A PBMC-Based System to Assess Human T Cell Responses to Influenza Vaccine Candidates In Vitro

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Vaccines
2.2. Peptides
2.3. Human Primary Cells
2.4. In Vitro Stimulation Using Influenza Vaccines and Peptides
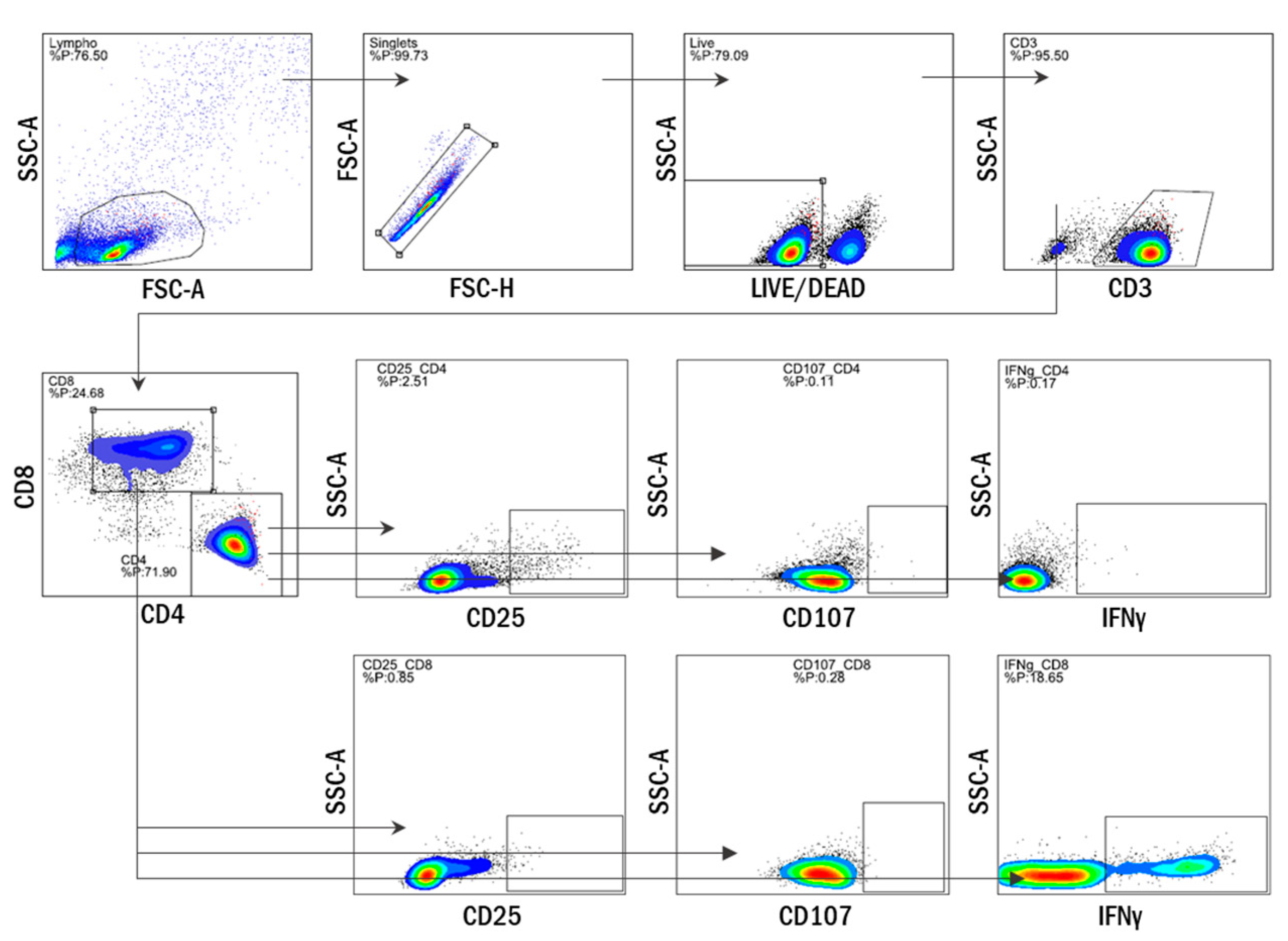
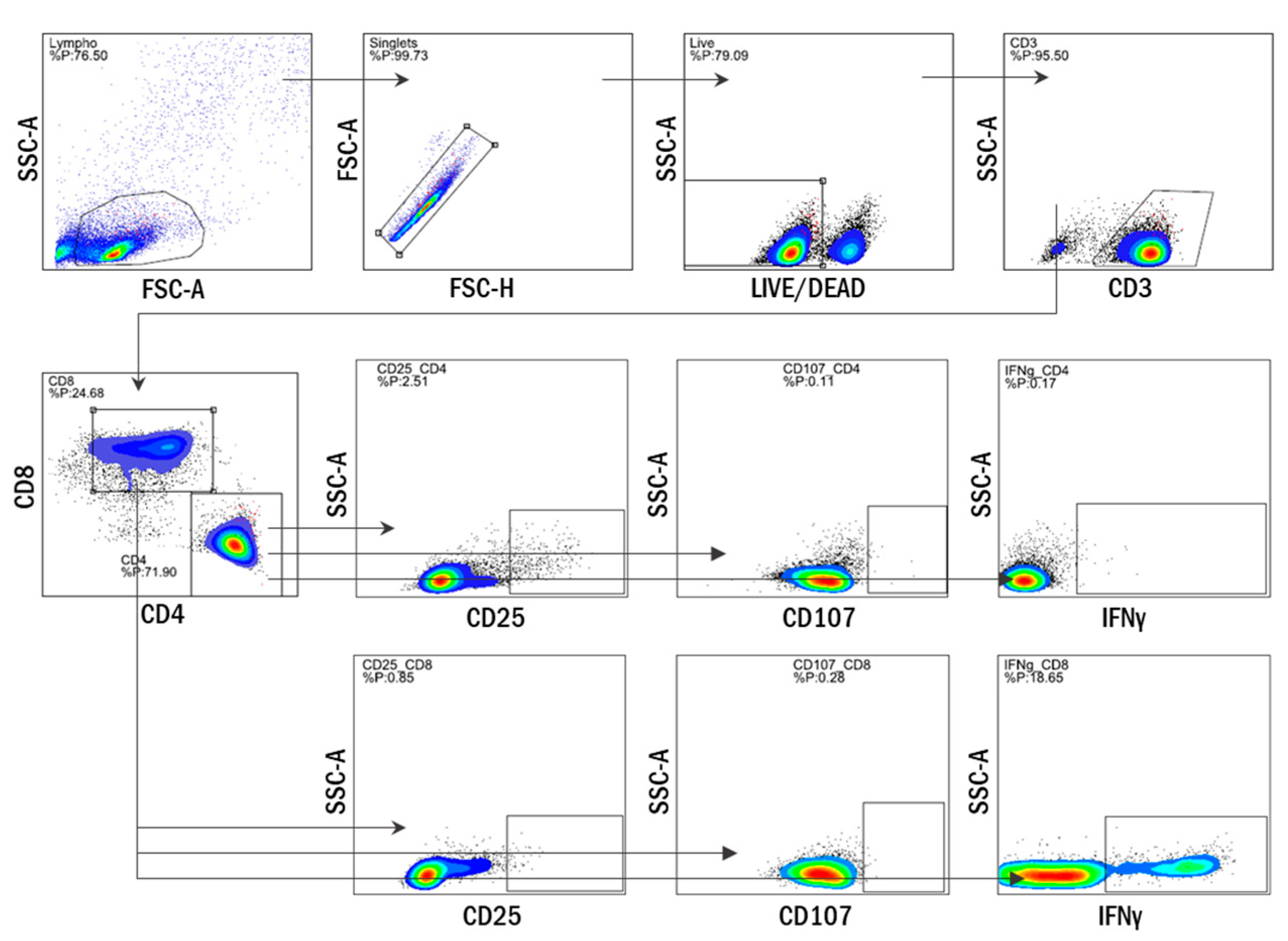
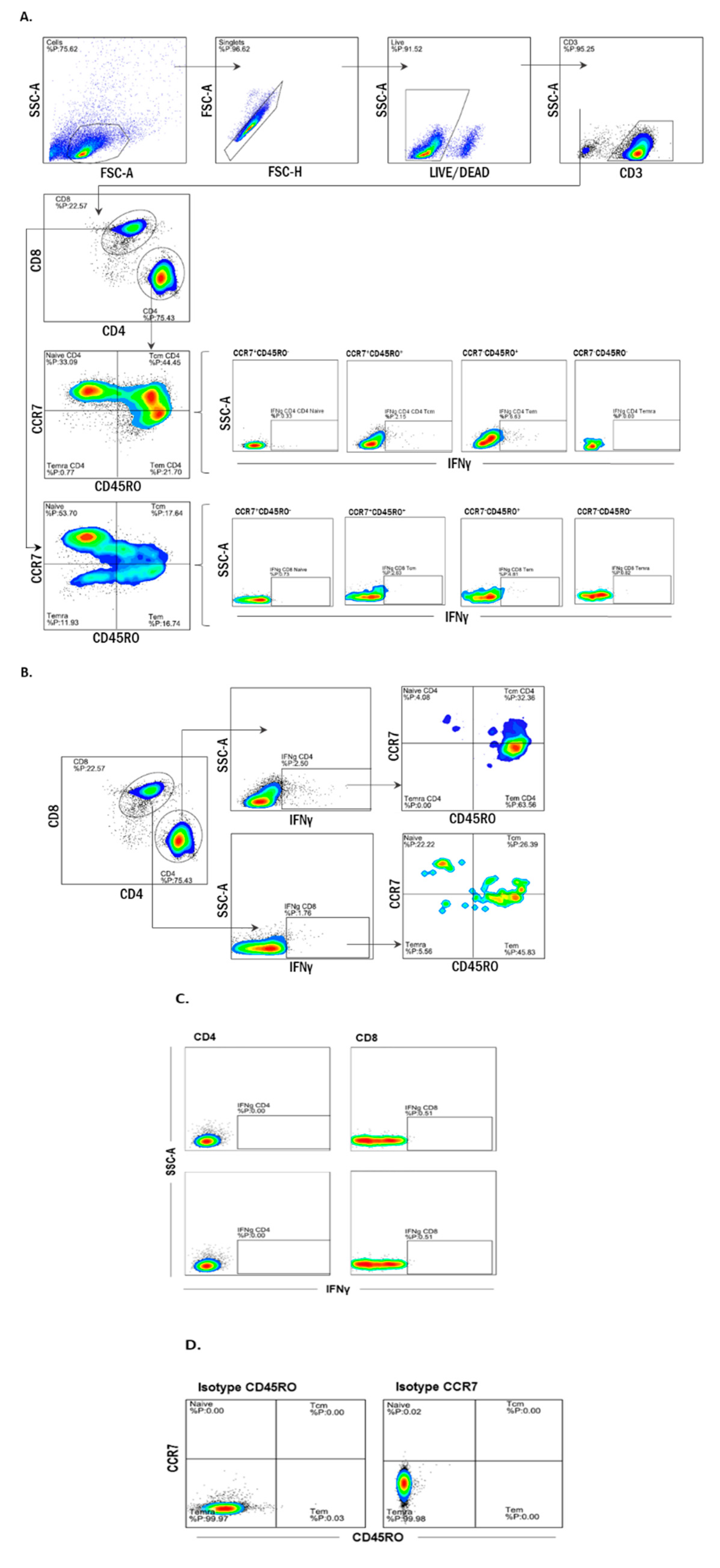
2.5. Multiparameter Flow Cytometry
2.6. ELISpot
2.7. Statistical Analysis
3. Results
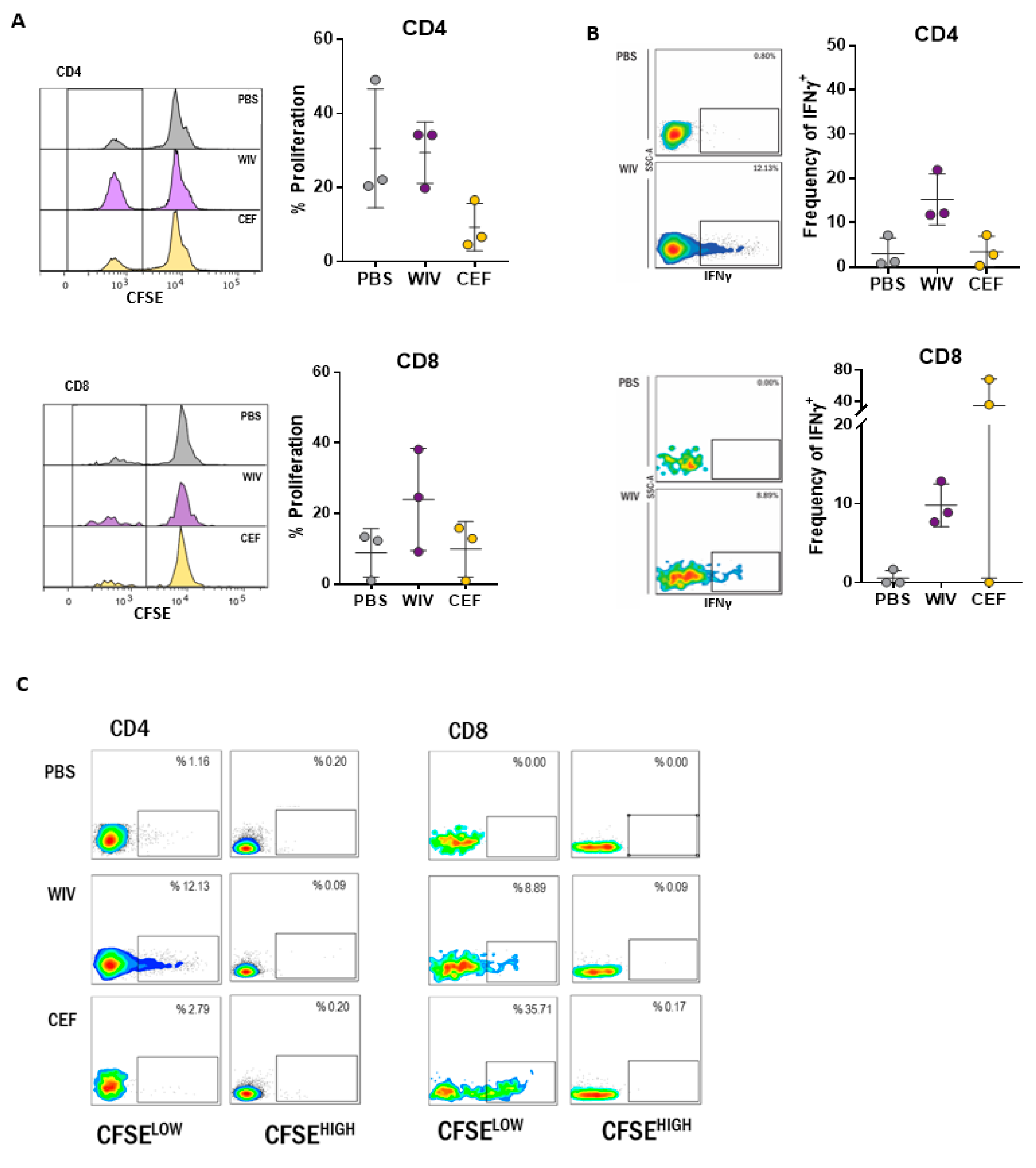
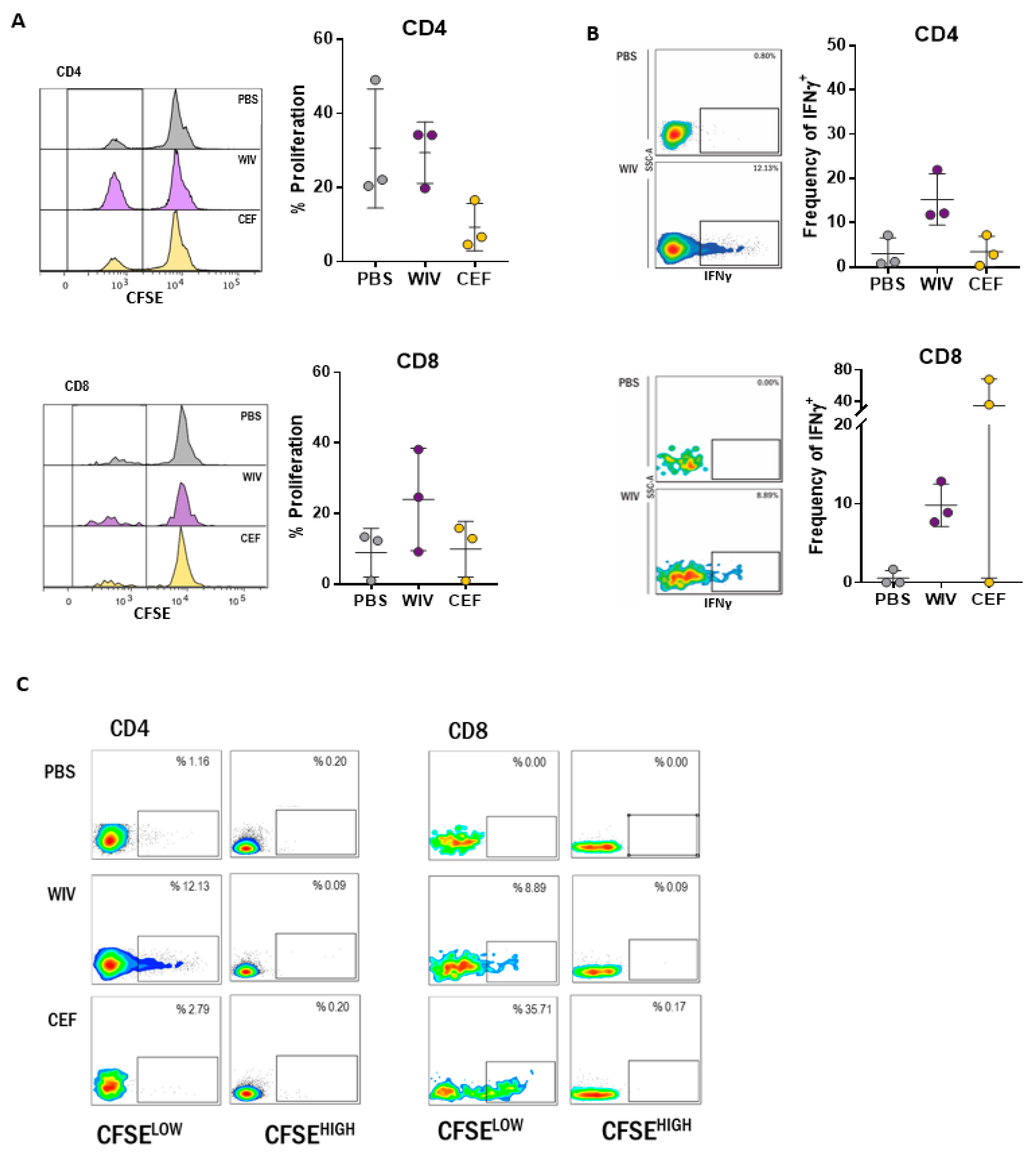
3.1. Antigen-Specific T Cells Expand Upon In Vitro Stimulation with WIV Vaccines
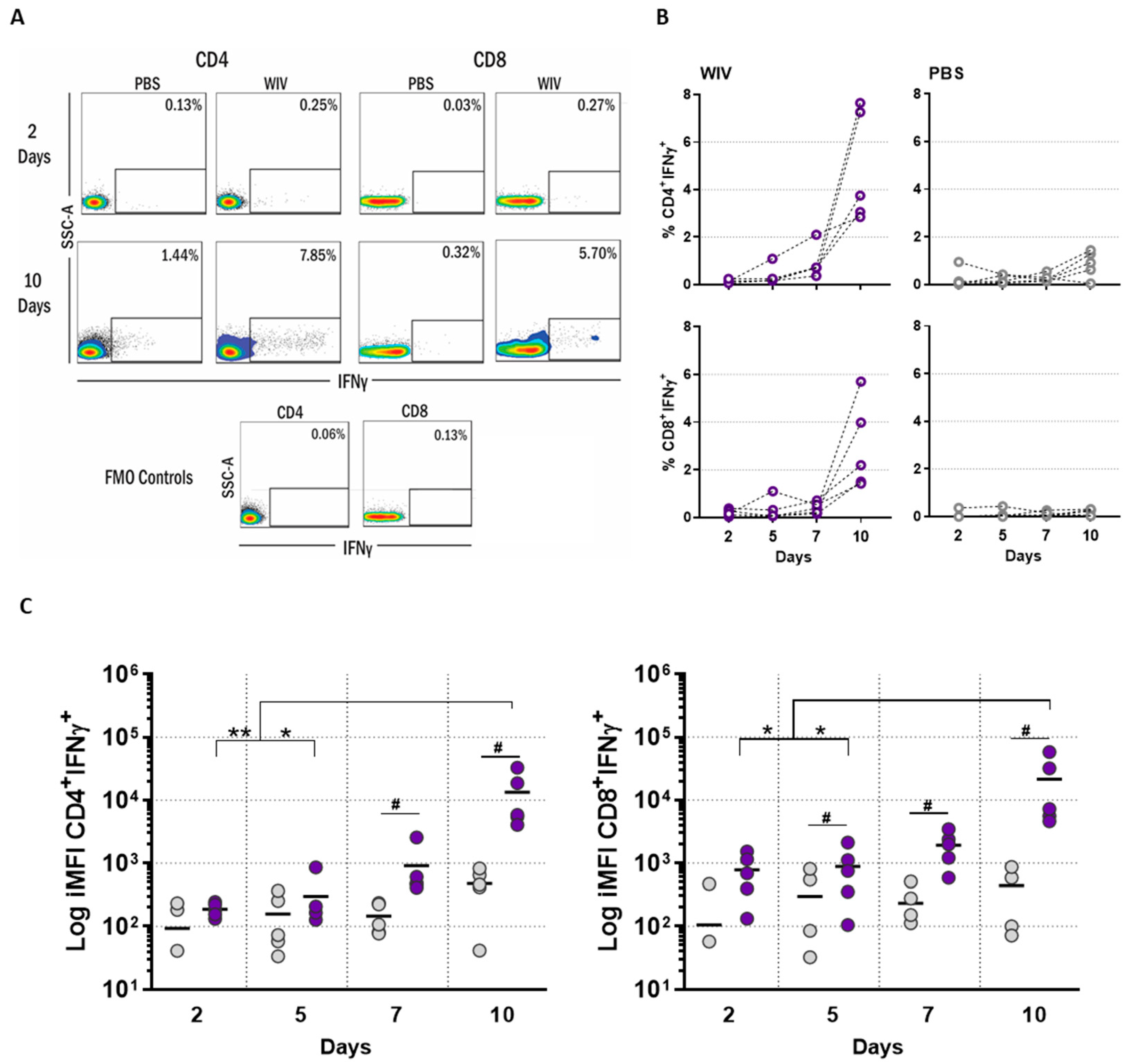
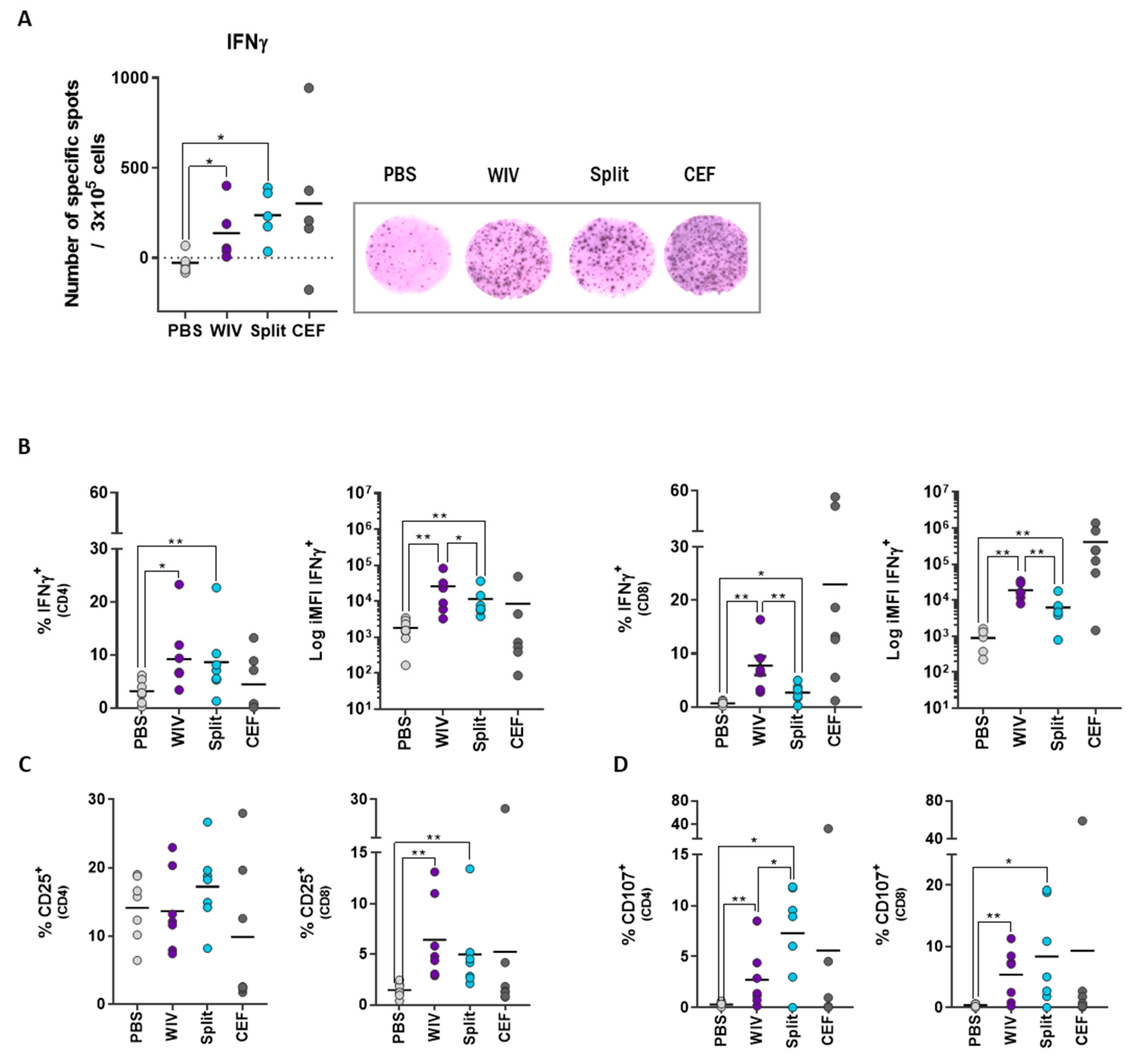
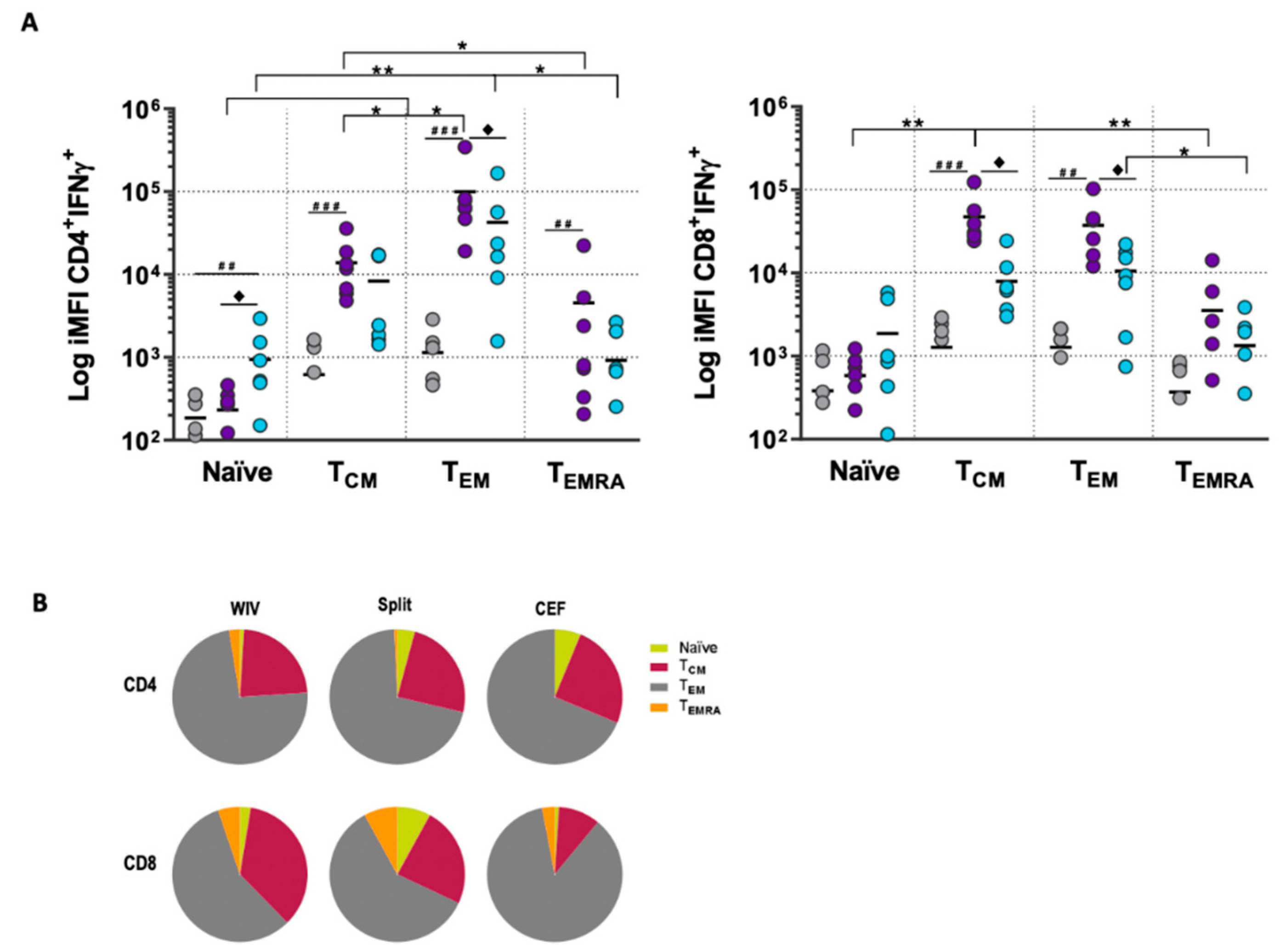
3.2. T Cell Responses in Long-Term PBMC Cultures Are Vaccine Formulation-Specific
3.3. Memory T Cell Subsets Are the Main Source of Antigen-Specific Responses In Vitro
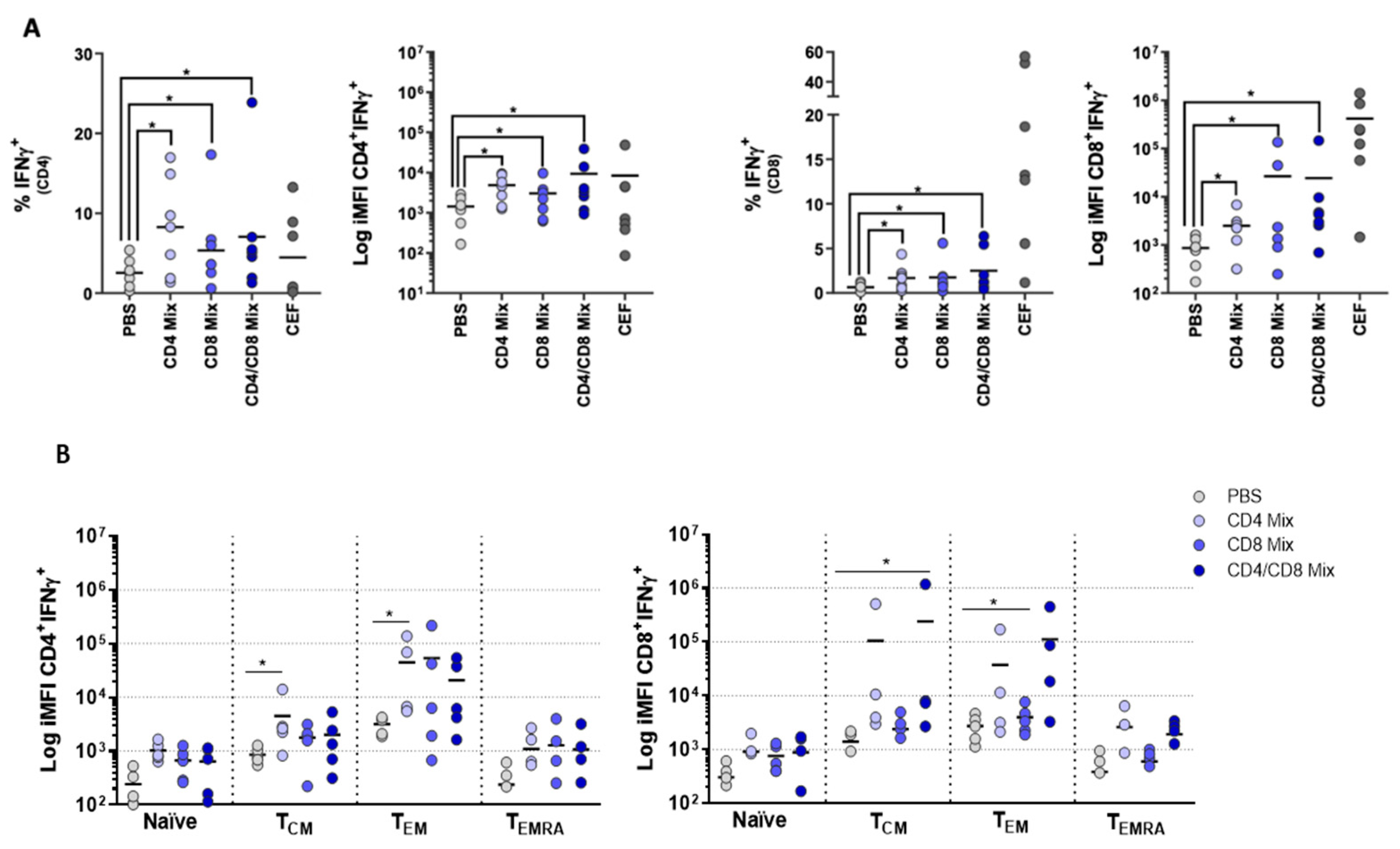
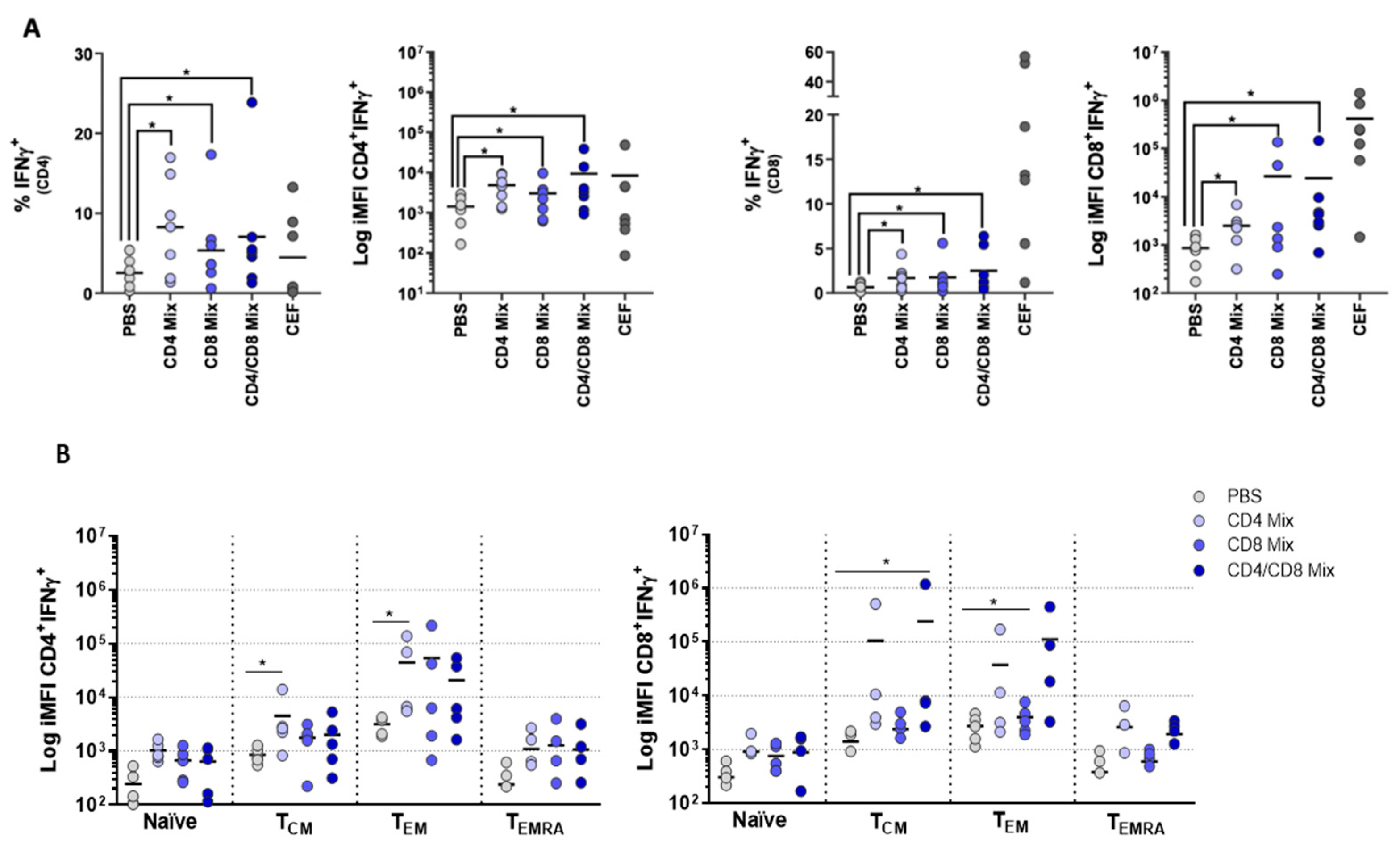
3.4. PBMCs Display Antigen-Specific Responses upon Stimulation with Influenza Peptides
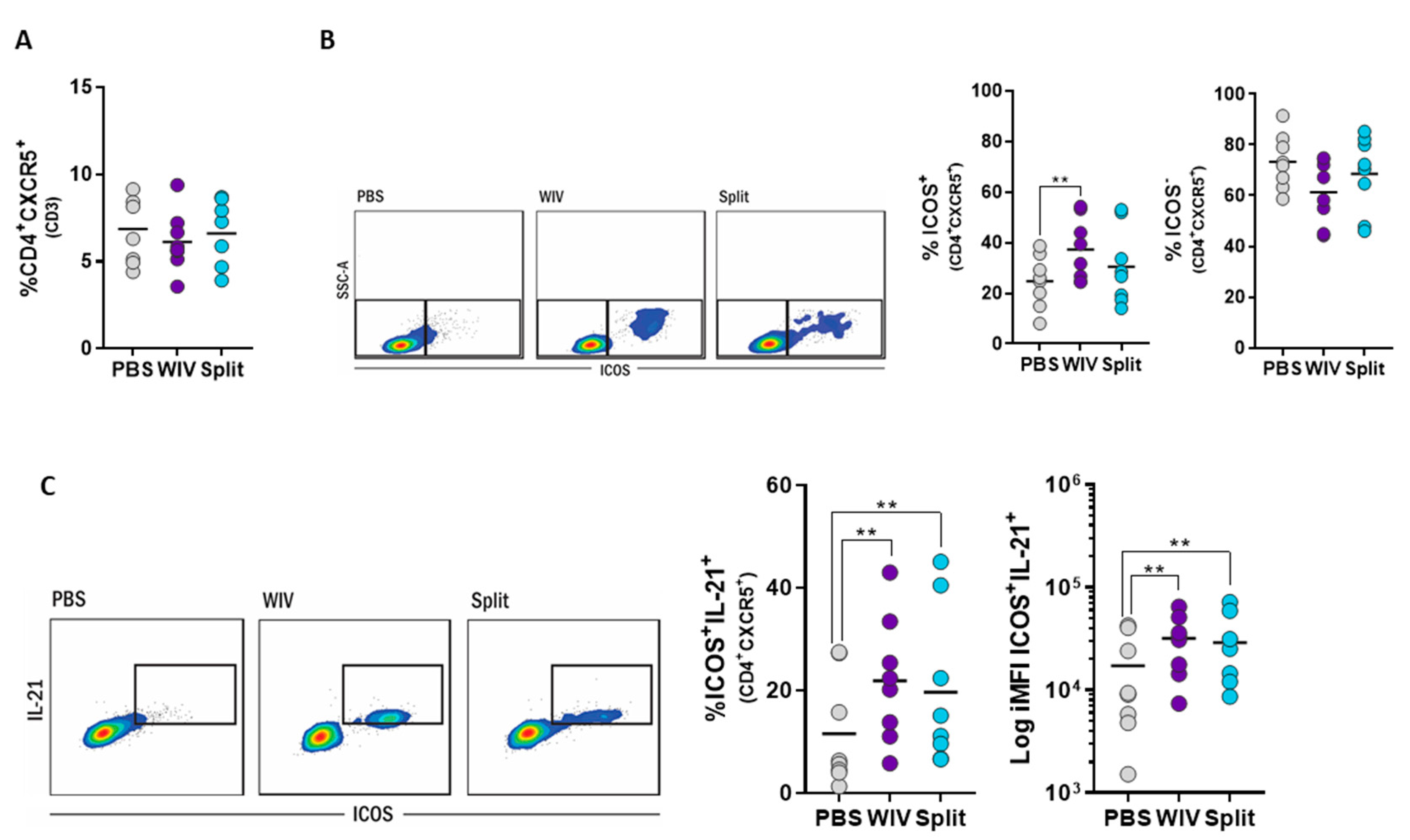
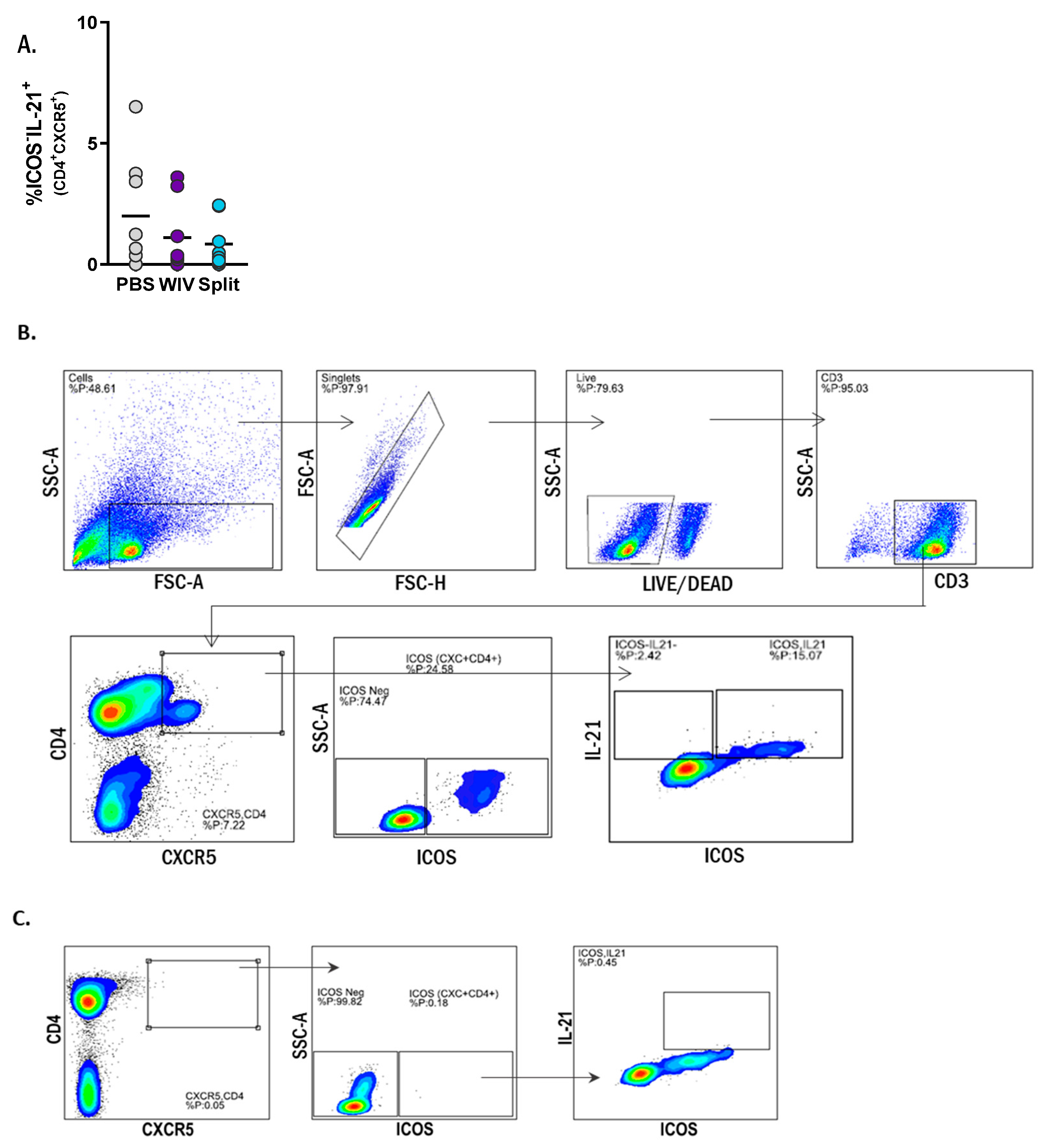
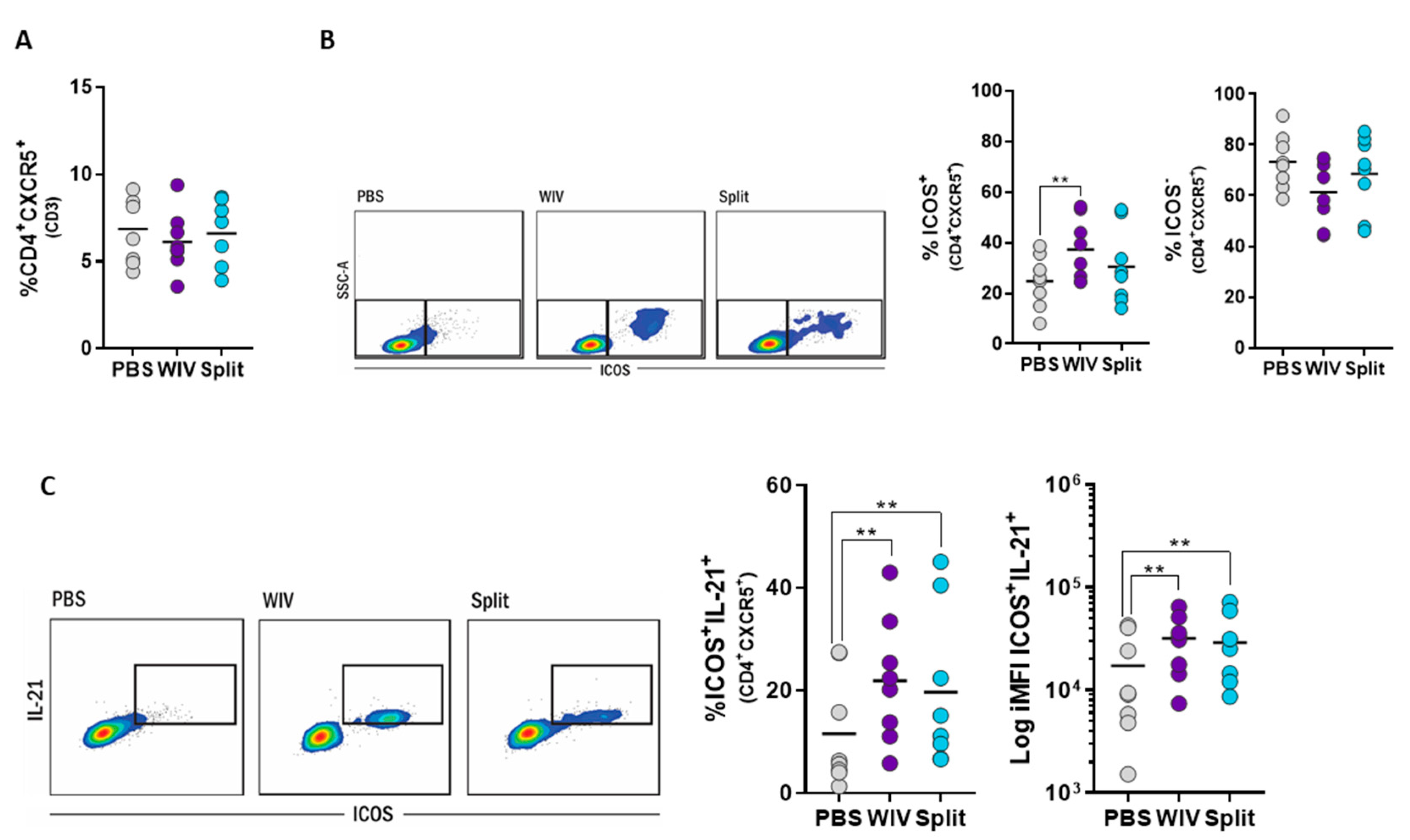
3.5. Induction of Follicular T Helper Cells by Influenza Vaccines In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epitope | Protein | Amino Acid Position | CD4/CD8 | Length |
|---|---|---|---|---|
| LKREITFHGAKEIALSY | M | 103–119 | CD4 | 16 |
| TYQRTRALVRTGMDPRM | NP | 147–163 | CD8 | 16 |
| EALMEWLKTRPILSPLTK | M | 40–57 | CD8 | 17 |
| RMCNILKGKFQTAAQRAM | NP | 221–238 | CD4 | 17 |
| Panel 1. Activation, Cytotoxicity and IFNγ of T Cells T Cells | Panel 2. T Cell Subsets and IFNγ | Panel 3. T Follicular Helper Cells |
|---|---|---|
| Live/Dead-Pacific Orange | Live/Dead - Pacific Orange | Live/Dead - Pacific Orange |
| CD3-Pacific Blue | CD3-Pacific Blue | CD3-Pacific Blue |
| CD4-APCCy7 | CD4-APCCy7 | CD4-APCCy7 |
| CD8-PerCPCy5.5 | CD8-PerCPCy5.5 | ICOS-PerCPCy5.5 |
| IFNγ-PeCy7 | IFNγ-PeCy7 | CXCR5-PeCy7 |
| CD25-PE | CCR7-PE | IL21-PE |
| CD107-APC | CD45RO-APC | - |
References
- Smith, S.G.; Kleinnijenhuis, J.; Netea, M.G.; Dockrell, H.M. Whole Blood Profiling of Bacillus Calmette–Guérin-Induced Trained Innate Immunity in Infants Identifies Epidermal Growth Factor, IL-6, Platelet-Derived Growth Factor-AB/BB, and Natural Killer Cell Activation. Front. Immunol. 2017, 8, 644. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.F.; Coviello, S.; Monsalvo, A.C.; Melendi, G.A.; Hernandez, J.Z.; Batalle, J.P.; Diaz, L.; Trento, A.; Chang, H.-Y.; Mitzner, W.; et al. Lack of antibody affinity maturation due to poor Toll-like receptor stimulation leads to enhanced respiratory syncytial virus disease. Nat. Med. 2009, 15, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Watkins, D.I.; Burton, D.R.; Kallas, E.G.; Moore, J.P.; Koff, W.C. Nonhuman primate models and the failure of the Merck HIV-1 vaccine in humans. Nat. Med. 2008, 14, 617–621. [Google Scholar] [CrossRef]
