1. Introduction
Benzylisoquinoline alkaloids (BIAs) are a family of secondary metabolites with a wide variety of pharmaceutical applications. Important examples of this family are the opioid analgesics morphine and codeine, the antitussive noscapine or the antispasmodic drug papaverine [
1]. Opioids are classified as essential medicines by the World Health Organization (WHO), due to their usefulness in reducing pain for trauma patients and in palliative care [
2], with 10 billion standard daily doses consumed per year [
3]. For industrial production, typically a largely mechanized process is used in which fully mature plants are harvested, dried and then pulverized, before extracting the active component [
4], which is then refined for use in pharmaceuticals. Environmental factors such as pests, diseases, and extreme weather events make poppy supply chains unstable, threatening the secure availability of the medicines derived from them. Therefore, there is great interest in developing efficient and scalable production platforms that would not only secure supply but also overcome the capacity limitations of the plant-based production.
More than 30 chemical and chemo-enzymatic synthesis pathways of morphine and derivatives are known, but none of them could be developed to the point where industrial production at market prices would be possible, as the synthesis of these chiral molecules is very complex and currently not cost-competitive [
5]. The first full biosynthesis of opioids in yeast was published in 2015 but was also not able to deter industrial production away from using the opium poppy as the raw material, due to the very low titers achieved [
6]. In the same year, an elegant biocatalytic route to the important BIA scaffold structure (S)-norlaudanosoline ((S)-NLS) was published [
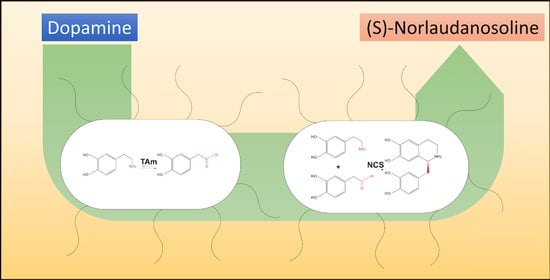
7], which involves two enzymatic transformations and the affordable starting materials dopamine (DA) and pyruvate, as shown in
Scheme 1. Since (S)-NLS can be diversified into various target structures, including the pharmaceuticals morphine and codeine or the neuromuscular-blocking drug cisatracurium [
1,
8,
9], the efficient industrial-scale production of this molecule would represent a milestone on the way to a plant-independent synthesis platform.
In the two-step enzymatic synthesis of (S)-NLS, dopamine is first converted to 3,4-dihydroxyphenylacetaldehyde (DOPAL) at the expense of pyruvate using transaminase (EC 2.6.1.B16), which catalyzes the transfer of an amine group from an amine donor to either a ketone or aldehyde with 5′-pyridoxalphosphate (PLP) as the amine shuttling cofactor [
10]. A transaminase subgroup, the so-called ω-transaminases (TAms), use non-α position amino acids or amine compounds without carboxylic group as donors and have been studied intensively due to their high relevance for the production of chiral molecules, e.g., in fine chemistry and the pharmaceutical industry. The TAm-catalyzed reactions are generally reversible [
10].
In the triangular cascade to (S)-norlaudanosoline, the second step is catalyzed by norcoclaurine synthase (NCS, EC 4.2.1.78), which combines dopamine and the formed aldehyde, thereby creating the BIA core structure. NCS, which is found in plants, catalyzes a stereoselective Pictet–Spengler cyclisation, where arylethylamine and aldehyde/ketone condense followed by a ring closure.
One general problem of the reaction is the oxidative degeneration of DA and DOPAL, which can be reduced by the exclusion of oxygen [
11,
12,
13]. Another challenge is the inherent instability of DOPAL and its tendency to create polymers with the surrounding molecules via a radical reaction [
11], often creating black eumelanin [
14]. This can be mitigated by adding the antioxidative ascorbate to the reaction solution [
15].
A spontaneous reaction of DA and DOPAL to racemic NLS can be observed in the presence of several conventional buffer ions, for example, phosphate. This unwanted spontaneous reaction can be minimized by using only specific buffers, for example, HEPES buffer, and by keeping the DOPAL (or theoretically DA) concentration low [
11,
16,
17]. Here, the triangular cascade synthesis of (S)-NLS using TAm and NCS is beneficial since it generates and utilizes the reactive aldehyde DOPAL in situ and achieved yields of up to 87% using (partially) purified enzymes [
7].
To further optimize the biocatalytic synthesis of (S)-NLS from dopamine and pyruvate and its cost effectiveness, the use of whole cells instead of (partially) purified enzymes and the establishment of a continuous process in flow are two options. While the use of whole cells lowers the cost of the catalyst preparation, continuous processing can intensify biocatalytic reactions, thereby reducing the size of the process equipment, improving product quality and lowering energy and solvent consumption, as well as the waste generated [
18]. Ultimately, all these aspects reduce the footprint of the process and might also improve its safety [
19]. While diverse TAm reactions (e.g., [
20,
21,
22]) and also the NCS reaction [
15] have been successfully performed in flow setups, the triangular synthesis of (S)-NLS has not been investigated in flow so far.
