Big Lessons from Tiny Flies: Drosophila melanogaster as a Model to Explore Dysfunction of Dopaminergic and Serotonergic Neurotransmitter Systems


Abstract
:1. Introduction
2. The Dopaminergic System
3. The Serotonergic System
4. Cross-Talk between the Dopaminergic and Serotonergic Systems
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bellen, H.J.; Yamamoto, S. Morgan’s Legacy: Fruit flies and the functional annotation of conserved genes. Cell 2015, 163, 12–14. [Google Scholar] [CrossRef] [PubMed]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef] [PubMed]
- Lenz, S.; Karsten, P.; Schulz, J.B.; Voigt, A. Drosophila as a screening tool to study human neurodegenerative diseases. J. Neurochem. 2013, 127, 453–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nüsslein-volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Jaiswal, M.; Charng, W.L.; Gambin, T.; Karaca, E.; Mirzaa, G.; Wiszniewski, W.; Sandoval, H.; Haelterman, N.A.; Xiong, B.; et al. A drosophila genetic resource of mutants to study mechanisms underlying human genetic diseases. Cell 2014, 159, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Rubin, G.; Spradling, A. Genetic transformation of Drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [PubMed]
- Perrimon, N. Drosophila Developmental Biology Methods. Methods 2014, 68, 1. [Google Scholar] [CrossRef] [PubMed]
- Hummel, T.; Klämbt, C. P-element mutagenesis. Methods Mol. Biol. 2008, 420, 97–117. [Google Scholar] [PubMed]
- Gratz, S.J.; Rubinstein, C.D.; Harrison, M.M.; Wildonger, J.; O’Connor-Giles, K.M. CRISPR-Cas9 genome editing in Drosophila. Curr. Protoc. Mol. Biol. 2015, 111, 31. [Google Scholar] [PubMed]
- Koundakjian, E.J.; Cowan, D.M.; Hardy, R.W.; Becker, A.H. The Zuker collection: A resource for the analysis of autosomal gene function in Drosophila melanogaster. Genetics 2004, 167, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Bauer, H.; Demerec, M.; Kaufmann, B.P. X-Ray induced chromosomal alterations in Drosophila Melanogaster. Genetics 1938, 23, 610–630. [Google Scholar] [PubMed]
- Berger, J.; Suzuki, T.; Senti, K.A.; Stubbs, J.; Schaffner, G.; Dickson, B.J. Genetic mapping with SNP markers in Drosophila. Nat. Genet. 2001, 29, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Blumenstiel, J.P.; Noll, A.C.; Griffiths, J.A.; Perera, A.G.; Walton, K.N.; Gilliland, W.D.; Hawley, R.S.; Staehling-Hampton, K. Identification of EMS-induced mutations in Drosophila melanogaster by whole-genome sequencing. Genetics 2009, 182, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.S.; Titen, S.W.; Xie, H.B.; Golic, M.M.; Bastiani, M.; Bandyopadhyay, P.; Olivera, B.M.; Brodsky, M.; Rubin, G.M.; Golic, K.G. Targeted mutagenesis by homologous recombination in D. melanogaster. Genes Dev. 2002, 16, 1568–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennerdell, J.R.; Carthew, R.W. Heritable gene silencing in Drosophila using double-stranded RNA. Nat. Biotechnol. 2000, 18, 896–898. [Google Scholar] [CrossRef] [PubMed]
- Port, F.; Chen, H.M.; Lee, T.; Bullock, S.L. Optimized CRISPR/Cas tools for efficient germline and somatic genome engineering in Drosophila. Proc. Natl. Acad. Sci. USA 2014, 111, E2967–E2976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Z.; Davis, R.L. Eight different types of dopaminergic neurons innervate the Drosophila mushroom body neuropil: Anatomical and physiological heterogeneity. Front. Neural Circuits 2009, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Schwaerzel, M.; Monastirioti, M.; Scholz, H.; Friggi-Grelin, F.; Birman, S.; Heisenberg, M. Dopamine and octopamine differentiate between aversive and appetitive olfactory memories in Drosophila. J. Neurosci. 2003, 23, 10495–10502. [Google Scholar] [CrossRef] [PubMed]
- Nitz, D.A.; van Swinderen, B.; Tononi, G.; Greenspan, R.J. Electrophysiological correlates of rest and activity in Drosophila melanogaster. Curr. Biol. 2002, 12, 1934–1940. [Google Scholar] [CrossRef]
- Alekseyenko, O.V.; Chan, Y.B.; Ran, L.; Kravitz, E.A. Single dopaminergic neurons that modulate aggression in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 10, 6151–6156. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, N.; Ichinose, T.; Aso, Y.; Plaçais, P.-Y.; Friedrich, A.B.; Sima, R.J.; Preat, T.; Rubin, G.M.; Tanimoto, H. Distinct dopamine neurons mediate reward signals for short- and long-term memories. Proc. Natl. Acad. Sci. USA 2015, 112, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Riemensperger, T.; Isabel, G.; Coulom, H.; Neuser, K.; Seugnet, L.; Kume, K.; Iche-Torres, M.; Cassar, M.; Strauss, R.; Preat, T.; et al. Behavioral consequences of dopamine deficiency in the Drosophila central nervous system. Proc. Natl. Acad. Sci. USA 2011, 108, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.A.; Cervantes-Sandoval, I.; Chakraborty, M.; Davis, R.L. Sleep facilitates memory by blocking dopamine neuron-mediated forgetting. Cell 2015, 161, 1656–1667. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Rubin, G.M. Dopaminergic neurons write and update memories with cell-type-specific rules. Elife 2016, 5, e16135. [Google Scholar] [CrossRef] [PubMed]