- Tapia-Calle, G.; Stoel, M.; de Vries-Idema, J.; Huckriede, A. Distinctive Responses in an In Vitro Human Dendritic Cell-Based System upon Stimulation with Different Influenza Vaccine Formulations. Vaccines 2017, 5, 21. [Google Scholar] [CrossRef]
- Hoonakker, M.E.; Verhagen, L.M.; Hendriksen, C.F.M.; van Els, C.A.C.M.; Vandebriel, R.J.; Sloots, A.; Han, W.G.H. In Vitro Innate Immune Cell Based Models To Assess Whole Cell Bordetella Pertussis Vaccine Quality: A Proof Of Principle. Biologicals 2015, 43, 100–109. [Google Scholar] [CrossRef]
- Kon, T.C.; Onu, A.; Berbecila, L.; Lupulescu, E.; Ghiorgisor, A.; Kersten, G.F.; Cui, Y.-Q.; Amorij, J.-P.; Van der Pol, L. Influenza Vaccine Manufacturing: Effect of Inactivation, Splitting and Site of Manufacturing. Comparison of Influenza Vaccine Production Processes. PLoS ONE 2016, 11, e0150700. [Google Scholar] [CrossRef]
- Wilkinson, T.M.; Li, C.K.F.; Chui, C.S.C.; Huang, A.K.Y.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B.; et al. Preexisting Influenza-Specific CD4+ T Cells Correlate With Disease Protection Against Influenza Challenge In Humans. Nat. Med. 2012, 18, 274–280. [Google Scholar] [CrossRef]
- Darrah, P.A.; Patel, D.T.; De Luca, P.M.; Lindsay, R.W.B.; Davey, D.F.; Flynn, B.J.; Hoff, S.T.; Andersen, P.; Reed, S.G.; Morris, S.L.; et al. Multifunctional TH1 Cells Define A Correlate Of Vaccine-Mediated Protection Against Leishmania Major. Nat. Med. 2007, 13, 843–850. [Google Scholar] [CrossRef]
- Budimir, N.; de Haan, A.; Meijerhof, T.; Waijer, S.; Boon, L.; Gostick, E.; Price, D.A.; Wilschut, J.; Huckriede, A. Critical Role of TLR7 Signaling in the Priming of Cross-Protective Cytotoxic T Lymphocyte Responses by a Whole Inactivated Influenza Virus Vaccine. PLoS ONE 2013, 8, e63163. [Google Scholar] [CrossRef]
- Huckriede, A.; Bungener, L.; ter Veer, W.; Holtrop, M.; Daemen, T.; Palache, A.M.; Wilschut, J. Influenza Virosomes: Combining Optimal Presentation Of Hemagglutinin With Immunopotentiating Activity. Vaccine 2003, 21, 925–931. [Google Scholar] [CrossRef]
- Budimir, N.; Huckriede, A.; Meijerhof, T.; Boon, L.; Gostick, E.; Price, D.A.; Wilschut, J.; de Haan, A. Induction of Heterosubtypic Cross-Protection against Influenza by a Whole Inactivated Virus Vaccine: The Role of Viral Membrane Fusion Activity. PLoS ONE 2012, 7, e30898. [Google Scholar] [CrossRef]
