
Novel In Vitro Multienzyme Cascade for Efficient Synthesis of d-Tagatose from Sucrose

Key Laboratory of Industrial Biotechnology, School of Biotechnology, Jiangnan University, Wuxi 214122, China
*
Author to whom correspondence should be addressed.
Catalysts 2023, 13(12), 1515; https://doi.org/10.3390/catal13121515
Submission received: 19 October 2023
/
Revised: 13 December 2023
/
Accepted: 13 December 2023
/
Published: 15 December 2023
(This article belongs to the Special Issue Advances in Enzyme Engineering, Biocatalysis and Biosynthesis)
Abstract
:d-Tagatose is a rare sugar with low calories, and is extensively used in food, beverage, and drug additives. In this study, an in vitro multienzyme cascade route for d-tagatose synthesis from sucrose (MCTS) was designed, which contains five enzymes (sucrose phosphorylase, fructokinase, d-fructose 6-phosphate 4-epimerase, d-tagatose 6-phosphate phosphatase, and polyphosphate kinase). The whole MCTS route comprised a sucrose phosphorylation reaction, and a phosphorylation–dephosphorylation reaction coupled with an ATP regeneration system. After optimization, the conversion of d-tagatose from 10 mM sucrose reached 82.3%. At an elevated sucrose concentration of 50 mM, 72.4% of d-tagatose conversion and 0.27 g·L–1·h−1 of space–time yield were obtained. Furthermore, ADP consumption decreased to 1% of the sucrose concentration after introducing the ATP regeneration system. The MCTS strategy is an efficient and cost-effective approach for d-tagatose production.
1. Introduction
d-Tagatose (the C-4 epimer of d-fructose) is a common rare natural monosaccharide, which has up to 92% of the sweetness and only 30% of the calories of sucrose [1]. It was certified as a safe food by the U.S. Food and Drug Administration in 2001, and has been used as a sweetener in food and beverages (e.g., chocolate candies, ready-to-eat cereals, ice cream, chewing gum, diet sodas, and health bars) [2]. d-Tagatose has various pharmacological functions in preventing obesity [3,4] and caries [5], and promoting probiotic activity [6]. Accordingly, it can be used as a pharmaceutical additive to mask undesirable tastes (e.g., toothpaste and lipstick). In addition, d-tagatose can be also used as an anti-type 2 diabetes drug due to its anti-glycemic effects [7,8].
Since the first synthesis of d-tagatose in the early 1990s [9], many chemosynthesis and biosynthesis methods for producing d-tagatose have been proposed. In general, chemical methods for d-tagatose synthesis primarily depend on alkaline conditions, which are favorable for the isomerization of d-galactose with alkali metal salts and subsequent formation of metal hydroxide-tagatose intermediate precipitates. Subsequently, d-tagatose is released from the intermediate by adding acid [10]. However, this method generates numerous by-products, has a difficult downstream process, and has adverse effects on the environment. The enzymatic biosynthesis of d-tagatose has become a major interest in research recently due to its low cost, high purity, and environmental friendliness. For instance, a two-enzyme (β-galactosidase and l-arabinose isomerase) catalysis has poor enzyme specificity and a low conversion (less than 45%) for d-tagatose synthesis using lactose as a substrate [11,12]. Approximately 72% conversion was obtained after galactosol-2-dehydrogenase derived from Rhizobium leguminosarum bv. viciae 3841 was used to catalyze the synthesis of d-tagatose from galactitol [13]. However, the industrialization of this approach is hindered by many factors, such as an expensive substrate (galactitol), excessive amount of cofactor NAD+, and low enzymatic thermostability. Owing to the reversibility of d-tagatose 3-epimerase, a low conversion of d-tagatose (37.2%) was obtained from sorbose [14]. To break the thermodynamic equilibrium, many multienzyme cascade routes for d-tagatose biosynthesis have been proposed [15,16]. A three-enzyme cascade route achieved 80% d-tagatose conversion using d-fructose as a substrate [17]. However, a large amount of cofactors (ATP) increased the cost. A five-enzyme cascade route (e.g., α-glucan phosphorylase, phosphoglucomutase, glucose 6-phosphate isomerase, d-tagatose 1,6-bisphosphate aldolase, and phosphoglycolate phosphatase) designed to generate d-tagatose from maltodextrin led to a final conversion of 20.8% [18,19,20].
ATP is an energy currency required to provide energy for the synthesis of many important substances [21]. However, the high cost of ATP is usually a major limitation for industrialization. Consequently, ATP regeneration is essential for various reactions, and many approaches for regenerating ATP have been proposed. For instance, acetate kinase was employed for the generation of ATP from ADP by using acetyl phosphate as a phosphate donor [22], and pyruvate kinase was used to produce ATP from ADP with phosphoenolpyruvate as a phosphate donor [23]. Polyphosphates, as inexpensive phosphagens, are often used as donors to replace expensive phosphates in most ATP regeneration systems [24,25,26].
In this study, a novel in vitro multienzyme cascade approach for d-tagatose synthesis was designed using cheap and abundant sucrose as the substrate. This route primarily comprised four enzymes, namely sucrose phosphorylase (SP), fructokinase (FRK), fructose-6-phosphate 4-epimerase (F6PE), and tagatose 6-phosphoate phosphatase (T6PP). In addition, an ATP regeneration system caused by phosphokinase (PPK) was introduced to reduce the cost. Finally, d-tagatose conversion was improved by optimizing multienzyme reaction conditions. This route provides an economical and efficient method for synthesizing d-tagatose from sucrose.