The aim of this work was to elucidate whether the asymmetric two-step synthesis of (S)-NLS can be performed with whole-cell catalysts and whether the joint implementation of the TAm and NCS reactions in a continuous flow system provides access to a more efficient synthesis of the BIA scaffold structure. Possible benefits could be a higher space–time yield, a reduced spontaneous background reaction, and thus, a higher optical purity of the product, as proposed by Lechner et al. [
15]. Furthermore, simple 3D-printed columns should be used as fixed-bed reactors. The method of 3D printing has long been used in the manufacturing industry to produce design prototypes and accelerate developments, made possible by the design freedom inherent in additive manufacturing in combination with the low production costs [
23], and we wanted to take advantage of these benefits in this study as well.
2. Results
To investigate the asymmetric two-step enzymatic synthesis of the BIA scaffold structure (S)-NLS with whole cell catalysts in batch biotransformations and flow setups, two recombinant
E. coli strains expressing the genes for the enzymes TAm and NCS were cultivated separately. Subsequently, the cells were washed and lyophilized. Both biocatalysts showed good expression levels of the target proteins (
Supporting Figures S1 and S2). Typical specific activities of TAm-containing cells were in the range of 30 U per gram cell dry mass. Due to the high (non-specific) reactivity of the aldehyde DOPAL, the specific activity of NCS cells was not determined directly with the substrate in excess, but as combined activity in batch experiments to identify the optimal cell ratio.
2.1. Batch Biotransformations
To achieve the maximum enzymatic activity and to test the interaction of both enzymes, different mass ratios of lyophilized TAm- and NCS-containing cells were combined into mixtures. The different cell mixtures were suspended in the reaction buffer, and the reactions were performed in miniaturized stirred-tank reactors at the 12 mL scale. To minimize the oxidation and subsequent polymerization of substrates and intermediates, ascorbate was used, and the reactors’ headspaces were gassed with nitrogen. The antibiotic gentamicin was added to prevent bacterial growth, e.g., on pyruvate as the carbon source. The enzyme activity of TAm and NCS in the cell mixtures was measured in tandem, as the NCS reaction is dependent on the supply of the intermediate DOPAL via TAm. The TAm (1st cascade reaction) activity is in turn increased by the presence of NCS (2nd cascade reaction), which uses DOPAL in its reaction. To obtain results that are comparable to those of the cell-free enzyme system used by Lichman et al. [
7], samples were taken after 3 h. Measurements were done for (S)-NLS and (R)-NLS, to determine the yield (referring to dopamine) and enantiomeric excess (ee), as well as for alanine and pyruvate, to gain a measure of the TAm activity, as summarized in
Table 1.
The central finding of this experiment was the fact that mainly the mass of NCS-containing cells in the mixture was crucial for the yield, while the TAm-containing cell mass above a minimal amount nearly did not factor in. In the assays with only 12 mg of TAm-containing cells, residual pyruvate concentrations of 9 mM, 11 mM and 18 mM were found. Increased amounts of TAm led to a complete pyruvate depletion during the reaction. Pyruvate is used up much more than would be expected by the reaction scheme. This has been anticipated as a necessary cost of using intact cells, which still contain metabolic enzymes. These can metabolize some of the pyruvate as it is a central part of their carbon metabolism [
24]. Although gentamicin inhibits the protein biosynthesis and prevents the growth of bacteria, metabolic enzymes already present in the cells can still degrade the carbon source pyruvate. This was also shown in a previous experiment, where tests with 25 mM pyruvate and without gentamicin led to a complete pyruvate depletion in all samples. The increasing residual pyruvate concentration with the increasing concentration of NCS-containing cells could indicate an inhibiting effect of the final product on the cellular metabolism of
E. coli, preventing the additional use of pyruvate as a carbon source.
While increased amounts of TAm-containing cells with a given amount of NCS-containing cells led to only an insignificant increase in the total yield in two of the three assays, for 120 mg of NCS-containing cells, higher amounts of TAm led to an even lower yield (from 67 over 62 to 54%). This could indicate a negative effect of DOPAL accumulation created by excess TAm. The synergy between the catechol and aldehyde groups of DOPAL strongly enhances its reactivity, possibly leading to enzyme inactivation by modification of functional protein residues, protein cross-linking and protein aggregation [
25].
Increased amounts of NCS-containing cells in the reaction setup resulted in higher yields: 12 mg of catalyst gave 11% yield, while 66 mg resulted in 35%. In the experiments with 36 mg of TAm-containing cells, the yield seemed to linearly increase with the NCS-containing cell mass until reaching 91% at 228 mg. Thereafter, the yield stopped increasing steadily. The observed maximum yield of about 90% could be due to product inhibition of NCS, which could be analogous to the inhibitory effect of (S)-norcoclaurine detected in an earlier study [
15]. This trihydroxylated compound represents the BIA scaffold structure occurring in plant BIA syntheses and differs from the tetrahydroxylated (S)-NLS by the absence of a hydroxy group at the isoquinoline moiety.
