) and exist in an equilibrium between oxidized and reduced states. This balance is known as lipid redox homeostasis, a process which is essential for ensuring cell survival [6]. One of the lethal outcomes of redox imbalance is ferroptosis, a unique form of cell death induced by iron-dependent lipid peroxidation [7]. The term “ferroptosis” was only recently described, although the phenomenon of lipid oxidative damage has been observed for decades [8]. By the 1980s, the role of lipid oxidation in cell stress was well-described [9,10]. It was not until more recently that it was discovered that excessive lipid oxidation leads to cell death. However, the precise mechanism through which lipid oxidation causes ferroptosis is still unknown [11,12]. Lipid peroxidation is the key driver of ferroptosis [13,14]. Specifically, oxidation of polyunsaturated fatty acids (PUFA), mediated by lipoxygenases and the intracellular iron pool, which is in turn regulated by phosphorylase kinase catalytic subunit gamma 2 (PHKG2), drives ferroptosis [15]. Synthesis of PUFAs containing ether phospholipids provides substrates that are then peroxidated to drive ferroptosis [16]. Acyl-coenzyme A synthetase long-chain family member 4 (ACSL4) esterifies CoA to free fatty acids, with a preference for long PUFAs [17]. ACSL4 inhibits redox homeostasis by allowing for accumulation of oxidized lipids within the plasma membrane [18]. In contrast, it has been shown that introduction of monounsaturated fatty acids inhibits ferroptosis by decreasing membrane ROS content via acyl-CoA synthetase long-chain family member 3 (ACSL3) activity [19]. Another aspect to consider is mitochondrial electron transport chain activity, which contributes to the generation of endogenous lipid radicals to induce ferroptosis [20]. Thus, lipid metabolism and biosynthesis regulate ferroptosis.
Lipid oxidation and ferroptosis are limited by multiple pathways. One critical regulator is glutathione peroxidase 4 (GPX4), a mammalian glutathione peroxidase which inhibits lipid peroxidation and contributes to redox homeostasis by catalyzing the reduction of lipid peroxides [21,22,23]. In the context of B cell lymphomas and renal cell carcinomas, GPX4 regulates induction of ferroptosis. RSL-3, a pharmacological ferroptosis inducer, drives colorectal cancer cell death via direct binding and inactivation of GPX4, resulting in loss of redox homeostasis [22]. Other defenses include ferroptosis suppressor protein 1 (FSP1), which functions as a coenzyme Q10 oxidoreductase and restores the antioxidant pool to suppress ferroptosis [24]. These data highlight that multiple enzymatic pathways act in concert to limit lipid peroxidation induced cell death. Antioxidants are required for the enzymatic reduction of lipids and prevention of ferroptosis. Tetrahydrobiopterin, generated by GTP cyclohydrolase I, is a hydroxylase cofactor and antioxidant which limits ferroptosis [25,26]. Cysteine, the reduced form of cystine, is the rate-limiting precursor for the antioxidant glutathione (GSH). In the 1950s, Eagle demonstrated the importance of cystine presence for Hela cell survival [27]. Depletion of GSH has been linked to diminished GPX4 activity and ferroptosis in cancer cells [28,29]. Cystine import is regulated by system xc−, a cystine-glutamine anti-transporter [11] composed of a heavy chain (Solute Light Chain, SLC3A2) and a light chain (SLC7A11) [30]. Interestingly, tumor protein 53 (p53) increases expression of SLC7A11, driving cystine import to restrain oxidative stress and thus prevent ferroptotic cell death in cancer cells [31]. In murine pancreatic cancer cells, deletion of SLC7A11 was sufficient to decrease cystine import, downregulate GSH activity, and induce tumor ferroptosis [28]. Additionally, interferon-γ from CD8+ T cells can promote tumor cell ferroptosis by downregulating SLC3A2 and SLC7A11, impairing tumor cystine uptake and disrupting tumor cell redox homeostasis [32]. Other regulators of system xc− include nuclear factor erythroid factor 2-related factor 2 and kelch-like ECH-associated protein 1 (NRF2-KEAP1) signaling [33]. This may be exploited to treat cancer by sensitizing it to radiotherapy [5]. Further evidence of the importance of antioxidants to ferroptosis comes from metabolic perturbations. Use of a glutamine antagonist increased T cell-mediated antitumor immunity in several murine cancer models [34]. However, Gao et al. showed that glutamine supplementation induced ferroptosis in mouse embryonic fibroblasts, and Rossler et al. demonstrated that elevated glutamate contributes to cell death in HT22 cells [35,36]. More work is needed to better define the role of glutamine on ferroptosis in cancer. Thus, metabolic import and antioxidant biosynthetic pathways regulate ferroptosis.
Labile iron radicals can directly generate oxygen radicals via Fenton chemistry [37] and promote lipid peroxidation directly [38]. Iron can also indirectly promote ferroptosis as a cofactor in the enzymes which promote lipid oxidation [7]. For example, cytochrome P450 oxidoreductase depends on the cycling of iron between ferric and ferrous states to enable lipid peroxidation [39]. Lipoxygenase enzymes, which are iron dependent and promote lipid oxidation, also promote ferroptosis [40]. It has been shown that pharmacologic iron chelators as well as physiologic iron chelators can inhibit ferroptosis, although the precise mechanism by which this occurs is not yet known [7,41]. These data underscore the necessity of iron to induce ferroptosis. Collectively, this work highlights the multiple modules that regulate lipid redox homeostasis (Figure 1).
3. Evidence of Lipid Oxidation and Ferroptosis Following Radiotherapy
Recent work has demonstrated a clear connection between radiotherapy, lipid redox homeostasis, and ferroptosis. Biochemical studies show that multilamellar liposomes treated with therapeutically relevant doses of radiotherapy undergo peroxidation as well as lipid fragmentation, leading to the rupture of a model membrane [42]. In vitro studies have shown that absorption of radiotherapy by water leads to the formation of oxygen radicals, which subsequently attack PUFAs to cause lipid peroxidation [37,43]. Radiotherapy has been shown to increase lipid oxidation in a dose-dependent manner when quantified by ROS sensitive fluorescent dyes which localize to lipid membranes [5]. These studies demonstrate a clear link between radiotherapy and lipid peroxidation.
