2.1. Potential Optimization of the Current-Time Measurement Method
The prepared sensor was measured by the current–time method at different potentials (the test substrate was ultrapure water). The steady-state current differences before and after the addition of 10
−5 mol/L glucose were used to measure the influence of different potentials on the electrochemical responses of the sensor. The results showed that the value of the current change was the largest under the condition of −0.38 V, as shown in
Figure 2A. Therefore, −0.38 V was chosen as the constant potential to investigate the response characteristics of the biosensor to glucose, sodium glutamate, disodium inosinate, and sodium lactate.
The redox peak potential difference is less than 80 mV, and the peak current ratio is close to 1, as shown in
Figure 2B. As shown in
Figure 2C,D, under different scanning rate conditions (25, 50, 75, 100, 125, 150, and 200 mV/s), the reduction peak and oxidation peak current of the electrode after activation have a good linear relationship with the square root of the scanning rate, indicating that the glassy carbon electrode after the treatment, the redox peak current, is controlled only by the diffusion conditions, and the pretreatment effect of the glassy carbon electrode is up to the standard, and subsequent research can be carried out.
As shown in
Figure 2E, under the conditions of a scanning rate of 50 mV/s and scanning range of −0.1~0.6 V, the peak current value after the assembly of the nuclear microporous membrane is lower than that of the bare electrode. This is because the nuclear microporous membrane impedes the transfer of electrons to the electrode surface; the peak current value is further reduced after the assembly of the colon cancer tissue or adjacent tissue, because the colon cancer tissue or adjacent tissue impedes the electron transfer. This shows that the electrode assembly is successful.
2.3. Activation Constant of the Receptor and the Ligand
The hyperbolic fit curve obtained from
Figure 4 was very similar to the Michaelis–Menten equation for enzymatic reactions. Therefore, the reciprocals of the molar concentrations of the four substances and the reciprocals of the current rate of change could be fitted to the double reciprocal linear fit according to
Figure 5 to obtain the linear regression equation, which calculated an activation constant Ka similar to the Michaelis constant (
Table 5). The essence of this constant is the ligand concentration at which the interrelated allosteric effect generated by ligand and receptor recognition triggers activation and amplification of the intracellular signal to half the maximum signal output. In other words, the smaller the activation constant, the greater the sensitivity of the ligand to the receptor activation. From this, the response concentrations of neighboring tissues to different types of ligands could be deduced.
The results in
Table 5 show that the activation constants differed between the adjacent tissues and the cancerous tissues. For three substances, including glucose, sodium glutamate, and disodium inosinate, the differences in Ka values between the cancer tissues and adjacent tissues were 14 times, 115 times, and 3 times, respectively. Sodium lactate, however, showed a different pattern; the cancer tissues had almost no response, whereas the adjacent tissues showed a strong sensing ability, with a sensitivity only lower than that of glucose. We were particularly interested in the way in which these dramatic differences in the sensing capabilities of the cancer and adjacent tissues were expressed. We proposed two possible explanations for this: differences in the number of receptors on the cell surface and changes in the intracellular cascade amplification system. We estimated the intracellular amplification of the signaling cascade and the minimum number of receptors required to activate the maximum output signal on the cell surface to identify the causes of these differences.
2.4. Estimation of Cell Cascade Magnification
The common feature of GPCRs is the seven-transmembrane structure. If the process of recognition between the extracellular domain and the ligand occurs through a “signature” method—namely, through the interconnected allosteric interactions between the seven transmembrane domains—the signature is in the G protein activation mode [
23] and the simultaneous activation of genomic and non-genomic pathways. The kinetic laws and parameters determined in this study included the control of plasma membrane ion channels by non-genomic signaling pathways by G proteins. Studies have shown that the ligands used in this study mainly sense and amplify signals through T1R1 and T1R2. The process of receptors and ligand recognition activated the heterotrimeric G proteins through interrelated allosteric effects, and activation of the βγ subunit additionally activated PLCβ2, causing Ca
2+ to flow in from the transient receptor potential ion channel protein 5 (TRPM5), resulting in the depolarization of the cell. These changes in electrochemical signals can be detected and collected by the glassy carbon electrode. We were also able to quantitatively measure and estimate these electrical signal changes using bare electrodes as a control. Similarly, the logarithms of the concentrations of the four substance solutions were used as the abscissa, and the current change rates were plotted on the ordinate to perform a linear fit. The interaction equations of the four ligands of glucose, sodium glutamate, disodium inosinate, and sodium lactate with the bare electrode were as described below.