In general, ee followed the trend of the yield. An increasing amount of NCS in the reaction mixture led to an increase in enantioselective enzyme activity compared to the non-selective spontaneous Pictet–Spengler reaction and thus to a higher optical purity. Due to the single measurements, the effects that were observed with very high cell concentrations are of limited significance. Fluctuations in the yield and ee could also be due to measurement uncertainties. This implies that amounts of the NCS catalyst higher than 228 mg do not reliably improve the output under these conditions. The mixture of 36 mg TAm-containing and 228 mg NCS-containing cell mass, corresponding to a mass ratio of 3:19, seemed optimal, although as shown by the pyruvate depletion, even lower ratios of TAm to NCS cells could also be successful.
Compared to the study performed by Lichman et al. [
7], who used
E. coli cell extract for TAm activity in combination with purified NCS enzymes, slightly higher yields were reached in this work. At substrate concentrations of 50 mM DA and 25 mM pyruvate, an 85% conversion (referring to DA) was reported after 3 h with a product ee of 96% (S). Due to the differences in catalyst preparations, the enzyme loadings are hard to compare. However, this result underlines that cheaply producible whole-cell catalysts are suitable for use in this two-step enzyme reaction and there is no need for enzyme purification. Whole cells were also employed in the study performed by Lechner et al. [
15], who investigated the NCS-catalyzed reaction of DA and 4-hydroxyphenylacetaldehyde to (S)-norcoclaurine. At a catalyst concentration of 10 mg mL
−1, 27% conversion with >98% ee was obtained. This catalyst loading corresponds well to the experiments performed in this study, with 120 mg NCS-containing cells in a reaction volume of 12 mL, where yields of ≥54% with 90–99% ee were obtained, depending on the amount of TAm used. Although the syntheses are not directly comparable because of the different enzymatic reactions, the mass transport across two cell walls, which is necessary due to the use of catalysts with separately expressed TAm and NCS genes, does not seem to be severely rate-limiting since the reaction is faster than the one-step biotransformation to (S)-norcoclaurine, where the substrates must cross only one cell wall. This is supported by the fact that the enzymatic activities of NCS with DOPAL and 4-hydroxyphenylacetaldehyde, respectively, are typically in the same order of magnitude [
26].
The comparison with other data from the literature is difficult in general as early reports either (1) used phosphate buffer leading to significant amounts of racemic NLS [
16,
27,
28], (2) aim at slightly different BIAs such as norcoclaurine [
15,
29] or (S)-reticuline [
30,
31,
32] or (3) only use a single-enzyme reaction [
15,
20,
33].
2.2. Whole Cell Catalyst Immobilization on Carriers
To establish the two-step enzymatic reaction using cells in a flow setup, the cell mixture must be immobilized in the reactor, e.g., on a suitable carrier material. Immobilization has several advantages pertaining to (1) product purification, as less biomass must be removed, (2) reusability, as enzymes can theoretically be used until fully degraded, and (3) volumetric activity, as higher local cell concentrations can theoretically be created [
34,
35,
36]. Thus, the immobilization of the cell mixture on carrier particles was investigated.
Five distinct types of catalyst particles were tested. Diaion HP-2MG represents methacrylic ester copolymer particles, which are hydrophilic and allow cells to adsorb to their surface. Two different methods of cell adsorption on Diaion particles were assessed here: (1) The incubation of particles with suspended lyophilized cells (Diaion incubated) or (2) the addition of particles during the growth of cells expressing the enzymes (Diaion overgrown). The dried particles with cells adsorbed on them were subsequently mixed. (3) As an even cheaper adsorber material, glass beads were selected and also incubated with the lyophilized cells [
37]. For an established encapsulation option, calcium alginate beads were chosen. The high porosity of cross-linked alginate combined with the potential to use the full particle rather than just the surface implies the possibility of achieving high volumetric reaction rates, especially compared to the adsorption-based particles. The alginate beads were created using two types of preparation: (4) pure calcium alginate beads (uncoated alginate) or (5) calcium alginate beads additionally coated twice, first with polyDADMAC, which binds as cation to the anionic alginate on the bead surface, and subsequently in sodium-alginate, which binds to the polyDADMAC now on the surface, creating layers that are linked together at multiple sites (coated alginate).
For the five different types of particles loaded with the (3:19) mixture of TAm and NCS cells, respectively, 1.5 g of each were added to the miniaturized stirred-tank reactors containing 12 mL reaction medium in a batch setup. Headspace aeration with nitrogen was no longer carried out, as ascorbate was found to be sufficient to prevent oxidation, while contact with oxygen was still minimized for the solutions. The setup and reaction medium were otherwise kept the same as with the screening of cell ratios.