The consequence of excessive lipid oxidation is ferroptosis. Lang et al. found that radiotherapy treatment of ID8 ovarian cancer cells increases lipid oxidative damage by activating the ataxia-telangiectasia mutated gene (ATM) to suppress SLC7A11 expression. This resulted in loss of lipid redox homeostasis and initiation of ferroptosis. Inhibition of ATM rescued the cells from radiotherapy-induced ferroptosis [5]. Lei et al. showed that radiotherapy treatment of different non-small cell lung cancer (NSCLC) lines induced ROS production, lipid peroxidation, and increased ACSL4 expression. Deletion of ACSL4 inhibited ferroptosis by reducing lipid peroxidation [3]. Ye et al. provided functional evidence that administration of ferroptosis inducers improved the cell-killing effects of radiotherapy, both in fibrosarcoma cells in vitro and human patient murine xenografts of adenocarcinoma and glioma [4]. Shrunken mitochondria are a hallmark of ferroptosis, and Lei et al. demonstrated that cancer cells treated with ionizing radiotherapy were characterized by shrunken mitochondria, further suggesting a link between radiotherapy and ferroptosis induction [3]. Finally, increases in tumoral lipid oxidation following radiotherapy based neoadjuvant treatment in patients with esophageal cancer is associated with improved locoregional control and OS [3]. Taken together, these data suggest that radiotherapy promotes lipid oxidation and ferroptosis.
4. Therapeutic Opportunities to Enhance Radiotherapy Efficacy via Ferroptosis Induction
Ferroptotic sensitivity is dictated by the proportion of PUFA in the lipid membrane [44]. Interestingly, genetic perturbation of lipid composition modulates radiotherapy sensitivity. ACSL4 knockout, which results in diminished PUFA lipid synthesis, abolishes radiotherapy efficacy in vitro and in vivo [5]. Conversely, ACSL3 knockout, which limits monounsaturated fatty acid lipid synthesis in the cell membrane, augments radiotherapy efficacy [5]. Additionally, lipid metabolism regulates lipid membrane composition. Repeated radiotherapy exposure to fractionated radiotherapy can generate cancer cell lines resistant to radiation [45]. Work has shown that in cervical cancer cells, upregulation of MiR-7-5p promotes radiotherapy resistance by silencing arachidonate 12-lipoxygenase (ALOX12) and other components of ferroptosis signaling, thus limiting radiotherapy efficacy [45]. Together, these data suggest that lipid metabolism can be therapeutically targeted to improve radiotherapy efficacy.
Cystine import is critical for glutathione biosynthesis and maintenance of the antioxidant pool within the cell. Upregulation of SLC family members that regulate cystine import has been tied to acquired radiotherapy resistance in vitro [46]. Recombinant enzymes that degrade cysteine and cystine promote radiotherapy sensitivity of melanoma and ovarian tumors in vivo [5]. Further, pharmacologic inhibitors of SLC7A11, the critical antiporter responsible for cystine uptake, increase radiotherapy sensitivity [4]. Erastin, an SLC7A11 inhibitor, sensitizes radiotherapy resistant NSCLC cancer lines [47]. Another SLC7A11 inhibitor, sulfasalazine, has been shown to promote radiotherapy efficacy in cell line xenograft and patient derived xenograft murine models of lung cancer. Interestingly, sulfasalazine alone did not impact the size of tumors in the absence of radiation in this study [3]. Further, sorafenib, a tyrosine kinase inhibitor that inhibits SLC7A11, sensitizes fibrosarcoma and lung adenocarcinoma xenografts to radiotherapy by increasing lipid peroxidation without increasing the DNA damage profile compared to radiation alone [4]. Radiation efficacy was shown to synergize with direct GPX4 inhibition via RSL-3 to diminish clonogenic survival in multiple models when compared to radiation or RSL-3 alone. This work provides functional evidence that administration of ferroptosis inducers improves radiotherapy efficacy in different contexts, including fibrosarcoma cells in vitro, murine xenografts of lung adenocarcinoma and fibrosarcoma in vivo, and glioma patient derived slice cultures ex vivo [4].
Corroborating this preclinical data, high NRF2 and SLC7A11 expression has been associated with diminished radiotherapy induced ferroptosis and decreased lipid oxidation, as well as radiotherapy resistance in patients with esophageal cancer [48]. In head and neck cancer, treatment with artesunate to inhibit NRF2 increased ferroptosis in cancer cells [49]. KRAS, an oncogene mutated in approximately 25% of human cancers, regulates NRF2 signaling [50]. In pancreatic cancer, KRAS signaling upregulating NRF2 led to chemoresistance via increases in glutaminolysis [51]. KRAS mutant lung cancer cells showed resistance to erastin-induced ferroptosis [52]. This suggests that KRAS can promote cancer resistance to ferroptosis by signaling through NRF2 to upregulate system xc−. Together, these data highlight that cystine transporters are a novel and targetable mechanism to augment radiotherapy efficacy.
Iron metabolism and oxidation have been tied not only to ferroptosis but also radiotherapy efficacy. Holo-Lactoferrin is a radiosensitizer which increases total iron content, promotes ROS, and facilitates lipid oxidation to enhance radiotherapy efficacy through ferroptosis [53]. Depletion of mitochondrial antioxidants including Coenzyme Q with FIN56, a known ferroptosis inducer, diminishes cancer cell survival following radiotherapy. The glycoprotein collectrin improves radiotherapy sensitivity through ferroptosis induction in the setting of hepatocellular carcinoma [54].
5. Therapeutic Opportunities to Limit Radiotherapy Toxicity via Ferroptosis Inhibition
Normal tissue toxicity following radiotherapy can cause significant morbidity in cancer patients. Technological advances in radiotherapy planning and delivery have led to striking decreases in patient adverse events. However, for patients in which large treatment fields or ablative doses are required for tumor control, radiotherapy can produce significant normal tissue toxicity in adjacent, healthy organs. Focal administration of therapeutic doses of radiotherapy can induce a hyperinflammatory cytokine response via the release of cardiolipin and phosphatidylserine lipid oxidation in the lung [55] as well as malondialdehyde lipid oxidation in the liver [56]. In preclinical models receiving thoracic irradiation, ferroptosis inhibitors limit cytokine release following radiotherapy, thereby decreasing inflammation and reducing lung fibrosis [57,58]. Thus, radiotherapy-induced ferroptosis may contribute to late effects following radiotherapy in slow-growing tissues.