2. Results and Discussion
2.1. Design of Multienzyme Cascade Route for d-Tagatose Synthesis from Sucrose
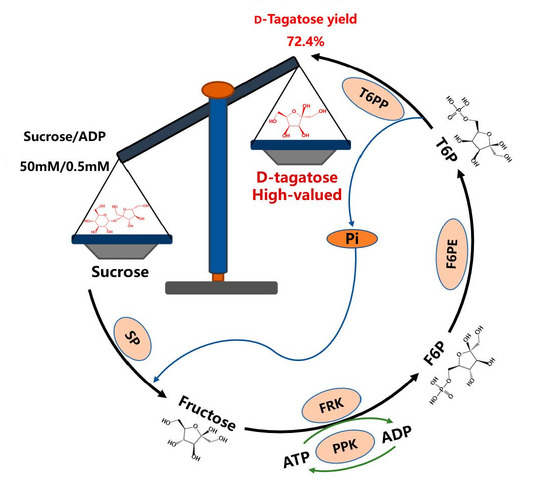
In this study, one novel multienzyme cascade route was designed to synthesize d-tagatose using sucrose as an initial substrate (named the MCTS route). It comprises five reactions (Figure 1): (I) SP catalyzes sucrose phosphorolysis for the synthesis of d-fructose and α-d-glucose-1-phosphate. (II) d-Fructose is phosphorylated by FRK in the presence of ATP to generate d-fructose-6-phosphate (F6P). (III) F6P is then epimerized by F6PE for d-tagatose-6-phosphate (T6P) synthesis. (IV) d-Tagatose is produced from T6P through dephosphorylation of T6PP. (V) To decrease ATP consumption, PPK is introduced for ATP regeneration. Compared with the reaction catalyzed by d-tagatose 3-epimerase [14], the MCTS route theoretically breaks the thermodynamic equilibrium due to the irreversibility of the last step (dephosphorylation of T6PP to d-tagatose), potentially producing high d-tagatose conversion. Additionally, the MCTS route can reduce the cost due to its inexpensive substrate (sucrose) and intruding ATP regeneration system.
The change in Gibbs free energy is used to reflect the stability of a reaction system in thermodynamics, and a lower Gibbs free energy usually leads to a more stable system [27]. Here, to identify the thermodynamic feasibility of the MCTS route, the standard Gibbs free energy change (ΔG′0) was calculated (https://equilibrator.weizmann.ac.il/ (accessed on 15 September 2023)). As shown in Figure 1, the overall ΔG′0 of MCTS (from sucrose to d-tagatose) was −37.6 kJ·mol−1, indicating the thermodynamic favorableness of the MCTS route.
2.2. Feasibility Verification of MCTS
To verify the feasibility of the MCTS route (without ATP regeneration of PPK), the following enzymes were heterologously expressed in Escherichia coli BL21 (DE3) and purified: SP from Bifidobacterium adolescentis (BaSP), FRK from Clostridium acetobutylicum (CaFRK), F6PE from Caldilinea aerophila DSM 14535 (CaF6PE), and T6PP from Methanothermobacter marburgensis (MmT6PP). The sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) results showed that the molecular weights of BaSP, CaFRK, CaF6PE, and MmT6PP were approximately 61, 36, 50, and 29 kDa, respectively (Figure 2). Moreover, the specific activities of BaSP, CaFRK, CaF6PE, and MmT6PP were summarized in Table 1. BaSP and CaFRK showed much higher enzymatic activities than CaF6PE and MmT6PP, suggesting a higher catalytic efficiency of steps I (sucrose phosphorolysis) and II (d-fructose phosphorylation) than that of steps III (F6P epimerization) and IV (T6P dephosphorylation).
Subsequently, a 1 mL reaction mixture comprising the above four enzymes was prepared, and the products were determined by high performance liquid chromatography (HPLC). As shown in Figure 3, the HPLC result for the reaction mixture showed a peak at a retention time of 12.5 min (consistent with the d-tagatose standard), suggesting the feasibility of d-tagatose synthesis from sucrose. In addition, no sucrose peak in the reaction mixture was observed, indicating the high efficiency of BaSP. However, some other peaks (e.g., unknown peak 1, unknown peak 2, and d-fructose) were detected, indicating that some intermediate products (e.g., d-fructose, F6P, and T6P) were not sufficiently converted to the final product d-tagatose. To improve the catalytic efficiency of the MCTS route and reduce the cost, the following strategies and optimization were proposed.
2.3. Construction of ATP Regeneration System
In the MCTS route, a large amount of ATP is required, which is unaffordable for industrialization. To reduce the cost, an ATP regeneration system is urgently necessary. Currently, three strategies (substrate-level phosphorylation, oxidative phosphorylation, and photophosphorylation) are often used for ATP regeneration, and substrate level phosphorylation is regarded as an efficient method [28]. Notably, PPKs are usually used to catalyze ATP generation with inexpensive polyphosphates as phosphate donors, which are easily obtained by treating biological wastewater with phosphorus accumulation [29].
As shown in Figure 4a, PPKs are usually divided into two types: PPK1 and PPK2. PPK1 enzymes mainly catalyze the dephosphorylation of ATP for the ADP generation [30], and PPK2 enzymes primarily catalyze the ATP generation from ADP and polyphosphates [31,32]. In this study, to screen an efficient PPK2 for ATP regeneration, PPK2 enzymes from Deinococcus radiodurans (DrPPK) and Delftia acidovorans (DaPPK) were purified and investigated (Figure S1). As shown in Figure 4b, the specific activity of DrPPK was 1.5-fold of that of DaPPK. Therefore, DrPPK was selected for ATP regeneration in the MCTS route.
2.4. Optimization of MCTS Route
2.4.1. Enzyme Dosage
In general, in a multienzyme cascade route, all enzymes exert synergistic performance at a certain dosage ratio. Here, multienzyme dosages in the MCTS route were optimized, and the initial dosage of each enzyme was determined through their activities and performance in the MCTS route. First, a small amount of BaSP and CaFRK (0.01 mg·mL−1) were used in the MCTS route due to its relatively high activity (Table 1). Then, 0.05 mg·mL−1 (CaF6PE and MmT6PP) and 0.04 mg·mL−1 (DrPPK) were selected as the initial dosages. Based on the initial condition, the dosages of three cascade enzymes (CaFRK, CaF6PE, and MmT6PP) were optimized. As shown in Figure 5a,b, different dosages of CaFRK and CaF6PE had insignificant effects on d-tagatose conversion. Here, 0.02 mg·mL-1 CaFRK and 0.05 mg·mL−1 CaF6PE were selected due to their relatively higher d-tagatose conversions compared with those at other concentrations. As shown in Figure 5c, d-tagatose conversion considerably increased with the increasing dosage of MmT6PP, indicating the rate-limiting role of MmT6PP. Here, 0.2 mg·mL−1 MmT6PP was selected due to its good performance in terms of d-tagatose conversion (29.9%).
2.4.2. Reaction Temperature and pH
Reaction temperature plays an important role in biocatalytic reactions by affecting enzymatic activity and stability [33]. Here, many enzymes (e.g., CaF6PE, and MmT6PP) involved in the MCTS route originated from thermophilic bacteria. Therefore, d-tagatose conversions obtained at high temperatures were higher than those at low temperatures. Especially, the MCTS route displayed an optimal temperature of 45 °C, resulting in the best d-tagatose conversion (51.9%, Figure 6a).