- Waddell, S. Reinforcement signalling in Drosophila; dopamine does it all after all. Curr. Opin. Neurobiol. 2013, 23, 324–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, R.W.; Akinleye, A.A.; Nurilov, M.; Feliciano, O.; Lollar, M.; Aijuri, R.R.; O’Donnell, J.M.; Simon, A.F. Modulation of social space by dopamine in Drosophila melanogaster, but no effect on the avoidance of the Drosophila stress odorant. Biol. Lett. 2017, 13, 20170369. [Google Scholar] [CrossRef] [PubMed]
- Hearn, M.G.; Ren, Y.; McBride, E.W.; Reveillaud, I.; Beinborn, M.; Kopin, A.S. A Drosophila dopamine 2-like receptor: Molecular characterization and identification of multiple alternatively spliced variants. Proc. Natl. Acad. Sci. USA 2002, 99, 14554–14559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimoto, H.; Wang, Z.; Rao, Y.; Wu, C.F.; Kitamoto, T. A Novel Role for Ecdysone in Drosophila Conditioned Behavior: Linking GPCR-Mediated Non-canonical Steroid Action to cAMP Signaling in the Adult Brain. PLoS Genet. 2013, 9, e1003843. [Google Scholar] [CrossRef] [PubMed]
- Neckameyer, W.S.; White, K. drosophila tyrosine hydroxylase is encoded by the pale locus. J. Neurogenet. 1993, 8, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Cichewicz, K.; Garren, E.J.; Adiele, C.; Aso, Y.; Wang, Z.; Wu, M.; Birman, S.; Rubin, G.M.; Hirsh, J. A new brain dopamine-deficient Drosophila and its pharmacological and genetic rescue. Genes Brain Behav. 2017, 16, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Dartevelle, L.; Yuan, C.; Wei, H.; Wang, Y.; Ferveur, J.F.; Guo, A. Increased Dopamine Level Enhances Male-Male Courtship in Drosophila. J. Neurosci. 2008, 28, 5539–5546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakely, R.D.; Edwards, R.H. Vesicular and plasma membrane transporters for neurotransmitters. Cold Spring Harb. Perspect. Biol. 2012, 4, a005595. [Google Scholar] [CrossRef] [PubMed]
- Greer, C.L.; Grygoruk, A.; Patton, D.E.; Ley, B.; Romero-Calderon, R.; Chang, H.Y.; Houshyar, R.; Bainton, R.J.; DiAntonio, A.; Krantz, D.E. A splice variant of the Drosophila vesicular monoamine transporter contains a conserved trafficking domain and functions in the storage of dopamine, serotonin, and octopamine. J. Neurobiol. 2005, 64, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Romero-Calderón, R.; Uhlenbrock, G.; Borycz, J.; Simon, A.F.; Grygoruk, A.; Yee, S.K.; Shyer, A.; Ackerson, L.C.; Maidment, N.T.; Meinertzhagen, I.A.; et al. A glial variant of the vesicular monoamine transporter is required to store histamine in the Drosophila visual system. PLoS Genet. 2008, 4, e1000245. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Ng, F.; Lebestky, T.; Grygoruk, A.; Djapri, C.; Lawal, H.O.; Zaveri, H.A.; Mehanzel, F.; Najibi, R.; Seidman, G.; et al. Dispensable, redundant, complementary, and cooperative roles of dopamine, octopamine, and serotonin in Drosophila melanogaster. Genetics 2013, 193, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Sitte, H.H.; Freissmuth, M. Amphetamines, new psychoactive drugs and the monoamine transporter cycle. Trends Pharmacol. Sci. 2015, 36, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Pörzgen, P.; Park, S.K.; Hirsh, J.; Sonders, M.S.; Amara, S.G. The antidepressant-sensitive dopamine transporter in Drosophila melanogaster: A primordial carrier for catecholamines. Mol. Pharmacol. 2001, 59, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Vickrey, T.L.; Condron, B.; Venton, B.J. Detection of endogenous dopamine changes in Drosophila melanogaster using fast-scan cyclic voltammetry. Anal. Chem. 2009, 81, 9306–9313. [Google Scholar] [CrossRef] [PubMed]
- Makos, M.A.; Han, K.A.; Heien, M.L.; Ewing, A.G. Using In Vivo Electrochemistry to Study the Physiological Effects of Cocaine and Other Stimulants on the Drosophila melanogaster Dopamine Transporter. ACS Chem. Neurosci 2010, 1, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, A.B.; Karam, C.S.; Zhang, Y.; Yano, H.; Freyberg, R.J.; Karam, D.S.; Freyberg, Z.; Yamamoto, A.; McCabe, B.D.; Javitch, J.A. The membrane raft protein Flotillin-1 is essential in dopamine neurons for amphetamine-induced behavior in Drosophila. Mol. Psychiatry 2013, 18, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, A.B.; Karam, C.S.; Zhang, Y.; Ma, C.L.; Mccabe, B.D.; Javitch, J.A. Amphetamine-induced behavior requires CaMKII-dependent dopamine transporter phosphorylation. Mol. Psychiatry 2014, 19, 279–281. [Google Scholar] [CrossRef] [PubMed]