- Karlsson, A.C.; Martin, J.N.; Younger, S.R.; Bredt, B.M.; Epling, L.; Ronquillo, R.; Varma, A.; Deeks, S.G.; McCune, J.M.; Nixon, D.F.; et al. Comparison Of The ELISPOT And Cytokine Flow Cytometry Assays For The Enumeration Of Antigen-Specific T Cells. J. Immunol. Methods 2003, 283, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.; Eirikis, E.; Davis, C.; Davis, H.M.; Prabhakar, U. Comparative analysis of lymphocyte activation marker expression and cytokine secretion profile in stimulated human peripheral blood mononuclear cell cultures: An in vitro model to monitor cellular immune function. J. Immunol. Methods 2004, 293, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Betts, M.R.; Koup, R.A. Detection of T-Cell Degranulation: CD107a and b. Methods Cell Biol. 2004, 75, 497–512. [Google Scholar] [PubMed]
- Sridhar, S.; Begom, S.; Bermingham, A.; Hoschler, K.; Adamson, W.; Carman, W.; Bean, T.; Barclay, W.; Deeks, J.J.; Lalvani, A. Cellular Immune Correlates Of Protection Against Symptomatic Pandemic Influenza. Nat. Med. 2013, 19, 1305–1312. [Google Scholar] [CrossRef]
- Wagar, L.E.; Rosella, L.; Crowcroft, N.; Lowcock, B.; Drohomyrecky, P.C.; Foisy, J.; Gubbay, J.; Rebbapragada, A.; Winter, A.-L.; Achonu, C.; et al. Humoral and Cell-Mediated Immunity to Pandemic H1N1 Influenza in a Canadian Cohort One Year Post-Pandemic: Implications for Vaccination. PLoS ONE 2011, 6, e28063. [Google Scholar] [CrossRef]
- van de Sandt, C.E.; Hillaire, M.L.B.; Geelhoed-Mieras, M.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M.; Rimmelzwaan, G.F. Human Influenza A Virus–Specific CD8+ T-Cell Response Is Long-lived. J. Infect. Dis. 2015, 212, 81–85. [Google Scholar] [CrossRef]
- Boots, A.M.H.; Maier, A.B.; Stinissen, P.; Masson, P.; Lories, R.J.; De Keyser, F. The Influence Of Ageing On The Development And Management Of Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2013, 9, 604–613. [Google Scholar] [CrossRef]
- Schmitt, N.; Bentebibel, S.-E.; Ueno, H. Phenotype And Functions Of Memory Tfh Cells In Human Blood. Trends Immunol. 2014, 35, 436–442. [Google Scholar] [CrossRef]
- Chevalier, N.; Jarrossay, D.; Ho, E.; Avery, D.T.; Ma, C.S.; Yu, D.; Sallusto, F.; Tangye, S.G.; Mackay, C.R. CXCR5 Expressing Human Central Memory CD4 T Cells and Their Relevance for Humoral Immune Responses. J. Immunol. 2011, 186, 5556–5568. [Google Scholar] [CrossRef]
- Morita, R.; Schmitt, N.; Bentebibel, S.-E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human Blood CXCR5+CD4+ T Cells Are Counterparts of T Follicular Cells and Contain Specific Subsets that Differentially Support Antibody Secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef]