The pH can usually alter reaction rates by affecting the dissociation of key residues of enzymes and the binding of enzymes to substrates [34]. As shown in Figure 6b, the MCTS route showed higher d-tagatose conversions at pH 7.5 than at other pHs. In addition, different buffers showed varied effects on d-tagatose conversion. Compared with the Tris-HCl buffer (50 mM), the PBS buffer (50 mM) showed higher d-tagatose conversions possibly because of the essential role of phosphate ions as donors for BaSP in sucrose phosphorylation. In the Tris-HCl buffer (50 mM), the phosphate ions used for BaSP catalysis may come from the hydrolysis of sodium hexametaphosphate or ADP, which may inhibit ATP regeneration and result in low tagatose conversions. At the optimum pH (7.5), mixed buffers (various concentrations of PBS with a fixed concentration (50 mM) of Tris-HCl) were also investigated. As shown in Figure 6c, the mixed buffer (10 mM PBS and 50 mM Tris-HCl) displayed a better d-tagatose conversion (e.g., 70.7%) than other concentrations of PBS with 50 mM Tris-HCl buffer (Figure 6c). The possible reason is that excess phosphate ions can inhibit the activity of MmT6PP for T6P dephosphorylation to generate d-tagatose. Therefore, a mixed buffer (10 mM PBS and 50 mM Tris-HCl) was used for the MCTS cascade reaction.
2.4.3. Metal Ions
Metal ions are necessary for most metal enzymes by locating the substrate, exchanging electrons, and stabilizing the charge [35]. To investigate the effect of metal ions on d-tagatose conversion in the MCTS route, different metal ions (e.g., Mg2+, Co2+, Mn2+, Ni2+, Ca2+, Na+, Ba2+, Fe3+, Al3+, and Cu2+) were used (Figure 6d). Some bivalent metal ions (e.g., Mg2+, Mn2+, and Co2+) showed relatively better effects than other ions for the reaction, and the synergistic effect of mixed metal ions (e.g., Mix1 (Mg2+ + Mn2+), Mix2 (Mg2+ + Co2+), and Mix3 (Mn2+ + Co2+)) was also investigated. Mn2+ showed the best activation effect for the MCTS route. Then, the effect of various Mn2+ concentrations (e.g., 0.5, 1, 2, 4, and 8 mM) on the d-tagatose conversion was investigated (Figure 6e). The highest d-tagatose conversion (82.3%) was obtained at 4 mM Mn2+, and slightly decreased when the Mn2+ concentration increased to 8 mM. These effects may be ascribed to the consumption of phosphate ions by excessive Mn2+, which generated flocculent precipitate.
2.4.4. Additional ADP Concentration
In the MCTS route, DrPPK was used for ATP regeneration with ADP and polyphosphates as substrates. To further reduce the cost, the concentrations of additional ADP were optimized. As shown in Figure 6f, 82.3% of d-tagatose conversion was obtained at an ADP:sucrose ratio of 10%. Although the ADP:sucrose ratio decreased to 0.5%–5%, the d-tagatose conversions were maintained at over 70%. These results indicate that introducing an ATP regeneration system can significantly reduce the dosage of additional ADP, and thereby further reduce the cost of d-tagatose production.
2.5. Reaction Time Course
A time course was investigated to evaluate the trend of produced d-tagatose and the intermediate product (d-fructose) as the reaction time (Figure 7). The d-tagatose conversion dramatically increased to 52% in the first two hours, then slowly increased to 82.3% at 24 h. However, there was approximately 13% unutilized d-fructose in the mixture. As shown in Figure S2, in addition to d-fructose, other intermediate products (e.g., F6P) were also detected in the reaction mixture, suggesting that the cascade enzymes (e.g., CaFRK, and CaF6PE) need to be further improved in our future work.
2.6. d-Tagatose Synthesis at Elevated Concentration of Sucrose
An elevated concentration (50 mM) of sucrose was used in the MCTS route for d-tagatose synthesis. To further reduce ADP consumption, the effects of different ADP:sucrose ratios (0.2%, 0.5%, and 1%) on d-tagatose conversions were investigated. As shown in Table 2, a d-tagatose conversion of 72.4% was achieved at an ADP:sucrose ratio of 1%, leading to a space–time yield of 0.27 g·L−1 h−1 at a substrate:catalyst (S:C) ratio of 18.5. Compared with the previously reported three-enzyme cascade (e.g., hexokinase, fructose-1,6-biphosphate aldolase, and phytase) synthesis of d-tagatose (ATP: d-fructose = 100%) [17], the MCTS route greatly reduced the consumption of the expensive cofactor (ADP) by introducing ATP regeneration. Moreover, when the ADP:sucrose ratio decreased to 0.2% and 0.5%, the conversion of d-tagatose was still over 60%. In addition, compared with the d-tagatose synthesis route using maltodextrin as the substrate (involving five enzymes and leading to 37.6% conversion) [36], the MCTS route resulted in a much higher d-tagatose conversion (72.4%), providing an economical and efficient approach for d-tagatose production.
Although the d-tagatose conversion was significantly improved by the optimized MCTS route, some by-products (e.g., d-fructose and F6P) remained in the reaction mixture (Figure S2). Therefore, efficient strategies for d-tagatose recovery and purification are the focus in our future work. The following current strategies can be proposed: (I) ethanol crystallization is usually used for d-tagatose recovery [17]; (II) Ca2+ cation exchange resin chromatography is commonly used for d-tagatose purification [37]; (III) boronate affinity adsorption can be employed for recycling preparation of high-purity d-tagatose [38].
3. Materials and Methods
3.1. Genes, Plasmid, and Bacterial Strains
The plasmids and strains used in this study are summarized in Table S1. Genes encoding BaSP (GeneBank: WP_011742626.1), CaFRK (GeneBank: KHD36265.1), CaF6PE (GeneBank: WP_014433578), MmT6PP (GeneBank: WP_013296249.1), DaPPK, (GeneBank: WP_013800100.1), and DrPPK (GeneBank: WP_010886780.1) were synthesized by Exsyn Bio Co., Ltd. (Shanghai, China). All the genes were individually inserted into a pET-28a(+) vector (Takara Co., Ltd, Dalian, China) between the restriction sites of Bam HI and Xho I. E. coli BL21 (DE3) (Takara Co., Ltd, Dalian, China) was used as the host for recombinant protein expression.