- Sucic, S.; Dallinger, S.; Zdrazil, B.; Weissensteiner, R.; Jørgensen, T.N.; Holy, M.; Kudlacek, O.; Seidel, S.; Hwan Cha, J.; Gether, U.; et al. The N terminus of monoamine transporters is a lever required for the action of amphetamines. J. Biol. Chem. 2010, 285, 10924–10938. [Google Scholar] [CrossRef] [PubMed]
- Kern, C.; Erdem, F.A.; El-Kasaby, A.; Sandtner, W.; Freissmuth, M.; Sucic, S. The N terminus specifies the switch between transport modes of the human serotonin transporter. J. Biol. Chem. 2017, 292, 3603–3613. [Google Scholar] [CrossRef] [PubMed]
- Kasture, A.; El-Kasaby, A.; Szöllosi, D.; Mazhar Asjad, H.M.; Grimm, A.; Stockner, T.; Hummel, T.; Freissmuth, M.; Sucic, S. Functional rescue of a misfolded Drosophila melanogaster dopamine transporter mutant associated with a sleepless phenotype by pharmacological chaperones. J. Biol. Chem. 2016, 291, 20876–20890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.N.; Koh, K.; Yue, Z.; Joiner, W.J.; Sehgal, A. A genetic screen for sleep and circadian mutants reveals mechanisms underlying regulation of sleep in Drosophila. Sleep 2008, 31, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Kume, K.; Kume, S.; Park, S.K.; Hirsh, J.; Jackson, F.R. Dopamine Is a Regulator of Arousal in the Fruit Fly. J. Neurosci. 2005, 25, 7377–7384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devlin, B.; Boone, B.E.; Levy, S.E.; Lihm, J.; Buxbaum, J.D.; Wu, Y.; Lewis, L.; Han, Y.; Boerwinkle, E.; Gibbs, R.A.; et al. Patterns and rates of exonic de novo mutations in autism spectrum disorders. Nature 2012, 485, 242–246. [Google Scholar]
- Hamilton, P.J.; Campbell, N.G.; Sharma, S.; Erreger, K.; Herborg Hansen, F.; Saunders, C.; Belovich, A.N.; Sahai, M.A.; Cook, E.H.; Gether, U.; et al. De novo mutation in the dopamine transporter gene associates dopamine dysfunction with autism spectrum disorder. Mol. Psychiatry 2013, 18, 1315–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kasaby, A.; Koban, F.; Sitte, H.H.; Freissmuth, M.; Sucic, S. A cytosolic relay of heat shock proteins HSP70-1A and HSP90β Monitors the Folding trajectory of the serotonin transporter. J. Biol. Chem. 2014, 289, 28987–29000. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.T.; Zhang, Y.W.; Campbell, S.D.; Rudnick, G. Ibogaine, a noncompetitive inhibitor of serotonin transport, acts by stabilizing the cytoplasm-facing state of the transporter. J. Biol. Chem. 2007, 282, 29441–29447. [Google Scholar] [CrossRef] [PubMed]
- El-Kasaby, A.; Just, H.; Malle, E.; Stolt-Bergner, P.C.; Sitte, H.H.; Freissmuth, M.; Kudlacek, O. Mutations in the carboxyl-terminal SEC24 binding motif of the serotonin transporter impair folding of the transporter. J. Biol. Chem. 2010, 285, 39201–39210. [Google Scholar] [CrossRef] [PubMed]
- Sucic, S.; El-Kasaby, A.; Kudlacek, O.; Sarker, S.; Sitte, H.H.; Marin, P.; Freissmuth, M. The serotonin transporter is an exclusive client of the coat protein complex II (COPII) component SEC24C. J. Biol. Chem. 2011, 286, 16482–16490. [Google Scholar] [CrossRef] [PubMed]
- Sucic, S.; Koban, F.; El-Kasaby, A.; Kudlacek, O.; Stockner, T.; Sitte, H.H.; Freissmuth, M. Switching the clientele: A lysine residing in the c terminus of the serotonin transporter specifies its preference for the coat protein complex ii component SEC24C. J. Biol. Chem. 2013, 288, 5330–5341. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, T.R.; Steinkellner, T.; Sucic, S.; Koban, F.; Schuchner, S.; Ogris, E.; Sitte, H.H.; Freissmuth, M. Axonal targeting of the serotonin transporter in cultured rat dorsal raphe neurons is specified by SEC24C-dependent export from the endoplasmic reticulum. J. Neurosci. 2014, 34, 6344–6351. [Google Scholar] [CrossRef] [PubMed]
- Kurian, M.A.; Zhen, J.; Cheng, S.; Li, Y.; Mordekar, S.R.; Jardine, P.; Morgan, N.V.; Meyer, E.; Tee, L.; Pasha, S.; et al. Homozygous loss-of-function mutations in the gene encoding the dopamine transporter are associated with infantile parkinsonism-dystonia. J. Clin. Investig. 2009, 119, 1595–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurian, M.A.; Gissen, P.; Smith, M.; Heales, S.J.R.; Clayton, P.T. The monoamine neurotransmitter disorders : An expanding range of neurological syndromes. Lancet Neurol. 2011, 10, 721–733. [Google Scholar] [CrossRef]