Exposure to whole body radiation in the setting of extraterrestrial travel or incidental radiation exposure can be potentially lethal. Multiorgan dysfunction contributes to morbidity and mortality in this setting, including gastrointestinal and hematopoietic injuries [59]. Additionally, multiple cancers required low dose total body irradiation (TBI) as adjuvant or ablative treatment, and this also has associated toxicities [60]. It has long been understood that organs with rapidly dividing cells, including the intestinal epithelium and hematopoietic precursors, respond acutely to radiotherapy. It has been suggested that total body irradiation increases DNA damage and antioxidant responses to induce normal tissue toxicity [61]. Interestingly, TBI has been shown to increase bone marrow lipid oxidation and decrease bone marrow Vitamin E, a lipophilic antioxidant, in a dose-dependent manner [62]. This suggests that ferroptosis may contribute to total body irradiation toxicity and may also contribute to acute effects following radiotherapy treatment in rapidly dividing tissues.
Consistent with this, polycysteine derivatives, which increase the antioxidant pool and promote GPX4 activity, limit lethal whole body radiotherapy toxicity in preclinical models. Further, treatment with polycysteine derivatives limited gastrointestinal and hematopoietic toxicity as well as radiotherapy induced lung disease in murine models. Mechanistic studies showed that polycysteine diminished lipid oxidation and restored the GSH pool following radiotherapy [63]. Yet another molecule, arachidonate-15-lipoxygenase-1 (ALOX15), promotes lipid oxidation and contributes to the induction of ferroptosis [64]. Baicalein, an ALOX15 inhibitor, has been shown to normalize inflammatory cytokines induced by total body irradiation and improves the survival of mice treated with TBI even when administered post radiation exposure [65]. Collectively, these data suggest that ferroptosis contributes to normal tissue toxicity after radiation exposure and suggests that manipulation of ferroptotic induction may provide radioprotection for healthy tissues. At present, lipophilic antioxidant ferroptosis inhibitors include liproxstatin-1 and ferrostatin-1, which are currently unsuitable for in vivo administration, although medicinal chemistry approaches are improving the pharmacodynamics of these compounds [66].
6. Conclusions and Future Directions
There is now compelling literature that radiotherapy induces lipid oxidation and ferroptosis in tumors. This adds to the growing body of evidence that radiotherapy can provide tumor control by inducing programmed forms of cell death, including apoptosis [67,68], necroptosis [69], autophagy [70], and now ferroptosis. Additionally, radiotherapy can induce unregulated forms of tumor cell death including mitotic catastrophe, necrosis, and senescence [71,72]. Recent work has demonstrated that stress granules are associated with cancer chemoresistance and may provide a link between ferroptosis and radioresistance [73]. Additional work is required to understand whether radiotherapy dose, fractionation, sequencing, and source as well as cancer type and tumor microenvironment alter the relative contribution of each form of cell death following radiotherapy.
The radiosensitization agents currently most clinically utilized include platinum compounds, alkylating agents, inhibitors of DNA synthesis, and topoisomerase inhibitors [74]. Emerging strategies for radiosensitization include targeting the DNA damage response [75]. However, these strategies converge on augmenting DNA damage. Ferroptosis inducers have been shown to sensitize preclinical cancer models to radiotherapy, suggesting that augmentation of lipid damage may offer a novel therapeutic target that may prove to be an invaluable addition to the anticancer therapeutic armamentarium. Importantly, studies suggest that ferroptosis following radiotherapy is independent of DNA damage [4]. As the DNA damage response and lipid oxidation both induce a cellular stress response with common molecular modulators including p53 and ATM [76], additional work is required to understand the nature of cross talk between DNA and lipid oxidative damage following radiotherapy. Additional studies are also required to establish the optimal ferroptotic agent, administration schedule, and dose to advance to clinical trials in combination with radiotherapy (Table 1).
Radiation damage to adjacent healthy organs and tissues can cause significant morbidity in cancer patients. This radiotherapy toxicity in normal tissue may also rely on lipid oxidation and ferroptosis. Additional studies are required to understand whether targeting ferroptosis to augment radiotherapy efficacy widens or narrows the therapeutic index.