- Rasheed, A.-U.; Rahn, H.-P.; Sallusto, F.; Lipp, M.; Müller, G. Follicular B Helper T Cell Activity Is Confined To CXCR5hi ICOShi CD4 T Cells And Is Independent Of CD57 Expression. Eur. J. Immunol. 2006, 36, 1892–1903. [Google Scholar] [CrossRef]
- Bentebibel, S.-E.; Lopez, S.; Obermoser, G.; Schmitt, N.; Mueller, C.; Harrod, C.; Flano, E.; Mejias, A.; Albrecht, R.A.; Blankenship, D.; et al. Induction Of ICOS+CXCR3+CXCR5+ TH Cells Correlates With Antibody Responses To Influenza Vaccination. Sci. Transl. Med. 2013, 5, 176ra32. [Google Scholar] [CrossRef] [PubMed]
- Bentebibel, S.-E.; Khurana, S.; Schmitt, N.; Kurup, P.; Mueller, C.; Obermoser, G.; Palucka, A.K.; Albrecht, R.A.; Garcia-Sastre, A.; Golding, H.; et al. ICOS+PD-1+CXCR3+ T Follicular Helper Cells Contribute to the Generation of High-Avidity Antibodies Following Influenza Vaccination. Sci. Rep. 2016, 6, 26494. [Google Scholar] [CrossRef]
- Spensieri, F.; Borgogni, E.; Zedda, L.; Bardelli, M.; Buricchi, F.; Volpini, G.; Fragapane, E.; Tavarini, S.; Finco, O.; Rappuoli, R.; et al. Human Circulating Influenza-CD4+ ICOS1+IL-21+ T Cells Expand After Vaccination, Exert Helper Function, And Predict Antibody Responses. Proc. Natl. Acad. Sci. USA 2013, 110, 14330–14335. [Google Scholar] [CrossRef]
- Spensieri, F.; Siena, E.; Borgogni, E.; Zedda, L.; Cantisani, R.; Chiappini, N.; Schiavetti, F.; Rosa, D.; Castellino, F.; Montomoli, E.; et al. Early Rise of Blood T Follicular Helper Cell Subsets and Baseline Immunity as Predictors of Persisting Late Functional Antibody Responses to Vaccination in Humans. PLoS ONE 2016, 11, e0157066. [Google Scholar] [CrossRef]
- Moser, J.M.; Sassano, E.R.; Leistritz, D.C.; Eatrides, J.M.; Phogat, S.; Koff, W.; Drake, D.R., III. Optimization Of A Dendritic Cell-Based Assay For The In Vitro Priming Of Naïve Human CD4+ T Cells. J. Immunol. Methods 2010, 353, 8–19. [Google Scholar] [CrossRef]
- Komanduri, K.V.; Thomas, M.; St. John, L.S.; Xing, D.; Decker, W.; Robinson, S.; Yang, H.; Molldrem, J.J.; Champlin, R.E.; McMannis, J.D.; et al. Characterization of optimal T Cell/Dendritic Cell (DC) Co-Culture Conditions for Ex Vivo Expansion of Antigen-Specific Human T Cells. Blood 2006, 108, 3654. [Google Scholar] [CrossRef]
- Höpken, U.E.; Lehmann, I.; Droese, J.; Lipp, M.; Schüler, T.; Rehm, A. The Ratio Between Dendritic Cells And T Cells Determines The Outcome Of Their Encounter: Proliferation Versus Deletion. Eur. J. Immunol. 2005, 35, 2851–2863. [Google Scholar] [CrossRef]