- Ng, J.; Zhen, J.; Meyer, E.; Erreger, K.; Li, Y.; Kakar, N.; Ahmad, J.; Thiele, H.; Kubisch, C.; Rider, N.L.; et al. Dopamine transporter deficiency syndrome: Phenotypic spectrum from infancy to adulthood. Brain 2014, 137, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Barral, S.; Kurian, M.A. Utility of induced pluripotent stem cells for the study and treatment of genetic diseases: Focus on childhood neurological disorders. Front. Mol. Neurosci. 2016, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Freissmuth, M.; Stockner, T.; Sucic, S. SLC6 transporter folding diseases and pharmacochaperoning. Target. Traffick. Drug Dev. 2018, 245, 249–270. [Google Scholar]
- Kasture, A.; Stockner, T.; Freissmuth, M.; Sucic, S. An unfolding story: Small molecules remedy misfolded monoamine transporters. Int. J. Biochem. Cell Biol. 2017, 92, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sucic, S.; Kasture, A.; Asjad, H.M.M.; Kern, C.; El-kasaby, A.; Freissmuth, M. When transporters fail to be transported : How to rescue folding-deficient SLC6 transporters. J. Neurol. Neuromed. 2016, 1, 34–40. [Google Scholar]
- Mazhar Asjad, H.M.; Kasture, A.; El-Kasaby, A.; Sackel, M.; Hummel, T.; Freissmuth, M.; Sucic, S. Pharmacochaperoning in a Drosophila model system rescues human dopamine transporter variants associated with infantile/juvenile parkinsonism. J. Biol. Chem. 2017, 292, 19250–19265. [Google Scholar] [CrossRef] [PubMed]
- Beerepoot, P.; Lam, V.M.; Salahpour, A. Pharmacological chaperones of the dopamine transporter rescue dopamine transporter deficiency syndrome mutations in heterologous cells. J. Biol. Chem. 2016, 291, 22053–22062. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.; Hasenhuetl, P.S.; Kasture, A.; Ali, E.K.; Baumann, M.H.; Blough, B.E.; Sucic, S.; Sandtner, W.; Freissmuth, M. Conformational state interactions provide clues to the pharmacochaperone potential of serotonin transporter partial substrates. J. Biol. Chem. 2017, 292, 16773–16786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulin, J.F.; Zou, J.; Drouin-Ouellet, J.; Kim, K.Y.A.; Cicchetti, F.; Awatramani, R.B. Defining midbrain dopaminergic neuron diversity by single-cell gene expression profiling. Cell Rep. 2014, 9, 930–943. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Hikosaka, O. Two types of dopamine neuron distinctly convey positive and negative motivational signals. Nature 2009, 459, 837–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Distinct tonic and phasic anticipatory activity in lateral habenula and dopamine neurons. Neuron 2010, 67, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Sandoval, I.; Phan, A.; Chakraborty, M.; Davis, R.L. Reciprocal synapses between mushroom body and dopamine neurons form a positive feedback loop required for learning. Elife 2017, 6, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.A.; Cervantes-Sandoval, I.; Nicholas, E.P.; Davis, R.L. Dopamine is required for learning and forgetting in Drosophila. Neuron 2012, 74, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Plaa̧ais, P.Y.; Yamagata, N.; Pfeiffer, B.D.; Aso, Y.; Friedrich, A.B.; Siwanowicz, I.; Rubin, G.M.; Preat, T.; Tanimoto, H. A subset of dopamine neurons signals reward for odour memory in Drosophila. Nature 2012, 488, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Burke, C.J.; Huetteroth, W.; Owald, D.; Perisse, E.; Krashes, M.J.; Das, G.; Gohl, D.; Silies, M.; Certel, S.; Waddell, S. Layered reward signalling through octopamine and dopamine in Drosophila. Nature 2012, 492, 433–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krashes, M.J.; DasGupta, S.; Vreede, A.; White, B.; Armstrong, J.D.; Waddell, S. A neural circuit mechanism integrating motivational state with memory expression in Drosophila. Cell 2009, 139, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Huetteroth, W.; Perisse, E.; Lin, S.; Klappenbach, M.; Burke, C.; Waddell, S. Sweet taste and nutrient value subdivide rewarding dopaminergic neurons in drosophila. Curr. Biol. 2015, 25, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Felsenberg, J.; Barnstedt, O.; Cognigni, P.; Lin, S.; Waddell, S. Re-evaluation of learned information in Drosophila. Nature 2017, 544, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claridge-Chang, A.; Roorda, R.D.; Vrontou, E.; Sjulson, L.; Li, H.; Hirsh, J.; Miesenböck, G. Writing Memories with Light-Addressable Reinforcement Circuitry. Cell 2009, 139, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Herb, A.; Ogueta, M.; Siwanowicz, I.; Templier, T.; Friedrich, A.B.; Ito, K.; Scholz, H.; Tanimoto, H. Three Dopamine pathways induce aversive odor memories with different stability. PLoS Genet. 2012, 8, e1002768. [Google Scholar] [CrossRef] [PubMed]