Author Contributions
A.N.P.: investigation, writing—original draft preparation, visualization, revised draft preparation. L.J.: writing—reviewing and editing. J.C.: writing reviewing and editing. Y.L.L.: writing—review and editing. M.D.G.: conceptualization, investigation, writing—original draft preparation, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by R01 DE026728 to Y.L.L as well as R21 CA252010 and I01 BX005267 to M.D.G.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thariat, J.; Hannoun-Levi, J.-M.; Myint, A.S.; Vuong, T.; Gérard, J.-P. Past, present, and future of radiotherapy for the benefit of patients. Nat. Rev. Clin. Oncol. 2013, 10, 52–60. [Google Scholar] [CrossRef]
- Adjemian, S.; Oltean, T.; Martens, S.; Wiernicki, B.; Goossens, V.; Berghe, T.V.; Cappe, B.; Ladik, M.; Riquet, F.B.; Heyndrickx, L.; et al. Ionizing radiation results in a mixture of cellular outcomes including mitotic catastrophe, senescence, methuosis, and iron-dependent cell death. Cell Death Dis. 2020, 11, 1003. [Google Scholar] [CrossRef]
- Lei, G.; Zhang, Y.; Koppula, P.; Liu, X.; Zhang, J.; Lin, S.H.; Ajani, J.A.; Xiao, Q.; Liao, Z.; Wang, H.; et al. The role of ferroptosis in ionizing radiation-induced cell death and tumor suppression. Cell Res. 2020, 30, 146–162. [Google Scholar] [CrossRef]
- Ye, L.F.; Chaudhary, K.R.; Zandkarimi, F.; Harken, A.D.; Kinslow, C.J.; Upadhyayula, P.S.; Dovas, A.; Higgins, D.M.; Tan, H.; Zhang, Y.; et al. Radiation-Induced Lipid Peroxidation Triggers Ferroptosis and Synergizes with Ferroptosis Inducers. ACS Chem. Biol. 2020, 15, 469–484. [Google Scholar] [CrossRef]
- Lang, X.; Green, M.D.; Wang, W.; Yu, J.; Choi, J.E.; Jiang, L.; Liao, P.; Zhou, J.; Zhang, Q.; Dow, A.; et al. Radiotherapy and Immunotherapy Promote Tumoral Lipid Oxidation and Ferroptosis via Synergistic Repression of SLC7A11. Cancer Discov. 2019, 9, 1673–1685. [Google Scholar] [CrossRef] [Green Version]
- Łuczaj, W.; Gęgotek, A.; Skrzydlewska, E. Antioxidants and HNE in redox homeostasis. Free Radic. Biol. Med. 2017, 111, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhorn, T.; Stockwell, B.R. The development of the concept of ferroptosis. Free Radic. Biol. Med. 2019, 133, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Comporti, M.; Saccocci, C.; Dianzani, M.U. Effect of CCl-4 in vitro and in vivo on lipid peroxidation of rat liver homogenates and subcellular fractions. Enzymologia 1965, 29, 185–204. [Google Scholar]
- Ghoshal, A.K.; Recknagel, R.O. Positive evidence of acceleration of lipoperoxidation in rat liver by carbon tetrachloride: In Vitro experiments. Life Sci. 1965, 4, 1521–1530. [Google Scholar] [CrossRef]
- Zheng, J.; Conrad, M. The Metabolic Underpinnings of Ferroptosis. Cell Metab. 2020, 32, 920–937. [Google Scholar] [CrossRef]
- Feng, H.; Stockwell, B.R. Unsolved mysteries: How does lipid peroxidation cause ferroptosis? PLoS Biol. 2018, 16, e2006203. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.R.; Angeli, J.P.F.; Bayir, H.; Bush, A.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Wiernicki, B.; Dubois, H.; Tyurina, Y.Y.; Hassannia, B.; Bayir, H.; Kagan, V.E.; Vandenabeele, P.; Wullaert, A.; Berghe, T.V. Excessive phospholipid peroxidation distinguishes ferroptosis from other cell death modes including pyroptosis. Cell Death Dis. 2020, 11, 922. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Kim, K.J.; Gaschler, M.M.; Patel, M.; Shchepinov, M.S.; Stockwell, B.R. Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc. Natl. Acad. Sci. USA 2016, 113, E4966–E4975. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Henry, W.S.; Eaton, J.K.; Ferguson, B.; Wang, W.; Fairman, J.; Keys, H.R.; Dančík, V.; Clish, C.B.; Clemons, P.A.; et al. Plasticity of ether lipids promotes ferroptosis susceptibility and evasion. Nature 2020, 585, 603–608. [Google Scholar] [CrossRef]
- Doll, S.; Proneth, B.; Tyurina, Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, M.I.J.; Aichler, M.; Walch, M.A.A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Y. The interaction between ferroptosis and lipid metabolism in cancer. Signal Transduct. Target. Ther. 2020, 5, 108. [Google Scholar] [CrossRef] [PubMed]