3.2. Chemicals
d-Tagatose was purchased from Yuanye Bio-Technology Co., Ltd. (Shanghai, China). Sucrose and d-fructose were procured from Sinopharm Co., Ltd (Shanghai, China). Sodium hexametaphosphate was purchased from J&K Scientific Co., Ltd (Shanghai, China). ATP and ADP were procured from Bidepharm Co., Ltd (Shanghai, China) and Adamas-beta Co., Ltd (Shanghai, China), respectively. d-Fructose 6-phosphate was obtained from Solarbio Co., Ltd (Beijing, China), and the ATP test kit was purchased from Jing Key Bio-Technology Co., Ltd. (Shanghai, China). Materials used for culture media (e.g., yeast extract, tryptone, and NaCl) were obtained from Sangon Biotech Co., Ltd (Shanghai, China).
3.3. Expression and Purification of Enzymes
All recombinant plasmids carrying target genes were transformed into E. coli BL21 (DE3), and the recombinant strains were coated on Luria-Bertani (LB) plates containing 50 μg·mL−1 kanamycin and incubated at 37 °C for 12 h. Single colonies were inoculated into 5 mL of LB medium containing 50 μg·mL−1 kanamycin and cultured at 37 °C with shaking at 180 rpm for 12 h. Then, 1 mL of culture was transferred to 100 mL of LB medium containing 50 μg·mL−1 kanamycin and cultured at 37 °C. At an OD600 of 0.6–0.8, isopropyl β-d-1-thiogalactopyranoside (IPTG, 0.2 mM) was added, and then the culture was incubated at 16 °C with shaking at 180 rpm for 16 h. Subsequently, the recombinant cells were collected by centrifugation at 8000× g for 5 min at 4 °C. The harvested pellets were resuspended in Tris-HCl buffer (pH 7.4). Resuspended cells were lysed by sonication on ice, and the broken solution was centrifuged at 8000× g for 30 min at 4 °C to remove impurities. Then the supernatant was filtered through a 0.45 μm filter, and then loaded onto a 2 mL of nickel tris ethylenediamine agarose (Ni-TED) resin at 4 °C, which was beforehand equilibrated with buffer A (25 mM Tris, 20 mM imidazole, 500 mM NaCl, 10% v/v glycerol, pH 7.4). The bound protein was washed with 25 mM imidazole, and eluted with a certain concentration of imidazole. Pure enzyme was estimated by SDS-PAGE, and the concentration was measured by using the NanoDrop 2000c equipment (Thermo Fisher Scientific Co., Ltd., Shanghai, China).
3.4. Determination of Sucrose, d-Fructose, d-Tagatose, and ATP
Analyses of sucrose, d-fructose, and d-tagatose were performed by HPLC using a HPLC 1260 system (Agilent Technologies, Santa Clara, CA, USA) with a refractive index detector and a CarboPac Ca2+ column (Dima Technology Co., Ltd., Beijing, China), the mobile phase used was pure water, the flow rate was set at 0.7 mL·min−1, and the column temperature was 80 °C. ATP content was determined by an ATP kit (Jing Key Bio-Technology Co., Ltd., Shanghai, China). Briefly, in the presence of ATP, hexokinase catalyzes the generation of glucose to glucose-6-phosphate, and then glucose-6-phosphate dehydrogenase further catalyzes the generation of NADPH from NAD+. NADPH has a characteristic absorption peak at 340 nm, and ATP concentration is proportional to NADPH content [39].
The conversion of d-tagatose or d-fructose is calculated as follows:
Produced d-tagatose/d-fructose: the amount of produced d-tagatose or d-fructose (mM); initial sucrose: the amount of initial sucrose (mM).
3.5. Activity Assay
BaSP activity was determined as follows. A sucrose hydrolysis reaction mixture (1 mL) containing 50 mM PBS buffer (pH 7.5), 10 mM sucrose, and 0.01 mg·mL−1 purified BaSP was incubated at 40 °C for 10 min, and then the reaction was terminated by heating at 100 °C in a metal bath for 10 min. The concentration of d-fructose was measured by HPLC. One unit of BaSP activity was defined as the amount of enzyme required for the production of 1 μmol of d-fructose per minute at pH 7.5 and 40 °C.
FRK activity was determined by the d-fructose phosphorylation reaction. A 1 mL mixture containing 50 mM PBS buffer (pH 7.5), containing 10 mM d-fructose, 10 mM ATP, 4 mM MgCl2, and 0.005 mg·mL−1 purified CaFRK. After incubation at 40 °C for 10 min, the reaction was terminated by heating at 100 °C in a metal bath for 10 min. The concentration of d-fructose was measured by HPLC. One unit of enzyme activity was defined as the amount of enzyme that consumed 1 μmol of d-fructose per minute at pH 7.5 and 40 °C.
The isomerization reaction from d-fructose-6-phosphate to tagatose-6-phosphate was used for the determination of CaF6PE activity. A 1 mL mixture contained 50 mM PBS buffer (pH 7.5), 10 mM d-fructose-6-phosphate, 4 mM MgCl2, 0.2 mg·mL−1 MmT6PP, and 2 mg·mL−1 CaF6PE. After incubation at 40 °C for 30 min, the reaction was terminated by heating at 100 °C in a metal bath for 10 min. One unit of enzyme activity was defined as the amount of enzyme that produced 1 μmol of d-tagatose-6-phosphate per minute at pH 7.5 and 40 °C.
MmT6PP activity was determined in a 1 mL mixture containing in 50 mM PBS buffer (pH 7.5), 10 mM d-fructose-6-phosphate, 4 mM MgCl2, 2 mg·mL−1 CaF6PE, and 0.2 mg·mL−1 MmT6PP. After incubation at 40 °C for 30 min, the reaction was terminated by heating at 100 °C in a metal bath for 10 min. The concentration of d-tagatose was measured by HPLC. One unit of MmT6PP activity was defined as the amount of enzyme that produced 1 μmol of d-tagatose per minute at pH 7.5 and 40 °C.
The activity of PPK was determined using an ATP test kit. The reaction (200 μL) was performed in 50 mM PBS buffer (pH 7.5) at 40 °C, which contained 5 mM ADP, 1 mM sodium hexametaphosphate, 4 mM MgCl2, and 0.1 mg·mL−1 PPK. Then, the change in absorbance at 340 nm within 5 min was measured by a multifunctional enzyme marker. One unit of PPK activity was defined as the amount of enzyme that produced 1 μmol of ATP per minute at pH 7.5 and 40 °C.