- Halbroth, B.R.; Heil, A.; Distler, E.; Dass, M.; Wagner, E.M.; Plachter, B.; Probst, H.C.; Strand, D.; Hartwig, U.F.; Karner, A.; et al. Superior In Vitro Stimulation of Human CD8+ T-Cells by Whole Virus versus Split Virus Influenza Vaccines. PLoS ONE 2014, 9, e103392. [Google Scholar] [CrossRef]
- Schultz, H.S.; Reedtz-Runge, S.L.; Bäckström, B.T.; Lamberth, K.; Pedersen, C.R.; Kvarnhammar, A.M.; ABIRISK consortium. Quantitative Analysis Of The CD4+ T Cell Response To Therapeutic Antibodies in Healthy Donors Using A Novel T Cell:PBMC Assay. PLoS ONE 2017, 12, e0178544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-W.; Liao, C.-H.; Ying, C.; Chang, C.-J.; Lin, C.-M. Ex Vivo Expansion Of Dendritic-Cell-Activated Antigen-Specific CD4+ T Cells With Anti-CD3/CD28, Interleukin-7, And Interleukin-15: Potential For Adoptive T Cell Immunotherapy. Clin. Immunol. 2006, 119, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.C.; Dayball, K.; Wan, Y.H.; Bramson, J. Detailed Analysis of the CD8+ T-Cell Response following Adenovirus Vaccination. J. Virol. 2003, 77, 13407–13411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Wang, Y.; Shuk-Yee Lo, A.; Gomes, E.M.; Junghans, R.P. Cell Density Plays a Critical Role in Ex Vivo Expansion of T Cells for Adoptive Immunotherapy. J. Biomed. Biotechnol. 2010, 2010, 386545. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Økern, G.; Rehan, S.; Beagley, L.; Lee, S.K.; Aarvak, T.; Schjetne, K.W.; Khanna, R. Ex Vivo Expansion Of Human T Cells For Adoptive Immunotherapy Using The Novel Xeno-Free CTS Immune Cell Serum Replacement. Clin. Transl. Immunol. 2015, 4, e31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canderan, G.; Gruarin, P.; Montagna, D.; Fontana, R.; Melloni, G.; Traversari, C.; Dellabona, P.; Casorati, G. An Efficient Strategy to Induce and Maintain In Vitro Human T Cells Specific for Autologous Non-Small Cell Lung Carcinoma. PLoS ONE 2010, 5, e12014. [Google Scholar] [CrossRef] [Green Version]
- Au-Yeung, B.B.; Zikherman, J.; Mueller, J.L.; Ashouri, J.F.; Matloubian, M.; Cheng, D.A.; Chen, Y.; Shokat, K.M.; Weiss, A. A Sharp T-Cell Antigen Receptor Signaling Threshold For T-Cell Proliferation. Proc. Natl. Acad. Sci. USA 2014, 111, E3679–E3688. [Google Scholar] [CrossRef] [Green Version]
- Irvine, D.J.; Purbhoo, M.A.; Krogsgaard, M.; Davis, M.M. Direct Observation Of Ligand Recognition By T Cells. Nature 2002, 419, 845–849. [Google Scholar] [CrossRef]
- Huang, J.; Brameshuber, M.; Zeng, X.; Xie, J.; Li, Q.; Chien, Y.; Valitutti, S.; Davis, M.M. A Single Peptide-Major Histocompatibility Complex Ligand Triggers Digital Cytokine Secretion in CD4+ T Cells. Immunity 2013, 39, 846–857. [Google Scholar] [CrossRef] [Green Version]