- Magtanong, L.; Ko, P.-J.; To, M.; Cao, J.Y.; Forcina, G.C.; Tarangelo, A.; Ward, C.C.; Cho, K.; Patti, G.J.; Nomura, D.K.; et al. Exogenous Monounsaturated Fatty Acids Promote a Ferroptosis-Resistant Cell State. Cell Chem. Biol. 2019, 26, 420–432.e9. [Google Scholar] [CrossRef]
- Gao, M.; Yi, J.; Zhu, J.; Minikes, A.; Monian, P.; Thompson, C.B.; Jiang, X. Role of Mitochondria in Ferroptosis. Mol. Cell 2019, 73, 354–363.e3. [Google Scholar] [CrossRef] [Green Version]
- Cozza, G.; Rossetto, M.; Bosello-Travain, V.; Maiorino, M.; Roveri, A.; Toppo, S.; Zaccarin, M.; Zennaro, L.; Ursini, F. Glutathione peroxidase 4-catalyzed reduction of lipid hydroperoxides in membranes: The polar head of membrane phospholipids binds the enzyme and addresses the fatty acid hydroperoxide group toward the redox center. Free Radic. Biol. Med. 2017, 112, 1–11. [Google Scholar] [CrossRef]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of Ferroptotic Cancer Cell Death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeli, J.P.F.; Schneider, M.; Basavarajappa, D.; Rådmark, O.; Kobayashi, S.; Seibt, T.; Beck, H.; Neff, F.; Esposito, I.; Wanke, R.; et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nature 2014, 16, 1180–1191. [Google Scholar] [CrossRef] [Green Version]
- Doll, S.; Freitas, F.P.; Shah, R.; Aldrovandi, M.; Da, S.M.; Ingold, I.; Goya, G.A.; Xavier, D.S.T.; Panzilius, E.; Scheel, C.H.; et al. FSP1 is a glutathione-independent ferroptosis suppressor. Nature 2019, 575, 693–698. [Google Scholar] [CrossRef]
- Kraft, V.A.; Bezjian, C.T.; Pfeiffer, S.; Ringelstetter, L.; Müller, C.; Zandkarimi, F.; Merl-Pham, J.; Bao, X.; Anastasov, N.; Kössl, J.; et al. GTP cyclohydrolase 1/tetrahydrobiopterin counteract ferroptosis through lipid remodeling. ACS Cent. Sci. 2019, 6, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Soula, M.; Weber, R.A.; Zilka, O.; Alwaseem, H.; La, K.; Yen, F.; Molina, H.; Garcia-Bermudez, J.; Pratt, D.A.; Birsoy, K. Metabolic determinants of cancer cell sensitivity to canonical ferroptosis inducers. Nat. Chem. Biol. 2020, 16, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Eagle, H. Utilization of Dipeptides by Mammalian Cells in Tissue Culture. Exp. Biol. Med. 1955, 89, 96–99. [Google Scholar] [CrossRef]
- Badgley, M.A.; Kremer, D.M.; Maurer, H.C.; DelGiorno, K.E.; Lee, H.-J.; Purohit, V.; Sagalovskiy, I.R.; Ma, A.; Kapilian, J.; Firl, C.E.M.; et al. Cysteine depletion induces pancreatic tumor ferroptosis in mice. Science 2020, 368, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Koppula, P.; Zhuang, L.; Gan, B. Cystine transporter SLC7A11/xCT in cancer: Ferroptosis, nutrient dependency, and cancer therapy. Protein Cell 2021, 12, 599–620. [Google Scholar] [CrossRef]
- Sato, H.; Tamba, M.; Ishii, T.; Bannai, S. Cloning and Expression of a Plasma Membrane Cystine/Glutamate Exchange Transporter Composed of Two Distinct Proteins. J. Biol. Chem. 1999, 274, 11455–11458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Kon, N.; Li, T.; Wang, S.-J.; Su, T.; Hibshoosh, H.; Baer, R.; Gu, W. Ferroptosis as a p53-mediated activity during tumour suppression. Nature 2015, 520, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Green, M.; Xia, H.; Zhou, J.; Li, G.; Li, J.; Li, W.; Wei, S.; Vatan, L.; Zhang, H.; et al. CD8+ T cells regulate tumour ferroptosis during cancer immunotherapy. Nature 2019, 569, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Wirth, A.-K.; Chen, D.; Wruck, C.J.; Rauh, M.; Buchfelder, M.; Savaskan, N. Nrf2-Keap1 pathway promotes cell proliferation and diminishes ferroptosis. Oncogenesis 2017, 6, e371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leone, R.D.; Zhao, L.; Englert, J.M.; Sun, I.-M.; Oh, M.-H.; Arwood, M.L.; Bettencourt, I.A.; Patel, C.H.; Wen, J.; Tam, A.; et al. Glutamine blockade induces divergent metabolic programs to overcome tumor immune evasion. Science 2019, 366, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Monian, P.; Quadri, N.; Ramasamy, R.; Jiang, X. Glutaminolysis and Transferrin Regulate Ferroptosis. Mol. Cell 2015, 59, 298–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rössler, O.G.; Bauer, I.; Chung, H.Y.; Thiel, G. Glutamate-induced cell death of immortalized murine hippocampal neurons: Neuroprotective activity of heme oxygenase-1, heat shock protein 70, and sodium selenite. Neurosci. Lett. 2004, 362, 253–257. [Google Scholar] [CrossRef]