3.6. Optimization Reaction Conditions for d-Tagatose Synthesis
The initial reaction mixture (1 mL) included 10 mM sucrose, 4 mM MgCl2, 1 mM ADP, 2 mM sodium hexametaphosphate plus 0.01 mg·mL−1 BaSP, 0.01 mg·mL−1 CaFRK, 0.05 mg·mL−1 CaF6PE and 0.05 mg·mL−1 MmT6PP, and 0.04 mg·mL−1 DrPPK. Three enzymes, namely, CaFRK (0.01, 0.02, and 0.05 mg·mL−1), CaF6PE (0.05, 0.1, and 0.2 mg·mL−1), and MmT6PP (0.05, 0.1, and 0.2 mg·mL−1) were individually investigated according to the fixed concentrations of other enzymes. The initial temperature and pH of the reaction were 40 °C and 7.5, respectively.
The optimum temperature was investigated at 35–55 °C. A sample was collected periodically, and the concentration of d-tagatose was evaluated using HPLC. To determine the optimum pH of the reaction, the MCTS was assayed in different buffers (50 mM PBS buffer, pH 6.0–8.0; 50 mM Tris-HCl buffer, pH 7.0–9.0; 50 mM Tris-HCl buffer + different concentrations of PBS buffer (0, 5, 10, 20, 30, 50 mM), pH 7.5.). The reaction was performed at 45 °C for 24 h, and the concentration of d-tagatose was detected by HPLC.
The effect of metal ions (Mg2+, Co2+, Mn2+, Ni2+, Ca2+, Na+, Ba2+, Fe3+, Al3+, Cu2+) on the MCTS route for d-tagatose synthesis was investigated in the presence of certain metal ions at a concentration of 4 mM. The effect of metal ion concentration was also determined in the presence of Mn2+ at 0.5, 1, 2, 4, and 8 mM. The reaction was performed at 45 °C, pH 7.5 for 24 h, and the concentration of d-tagatose was detected by HPLC.
The effect of ADP concentrations on the multienzyme cascade synthesis of d-tagatose was evaluated. The reaction was conducted in 50 mM Tris-HCl buffer and 10 mM PBS buffer (pH 7.5) containing 0.005, 0.01, 0.02, 0.05, 0.1, 0.2, 0.5, and 1 mM ADP plus 4 mM MnCl2 solution at 45 °C for 24 h. d-Tagatose was detected by HPLC.
3.7. Reaction Time Course
In the MCTS route for d-tagatose synthesis from 10 mM sucrose, a 2 mL mixture contained 10 mM sucrose, 1 mM ADP, 2 mM sodium hexametaphosphate, 4 mM MnCl2, 0.01 mg·mL−1 BaSP, 0.02 mg·mL−1 CaFRK, 0.05 mg·mL−1 CaF6PE, 0.2 mg·mL−1 MmT6PP, and 0.04 mg·mL−1 DrPPK. The reaction was performed at pH 7.5 (50 mM Tris-HCl + 10 mM PBS buffer) and 45 °C for 30 h. A sample was collected periodically, and the concentration of d-tagatose was detected using HPLC.
3.8. D-Tagatose Production at an Elevated Sucrose Concentration of 50 mM
In the MCTS route for d-tagatose synthesis from 50 mM sucrose, 2 mL of the reaction mixture was placed in a water bath, and then its pH was adjusted to 7.5 with a 50 mM Tris-HCl + 10mM PBS buffer solution. The mixture contained 50 mM sucrose, 20 mM MnCl2, 0.1–0.5 mM ADP, 10 mM sodium hexametaphosphate, 0.025 mg·mL−1 BaSP, 0.05 mg·mL−1 CaFRK, 0.25 mg·mL−1 CaF6PE, 0.5 mg·mL−1 MmT6PP, and 0.1 mg·mL−1 DrPPK. The reaction was performed at 45 °C for 24 h, and the concentration of d-tagatose was detected by HPLC.
4. Conclusions
In this study, an in vitro multienzyme cascade synthesis of d-tagatose from sucrose (MCTS) was successfully constructed. It included sucrose phosphorylation, d-fructose phosphorylation, F6P epimerization, T6P dephosphorylation, and ATP regeneration reactions. The MCTS route broke the thermodynamic limitations and efficiently generated d-tagatose using inexpensive sucrose as a substrate. Moreover, the cost was further reduced by introducing an ATP regeneration system. After optimization, 82.3% d-tagatose conversion was achieved from 10 mM sucrose. At an elevated concentration of sucrose (50 mM), a 72.4% d-tagatose conversion was obtained with a reduced ADP dosage (1% of sucrose). The newly designed MCTS route is a potential route for the efficient and cost-effective production of d-tagatose from sucrose.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/catal13121515/s1, Table S1: Strains and plasmids used in this study. Figure S1: SDS-PAGE analysis of purified enzymes of PPK2. M, protein marker; line 1, DrPPK; line 2, DaPPK, Figure S2: HPLC profiles of MCTS and standard substances and (by)products.
Author Contributions
Data curation, methodology, writing—original draft preparation, S.L.; methodology, investigation, W.T.; writing—review and editing, project administration, funding acquisition, Y.N.; methodology, Y.G.; writing—review and editing, funding acquisition, supervision, project administration, R.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program of China (2021YFC2102700), the National Natural Science Foundation of China (31871738), the National First-Class Discipline Program of Light Industry Technology and Engineering (LITE2018-07), the Program of Introducing Talents of Discipline to Universities (111-2-06).
Data Availability Statement
The data presented in this study are available in the article and the supplementary materials.