The yields obtained with the different preparations of the immobilized cells are summarized in
Table 2. Preparations with glass did not lead to significant product formation (0.9% yield) and thus no ee could be determined. The optical purities obtained with all other catalyst particles were in the range between 89 and 97%. Due to measurement uncertainties, the differences between the ee values cannot be considered statistically significant. The selection of the catalyst preparation for further experiments was therefore based mainly on the yield obtained. The second lowest yield was observed for uncoated alginate particles (19.9%), followed by coated alginate particles (33.5%). The use of incubated Diaion led to a yield of 52.4%, whereas overgrown Diaion particles led to an even higher yield of 64.1%, corresponding to 55 mg of product. The addition of ascorbate and minimization of oxygen contact seemed to prevent the formation of insoluble polydopamine and eumelanin derivatives observed in other studies [
20], as no blackening of any solution was observed.
While the cellular dry mass for the experiment with glass was below the limit of detection, the value for the overgrown Diaion could not be determined because the proliferation of cells during preparation did not allow for a mass balance. When comparing the cellular dry mass for the other three immobilizations, it was lowest with uncoated alginate at 0.06 ± 0.01 mg per batch of 1.5 g carriers, followed by coated alginate at 0.14 ± 0.03 mg, while incubated Diaion HP2MG fixed 0.35 ± 0.03 mg cell dry mass. A slight turbidity, later identified by microscopy as intact cells, occurred in the solution of alginate particles suggesting that the alginate particles lose cells during the reaction. This could indicate a constant loss of catalyst in the solution of alginate particles through a combination of dissolved alginate matrix and cells slowly washing out through openings in the matrix. Binding E. coli on Diaion seemed to be comparatively more stable and no turbidity was observed.
Although glass beads seem to be a bad choice for this type of use, they are still the cheapest option of the investigated particle types and thus should not be easily discounted. To increase adsorption and thus yield in future studies, (1) beads could be overgrown as was done with Diaion in this study and (2) surfaces could be covered by a thin coating of Fe
2O
3 or other metal oxides, to increase the adsorption strength [
38] or (3) cells could be linked covalently by aminopropyltrimethoxy silane and glutaraldehyde to the carrier to increase cell density [
39].
The yield was related to the determined cellular dry mass to make statements about the productivity of the cells in the different batches. Here, uncoated alginate scored highest (1.52 mgS-NLS gCDM−1 h−1), followed by coated alginate (1.22 mgS-NLS gCDM−1 h−1) and incubated Diaion (0.72 mgS-NLS gCDM−1 h−1). While productivities were even higher in alginate, this is to be expected, as the lower amount of biomass in these preparations led to lower product accumulation and thus lower product inhibition, but also a lower space–time yield, which was deemed paramount for a later application.
Though the exact productivity values could not be calculated for overgrown Diaion particles, as it is unknown how much cell mass is deposited during incubation, they were found to reach the highest yield combined with an acceptable ee of 95% and were thus chosen as the biocatalyst for subsequent experiments.
To better interpret the higher yield of overgrown Diaion compared to incubated Diaion, both particle populations as well as untreated Diaion particles and Diaion particles left to hydrate in 0.9% NaCl for one day were measured using a Mastersizer 2000 (
Table 3). Sizes of wet Diaion (d (0.5) = 580 ± 2 µm), incubated Diaion (d (0.5) = 598 ± 4 µm) and overgrown Diaion (d (0.5) = 546 ± 7 µm) were different; the overgrown Diaion particles were smaller. The relatively lower diameter could be due to the abrasion of the particles by the shaking during cultivation.
2.3. Preliminary Experiments in Packed-Bed Flow
Having established an optimum ratio of TAm-containing to NCS-containing cells and having chosen overgrown Diaion particles as the best catalytic preparation, a preliminary test in a flow setup was carried out. The flow setup could be advantageous over a batch setup as it could allow production over a longer time scale than batch setups, if the enzyme activity can be maintained, while still keeping concentrations of DA and DOPAL relatively low. This in turn minimizes the unselective background reaction of DA and DOPAL to racemic NLS, which can occur spontaneously, and also reduces the risk of substrate polymerization to polydopamine or eumelanin derivatives.
For the preliminary experiment, a 5 mL-reactor (reactor A) with typical cylindrical geometry was designed and additively manufactured. To facilitate manual filling and assembly, the column was printed in two parts and complementary screw threads were added to both parts. For the inlet and outlet, the form of a male Luer-Lock connection was used, to allow an easy connection with the periphery.