- Su, L.-J.; Zhang, J.-H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.-Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [Green Version]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef]
- Zou, Y.; Li, H.; Graham, E.T.; Deik, A.A.; Eaton, J.K.; Wang, W.; Sandoval-Gomez, G.; Clish, C.B.; Doench, J.G.; Schreiber, S.L. Cytochrome P450 oxidoreductase contributes to phospholipid peroxidation in ferroptosis. Nat. Chem. Biol. 2020, 16, 302–309. [Google Scholar] [CrossRef]
- Wenzel, S.E.; Tyurina, Y.; Zhao, J.; Croix, C.M.S.; Dar, H.H.; Mao, G.; Tyurin, V.; Anthonymuthu, T.S.; Kapralov, O.; Amoscato, A.A.; et al. PEBP1 Wardens Ferroptosis by Enabling Lipoxygenase Generation of Lipid Death Signals. Cell 2017, 171, 628–641.e26. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Yu, C.; Kang, R.; Tang, D. Iron Metabolism in Ferroptosis. Front. Cell Dev. Biol. 2020, 8, 590226. [Google Scholar] [CrossRef]
- Shadyro, O.I.; Yurkova, I.L.; Kisel, M.A. Radiation-induced peroxidation and fragmentation of lipids in a model membrane. Int. J. Radiat. Biol. 2002, 78, 211–217. [Google Scholar] [CrossRef]
- Azzam, E.I.; Jay-Gerin, J.-P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, G.; Mao, C.; Yan, Y.; Zhuang, L.; Gan, B. Ferroptosis, radiotherapy, and combination therapeutic strategies. Protein Cell 2021, 12, 836–857. [Google Scholar] [CrossRef]
- Tomita, K.; Nagasawa, T.; Kuwahara, Y.; Torii, S.; Igarashi, K.; Roudkenar, M.; Roushandeh, A.; Kurimasa, A.; Sato, T. MiR-7-5p Is Involved in Ferroptosis Signaling and Radioresistance Thru the Generation of ROS in Radioresistant HeLa and SAS Cell Lines. Int. J. Mol. Sci. 2021, 22, 8300. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Song, X.; Yu, J.; Guo, W.; Wei, L.; Liu, Y.; Wang, X. Solute carrier protein family may involve in radiation-induced radioresistance of non-small cell lung cancer. J. Cancer Res. Clin. Oncol. 2011, 137, 1739–1747. [Google Scholar] [CrossRef]
- Pan, X.; Lin, Z.; Jiang, D.; Yu, Y.; Yang, D.; Zhou, H.; Zhan, D.; Liu, S.; Peng, G.; Chen, Z.; et al. Erastin decreases radioresistance of NSCLC cells partially by inducing GPX4-mediated ferroptosis. Oncol. Lett. 2019, 17, 3001–3008. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Zhao, K.; Sun, L.; Yin, X.; Zhang, J.; Liu, C.; Li, B. SLC7A11 regulated by NRF2 modulates esophageal squamous cell carcinoma radiosensitivity by inhibiting ferroptosis. J. Transl. Med. 2021, 19, 367. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.-L.; Kim, E.H.; Jang, H.; Shin, D. Nrf2 inhibition reverses the resistance of cisplatin-resistant head and neck cancer cells to artesunate-induced ferroptosis. Redox Biol. 2017, 11, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Wang, S.; Moghaddam, S.J.; Ooi, A.; Chapman, E.; Wong, P.K.; Zhang, D.D. Oncogenic KRAS Confers Chemoresistance by Upregulating NRF2. Cancer Res. 2014, 74, 7430–7441. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Goswami, D.; Adiseshaiah, P.P.; Burgan, W.; Yi, M.; Guerin, T.M.; Kozlov, S.V.; Nissley, D.V.; McCormick, F. Undermining Glutaminolysis Bolsters Chemotherapy While NRF2 Promotes Chemoresistance in KRAS-Driven Pancreatic Cancers. Cancer Res. 2020, 80, 1630–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, B.H.; Colon, T.; Jiang, J.; Lee, E.; Dai, W. Regulation of ferroptosis in KRas mutant cancer cells. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, M.; Chen, C.; Tong, X.; Li, Y.; Yang, K.; Lv, H.; Xu, J.; Qin, L. Holo-lactoferrin: The link between ferroptosis and radiotherapy in triple-negative breast cancer. Theranostics 2021, 11, 3167–3182. [Google Scholar] [CrossRef]
- Yuan, Y.; Cao, W.; Zhou, H.; Qian, H.; Wang, H. CLTRN, Regulated by NRF1/RAN/DLD Protein Complex, Enhances Radiation Sensitivity of Hepatocellular Carcinoma Cells through Ferroptosis Pathway. Int. J. Radiat. Oncol. 2021, 110, 859–871. [Google Scholar] [CrossRef]