The bare electrode concentrations could be calculated by using the current change rates corresponding to the activation constant Ka in
Table 5 in the bare electrode equation in
Table 6. The following equations could then be used to calculate the cascade amplification folds [
23]:
It is obvious that there was a significant difference in the amplification factor of the intracellular signals between the cancerous tissues and the peripheral cancerous tissues. The amplification factor of the intracellular signals in the cancer tissues was much higher than that of the peripheral cancer tissues. The differences in the signal amplification of glucose, sodium glutamate, and disodium inosinate were about 249 times, 270 times, and 16 times, respectively. The experimental results confirmed that, in comparison with the cancer-adjacent tissues, the colon cancer cells required nutrients for their reproduction, including glucose, sodium glutamate, and disodium inosinate. The increased sensing ability was primarily manifested by increased activation of the intracellular signal amplification system.
2.5. Estimation of the Minimum Number of Receptors Required to Activate Cancer-Adjacent Tissues or Cancerous Tissues to Achieve the Maximum Signal Output
Through the kinetic equation of ligand–receptor interactions, the minimum number of receptors (hereafter referred to as the minimum number of receptors) required to activate the adjacent or cancerous tissue to obtain the maximum signal output could be calculated. A reported method [
24] defined the minimum number of receptors required to activate the adjacent tissue or cancerous tissue to obtain the maximum signal output as follows:
In the equation, C
M is the concentration that gave the maximum response signal when the number of receptors on the cell was not one, and C
L is the concentration that gave the maximum response signal when there was only one receptor on the cell surface. After careful calculation, it was concluded that the minimum number of receptors for glucose, sodium glutamate, disodium inosinate, and sodium lactate on the colon cancer tissue cells were 1.49, 2.20, 9.21, and 0, respectively. The minimum number of receptors for glucose, sodium glutamate, disodium inosinate, and sodium lactate on the healthy cells adjacent to the cancer tissues were 2.02, 2.55, 2.85, and 1.83, respectively. The results showed that the number of receptors for the two substances of glucose and sodium glutamate did not differ much between the cancer tissues and the peripheral cancer tissues, whereas disodium inosinate was significantly different. Studies have reported that T1R1 can sense sodium glutamate and disodium inosinate, and another receptor, the metabotropic glutamate receptors (mGluR) [
25], can only sense glutamate sodium but not inosine. Therefore, based on the above information, we speculated that the increased sensitivity of colon cancer tissue to disodium inosinate would mainly depend on the increased intracellular signal amplification of disodium inosinate through T1R1 activation rather than the increased signal amplification system of inosine. In other words, the signal amplification systems for glucose and sodium glutamate showed a higher degree of improvement than disodium inosinate. By estimating the amplification of the cell signal cascade, the minimum number of receptors on the cell surface, it is clear that the sensing of glucose and sodium glutamate by the cancer tissues was primarily achieved by altering the intracellular signal amplification.