Before both reactor parts were screwed together, the inside was carefully filled with the overgrown Diaion particle mixture creating the particle bed. The reactor was then connected to tubes, placed in a temperature-controlled incubator at 37 °C and was filled with HEPES buffer. During the reaction, a HPLC pump was used to continuously feed substrate at a flow rate of 0.5 mL min−1, while a fraction collector was used to collect the outflowing fluid. The first fraction contained the outflow of the first 10 min (5 mL), while each subsequent fraction contained the outflow of 20 min (10 mL). Samples were taken from each fraction and the concentrations of both NLS enantiomers were measured to calculate the yield and ee.
After the reaction, the particle mass was measured again to be able to compute porosity. The bed volume was determined to be 4.26 mL accommodating 0.47 g of Diaion particles, which corresponds to 0.11 g cm
−3. The total fluid volume of the filled column A was determined as 1.6 mL resulting in a porosity of 37% (Φ = 0.37). This corresponds well with the literature values for dense random packing (Φ = 0.359–0.375) and thus, this column packing was deemed acceptable [
40].
The advantage of additive manufacturing is the fast and inexpensive design of prototypes for preliminary experiments to clarify individual questions. The small column was used to investigate if any adverse effects of the “Figure 4® PRO-BLK 10” plastic were detected (which was not the case) and if the immobilized cell preparation appears to be sufficiently stable in the flow experiments to allow for larger-scale experiments needing more material. From the combination of the flow rate and the hold-up volume of the column, a residence time of 3.2 min was calculated. Compared to the batch experiment with Diaion (overgrown), which was performed with 3-times more catalyst and a 56-times longer reaction time, a much lower yield than 64.1% (
Table 2) was expected. As shown in
Figure 1, a yield of around 6% was reached over 80 column volumes (CV), with a consistent ee of 97% and an overall productivity of more than 118 mg
(S)-NLS g
CDM−1 h
−1, assuming that these particles had at least the same cell loading as the incubated Diaion.
The yield dropped from about 6.5% at the start to 5.8% at CV 80, corresponding to a trend of constantly decreasing yield. Over the time course of the experiment of 5 h, the product concentration measured at the outlet decreased by 18%, pointing to the fact that the system stability is not high enough for long-time operation. To further substantiate this finding, a confirmational large scale-experiment was conducted.
2.4. Rescaled Packed Bed Reactor
To create comparability to previous experiments performed in batch mode, a residence time of about 180 min was envisioned, which would correspond to a 60-fold increase in contact time compared to the preliminary experiment with a residence time of 3.2 min. This was achieved by lowering the flow rate from 0.5 mL min−1 to 0.1 mL min−1, which was the minimum flow rate compatible with the fraction collector and designing a column with a 12-times increased inner volume of 48 mL. A cone shaped part was added to the inlet and outlet section, to accommodate flow through the wider main cylindrical column part. The total bed volume was determined to be 51.1 mL, which was filled with 15.05 g of a 1:2 mixture of overgrown Diaion particles carrying TAm or NCS and untreated Diaion particles carrying no cells. For an industrial application, of course, the column should be filled with active material only and the flow rate should be increased proportionally, while minding the increased pressure. This was only carried out in this way to cope with the system restriction regarding the minimum flow rate without using too much catalyst. Nevertheless, a 3.7-fold higher catalyst load was used compared to the batch process performed with Diaion (overgrown) (5.5 g versus 1.5 g). The total fluid volume was computed as 18.9 mL, resulting in an average residence time of 189 min at a flow rate of 0.1 mL min−1.
Figure 2 shows the progression of the yield of the reaction and the ee of the product as a function of column volumes. After replacing NaCl in the void volume of the column with a substrate solution during CV 1, the yield rose rapidly to 88.3%. This is higher than the yield obtained in the batch process with Diaion (overgrown), which can be explained by the higher amount of catalysts in the system. The yield stayed stable for about 1–2 CVs (~3–6 h). Afterwards, the yield decreased consistently and with increasing intensity until the end of the experiment to 58%. This means a proportional loss of yield of about 34% from CV 1.1 to 4.3. The achieved ee was consistent at 94 ± 4%. The system achieved a production of 318 mg of (S)-NLS, corresponding to an overall productivity of more than 29 mg
(S)-NLS g
CDM−1 h
−1, assuming that these particles had at least the same cell loading as the incubated Diaion. The deducted productivity dropped 4-fold compared to the small column, mainly because more biomass was used over a longer run-time of the experiment. Analyzing the data further, the observed trend was best fitted as an increasing exponential decay, which seemed unusual. An extrapolation showed that the yield would approach zero after 5.5 CVs of active production, confirming the finding that this setup is not suitable for long-term operation.
MTT-testing in the collected fractions showed a continuous loss of biomass from CV 1 when starting at 0.4 mg mL
−1 at CV 1 and increasing to 2 mg mL
−1 at CV 4. This resulted in a loss of 171 mg of biomass over the whole experiment, assuming the same cell loading for these particles as it was determined for incubated Diaion. This corresponds to a loss of at least 16% of biomass over the course of the reaction as shown in
Figure 3.