- Rohwedder, A.; Wenz, N.L.; Stehle, B.; Huser, A.; Yamagata, N.; Zlatic, M.; Truman, J.W.; Tanimoto, H.; Saumweber, T.; Gerber, B.; et al. Four Individually Identified Paired Dopamine Neurons Signal Reward in Larval Drosophila. Curr. Biol. 2016, 26, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Himmelreich, S.; Masuho, I.; Berry, J.A.; MacMullen, C.; Skamangas, N.K.; Martemyanov, K.A.; Davis, R.L. Dopamine Receptor DAMB Signals via Gq to Mediate Forgetting in Drosophila. Cell Rep. 2017, 21, 2074–2081. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, D.; Donlea, J.M.; Talbot, C.B.; Song, S.M.; Thurston, A.J.F.; Miesenböck, G. Operation of a homeostatic sleep switch. Nature 2016, 536, 333–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Liu, S.; Kodama, L.; Driscoll, M.R.; Wu, M.N. Two dopaminergic neurons signal to the dorsal fan-shaped body to promote wakefulness in Drosophila. Curr. Biol. 2012, 22, 2114–2123. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Kume, K. Functional characterization of dopamine transporter in vivo using Drosophila melanogaster behavioral assays. Front. Behav. Neurosci. 2014, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Tomita, J.; Tanimoto, H.; Endo, K.; Ito, K.; Kume, S.; Kume, K. Identification of a dopamine pathway that regulates sleep and arousal in Drosophila. Nat. Neurosci. 2012, 15, 1516–1523. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tang, B.; Zhao, G.; Pan, Q.; Xia, K.; Bodmer, R.; Zhang, Z. Dispensable role of Drosophila ortholog of LRRK2 kinase activity in survival of dopaminergic neurons. Mol. Neurodegener. 2008, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.B.; Kim, W.; Lee, S.; Chung, J. Loss of LRRK2/PARK8 induces degeneration of dopaminergic neurons in Drosophila. Biochem. Biophys. Res. Commun. 2007, 358, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gehrke, S.; Imai, Y.; Huang, Z.; Ouyang, Y.; Wang, J.W.; Yang, L.; Beal, M.F.; Vogel, H.; Lu, B. Mitochondrial pathology and muscle and dopaminergic neuron degeneration caused by inactivation of Drosophila Pink1 is rescued by Parkin. Proc. Natl. Acad. Sci. USA 2006, 103, 10793–10798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Paik, D.; Bang, S.; Kang, J.; Chun, B.; Lee, S.; Bae, E.; Chung, J.; Kim, J. Loss of spastic paraplegia gene atlastin induces age-dependent death of dopaminergic neurons in Drosophila. Neurobiol. Aging 2008, 29, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R.; et al. Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [PubMed]
- Feany, M.B.; Bender, W.W. A Drosophila model of Parkinson’s disease. Nature 2000, 404, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Sitaraman, D.; Zars, M.; Laferriere, H.; Chen, Y.; Sable-smith, A.; Kitamoto, T.; Rottinghaus, G.E.; Zars, T. Serotonin is necessary for place memory in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 5579–5584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Q.; Lin, F.; Zheng, X.; Sehgal, A. Serotonin Modulates Circadian Entrainment in Drosophila. Neuron 2005, 47, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Alekseyenko, O.V.; Lee, C.; Kravitz, E.A. Targeted Manipulation of Serotonergic Neurotransmission Affects the Escalation of Aggression in Adult Male Drosophila melanogaster. PLoS ONE 2010, 5, e10806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majeed, Z.R.; Abdeljaber, E.; Soveland, R.; Cornwell, K.; Bankemper, A.; Koch, F.; Cooper, R.L. Modulatory action by the serotonergic system : Behavior and neurophysiology in Drosophila melanogaster. Neural Plast 2016, 2016, 7291438. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Marie Macara, A.; Li, R.; Hu, Y.; Iwasaki, K.; Dunnings, Z.; Firestone, E.; Horvatic, S.; Guntur, A.; Shafer, O.T.; et al. Serotonergic modulation enables pathway-specific plasticity in Drosophila. Neuron 2017, 95, 623–638. [Google Scholar] [CrossRef] [PubMed]
- Scheunemann, L.; Plaçais, P.Y.; Dromard, Y.; Schwärzel, M.; Preat, T. Dunce Phosphodiesterase Acts as a Checkpoint for Drosophila Long-Term Memory in a Pair of Serotonergic Neurons. Neuron 2018, 98, 350–365. [Google Scholar] [CrossRef] [PubMed]
- Giang, T.; Rauchfuss, S.; Ogueta, M.; Scholz, H. The serotonin transporter expression in Drosophila melanogaster. J. Neurogenet. 2011, 25, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Niens, J.; Reh, F.; Çoban, B.; Cichewicz, K.; Eckardt, J.; Liu, Y.-T.; Hirsh, J.; Riemensperger, T.D. Dopamine Modulates Serotonin Innervation in the Drosophila Brain. Front. Syst. Neurosci. 2017, 11, 76. [Google Scholar] [CrossRef] [PubMed]
- Saudou, F.; Boschert, U.; Amlaiky, N.; Plassat, J.L.; Hen, R. A family of Drosophila serotonin receptors with distinct intracellular signalling properties and expression patterns. EMBO J. 1992, 11, 7–17. [Google Scholar] [PubMed]