- Sojka, D.K.; Bruniquel, D.; Schwartz, R.H.; Singh, N.J. IL-2 Secretion By CD4+ T Cells In Vivo Is Rapid, Transient, and Influenced by TCR-Specific Competition. J. Immunol. 2004, 172, 6136–6143. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Norbury, C.C.; Greenwood, R.; Bennink, J.R.; Yewdell, J.W.; Frelinger, J.A. Multiple Paths For Activation Of Naive CD8+ T Cells: CD4-Independent Help. J. Immunol. 2001, 167, 1283–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.H.; Kim, T.; Song, S.Y.; Park, S.; Cho, H.H.; Jung, S.-H.; Ahn, J.-S.; Kim, H.-J.; Lee, J.-J.; Kim, H.-O.; et al. Naïve CD8+ T Cell Derived Tumor-Specific Cytotoxic Effectors As A Potential Remedy For Overcoming TGF-Β Immunosuppression In The Tumor Microenvironment. Sci. Rep. 2016, 6, 28208. [Google Scholar] [CrossRef] [PubMed]
- Geeraedts, F.; Goutagny, N.; Hornung, V.; Severa, M.; de Haan, A.; Pool, J.; Wilschut, J.; Fitzgerald, K.A.; Huckriede, A. Superior Immunogenicity of Inactivated Whole Virus H5N1 Influenza Vaccine is Primarily Controlled by Toll-like Receptor Signalling. PLoS Pathog. 2008, 4, e1000138. [Google Scholar] [CrossRef]
- Koyama, S.; Aoshi, T.; Tanimoto, T.; Kumagai, Y.; Kobiyama, K.; Tougan, T.; Sakurai, K.; Coban, C.; Horii, T.; Akira, S.; et al. Plasmacytoid Dendritic Cells Delineate Immunogenicity of Influenza Vaccine Subtypes. Sci. Transl. Med. 2010, 2, 25ra24. [Google Scholar] [CrossRef]
- Jeisy-Scott, V.; Kim, J.H.; Davis, W.G.; Cao, W.; Katz, J.M.; Sambhara, S. TLR7 Recognition Is Dispensable for Influenza Virus A Infection but Important for the Induction of Hemagglutinin-Specific Antibodies in Response to the 2009 Pandemic Split Vaccine in Mice. J. Virol. 2012, 86, 10988–10998. [Google Scholar] [CrossRef] [Green Version]
- Crowe, B.A.; Brühl, P.; Gerencer, M.; Schwendinger, M.G.; Pilz, A.; Kistner, O.; Koelling-Schlebusch, K.; Aichinger, G.; Singer, J.; Zeitlinger, M.; et al. Evaluation of the Cellular Immune Responses Induced by a Non-Adjuvanted Inactivated Whole Virus A/H5N1/VN/1203 Pandemic Influenza Vaccine in Humans. Vaccine 2010, 29, 166–173. [Google Scholar] [CrossRef]
- Gillard, P.; Chu, D.W.S.; Hwang, S.-J.; Yang, P.-C.; Thongcharoen, P.; Lim, F.S.; Dramé, M.; Walravens, K.; Roman, F. Long-Term Booster Schedules with AS03A-Adjuvanted Heterologous H5N1 Vaccines Induces Rapid and Broad Immune Responses in Asian Adults. BMC Infect. Dis. 2014, 14, 142. [Google Scholar] [CrossRef]