Other factors that were considered as possibly contributing to this loss of yield could be the loss of substrate due to polymerization and catalyst inactivation. Polymerization seems unlikely, since these effects should be prevented by the addition of ascorbate to the substrate solution and the mechanical barrier of the reactor walls, preventing direct contact with the atmosphere. Also, the substrate and product fluid were monitored for the emergence of black precipitates, which would indicate such reactions, but were not found. The stability of the biocatalysts was tested by incubating cells at 37 °C (without substrate) before their use in the reaction. In this experiment, a loss of activity of 16% after 24 h was observed, increasing to a loss of 48% activity after 48 h (
Supporting Figure S3). This indicates that the loss of activity is connected to enzyme inactivation. After four CVs (meaning 12 h), this could account for about 10% of the 25% yield loss, but not for all of it. This catalyst inactivation, in conjunction with the wash out of cellular material from the particles, could be the reason for the observed loss of activity. As the wash out was detected via an MTT-test and assuming that the loss of activity correlates directly to the loss of cellular material, a loss of 16% yield could be due to wash out. This seems the most likely explanation at least for the bulk of the activity loss that was observed.
Concerning experimental options to increase the performance, the enzyme degradation occurring over time might be due to the fact that NCS is susceptible to protease attacks originating from the
E. coli cells. In this case, the identification of the responsible protease(s), for example, by a screening of deletion mutants, and the use of the corresponding knock-out strain for protein expression might increase the performance greatly [
41], via a better long-term stability of the enzymes involved in the synthesis.
It would also be of interest to measure the maximum amount of cells washed out of the immobilization particles and to examine how many cells remain bound on the Diaion carriers, as it is possible that only the extra outer layers of bacterial cells that adsorb onto already bound cells and not directly onto the carrier itself are more easily washed away under the pressure of the column flow. If so, a “single layered” particle would create less yield loss during use. The resulting reduction of the cell number per particle could then be compensated for by increasing the amount of enzymatically active particles per column.
To increase the adsorption strength of and to decrease the wash-out from Diaion particles, the
silCoat technology proposed by Findeisen et al. could be adapted [
42]. Solid carriers are coated with a silicon layer containing cells or enzymes, thus reducing the wash-out. Alternatively, thin coatings with metal oxides or covalent linking, as proposed for glass beads, could also be utilized.
While the separation of enzyme-containing cells from Diaion particles and their subsequent wash-out as well as enzyme inactivation were identified as key challenges preventing a long-time use of the column system, it already produced, in its current state, a decent amount of (S)-NLS.
3. Materials and Methods
3.1. Chemicals, Medium and Buffers
All chemicals, Diaion HP-2MG (13601 Supelco Diaion®® HP-2MG), glass beads (18406–500G, diameter ~5 mm) and sodium alginate (W201502) were obtained from Merck KGaA (Darmstadt, Germany). Polydiallyldimethylammonium chloride (short: PolyDADMAC, Polyquat 40 U50 A) was obtained from Katpol-Chemie GmbH (Bitterfeld, Germany). For lysogeny broth (LB)-medium, 10 g L−1 tryptone, 10 g L−1 NaCl, 10 g L−1 yeast extract were dissolved in deionized water (DIW). The pH was adjusted to 6.8. The buffers used in this study are HEPES buffer (50 mM, pH 7.4) and MOPS buffer (10 mM MOPS, pH 7.0). Adjustments of pH were carried out with NaOH and HCl if needed.
3.2. Expression of Proteins
Table 4 gives an overview of the proteins recombinantly produced in
Escherichia coli in this study.
A plasmid encoding the NCS from
Papaver somniferum (Ps_NCS2) was kindly donated by the working group of Prof. Wolfgang Kroutil (Institute of Chemistry, University of Graz, Graz, Austria). The gene encoding TAm (CV_2025) was synthesized by Eurofins Genomics (Ebersberg, Germany) and codon optimized for the expression in
E. coli (using the GENEius tool of Eurofins Genomics). Both coding regions were cloned into pET28a(+) vectors and expressed separately in
E. coli BL21(DE3) (Novagen, Darmstadt, Germany). For the recombinant production of NCS, the cells were co-transformed with the plasmid pG-Tf2 from the Takara chaperone plasmid set (Takara Bio inc., Shiga, Japan). This plasmid bears the genes
groES-groEL-tig, which code for the GroES/GroEL chaperonins and the trigger factor. In a previous study, it has been shown that the simultaneous expression of chaperones had a positive effect on the specific NCS activity of the recombinant cells [
15].
LB-medium supplemented with 50 µg mL−1 kanamycin was used for (pre)culture. For the cells co-transformed with the pG-Tf2 plasmid, additionally, 25 µg mL−1 of chloramphenicol was used. The precultures were incubated overnight at 150 rpm at 37 °C in an INFORS HT Multitron incubator (50 mm shake stroke, Infors AG, Bottmingen). The main cultures were inoculated with the preculture to reach an optical density at 600 nm (OD600) of 0.1.