- Tyurina, Y.Y.; Tyurin, V.A.; Kapralova, V.I.; Wasserloos, K.; Mosher, M.; Epperly, M.W.; Greenberger, J.S.; Pitt, B.R.; Kagan, V.E. Oxidative Lipidomics of γ-Radiation-Induced Lung Injury: Mass Spectrometric Characterization of Cardiolipin and Phosphatidylserine Peroxidation. Radiat. Res. 2011, 175, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Ueda, T.; Toyoshima, Y.; Kushihashi, T.; Hishida, T.; Yasuhara, H. Effect of dimethyl sulfoxide pretreatment on activities of lipid peroxide formation, superoxide dismutase and glutathione peroxidase in the mouse liver after whole-body irradiation. J. Toxicol. Sci. 1993, 18, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Duan, L.; Yuan, S.; Zhuang, X.; Qiao, T.; He, J. Ferroptosis inhibitor alleviates Radiation-induced lung fibrosis (RILF) via down-regulation of TGF-β1. J. Inflamm. 2019, 16, 11. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhuang, X.; Qiao, T. Role of ferroptosis in the process of acute radiation-induced lung injury in mice. Biochem. Biophys. Res. Commun. 2019, 519, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Stewart, F.A.; Akleyev, A.; Hauerjensen, M.; Hendry, J.; Kleiman, N.; MacVittie, T.; Aleman, B.; Edgar, A.; Mabuchi, K.; Muirhead, C.; et al. ICRP Publication 118: ICRP Statement on Tissue Reactions and Early and Late Effects of Radiation in Normal Tissues and Organs—Threshold Doses for Tissue Reactions in a Radiation Protection Context. Ann. ICRP 2012, 41, 1–322. [Google Scholar] [CrossRef]
- Vogel, J.; Hui, S.; Hua, C.-H.; Dusenbery, K.; Rassiah, P.; Kalapurakal, J.; Constine, L.; Esiashvili, N. Pulmonary Toxicity after Total Body Irradiation—Critical Review of the Literature and Recommendations for Toxicity Reporting. Front. Oncol. 2021, 11, 708906. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Inaba, Y.; Sogo, Y.; Ito, A.; Bekal, M.; Chida, K.; Moritake, T. Total body irradiation causes a chronic decrease in antioxidant levels. Sci. Rep. 2021, 11, 6716. [Google Scholar] [CrossRef]
- Umegaki, K.; Ichikawa, T. Decrease in vitamin E levels in the bone marrow of mice receiving whole-body X-ray irradiation. Free Radic. Biol. Med. 1994, 17, 439–444. [Google Scholar] [CrossRef]
- Zhang, J.; Li, K.; Zhang, Q.; Zhu, Z.; Huang, G.; Tian, H. Polycysteine as a new type of radio-protector ameliorated tissue injury through inhibiting ferroptosis in mice. Cell Death Dis. 2021, 12, 195. [Google Scholar] [CrossRef]
- Shintoku, R.; Takigawa, Y.; Yamada, K.; Kubota, C.; Yoshimoto, Y.; Takeuchi, T.; Koshiishi, I.; Torii, S. Lipoxygenase-mediated generation of lipid peroxides enhances ferroptosis induced by erastin and RSL3. Cancer Sci. 2017, 108, 2187–2194. [Google Scholar] [CrossRef] [PubMed]
- Thermozier, S.; Hou, W.; Zhang, X.; Shields, D.; Fisher, R.; Bayir, H.; Kagan, V.; Yu, J.; Liu, B.; Bahar, I.; et al. Anti-Ferroptosis Drug Enhances Total-Body Irradiation Mitigation by Drugs that Block Apoptosis and Necroptosis. Radiat. Res. 2020, 193, 435. [Google Scholar] [CrossRef]
- Devisscher, L.; Van Coillie, S.; Hofmans, S.; Van Rompaey, D.; Goossens, K.; Meul, E.; Maes, L.; De Winter, H.; Van Der Veken, P.; Vandenabeele, P.; et al. Discovery of Novel, Drug-Like Ferroptosis Inhibitors with In Vivo Efficacy. J. Med. Chem. 2018, 61, 10126–10140. [Google Scholar] [CrossRef]
- Dewey, W.C.; Ling, C.C.; Meyn, R.E. Radiation-induced apoptosis: Relevance to radiotherapy. Int. J. Radiat. Oncol. 1995, 33, 781–796. [Google Scholar] [CrossRef]
- Verheij, M.; Bartelink, H. Radiation-induced apoptosis. Cell Tissue Res. 2000, 301, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Nehs, M.A.; Lin, C.-I.; Kozono, D.E.; Whang, E.E.; Cho, N.L.; Zhu, K.; Moalem, J.; Moore, F.D.; Ruan, D.T. Necroptosis is a novel mechanism of radiation-induced cell death in anaplastic thyroid and adrenocortical cancers. Surgery 2011, 150, 1032–1039. [Google Scholar] [CrossRef]
- Chaurasia, M.; Bhatt, A.N.; Das, A.; Dwarakanath, B.S.; Sharma, K. Radiation-induced autophagy: Mechanisms and consequences. Free Radic. Res. 2016, 50, 273–290. [Google Scholar] [CrossRef]