- Colas, J.F.; Launay, J.M.; Kellermann, O.; Rosay, P.; Maroteaux, L. Drosophila 5-HT2 serotonin receptor: Coexpression with fushi-tarazu during segmentation. Proc. Natl. Acad. Sci. USA 1995, 92, 5441–5445. [Google Scholar] [CrossRef] [PubMed]
- Witz, P.; Amlaiky, N.; Plassat, J.L.; Maroteaux, L.; Borrelli, E.; Hen, R. Cloning and characterization of a Drosophila serotonin receptor that activates adenylate cyclase. Proc. Natl. Acad. Sci. USA 1990, 87, 8940–8944. [Google Scholar] [CrossRef] [PubMed]
- Gasque, G.; Conway, S.; Huang, J.; Rao, Y.; Vosshall, L.B. Small molecule drug screening in Drosophila identifies the 5HT2A receptor as a feeding modulation target. Sci. Rep. 2013, 3, 2120. [Google Scholar] [CrossRef] [PubMed]
- Becnel, J.; Johnson, O.; Luo, J.; Nässel, D.R.; Nichols, C.D. The serotonin 5-HT7dro receptor is expressed in the brain of drosophila, and is essential for normal courtship and mating. PLoS ONE 2011, 6, e20800. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.M.; Neckameyer, W.S. Serotonin synthesis by two distinct enzymes in Drosophila melanogaster. Arch. Insect Biochem. Physiol. 2005, 59, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Colas, J.F.; Launay, J.M.; Vonesch, J.L.; Hickel, P.; Maroteaux, L. Serotonin synchronises convergent extension of ectoderm with morphogenetic gastrulation movements in Drosophila. Mech. Dev. 1999, 87, 77–91. [Google Scholar] [CrossRef]
- Neckameyer, W.S.; Coleman, C.M.; Eadie, S.; Goodwin, S.F. Compartmentalization of neuronal and peripheral serotonin synthesis in Drosophila melanogaster. Genes Brain Behav. 2007, 6, 756–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daubert, E.A.; Heffron, D.S.; Mandell, J.W.; Condron, B.G. Serotonergic dystrophy induced by excess serotonin. Mol. Cell. Neurosci. 2010, 44, 297–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budnik, V.; Wu, C.F.; White, K. Altered branching of serotonin-containing neurons in Drosophila mutants unable to synthesize serotonin and dopamine. J. Neurosci. 1989, 9, 2866–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demchyshyn, L.L.; Pristupa, Z.B.; Sugamori, K.S.; Barker, E.L.; Blakely, R.D.; Wolfgang, W.J.; Forte, M.A.; Niznik, H.B. Cloning, expression, and localization of a chloride-facilitated, cocaine-sensitive serotonin transporter from Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1994, 91, 5158–5162. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, S.; Molina-Mateo, D.; Escobedo, P.; Zárate, R.V.; Fritz, E.; Fierro, A.; Perez, E.G.; Iturriaga-Vasquez, P.; Reyes-Parada, M.; Varas, R.; et al. Characterization of a Novel Drosophila SERT Mutant: Insights on the Contribution of the Serotonin Neural System to Behaviors. ACS Chem. Neurosci. 2017, 8, 2168–2179. [Google Scholar] [CrossRef] [PubMed]
- Albin, S.D.; Kaun, K.R.; Knapp, J.M.; Chung, P.; Heberlein, U.; Simpson, J.H. A Subset of Serotonergic Neurons Evokes Hunger in Adult Drosophila. Curr. Biol. 2015, 25, 2435–2440. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; He, J.; Kaiser, A.; Gräber, N.; Schläger, L.; Ritze, Y.; Scholz, H. A single pair of serotonergic neurons counteracts serotonergic inhibition of ethanol attraction in Drosophila. PLoS ONE 2016, 11, e0167518. [Google Scholar] [CrossRef] [PubMed]
- Pooryasin, A.; Fiala, A. Identified Serotonin-Releasing Neurons Induce Behavioral Quiescence and Suppress Mating in Drosophila. J. Neurosci. 2015, 35, 12792–12812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, A.S.; Hermanns, T.; Poeck, B.; Strauss, R. Serotonin modulates a depression-like state in Drosophila responsive to lithium treatment. Nat. Commun. 2017, 8, 15738. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.T.H.; Feng, H.; Teo, W.L. Interaction of the dopaminergic and serotonergic systems in the rat striatum: Effects of selective antagonists and uptake inhibitors. Neurosci. Res. 1995, 23, 115–119. [Google Scholar] [CrossRef]
- Sasaki-Adams, D.M.; Kelley, A.E. Serotonin-dopamine interactions in the control of conditioned reinforcement and motor behavior. Neuropsychopharmacology 2001, 25, 440–452. [Google Scholar] [CrossRef]
- Daw, N.D.; Kakade, S.; Dayan, P. Opponent interactions between serotonin and dopamine. Neural Netw. 2002, 15, 603–616. [Google Scholar] [CrossRef]
- Niederkofler, V.; Asher, T.E.; Dymecki, S.M. Functional Interplay between Dopaminergic and Serotonergic Neuronal Systems during Development and Adulthood. ACS Chem. Neurosci. 2015, 6, 1055–1070. [Google Scholar] [CrossRef] [PubMed]