The main cultures were grown at 150 rpm and 37 °C in 2 L shake flasks without baffles filled with 400 mL media and 5 ng mL−1 tetracycline when NCS was to be expressed, until an OD600 of 0.5–0.7 was reached. Expression of NCS and TAm was started by adding 0.5 mM isopropyl-β-D-thiogalactopyranoside (IPTG). Cells were then grown for 16 h at 20 °C. Cells were harvested via centrifugation for 30 min, 4500 g, 4 °C, washed with HEPES buffer with 20% (v/v) glycerol and stored at −20 °C until use. For lyophilization, cells with TAm or NCS were pelleted and frozen in HEPES buffer at −20 °C for one day before freeze-drying at −80 °C for three days or until completely dry.
3.3. Batch Biotransformations
To investigate the whole cell biotransformations in batch and to identify the optimum ratio of TAm- and NCS-containing
E. coli biomass, different ratios of the lyophilized whole cell preparations were weighed with a precision balance and added to 12 mL of substrate solution containing 50 mM pyruvate, 50 mM dopamine and 4 g L
−1 ascorbic acid in 50 mM HEPES buffer (pH 7.4). The reactions were performed in miniaturized stirred-tank reactors (bioREACTOR48) from 2mag AG (Munich, Germany) equipped with S-type stirrers driven at 500 rpm [
43]. The reactor temperature was set to 37 °C, while headspace cooling (15 °C) reduced evaporation. Prior to the reaction, the headspace was flushed with nitrogen and during the reaction, >0.1 bar of nitrogen was attached to minimize oxidation. Sealing tape was used to seal contact surfaces. The sampling aperture was closed with a fitting PTFE plastic plug except during sampling. Yield was measured 3 h after starting the reaction.
3.4. Immobilization of Lyophilized Cells
Lyophilized cells were mixed in a ratio of 3 g TAm-containing cells to 19 g NCS-containing cells, representing the optimum ratio determined during the batch experiments. Different materials and methods were used for subsequent immobilization. While the immobilization of lyophilized cells on the medium polarity adsorbent resin Diaion HP-2MG and glass beads followed a single protocol, the confinement of cells in alginate was performed with and without additional coating with PolyDADMAC. After immobilization, Diaion and glass-based particles were lyophilized before use, while alginate particles were not.
3.5. Incubated Diaion and Glass Beads
For Diaion (incubated) and glass, the method used by Findeisen et al. was adapted [
42]. Thus, 22 g L
−1 of mixed catalyst was added to 30 mL (final volume) HEPES buffer containing 25 µg mL
−1 gentamicin and 50 g L
−1 of carrier. The dispersion was incubated for 16 h at 20 °C. The supernatant was removed, and the particles were washed twice with 50 mL HEPES buffer. Cellular dry mass immobilized on the particles was determined by separate lyophilization of supernatant and incubated particles. Added mass on the particles and reduced dry mass of the supernatant fraction were assessed as biological loading on the particles.
3.6. Uncoated Alginate
For uncoated alginate, 11 g L−1 of the mixed cells was dispersed into MOPS buffer. Subsequently, 10 g L−1 of sodium alginate was added. Beads were produced by adding droplets of the solution to a stirred (300 rpm, magnetic stirrer) 100 mM CaCl2, 10 mM MOPS (pH 7) solution using the Encapsulator Medical IEM-40 2 (nozzle 300, pump speed 6.8 mL min−1, 677 Hz) (Inotech, Nabburg, Germany). Particles were incubated for 30 min at 25 °C before washing with 50 mL MOPS buffer. Cellular dry mass in the particles was determined by dissolution of particles in a covering solution of 0.5 M citric acid for 8 h. The fluid was centrifuged, resuspended in 50 mL MOPS buffer, and centrifuged again. The pellet was lyophilized and weighed.
3.7. Coated Alginate
Coated alginate particles were created by using alginate particles (created as described above), which subsequently were incubated in 50 mL of 1% PolyDADMAC. Particles were stirred (300 rpm) for 30 min at 25 °C, before washing with 50 mL MOPS buffer. The incubation was repeated in a solution of sodium alginate (1%), then the washing was done again. Finally, the particles were incubated in 1% PolyDADMAC as before and subsequently washed again. Movement of the particles between containers and washing was done with a sieve.
3.8. Immobilization of Growing Cells
The Diaion adsorbent resin was also used in another protocol in which the cells were immobilized by adding the carrier material to the growing cells to achieve biofilm formation [
42]. For the preparation of Diaion (overgrown), 0.5 g of Diaion HP-2MG particles was added at the time point of induction to 400 mL of bacterial culture.
