Virus-Like Particles Are a Superior Platform for Presenting M2e Epitopes to Prime Humoral and Cellular Immunity against Influenza Virus

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Influenza 5xM2e Vaccines and Virus Reagents
2.2. Immunization and Challenges of Mice
2.3. Lung Viral Titers
2.4. Sample Preparation and Antibody Responses
2.5. Bone Marrow-Derived Dendritic Cell (BMDC) Culture and Cytokine ELISA
2.6. Enzyme-Linked Immunospot (ELISPOT) Assay
2.7. Flow Cytometry
2.8. Protective Assay of Immune Sera
2.9. Intraperitoneal Injection to Determine Acute Immune Responses
2.10. Statistics
3. Results
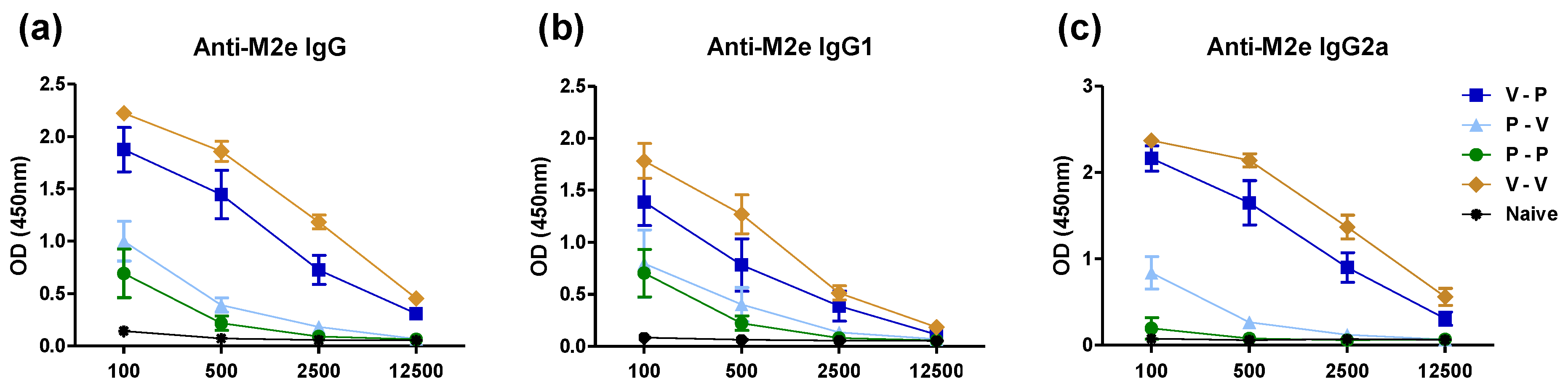
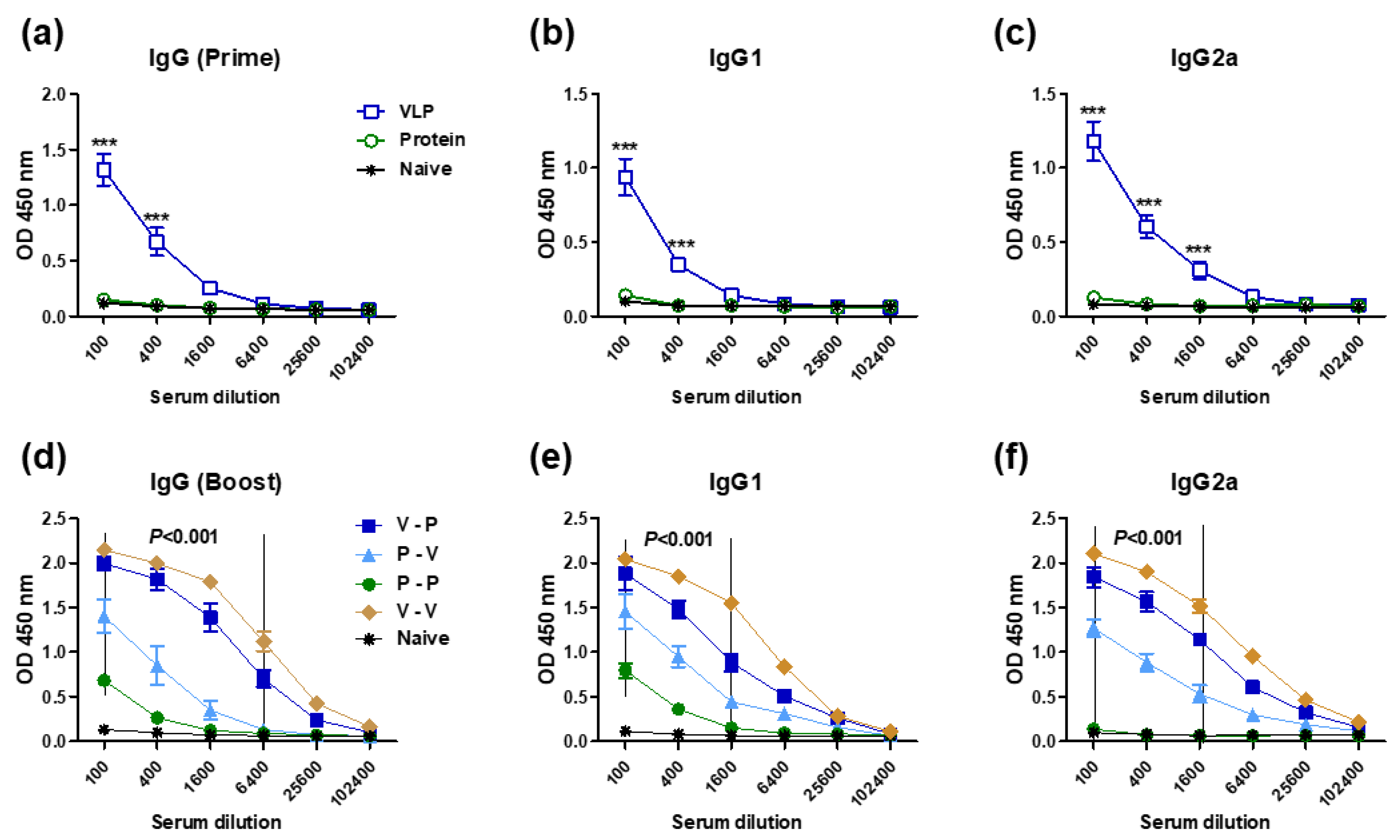
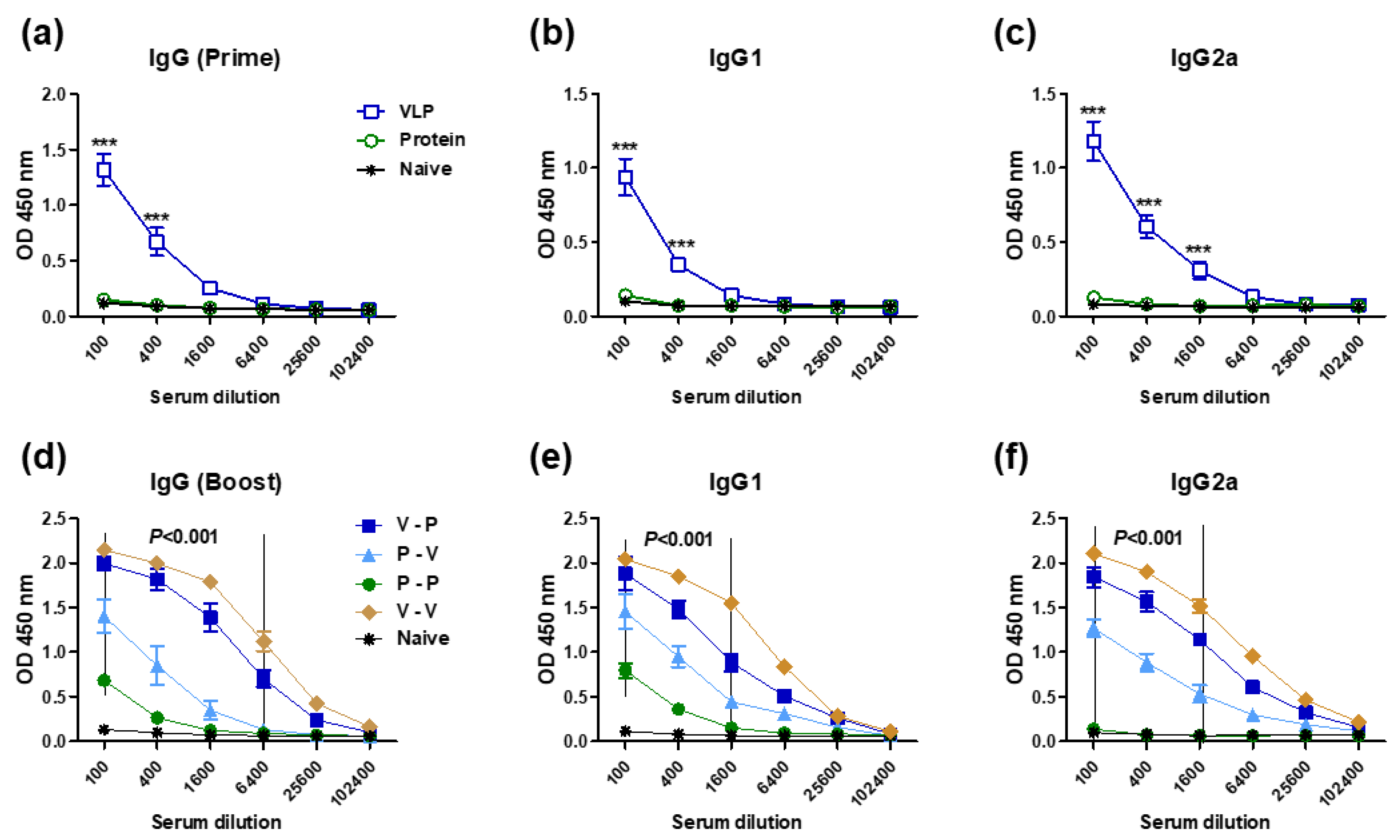