- Flor, A.C.; Doshi, A.P.; Kron, S.J. Modulation of therapy-induced senescence by reactive lipid aldehydes. Cell Death Discov. 2016, 2, 16045. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.; Giacci, A. Radiobiology for the Radiologist; Wolters Kluwer: Alphen aan den Rijn, The Netherlands, 2018. [Google Scholar]
- Mao, C.; Wang, X.; Liu, Y.; Wang, M.; Yan, B.; Jiang, Y.; Shi, Y.; Shen, Y.; Liu, X.; Liai, W.; et al. A G3BP1-interacting lncRNA promotes ferroptosis and apoptosis in cancer via nuclear sequestration of p53. Cancer Res. 2018, 78, 3484–3496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, T.S.; Blackstock, A.; McGinn, C. The mechanism of action of radiosensitization of conventional chemotherapeutic agents. Semin. Radiat. Oncol. 2003, 13, 13–21. [Google Scholar] [CrossRef]
- Parsels, L.A.; Zhang, Q.; Karnak, D.; Parsels, J.D.; Lam, K.; Willers, H.; Green, M.D.; Rehemtulla, A.; Lawrence, T.S.; Morgan, M.A. Translation of DNA Damage Response Inhibitors as Chemoradiation Sensitizers from the Laboratory to the Clinic. Int. J. Radiat. Oncol. 2021, 111, e38–e53. [Google Scholar] [CrossRef]
- Chen, P.-H.; Tseng, W.H.-S.; Chi, J.-T. The Intersection of DNA Damage Response and Ferroptosis—A Rationale for Combination Therapeutics. Biology 2020, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Scott Wilhelm, C.C.; Lynch, M.; Lowinger, T.; Dumas, J.; Smith, R.A.; Schwartz, B.; Simantov, R.; Kelley, S. Discovery and development of sorafenib: A multikinase inhibitor for treating cancer. Nat. Rev. Drug Discov. 2006, 5, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Taffet, S.L.; Das, K.M. Sulfasalazine. Dig. Dis. Sci. 1983, 28, 833–842. [Google Scholar] [CrossRef]
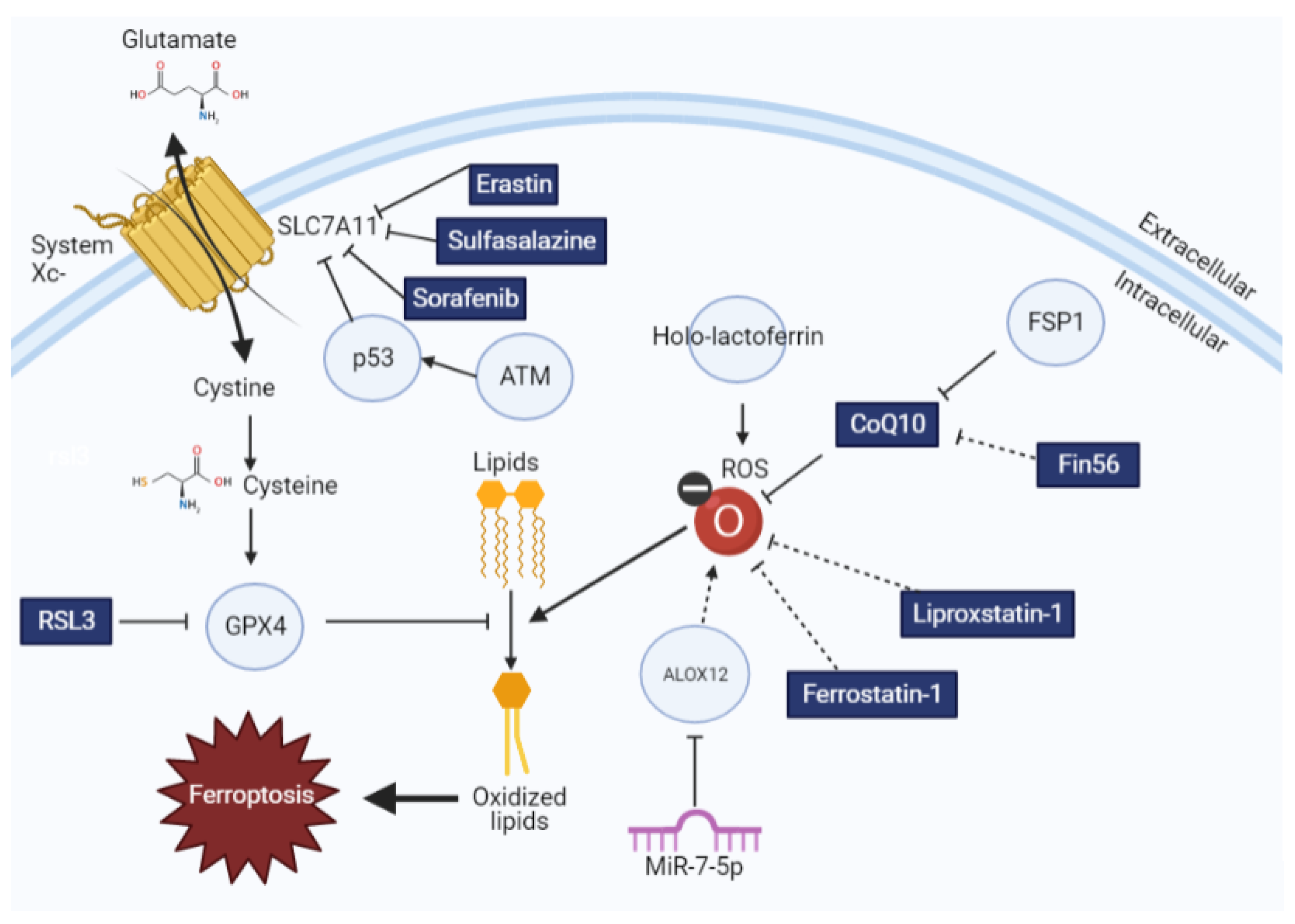
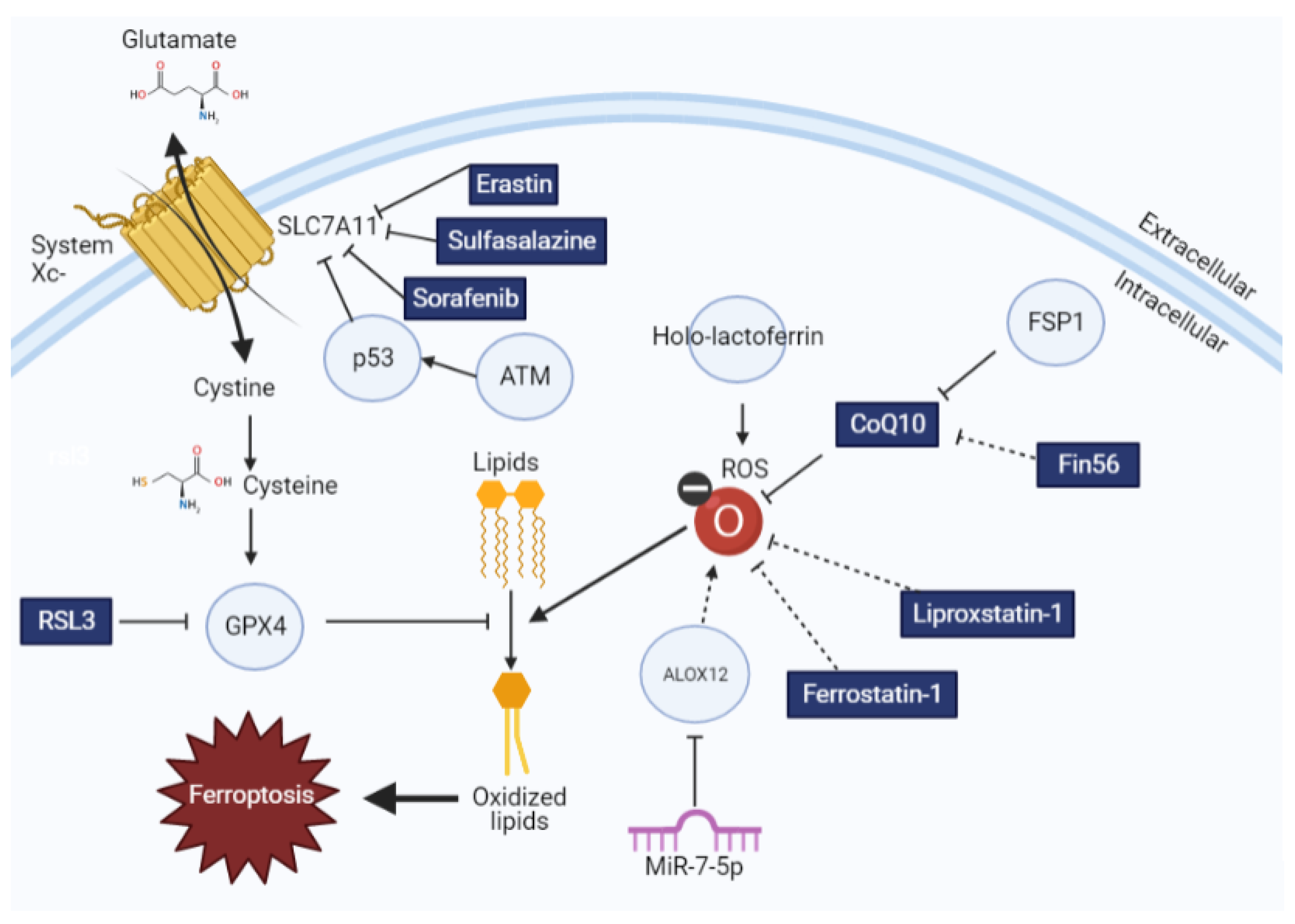
Figure 1.
The impact of ferroptotic inducers and inhibitors on lipid redox homeostasis. Glutathione synthesis is regulated by cystine import by system xc−, which is then used by GPX4 to oppose lipid oxidation. This process can be modulated by different ferroptosis inducers and inhibitors, as explained here. Drugs and small molecules are indicated in rectangles, whereas proteins are indicated with circles. Solid arrows indicate direct interactions, whereas dotted arrows indicate correlative interactions.
Figure 1.
The impact of ferroptotic inducers and inhibitors on lipid redox homeostasis. Glutathione synthesis is regulated by cystine import by system xc−, which is then used by GPX4 to oppose lipid oxidation. This process can be modulated by different ferroptosis inducers and inhibitors, as explained here. Drugs and small molecules are indicated in rectangles, whereas proteins are indicated with circles. Solid arrows indicate direct interactions, whereas dotted arrows indicate correlative interactions.

{kind=link}
Table 1.
Current clinical trials of drugs targeting ferroptosis. Sorafenib and sulfasalazine, neither of which were originally developed for the treatment of ferroptosis, may serve as ferroptosis inhibitors due to their ability to decrease the activity of SLC7A11 [77,78].
| Drug Name | Relevant Dates | Original Target | Current Number of Ongoing Clinical Trials | Ferroptotic Target |
|---|---|---|---|---|
| Sorafenib | 2000: Entered clinical trials | MAPK Cascade in Cancer | 92 | SLC7A11 |
| Sulfasalazine | 1950: Approved for clinical use | Rheumatoid arthritis | 13 | SLC7A11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pearson, A.N.; Carmicheal, J.; Jiang, L.; Lei, Y.L.; Green, M.D. Contribution of Lipid Oxidation and Ferroptosis to Radiotherapy Efficacy. Int. J. Mol. Sci. 2021, 22, 12603. https://doi.org/10.3390/ijms222212603
AMA Style
Pearson AN, Carmicheal J, Jiang L, Lei YL, Green MD. Contribution of Lipid Oxidation and Ferroptosis to Radiotherapy Efficacy. International Journal of Molecular Sciences. 2021; 22(22):12603. https://doi.org/10.3390/ijms222212603
Chicago/Turabian StylePearson, Ashley N., Joseph Carmicheal, Long Jiang, Yu Leo Lei, and Michael D. Green. 2021. "Contribution of Lipid Oxidation and Ferroptosis to Radiotherapy Efficacy" International Journal of Molecular Sciences 22, no. 22: 12603. https://doi.org/10.3390/ijms222212603
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.