3.9. Column Printing
The 3D-printing was done with a stereolithographic “Figure 4® Modular” printer (3D Systems, Moerfelden-Walldorf, Germany) using “Figure 4® PRO-BLK 10” biocompatible plastic (ISO10993-5 [
44], ISO10993-10 [
45]). Columns were precleaned manually with 70% EtOH before use.
Figure 4 shows a technical drawing of column A, while
Figure 5 shows column B.
3.10. Setup for Continuous Flow
Column A was filled with overgrown Diaion particles before screwing both parts of it together. Column B was filled with a 2:1 mixture of untreated Diaion particles and overgrown Diaion particles. For evaluation purposes, CV was defined as the volume inside of a packed column not occupied by the media. The column was connected using PEEK-tubing (inner diameter 0.75 mm, outer diameter 1/16″, from GE Healthcare (Uppsala, Sweden) to a LC-10ADVP-HPLC-pump from Shimadzu (Duisburg, Germany) and a F9R fraction collector from GE Healthcare.
Substrate was prepared with 50 mM dopamine, 50 mM pyruvate, 4 g L−1 ascorbic acid and 25 µg mL−1 gentamicin in 50 mM HEPES buffer (pH 7.4). A 0.9% NaCl solution and a 5 mL syringe were used to prefill columns and determine inner system volumes. Column and substrate feed were placed in an incubator at 37 °C while the further setup was stored at 25 °C.
3.11. Sample Preparation
Samples of 1 mL were taken at predetermined times and added to 200 µL of 1 M H
2SO
4. The samples were centrifuged at 10,000×
g for 5 min to remove cells and cell debris. A total of 100 µL of supernatant was evaporated and used for phenylisothiocyanate (PITC) derivatization. This was done following the established procedure of Gonzales et al. [
46]. PITC-derivatized samples were then dissolved in 80:20 HEPES buffer:MeOH before measurement. The remaining part of the sample was frozen at −20 °C for 16 h and again centrifuged after thawing. Before chiral norlaudanosoline measurements, samples were diluted 1:10 with 50 mM HEPES buffer (pH 7.4), and 20% MeOH was added before measurement.
3.12. Analytics
For all analytics, the Shimadzu Prominence HPLC 20A (Shimadzu, Duisburg, Germany) equipped with an UV detector was used. For norlaudanosoline analytics, an Astec Chirobiotic T column (25 cm) (Merck, Darmstadt, Germany) was used with 20:20:60 MeOH:ACN:25 mM phosphate buffer (pH 4.2) as mobile phase at a flow rate of 0.2 mL min
−1 at 20 °C. Retention times were 21 min for (S)-norlaudanosoline and 30 min for (R)-norlaudanosoline (
Supporting Figures S4 and S5). The detector was set to 230 nm. For pyruvate analytics, an Aminex HPX-87H column (30 cm) (Bio-Rad, Feldkirchen, Germany) was used with 5 mM sulfuric acid (pH 2.3) as mobile phase at a flow rate of 0.4 mL min
−1 at 30 °C. Retention time of pyruvate was 14.5 min. The detector was set to 210 nm. For alanine analytics, a Nucleodur C18 Gravity-SB column (15 cm) (Merck, Darmstadt, Germany) was used with 20:20:60 MeOH:ACN:25 mM phosphate buffer (pH 4.2) as mobile phase at a flow rate of 0.3 mL min
−1 at 35 °C. Retention time of alanine was 6.4 min. The detector was set to 254 nm.
3.13. Biomass Quantitation via Metabolic Activity
For the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT)-test of outflow of the rescaled packed bed reactor, 3 mL of each sample fraction was centrifuged at 11.000× g for 15 min, and the supernatant was carefully removed, before resuspending the pellet in 500 µL of 0.9% NaCl. Each sample was then split into 4 subsamples, each further diluted by half (undiluted, 1:2, 1:4, 1:8) with 0.9% NaCl. A total of 80 µL of each subsample was transferred into a microtiter plate and 20 µL MTT-solution (5 mg mL−1) was added to each well. After 1 h of incubation, the plates were centrifuged at 11.000× g for 5 min and the supernatant was removed and added to 400 µL solubilization buffer (405 µL isopropanol + 75 µL 20% w/v sodium dodecyl sulfate + 20 µL 1M HCl) and the solution was mixed thoroughly to solubilize the formazan crystals. Then, the absorbance at 575 nm and 660 nm was measured in an Infinite M Nano+ plate reader (Tecan, Männedorf, Switzerland). The difference of the absorbance at 575 nm and at 660 nm was calculated. The resulting signal was compared to the signal of catalyst suspended in 0.9% NaCl. This generated a relative value for the metabolic biomass activity. The samples up to 2.8 CV were processed immediately, while subsequent samples were processed the next day. For quantification of cell mass loss during the experiment, cells treated equally and of the same ratio as in the experiment were measured as reference during the experiment.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}