- Moris, P.; van der Most, R.; Leroux-Roels, I.; Clement, F.; Dramé, M.; Hanon, E.; Leroux-Roels, G.G.; Van Mechelen, M. H5N1 Influenza Vaccine Formulated with AS03A Induces Strong Cross-Reactive and Polyfunctional CD4 T-Cell Responses. J. Clin. Immunol. 2011, 31, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Begom, S.; Bermingham, A.; Ziegler, T.; Roberts, K.L.; Barclay, W.S.; Openshaw, P.; Lalvani, A. Predominance Of Heterosubtypic IFN-Γ-Only-Secreting Effector Memory T Cells In Pandemic H1N1 Naive Adults. Eur. J. Immunol. 2012, 42, 2913–2924. [Google Scholar] [CrossRef]
- Hillaire, M.L.B.; van Trierum, S.E.; Bodewes, R.; van Baalen, C.A.; van Binnendijk, R.S.; Koopmans, M.P.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Rimmelzwaan, G.F. Characterization Of The Human CD8+ T Cell Response Following Infection With 2009 Pandemic Influenza H1N1 Virus. J. Virol. 2011, 85, 12057–12061. [Google Scholar] [CrossRef] [Green Version]
- Wherry, E.J.; Teichgräber, V.; Becker, T.C.; Masopust, D.; Kaech, S.M.; Antia, R.; von Andrian, U.H.; Ahmed, R. Lineage Relationship And Protective Immunity Of Memory CD8 T Cell Subsets. Nat. Immunol. 2003, 4, 225–234. [Google Scholar] [CrossRef]
- Xie, D.; McElhaney, J.E. Lower GrB+ CD62Lhigh CD8 TCM effector lymphocyte response to influenza virus in older adults is associated with increased CD28null CD8 T lymphocytes. Mech. Ageing Dev. 2007, 128, 392–400. [Google Scholar] [CrossRef] [Green Version]
- Saurwein-Teissl, M.; Lung, T.L.; Marx, F.; Gschösser, C.; Asch, E.; Blasko, I.; Parson, W.; Böck, G.; Schönitzer, D.; Trannoy, E.; et al. Lack Of Antibody Production Following Immunization In Old Age: Association With CD8(+)CD28(−) T Cell Clonal Expansions And An Imbalance In The Production Of Th1 And Th2 Cytokines. J. Immunol. 2002, 168, 5893–5899. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Tsai, L.M.; Leong, Y.A.; Hu, X.; Ma, C.S.; Chevalier, N.; Sun, X.; Vandenberg, K.; Rockman, S.; Ding, Y.; et al. Circulating Precursor CCR7loPD-1hi CXCR5+ CD4+ T Cells Indicate Tfh Cell Activity and Promote Antibody Responses upon Antigen Reexposure. Immunity 2013, 39, 770–781. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, N.; Ueno, H. Blood Tfh Cells Come with Colors. Immunity 2013, 39, 629–630. [Google Scholar] [CrossRef] [Green Version]
- Abate, G.; Eslick, J.; Newman, F.K.; Frey, S.E.; Belshe, R.B.; Monath, T.P.; Hoft, D.F. Flow-cytometric detection of vaccinia-induced memory effector CD4(+), CD8(+), and gamma delta TCR(+) T cells capable of antigen-specific expansion and effector functions. J. Infect. Dis. 2005, 192, 1362–1371. [Google Scholar] [CrossRef] [Green Version]