- Depue, R.A.; Spoont, M.R. A Behavioral Dimension of Constraint. Ann. N. Y. Acad. Sci. 1986, 487, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Spoont, M.R. Modulatory role of serotonin in neural information processing: Implications for human psychopathology. Psychol. Bull. 1992, 112, 330–350. [Google Scholar] [CrossRef] [PubMed]
- Kume, K. A Drosophila dopamine transporter mutant, fumin (fmn), is defective in arousal regulation. Sleep Biol. Rhythems 2006, 4, 263–273. [Google Scholar] [CrossRef]
- Andretic, R.; Van Swinderen, B.; Greenspan, R.J. Dopaminergic modulation of arousal in Drosophila. Curr. Biol. 2005, 15, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.L.; Dauwalder, B. The Drosophila dunce locus: Learning and memory genes in the fly. Trends Genet. 1991, 7, 224–229. [Google Scholar] [CrossRef]
- Breese, G.R.; Traylor, T.D. Effect of 6-hydroxydopamine on brain norepinephrine and dopamine evidence for selective degeneration of catecholamine neurons. J. Pharmacol. Exp. Ther. 1970, 174, 413–420. [Google Scholar] [PubMed]
- Luthman, J.; Fredriksson, A.; Sundström, E.; Jonsson, G.; Archer, T. Selective lesion of central dopamine or noradrenaline neuron systems in the neonatal rat: Motor behavior and monoamine alterations at adult stage. Behav. Brain Res. 1989, 33, 267–277. [Google Scholar] [CrossRef]
- Stachowiak, M.K.; Bruno, J.P.; Snyder, A.M.; Stricker, E.M.; Zigmond, M.J. Apparent sprouting of striatal serotonergic terminals after dopamine-depleting brain lesions in neonatal rats. Brain Res. 1984, 291, 164–167. [Google Scholar] [CrossRef]
- Rozas, G.; Liste, I.; Guerra, M.J.; Labandeira-Garcia, J.L. Sprouting of the serotonergic afferents into striatum after selective lesion of the dopaminergic system by MPTP in adult mice. Neurosci. Lett. 1998, 245, 151–154. [Google Scholar] [CrossRef]
- Di Giovanni, G.; Esposito, E.; Di Matteo, V. Role of serotonin in central dopamine dysfunction. CNS Neurosci. Ther. 2010, 16, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bartolomeis, A.; Buonaguro, E.F.; Iasevoli, F. Serotonin-glutamate and serotonin-dopamine reciprocal interactions as putative molecular targets for novel antipsychotic treatments: From receptor heterodimers to postsynaptic scaffolding and effector proteins. Psychopharmacology 2013, 225, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, C.; Iasevoli, F.; Buonaguro, E.F.; De Berardis, D.; Fornaro, M.; Fiengo, A.L.C.; Martinotti, G.; Orsolini, L.; Valchera, A.; Di Giannantonio, M.; et al. Treating the synapse in major psychiatric disorders: The role of postsynaptic density network in dopamine-glutamate interplay and psychopharmacologic drugs molecular actions. Int. J. Mol. Sci. 2017, 18, 135. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Miyashita, T.; Fu, T.F.; Lin, W.Y.; Wu, C.L.; Pyzocha, L.; Lin, I.R.; Saitoe, M.; Tully, T.; Chiang, A.S. NMDA receptors mediate olfactory learning and memory in Drosophila. Curr. Biol. 2005, 15, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.W.; Wilson, R.I. Glutamate is an inhibitory neurotransmitter in the Drosophila olfactory system. Proc. Natl. Acad. Sci. USA 2013, 110, 10294–10299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, A.J. Glutamate-gated chloride channels. J. Biol. Chem. 2012, 287, 40232–40238. [Google Scholar] [CrossRef] [PubMed]
- Talbot, K. The sandy (sdy) mouse: A dysbindin-1 mutant relevant to schizophrenia research. Prog. Brain Res. 2009, 179, 87–94. [Google Scholar] [PubMed]
- Shao, L.; Shuai, Y.; Wang, J.; Feng, S.; Lu, B.; Li, Z.; Zhao, Y.; Wang, L.; Zhong, Y. Schizophrenia susceptibility gene dysbindin regulates glutamatergic and dopaminergic functions via distinctive mechanisms in Drosophila. Proc. Natl. Acad. Sci. USA 2011, 108, 18831–18836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, M. A null mutation in TGF-alpha leads to a reduction in midbrain dopaminergic neurons in the substantia nigra. Nat. Neurosci. 1998, 1, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Ishimura, K.; Takeuchi, Y.; Fujiwara, K.; Tominaga, M.; Yoshioka, H.; Sawada, T. Quantitative analysis of the distribution of serotonin-immunoreactive cell bodies in the mouse brain. Neurosci. Lett. 1988, 91, 265–270. [Google Scholar] [CrossRef]
- Martin, C.A.; Krantz, D.E. Drosophila melanogaster as a genetic model system to study neurotransmitter transporters. Neurochem. Int. 2014, 73, 71–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Xu, A.Q.; Giraud, J.; Poppinga, H.; Riemensperger, T.; Fiala, A.; Birman, S. Neural Control of Startle-Induced Locomotion by the Mushroom Bodies and Associated Neurons in Drosophila. Front. Syst. Neurosci. 2018, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Sitaraman, D.; Aso, Y.; Rubin, G.M.; Nitabach, M.N. Control of Sleep by Dopaminergic Inputs to the Drosophila Mushroom Body. Front. Neural Circuits 2015, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- Landayan, D.; Feldman, D.S.; Wolf, F.W. Satiation state-dependent dopaminergic control of foraging in Drosophila. Sci. Rep. 2018, 8, 5777. [Google Scholar] [CrossRef] [PubMed]