Acknowledgments
The authors would like to thank the Research platform of the School of Biotechnology, Jiangnan University for providing the equipment and technical support for the experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lu, Y.; Levin, G.V.; Donner, T.W. Tagatose, a new antidiabetic and obesity control drug. Diabetes. Obes. Metab. 2008, 10, 109–134. [Google Scholar] [CrossRef]
- Oh, D.-K. Tagatose: Properties, applications, and biotechnological processes. Appl. Microbiol. Biot. 2007, 76, 1–8. [Google Scholar] [CrossRef]
- Buemann, B.; Toubro, S.; Raben, A.; Blundell, J.; Astrup, A. The acute effect of d-tagatose on food intake in human subjects. Br. J. Nutr. 2000, 84, 227–231. [Google Scholar] [CrossRef]
- Mooradian, A.D. In search for an alternative to sugar to reduce obesity. Int. J. Vitam. Nutr. Res. 2019, 89, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Levin, G.V. Removal and prevention of dental plaque with d-tagatose. Int. J. Cosmet. Sci. 2010, 24, 225–234. [Google Scholar] [CrossRef]
- Laerke, H.N.; Jensen, B.B.; Højsgaard, S. In vitro fermentation pattern of d-tagatose is affected by adaptation of the microbiota from the gastrointestinal tract of pigs. J. Nutr. 2000, 130, 1772–1779. [Google Scholar] [CrossRef] [PubMed]
- Levin, G.V. Tagatose, the new GRAS sweetener and health product. J. Med. Food 2002, 5, 23–36. [Google Scholar] [CrossRef]
- Guerrero-Wyss, M.; Durán Agüero, S.; Angarita Dávila, L. d-Tagatose is a promising sweetener to control glycaemia: A new functional food. Biomed. Res. Int. 2018, 2018, 8718053. [Google Scholar] [CrossRef] [PubMed]
- Jayamuthunagai, J.; Gautam, P.; Srisowmeya, G.; Chakravarthy, M. Biocatalytic production of d-tagatose: A potential rare sugar with versatile applications. Crit. Rev. Food Sci. Nutr. 2017, 57, 3430–3437. [Google Scholar] [CrossRef] [PubMed]
- Oroskar, A.R.; Kulkarni, O.M.; House, D.W. Tagatose Production Using Simulated Moving Bed Separation. U.S. Patent 8,802,843, 12 August 2014. [Google Scholar]
- Zhan, Y.; Xu, Z.; Li, S.; Liu, X.; Xu, L.; Feng, X.; Xu, H. Coexpression of β-d-galactosidase and l-arabinose isomerase in the production of d-tagatose: A functional sweetener. J. Agric. Food Chem. 2014, 62, 2412–2417. [Google Scholar] [CrossRef]
- Zhang, X.; Lu, R.; Wang, Q.; Hu, M.; Li, Z.; Xu, M.; Yang, T.; Zhang, R.; Rao, Z. Production of d-tagatose by whole-cell conversion of recombinant Bacillus subtilis in the absence of antibiotics. Biology 2021, 10, 1343. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, S.S.; Singh, R.; Kang, Y.C.; Zhao, H.; Lee, J.-K. Cloning and characterization of a galactitol 2-dehydrogenase from Rhizobium legumenosarum and its application in d-tagatose production. Enzym. Microb. Technol. 2014, 58–59, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, Z.; Zhang, W.; Guang, C.; Mu, W. Characterization of a d-tagatose 3-epimerase from Caballeronia fortuita and its application in rare sugar production. Int. J. Biol. Macromol. 2019, 138, 536–545. [Google Scholar] [CrossRef]
- Papageorgiou, A.C. Structural characterization of multienzyme assemblies: An overview. Methods Mol. Biol. 2022, 2487, 51–72. [Google Scholar] [PubMed]
- Sun, Z.; Zhao, Q.; Haag, R.; Wu, C. Responsive emulsions for sequential multienzyme cascades. Angew. Chem. Int. Ed. Eng. 2021, 60, 8410–8414. [Google Scholar] [CrossRef]
- Lee, S.-H.; Hong, S.-H.; Kim, K.-R.; Oh, D.-K. High-yield production of pure tagatose from fructose by a three-step enzymatic cascade reaction. Biotechnol. Lett. 2017, 39, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Zhang, J.; Zhang, T.; Chen, J.; Hassanin, H.A.; Jiang, B. Characteristics of a fructose 6-phosphate 4-epimerase from Caldilinea aerophila DSM 14535 and its application for biosynthesis of tagatose. Enzym. Microb. Technol. 2020, 139, 109594. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, T.; Jiang, B.; Mu, W.; Chen, J.; Hassanin, H.A. Dictyoglomus turgidum DSM 6724 α-Glucan phosphorylase: Characterization and its application in multi-enzyme cascade reaction for d-tagatose production. Appl. Biochem. Biotechnol. 2021, 193, 3719–3731. [Google Scholar] [CrossRef]
- Dai, Y.; Li, M.; Jiang, B.; Zhang, T.; Chen, J. Whole-cell biosynthesis of d-tagatose from maltodextrin by engineered Escherichia coli with multi-enzyme co-expression system. Enzym. Microb. Technol. 2021, 145, 109747. [Google Scholar] [CrossRef]
- Bonora, M.; Patergnani, S.; Rimessi, A.; De Marchi, E.; Suski, J.M.; Bononi, A.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. ATP synthesis and storage. Purinergic Signal. 2012, 8, 343–357. [Google Scholar] [CrossRef]
- Andexer, J.N.; Richter, M. Emerging enzymes for ATP regeneration in biocatalytic processes. ChemBioChem 2015, 16, 380–386. [Google Scholar] [CrossRef]
- Wu, W.; Bergstrom, D.E.; Davisson, V.J. Chemoenzymatic preparation of nucleoside triphosphates. Curr. Protoc. Nucleic Acid. Chem. 2004, 16, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, S.; Motomura, K.; Shinoda, Y.; Urata, M.; Kato, J.; Takiguchi, N.; Ohtake, H.; Hirota, R.; Kuroda, A. Use of an Escherichia coli recombinant producing thermostable polyphosphate kinase as an ATP regenerator to produce fructose 1,6-diphosphate. Appl. Environ. Microbiol. 2007, 73, 5676–5678. [Google Scholar] [CrossRef] [PubMed]
- Aiguo, Z.; Ruiwen, D.; Meizhi, Z. Multi-enzymatic recycling of ATP and NADPH for the synthesis of 5-aminolevulinic acid using a semipermeable reaction system. Biosci. Biotechnol. Biochem. 2019, 83, 2213–2219. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Wang, Y.; Huang, H.; Jin, X.; Li, J.; Du, G.; Kang, Z. Closed-Loop system driven by ADP phosphorylation from pyrophosphate affords equimolar transformation of ATP to 3′-phosphoadenosine-5′-phosphosulfate. ACS Catal. 2021, 11, 10405–10415. [Google Scholar] [CrossRef]