2.7. Discussion
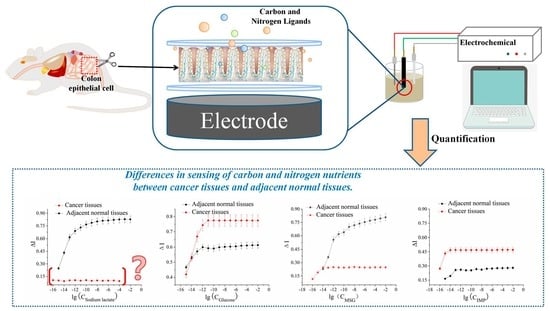
Indeed, the cancerous and peripheral cancerous tissues differed in their ability to sense carbon and nitrogen nutrients, whereas the cell surface receptors of the healthy and cancerous cells remained unchanged, eliminating the differences in ligand–receptor interactions. Therefore, our study focused only on intracellular signaling. Both sweet taste sensor receptors T1R2/T1R3 and umami taste sensor receptors T1R1/T1R3 are G protein-coupled receptors. Carbohydrates bind to T1R2/T1R3 to activate α-tastein, thereby activating adenosine acid cyclase (adenylyl cyclase, AC) to produce 3′, 5′-cyclic adenylate (cAMP), leading to an increase in intracellular cAMP concentration. This led to the activation of protein kinase A (PKA) and K+ phosphorylation of the channel to close the ion channel, inhibiting K+ outflow, membrane depolarization, and neurotransmitter release. The umami taste stimulant bound to T1R1/T1R3 to activate α-tastein, resulting in the separation of the β and γ subunits of the G protein. The released Gβ3 and Gγ13 subunits activated phospholipase-β2 (phospholipase-β2, PLC-β2), and PLC-β2 hydrolyzed phosphatidylinositol-4,5-bisphosphate [PI(4,5) P2] into diacylglycerol (DAG) and inositol triphosphate (IP3), which binds to the third type of inositol triphosphate receptor 3 (IP3R3), resulting in the opening of the IP3-gated Ca2+ channel on the intracellular organelle membrane and the intracellular Ca2+ storage. In turn, the increase in the concentration of Ca2+ in the cytoplasm led to the transient receptor potential melastatin 5 (TRPM5) channel to open, resulting in the inflow of Na+ to enter the cell. This will ultimately lead to membrane depolarization and neurotransmitter release, thereby generating electrical signals.
G proteins use the energy released by the hydrolysis of GTP to GDP to control the switching of ion channels. While controlling the rapid transmission of electrochemical signals, they also control the replication, transcription, internal and external materials, and energy of the genome through complex signaling pathways, nuclear factor and transcription factor networks, and energy and signal exchange [
26]. They also dominate the cell cycle, proliferation, and autophagy by controlling write; read; and erase (cell energy charge, reduction power, and modification of metabolic intermediates) on nucleosomes and apoptosis [
27].
The main difference between the cancerous and cancer-adjacent tissues in this study was the intracellular signal amplification. Although a rapid non-genomic approach was used for measurements, the difference in the signal amplification system was determined by the complex epigenetic modification of the colorectal cancer cells and the peripheral cancer tissue cells, as the fast pathway of colorectal cancer cell progression remained the same. Compared to healthy cells, the biggest difference in cancer cells is that they proliferate indefinitely [
28]. The premise is that they must continuously sense and absorb glucose to provide themselves with carbon skeletons and raw materials and thus continuously sense amino acids and nuclear glycolic acid to provide nitrogen nutrition for DNA replication, transcription, and protein synthesis. However, nucleotide, amino acid, and fatty acid synthesis should be involved in the TCA cycle, thereby interrupting the transfer of hydrogen protons to oxygen through the oxidized respiratory chain, interrupting oxidized phosphoric acid. The hydrogen protons can then only be transferred to pyruvate to produce lactate or removed from cells or tissues by the Warburg effect. Healthy cells undergo oxidative phosphorylation to obtain energy from the bloodstream [
29]. Recent studies have consistently shown that all cells must undergo these regulatory transformations in the process of proliferation. In particular, the proliferation of immune cells also exhibits a Warburg effect [