- Azanchi, R.; Kaun, K.R.; Heberlein, U. Competing dopamine neurons drive oviposition choice for ethanol in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 21153–21158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Philipsborn, A.C.; Jörchel, S.; Tirian, L.; Demir, E.; Morita, T.; Stern, D.L.; Dickson, B.J. Cellular and behavioral functions of fruitless isoforms in Drosophila courtship. Curr. Biol. 2014, 24, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Keleman, K.; Vrontou, E.; Kruttner, S.; Yu, J.Y.; Kurtovic-Kozaric, A.; Dickson, B.J. Dopamine neurons modulate pheromone responses in Drosophila courtship learning. Nature 2012, 489, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.X.; Rogulja, D.; Crickmore, M.A. Dopaminergic circuitry underlying mating drive. Neuron 2016, 91, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, A.; Issa, A.R.; Seugnet, L.; Birman, S.; Klarsfeld, A. Drosophila clock is required in brain pacemaker neurons to prevent premature locomotor aging independently of its circadian function. PLoS Genet. 2017, 13, e1006507. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tabuchi, M.; Liu, S.; Kodama, L.; Horiuchi, W.; Daniels, J.; Chiu, L.; Baldoni, D.; Wu, M.N. Branch-specific plasticity of a bifunctional dopamine circuit encodes protein hunger. Science 2017, 356, 534–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marella, S.; Mann, K.; Scott, K. Dopaminergic modulation of sucrose acceptance behavior in Drosophila. Neuron 2012, 73, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Alekseyenko, O.V.; Chan, Y.B.; De La Paz Fernandez, M.; Bülow, T.; Pankratz, M.J.; Kravitz, E.A. Single serotonergic neurons that modulate aggression in Drosophila. Curr. Biol. 2014, 24, 2700–2707. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.T.; Lin, H.W.; Chang, Y.H.; Fu, T.F.; Dubnau, J.; Hirsh, J.; Lee, T.; Chiang, A.S. Serotonin-mushroom body circuit modulating the formation of anesthesia-resistant memory in Drosophila. Proc. Natl. Acad. Sci. USA 2011, 108, 13794–13799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haynes, P.R.; Christmann, B.L.; Griffith, L.C. A single pair of neurons links sleep to memory consolidation in drosophila melanogaster. Elife 2015, 4, e03868. [Google Scholar] [CrossRef] [PubMed]
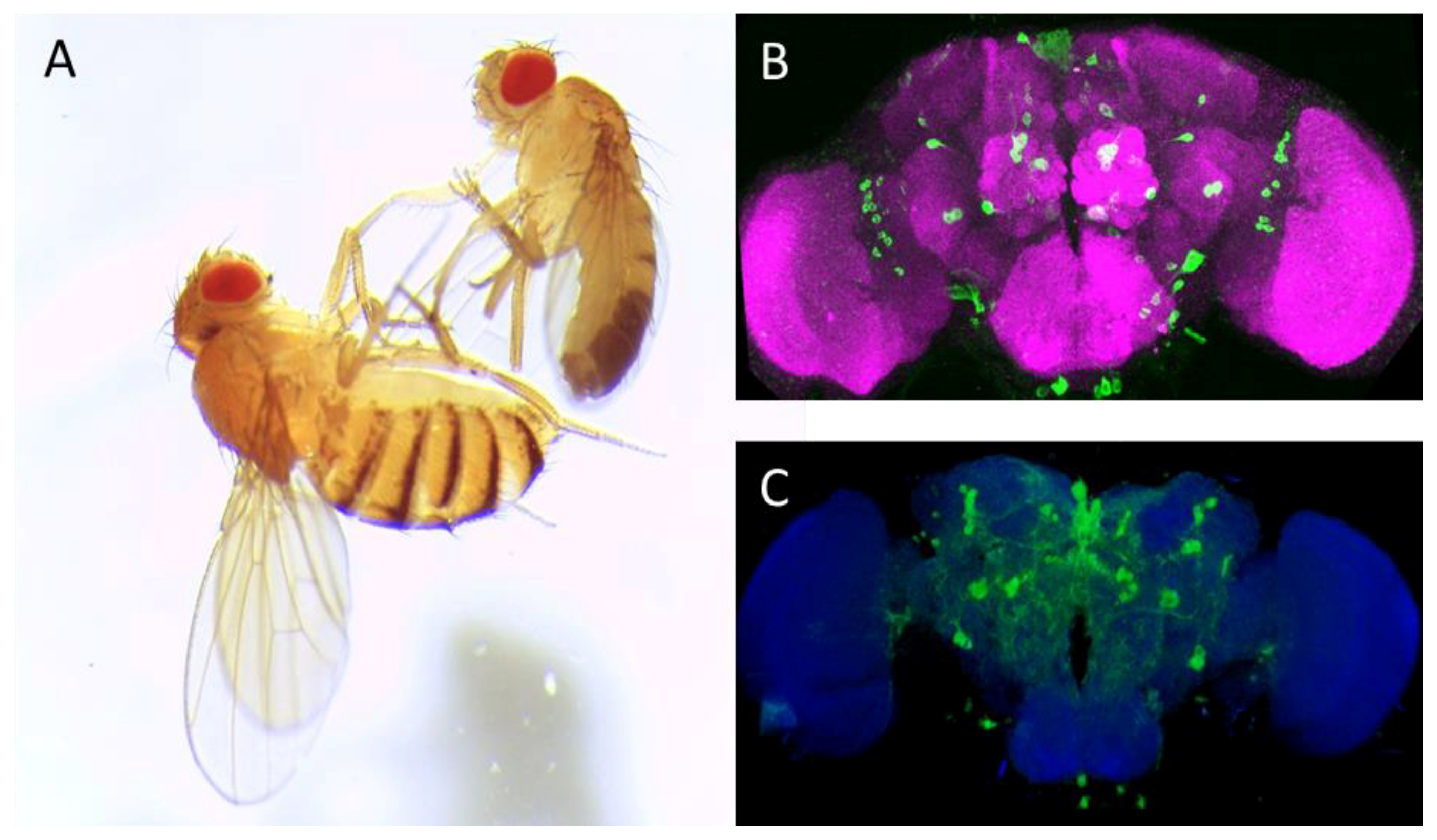

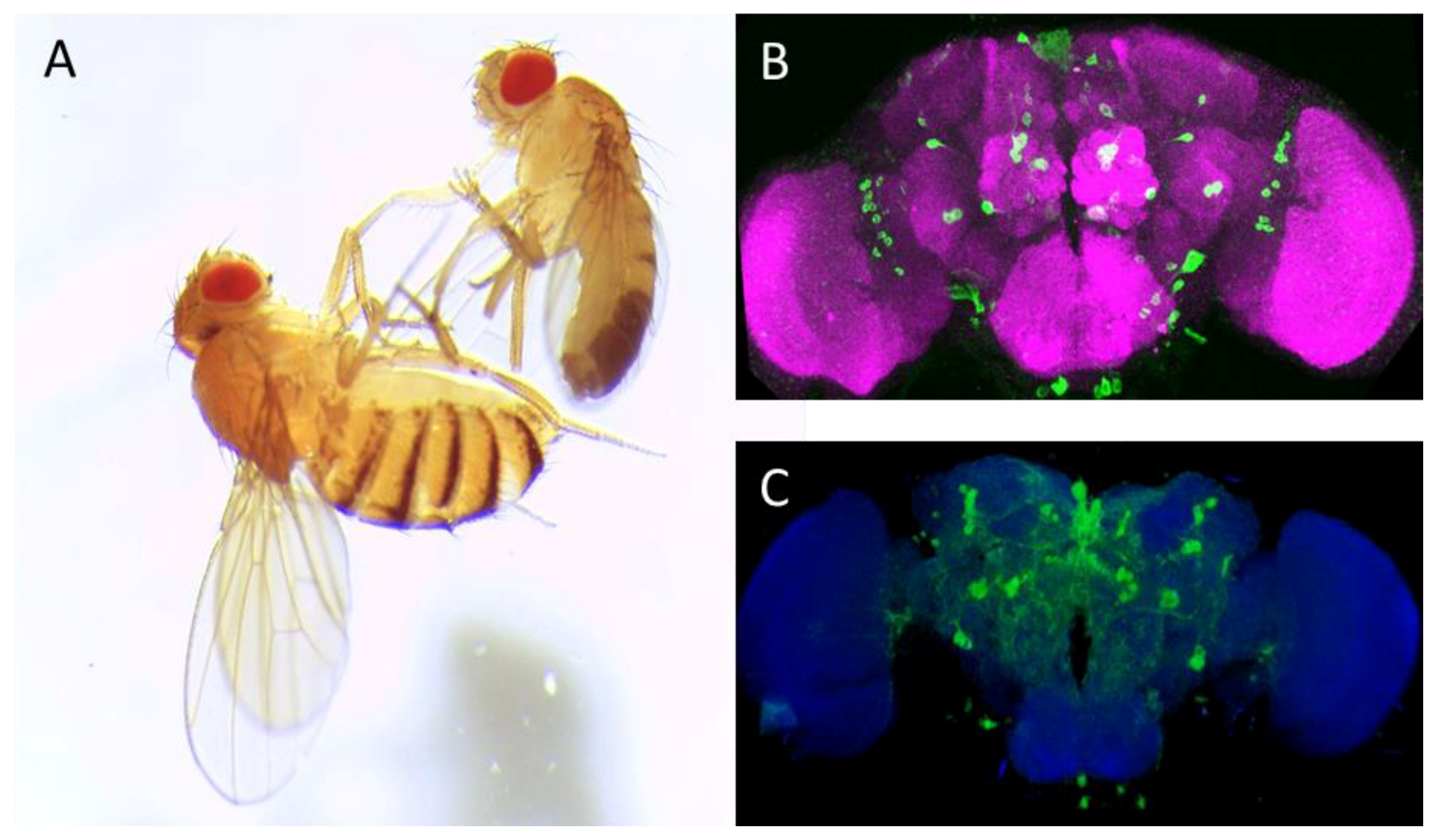{kind=link}
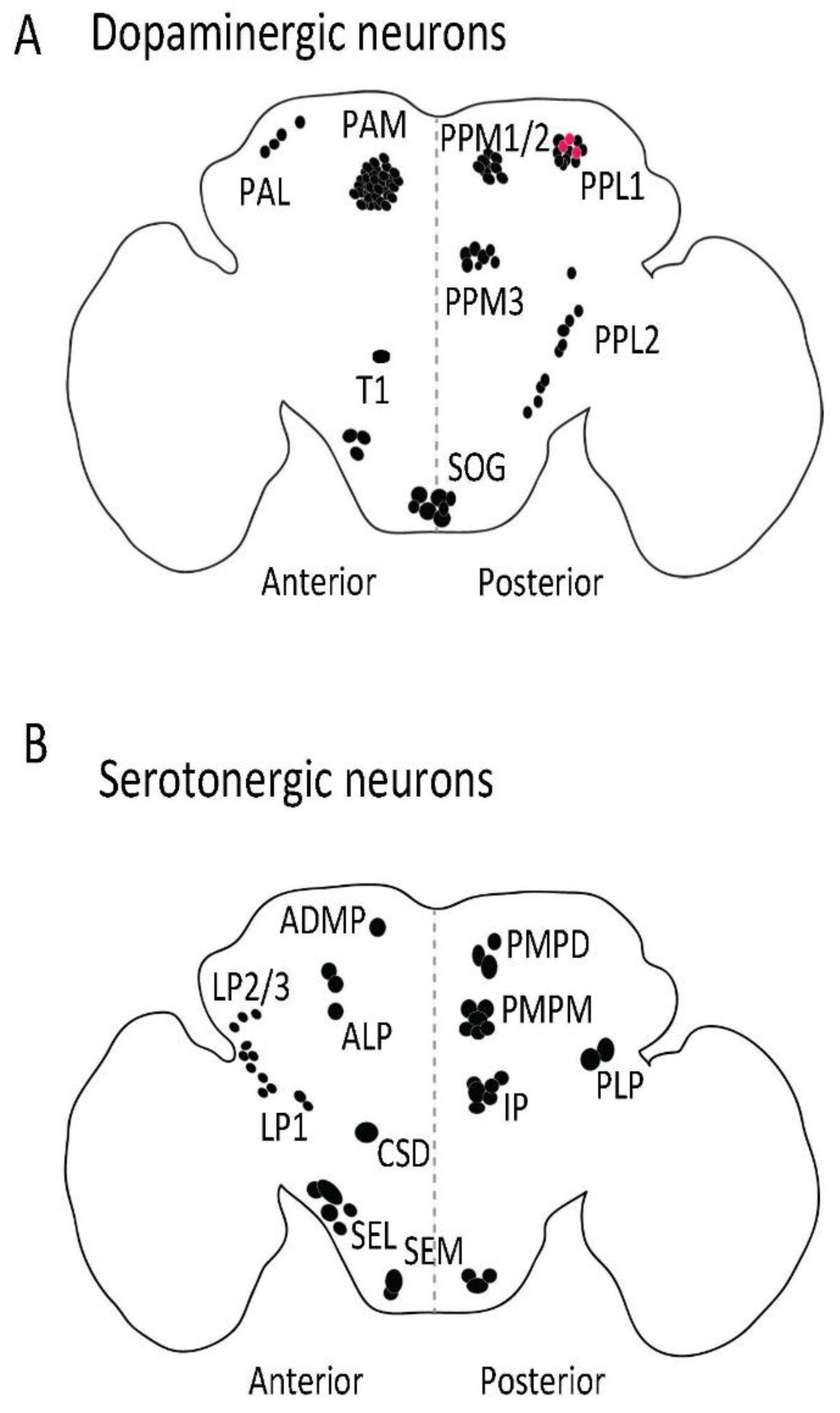
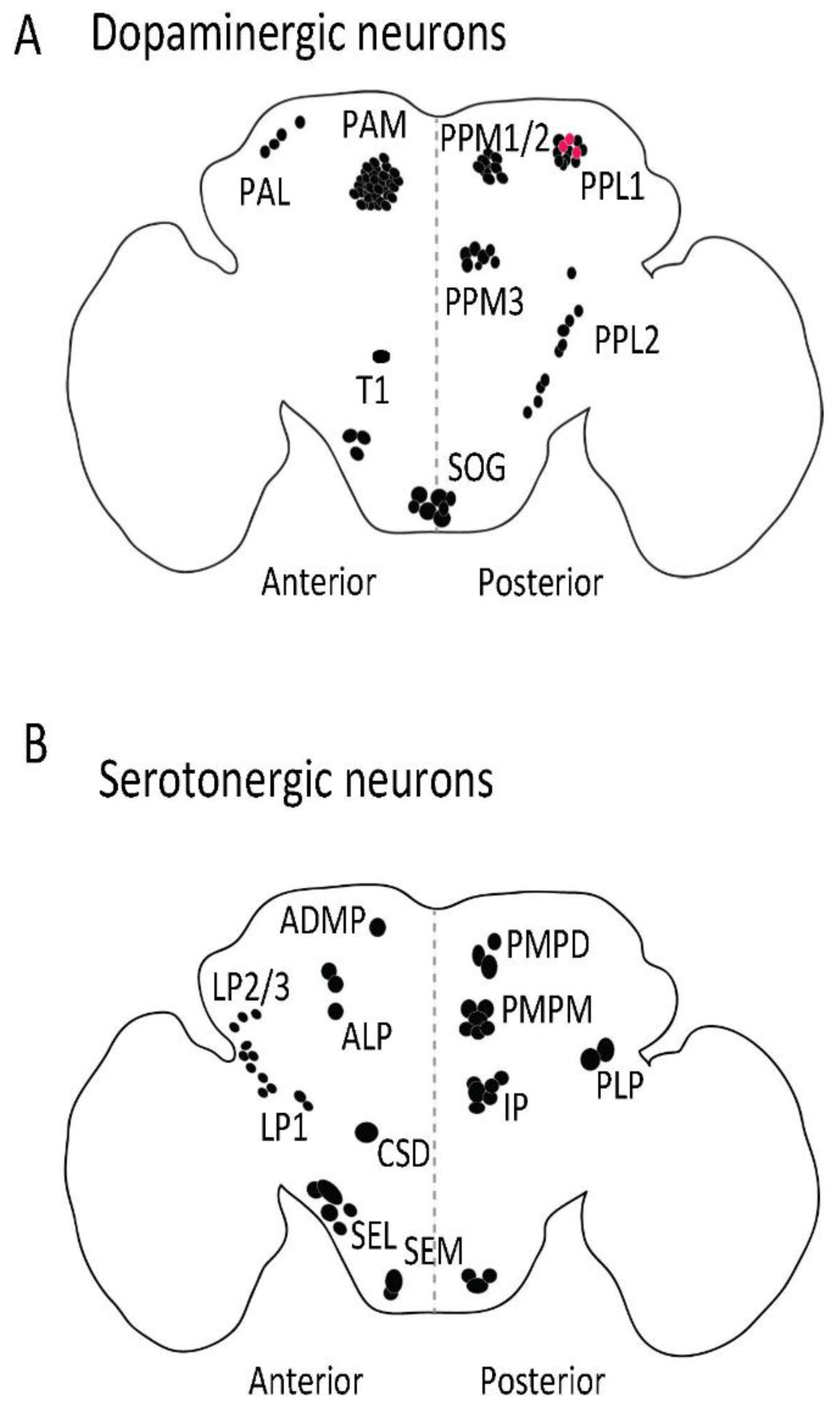{kind=link}
| Neuronal Cluster | Function | Ref. |
|---|---|---|
| Dopaminergic neurons | ||
| PAM | Sugar reward, reward signaling, long-term and short-term memory formation, aversive memory formation, negative geotaxis, promote wakefulness, foraging behavior, promote oviposition preference | [22,71,77,138,139,140,141] |
| PAL | Involved in mating drive | [142,143,144] |
| PPL1 | Aversive memory formation, sugar reward and nutrient value, negative geotaxis, modulate sleep, Inhibit oviposition preference | [1,3,4,5,7,11,12,145] |
| PPM2 | Protein consumption preference | [146] |
| PPM3 | Aggression phenotype Modulate sleep, promote oviposition preference | [7,14,15,16] |
| T1 | Aggression phenotype | [21] |
| SOG | Proboscis extension in response to satiety state | [147] |
| Serotonergic neurons | ||
| PLP | Aggression | [148] |
| PMPM | Modulate anesthesia resistant memory and sleep | [149,150] |
| CSD | Modulate ethanol perception | [111] |
| SEL | Long term memory formation | [95] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasture, A.S.; Hummel, T.; Sucic, S.; Freissmuth, M. Big Lessons from Tiny Flies: Drosophila melanogaster as a Model to Explore Dysfunction of Dopaminergic and Serotonergic Neurotransmitter Systems. Int. J. Mol. Sci. 2018, 19, 1788. https://doi.org/10.3390/ijms19061788
Kasture AS, Hummel T, Sucic S, Freissmuth M. Big Lessons from Tiny Flies: Drosophila melanogaster as a Model to Explore Dysfunction of Dopaminergic and Serotonergic Neurotransmitter Systems. International Journal of Molecular Sciences. 2018; 19(6):1788. https://doi.org/10.3390/ijms19061788
Chicago/Turabian StyleKasture, Ameya Sanjay, Thomas Hummel, Sonja Sucic, and Michael Freissmuth. 2018. "Big Lessons from Tiny Flies: Drosophila melanogaster as a Model to Explore Dysfunction of Dopaminergic and Serotonergic Neurotransmitter Systems" International Journal of Molecular Sciences 19, no. 6: 1788. https://doi.org/10.3390/ijms19061788