- Ji, Y.; Liu, J.; Wang, C.; Zhang, F.; Xu, X.; Zhu, L. Stability improvement of aerobic granular sludge (AGS) based on Gibbs free energy change (∆G) of sludge-water interface. Water Res. 2023, 240, 120059. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Y.P.J. Enzymatic regeneration and conservation of A1vTP: Challenges and opportunities. Crit. Rev. Biotechnol. 2021, 41, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Rudi, K.; Goa, I.A.; Saltnes, T.; Sørensen, G.; Angell, I.L.; Eikås, S. Microbial ecological processes in MBBR biofilms for biological phosphorus removal from wastewater. Water Sci. Technol. 2019, 79, 1467–1473. [Google Scholar] [CrossRef]
- Nahálka, J.; Gemeiner, P.; Bucko, M.; Wang, P.G. Bioenergy beads: A tool for regeneration of ATP/NTP in biocatalytic synthesis. Artif. Cells Blood Substit. Biotechnol. 2006, 34, 515–521. [Google Scholar] [CrossRef]
- Petchey, M.R.; Rowlinson, B.; Lloyd, R.C.; Fairlamb, I.J.S.; Grogan, G. Biocatalytic synthesis of moclobemide using the amide bond synthetase mcbA coupled with an ATP recycling system. ACS Catal. 2020, 10, 4659–4663. [Google Scholar] [CrossRef]
- Ogawa, M.; Uyeda, A.; Harada, K.; Sato, Y.; Kato, Y.; Watanabe, H.; Honda, K.; Matsuura, T. Class III polyphosphate kinase 2 enzymes catalyze the pyrophosphorylation of adenosine-5’-monophosphate. ChemBioChem 2019, 20, 2961–2967. [Google Scholar] [CrossRef]
- Arcus, V.L.; Prentice, E.J.; Hobbs, J.K.; Mulholland, A.J.; Van der Kamp, M.W.; Pudney, C.R.; Parker, E.J.; Schipper, L.A. On the temperature dependence of enzyme-catalyzed rates. Biochemistry 2016, 55, 1681–1688. [Google Scholar] [CrossRef]
- Alberty, R.A. Effects of pH in rapid-equilibrium enzyme kinetics. J. Phys. Chem. B 2007, 111, 14064–14068. [Google Scholar] [CrossRef]
- Prejanò, M.; Alberto, M.E.; Russo, N.; Toscano, M.; Marino, T. The effects of the metal ion substitution into the active site of metalloenzymes: A theoretical insight on some selected cases. Catalysts 2020, 10, 1038. [Google Scholar] [CrossRef]
- Hu, C.; Wei, X.; Song, Y. A thermophilic phosphatase from Methanothermobacter marburgensis and its application to in vitro biosynthesis. Enzym. Microb. Technol. 2022, 159, 110067. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Mu, W.; Jiang, B. Efficient biotransformation of d-galactose to d-tagatose by Acidothermus cellulolytics ATCC 43068. J. Food. Biochem. 2011, 35, 1298–1310. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, M.; Lyu, X.; Wang, C.; Tong, Y.; Hua, X.; Yang, R. Recycling preparation of high-purity tagatose from galactose using one-pot boronate affinity adsorbent-based adsorption-assisted isomerization and simultaneous purification. Chem. Eng. J. 2022, 446, 137089. [Google Scholar] [CrossRef]
- Gao, H.; Li, M.; Wang, Q.; Liu, T.; Zhang, X.; Yang, T.; Xu, M.; Rao, Z. A high-throughput dual system to screen polyphosphate kinase mutants for efficient ATP regeneration in L-theanine biocatalysis. Biotechnol. Biofuels Bioprod. 2023, 16, 122. [Google Scholar]
Figure 1.
Schematic diagram of the MCTS route for d-tagatose synthesis using sucrose as a substrate.
Figure 1.
Schematic diagram of the MCTS route for d-tagatose synthesis using sucrose as a substrate.

Figure 2.
SDS-PAGE analysis of purified enzymes in the MCTS route. M, protein marker; line 1, BaSP; line 2, CaFRK; line 3, CaF6PE; line 4, MmT6PP.
Figure 2.
SDS-PAGE analysis of purified enzymes in the MCTS route. M, protein marker; line 1, BaSP; line 2, CaFRK; line 3, CaF6PE; line 4, MmT6PP.

Figure 3.
HPLC profiles of multienzyme cascade catalytic reaction system and standard substances. Purple, sucrose standard; Green, d-tagatose standard; Red, fructose standard; Blue, reaction sample. A 1 mL mixture containing BaSP (0.01 mg·mL−1), CaFRK (0.01 mg·mL−1), CaF6PE (0.05 mg·mL−1), MmT6PP (0.05 mg·mL−1), 50 mM PBS buffer (pH 7.5), 10 mM sucrose, 4 mM MgCl2, and 10 mM ATP was incubated at 40 °C for 24 h. Unknown peak 1 may include ATP/PBS/F6P, and unknown peak 2 may include T6P.
Figure 3.
HPLC profiles of multienzyme cascade catalytic reaction system and standard substances. Purple, sucrose standard; Green, d-tagatose standard; Red, fructose standard; Blue, reaction sample. A 1 mL mixture containing BaSP (0.01 mg·mL−1), CaFRK (0.01 mg·mL−1), CaF6PE (0.05 mg·mL−1), MmT6PP (0.05 mg·mL−1), 50 mM PBS buffer (pH 7.5), 10 mM sucrose, 4 mM MgCl2, and 10 mM ATP was incubated at 40 °C for 24 h. Unknown peak 1 may include ATP/PBS/F6P, and unknown peak 2 may include T6P.

Figure 4.
Polyphosphate kinase’s catalyzing reactions. (a) Schematic diagram of reactions catalyzed by polyphosphate kinases (PPK1 and PPK2). (b) Specific enzyme activity of PPK2 from Deinococcus radiodurans and Delftia acidovorans. The error bar represents the standard deviation of triplicate measurements.
Figure 4.
Polyphosphate kinase’s catalyzing reactions. (a) Schematic diagram of reactions catalyzed by polyphosphate kinases (PPK1 and PPK2). (b) Specific enzyme activity of PPK2 from Deinococcus radiodurans and Delftia acidovorans. The error bar represents the standard deviation of triplicate measurements.

Figure 5.
Optimization of enzyme dosage for the synthesis of d-tagatose catalyzed by a multienzyme cascade. (a) CaFRK dosage optimization (based on 0.01 mg·mL−1 BaSP, 0.05 mg·mL−1 CaF6PE, 0.05 mg·mL−1 MmT6PP, and 0.04 mg·mL−1 DrPPK); (b) CaF6PE dosage optimization (based on 0.01 mg·mL−1 BaSP, 0.02 mg·mL−1 CaFRK, 0.05 mg·mL−1 MmT6PP, and 0.04 mg·mL−1DrPPK); (c) MmT6PP dosage optimization (based on 0.01 mg·mL−1 BaSP, 0.02 mg·mL−1 CaFRK, 0.05 mg·mL−1 CaF6PE, and 0.04 mg·mL−1 DrPPK). The error bar represents the standard deviation of triplicate measurements.