- Hoft, D.F.; Babusis, E.; Worku, S.; Spencer, C.T.; Lottenbach, K.; Truscott, S.M.; Abate, G.; Sakala, I.G.; Edwards, K.M.; Creech, C.B.; et al. Live and inactivated influenza vaccines induce similar humoral responses, but only live vaccines induce diverse T-cell responses in young children. J. Infect. Dis. 2011, 204, 845–853. [Google Scholar] [CrossRef]
- Hoft, D.F.; Blazevic, A.; Stanley, J.; Landry, B.; Sizemore, D.; Kpamegan, E.; Gearhart, J.; Scott, A.; Kik, S.; Pau, M.G.; et al. A recombinant adenovirus expressing immunodominant TB antigens can significantly enhance BCG-induced human immunity. Vaccine 2012, 30, 2098–2108. [Google Scholar] [CrossRef]
- Hoft, D.F.; Lottenbach, K.; Goll, J.B.; Hill, H.; Winokur, P.L.; Patel, S.M.; Brady, R.C.; Chen, W.H.; Edwards, K.; Creech, C.B.; et al. Priming Vaccination With Influenza Virus H5 Hemagglutinin Antigen Significantly Increases the Duration of T cell Responses Induced by a Heterologous H5 Booster Vaccination. J. Infect. Dis. 2016, 214, 1020–1029. [Google Scholar] [CrossRef] [Green Version]
- Giese, C.; Marx, U. Human immunity in vitro—Solving immunogenicity and more. Adv. Drug Deliv. Rev. 2014, 69–70, 103–122. [Google Scholar] [CrossRef]
- Wieczorek, L.; Brown, B.K.; DelSarto Macedo, C.; Wesberry-Schmierer, M.; Ngauy, V.; Rosa Borges, A.; Michael, N.L.; Marovich, M.A.; Montefiori, D.C.; Polonis, V.R. Mitigation Of Variation Observed In A Peripheral Blood Mononuclear Cell (PBMC) Based HIV-1 Neutralization Assay By Donor Cell Pooling. Virology 2013, 447, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Oosting, M.; Deelen, P.; Ricaño-Ponce, I.; Smeekens, S.; Jaeger, M.; Matzaraki, V.; Swertz, M.A.; Xavier, R.J.; Franke, L.; et al. Inter-Individual Variability and Genetic Influences on Cytokine Responses To Bacteria And Fungi. Nat. Med. 2016, 22, 952–960. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tapia-Calle, G.; Born, P.A.; Koutsoumpli, G.; Gonzalez-Rodriguez, M.I.; Hinrichs, W.L.J.; Huckriede, A.L.W. A PBMC-Based System to Assess Human T Cell Responses to Influenza Vaccine Candidates In Vitro. Vaccines 2019, 7, 181. https://doi.org/10.3390/vaccines7040181
Tapia-Calle G, Born PA, Koutsoumpli G, Gonzalez-Rodriguez MI, Hinrichs WLJ, Huckriede ALW. A PBMC-Based System to Assess Human T Cell Responses to Influenza Vaccine Candidates In Vitro. Vaccines. 2019; 7(4):181. https://doi.org/10.3390/vaccines7040181
Chicago/Turabian StyleTapia-Calle, Gabriela, Philip A. Born, Georgia Koutsoumpli, Martin Ignacio Gonzalez-Rodriguez, Wouter L. J. Hinrichs, and Anke L. W. Huckriede. 2019. "A PBMC-Based System to Assess Human T Cell Responses to Influenza Vaccine Candidates In Vitro" Vaccines 7, no. 4: 181. https://doi.org/10.3390/vaccines7040181