30], whereas the proliferation of healthy cells can be controlled. Specifically, only when cells need to proliferate do they continuously sense and absorb the nutrients necessary for their proliferation, and they will not continue to do so when they do not need to proliferate. Therefore, the manner by which healthy cells stop sensing and absorbing the nutrients necessary for their proliferation is an issue that must be clarified. Based on the results of a large number of recent studies, it is clear that the sensory control of essential nutrients is the key to this problem. The sensory control of cells is similar to the control of animal nutrition. Under starvation conditions, carbohydrates are rewarded with sweetness and amino acids, or nucleotides are rewarded with umami. In contrast, in satiated conditions, even if these nutrients and receptors are still present, they are no longer rewarded in the same way as when we are hungry. It is clear that the key to its control (especially negative inhibition) is not the receptor or the ligand but the control of the intracellular signal amplification process. The results of this study showed that the major differences between the colon cancer tissues and adjacent tissues were in the constitutive and heritable differences in signal transmission and amplification. In this regard, it could be inferred that continuous proliferation depends on continuous nutrient sensing and absorption, and continuous nutrient sensing and absorption depend on the continuous transcription and expression of the corresponding genes. Simply put, epigenetic modifications to the lysine side of nucleosome caused by repeated writing, reading, and erasing inhibit the energy charge (which is ATP) required to support cell proliferation; key metabolic intermediates (including lactate); coenzymes (including coenzymes I, II, and acetyl-CoA); heterotrimeric G protein; and the GTP substrate-level phosphorylation substrates (succinyl-CoA) required by the small G protein [
27] are the fundamental cause of cancer initiation and development. In general, the difference in carbon and nitrogen nutrition between cancer cells and para-cancerous cells is an illusion, and the reason lies in the changes in intracellular metabolism and the result of metabolic reprogramming. GPCRs play a key role in this process, accompanied by the exchange of GDP-GTP and ATP-cAMP, the phosphorylation of various kinases, and the activation of multiple downstream pathways by small G proteins. At the same time, the metabolism also changes. A large number of intermediate metabolites in the TCA cycle accumulate, are excreted from the mitochondria, and participate in anabolic metabolism, providing the proteins, lipids, and ribose required for the proliferation of cancer cell proliferation. Finally, the TCA cycle is disrupted, and glutamine is produced. A variety of coenzymes such as acetyl, succinyl, and malonyl also accumulate during this process to modify histones and feed back into the G protein signal amplification system. The intracellular signal amplification coefficient is increased, allowing lower concentrations of carbon and nitrogen nutrients to be taken up (
Figure 7).
Among the four carbon and nitrogen nutrients, sodium lactate is unique. Lactate can be transported by the monocarboxylate transporter (MCT) in tissues, cells, and organs and regulates body synthesis and catabolism in the body, where it plays a key role in the process. Lactate can also be used as a carbon source by the human body [
31,
32]. In addition, lactic acid has a specific sensor receptor, the HCAR1 receptor, through which lactic acid can directly inhibit the action of the NLRP3 inflammasome, which is activated by the action of TLR-4 and caspase1. As a result, the effect of NF-κB is reduced, and the conversion of PRO-IL-1b to active IL-1b is reduced [
33,
34]. This has a significant beneficial effect on disease severity in these two models. Lactic acid at physiological concentrations is a good anti-inflammatory agent, and this anti-inflammatory effect can effectively inhibit cell reproduction. However, the cancer tissues rejected the lactate. One of the most striking features of cancer cells is the Warburg effect, or aerobic glycolysis. This process produces a large amount of lactic acid, which is released into the circulation in the form of salt and is sensed, absorbed and utilized by healthy cells. For example, the TCA cycle is completely oxidized. For healthy cells, lactate is both a carbon nutrient and an energy substance, whereas, for cancerous tissues, which produce large amounts of lactic acid, lactate is no longer a nutrient. The loss of lactate-sensing ability in cancer cells is clearly an inevitable result, which was confirmed in this study.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}