Figure 5.
Optimization of enzyme dosage for the synthesis of d-tagatose catalyzed by a multienzyme cascade. (a) CaFRK dosage optimization (based on 0.01 mg·mL−1 BaSP, 0.05 mg·mL−1 CaF6PE, 0.05 mg·mL−1 MmT6PP, and 0.04 mg·mL−1 DrPPK); (b) CaF6PE dosage optimization (based on 0.01 mg·mL−1 BaSP, 0.02 mg·mL−1 CaFRK, 0.05 mg·mL−1 MmT6PP, and 0.04 mg·mL−1DrPPK); (c) MmT6PP dosage optimization (based on 0.01 mg·mL−1 BaSP, 0.02 mg·mL−1 CaFRK, 0.05 mg·mL−1 CaF6PE, and 0.04 mg·mL−1 DrPPK). The error bar represents the standard deviation of triplicate measurements.

Figure 6.
Optimization of reaction conditions for the synthesis of d-tagatose catalyzed by a multienzyme cascade. (a) Temperature; (b) pH; (c) different PBS concentrations with the fixed concentration of Tris-HCl buffer (50 mM); (d) metal ion, Mix1: Mg2+ (2 mM) + Mn2+ (2 mM), Mix2: Mg2+ (2 mM) + Co2+ (2 mM), Mix3: Mn2+ (2 mM) + Co2+ (2 mM); (e) metal ion concentration; (f) ADP:sucrose ratio. The error bar represents the standard deviation of triplicate measurements. A 1 mL reaction mixture contained 10 mM sucrose, 0.01 mg·mL−1 BaSP, 0.02 mg·mL−1 CaFRK, 0.05 mg·mL−1 CaF6PE, 0.2 mg·mL−1 MmT6PP, and 0.04 mg·mL−1 DrPPK.
Figure 6.
Optimization of reaction conditions for the synthesis of d-tagatose catalyzed by a multienzyme cascade. (a) Temperature; (b) pH; (c) different PBS concentrations with the fixed concentration of Tris-HCl buffer (50 mM); (d) metal ion, Mix1: Mg2+ (2 mM) + Mn2+ (2 mM), Mix2: Mg2+ (2 mM) + Co2+ (2 mM), Mix3: Mn2+ (2 mM) + Co2+ (2 mM); (e) metal ion concentration; (f) ADP:sucrose ratio. The error bar represents the standard deviation of triplicate measurements. A 1 mL reaction mixture contained 10 mM sucrose, 0.01 mg·mL−1 BaSP, 0.02 mg·mL−1 CaFRK, 0.05 mg·mL−1 CaF6PE, 0.2 mg·mL−1 MmT6PP, and 0.04 mg·mL−1 DrPPK.

Figure 7.
Time course of d-tagatose and d-fructose conversion using 10 mM sucrose as a substrate. A 2 mL mixture containing BaSP (0.01 mg·mL−1), CaFRK (0.02 mg·mL−1), CaF6PE (0.05 mg·mL−1), MmT6PP (0.2 mg·mL−1), DrPPK (0.04 mg·mL−1), 50 mM Tris-HCl + 10 mM PBS (pH 7.5), 10 mM sucrose, 4 mM MnCl2, 2 mM sodium hexametaphosphate, and 1 mM ADP was incubated at 45 °C.
Figure 7.
Time course of d-tagatose and d-fructose conversion using 10 mM sucrose as a substrate. A 2 mL mixture containing BaSP (0.01 mg·mL−1), CaFRK (0.02 mg·mL−1), CaF6PE (0.05 mg·mL−1), MmT6PP (0.2 mg·mL−1), DrPPK (0.04 mg·mL−1), 50 mM Tris-HCl + 10 mM PBS (pH 7.5), 10 mM sucrose, 4 mM MnCl2, 2 mM sodium hexametaphosphate, and 1 mM ADP was incubated at 45 °C.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of enzyme activity in a multienzyme cascade catalytic reaction.
| Enzyme Name | NCBI Accession Number | Specific Activity (U·mg−1) |
|---|---|---|
| BaSP | WP_011742626.1 | 84 ± 2 |
| CaFRK | KHD36265.1 | 113 ± 3 |
| CaF6PE | WP_014433578 | (2 ± 0.5) × 10–2 |
| MmT6PP | WP_013296249.1 | (1 ± 0.2) × 10–1 |
Table 2.
Summary of the conversion of d-tagatose at different ADP concentrations from sucrose [a].
| Sucrose (mM) | ADP (mM) | Conversion (%) | Production (g·L−1) | Space–Time Yield (g·L−1 h−1) | S:C [b] |
|---|---|---|---|---|---|
| 50 | 0.1 | 61.7 | 5.6 | 0.23 | 18.5 |
| 50 | 0.25 | 62.8 | 5.7 | 0.24 | 18.5 |
| 50 | 0.5 | 72.4 | 6.5 | 0.27 | 18.5 |
[a] Reaction system: sucrose (50 mM), sodium hexametaphosphate (10 mM), ADP (0.1, 0.25, and 0.5 mM), MnCl2 (20 mM), BaSP (0.025 mg·mL−1), CaFRK (0.05 mg·mL−1), CaF6PE (0.25 mg·mL−1), MmT6PP (0.5 mg·mL−1), DrPPK (0.1 mg·mL−1), the dosage of all enzymes was equal to 0.925 mg·mL−1, buffer (50 mM Tris-HCl + 10 mM PBS, pH 7.5); the reaction was performed at 45 °C for 24 h. [b] S:C of the system means substrate:catalyst (g/g).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, S.; Tu, W.; Ni, Y.; Guo, Y.; Han, R. Novel In Vitro Multienzyme Cascade for Efficient Synthesis of d-Tagatose from Sucrose. Catalysts 2023, 13, 1515. https://doi.org/10.3390/catal13121515
AMA Style
Liu S, Tu W, Ni Y, Guo Y, Han R. Novel In Vitro Multienzyme Cascade for Efficient Synthesis of d-Tagatose from Sucrose. Catalysts. 2023; 13(12):1515. https://doi.org/10.3390/catal13121515
Chicago/Turabian StyleLiu, Shuangyu, Wenyu Tu, Ye Ni, Yuanyi Guo, and Ruizhi Han. 2023. "Novel In Vitro Multienzyme Cascade for Efficient Synthesis of d-Tagatose from Sucrose" Catalysts 13, no. 12: 1515. https://doi.org/10.3390/catal13121515
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.