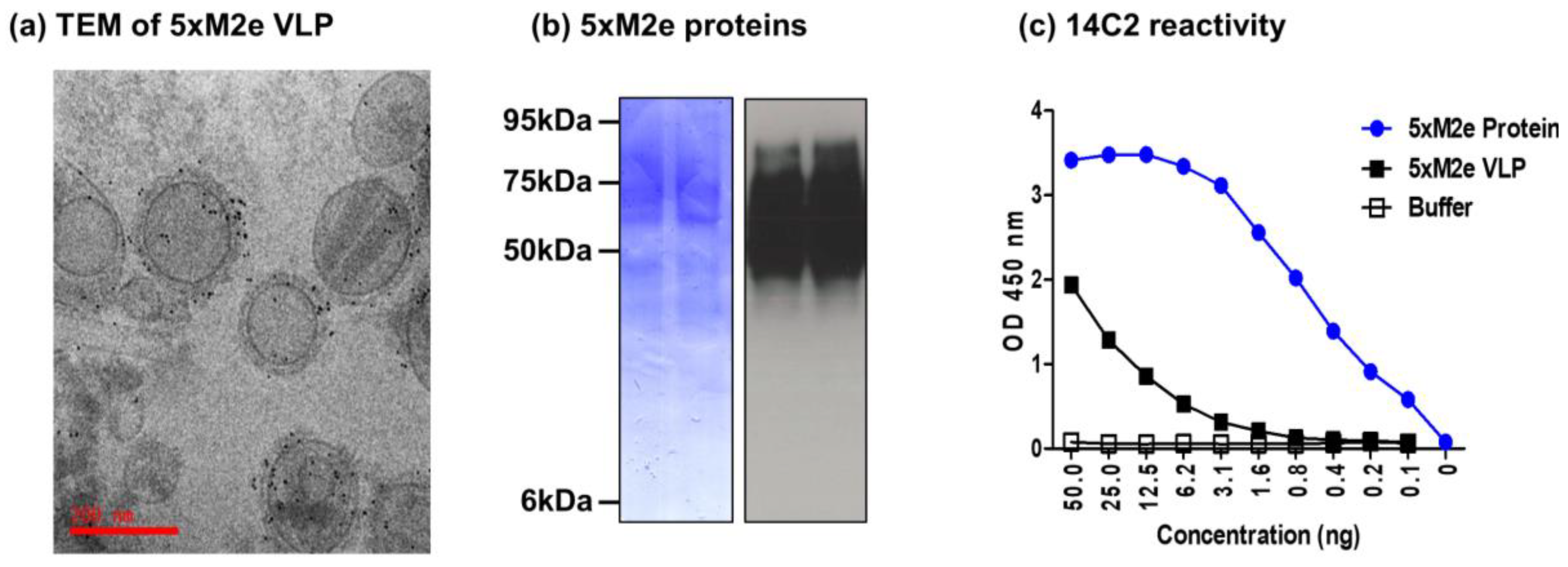
3.1. The 5xM2e VLP but Not the 5xM2e Protein Is Immunogenic and Effective in Eliciting IgG2a Isotype Antibodies
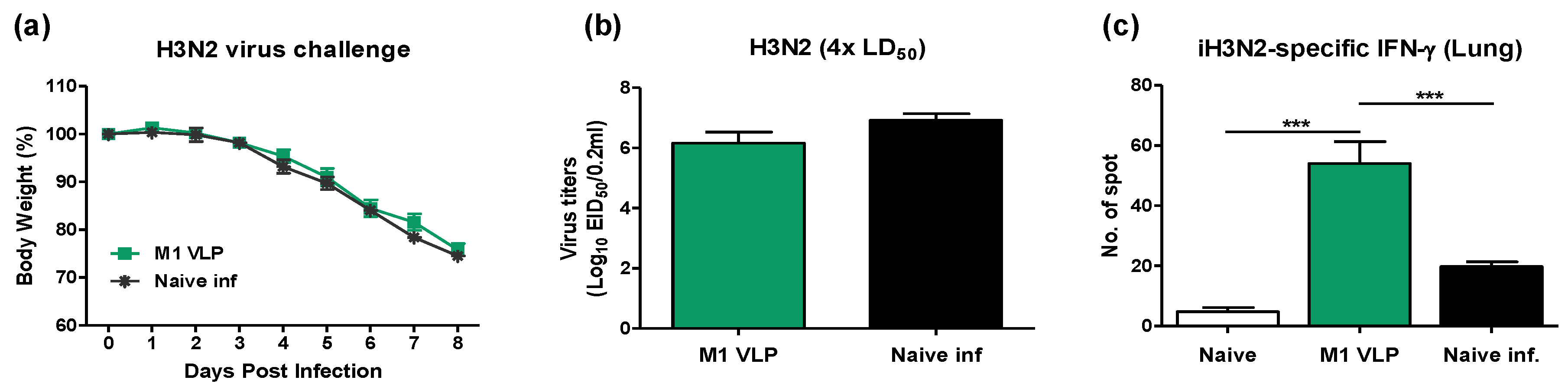
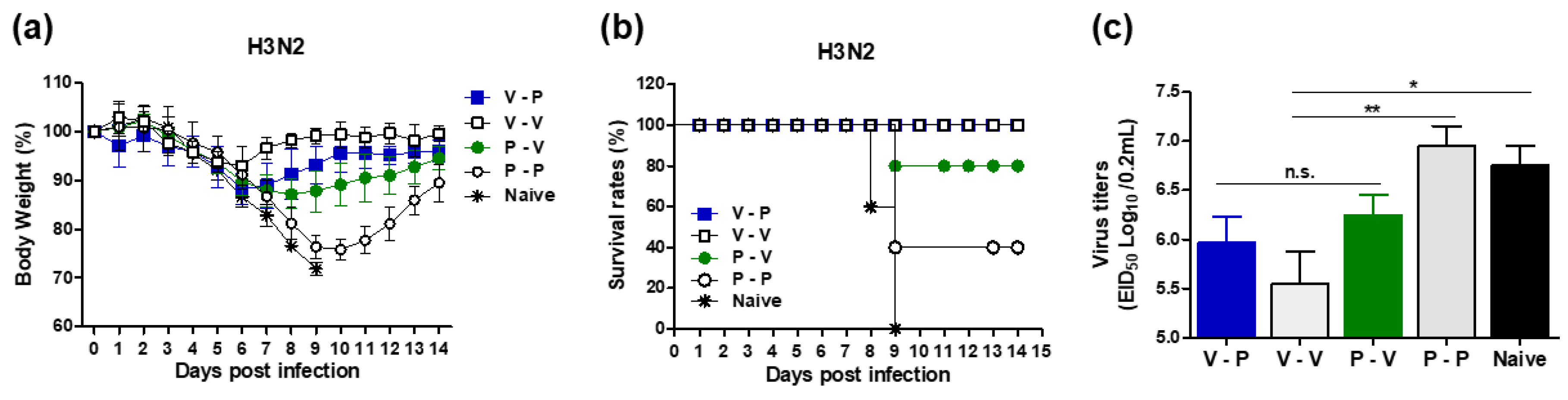
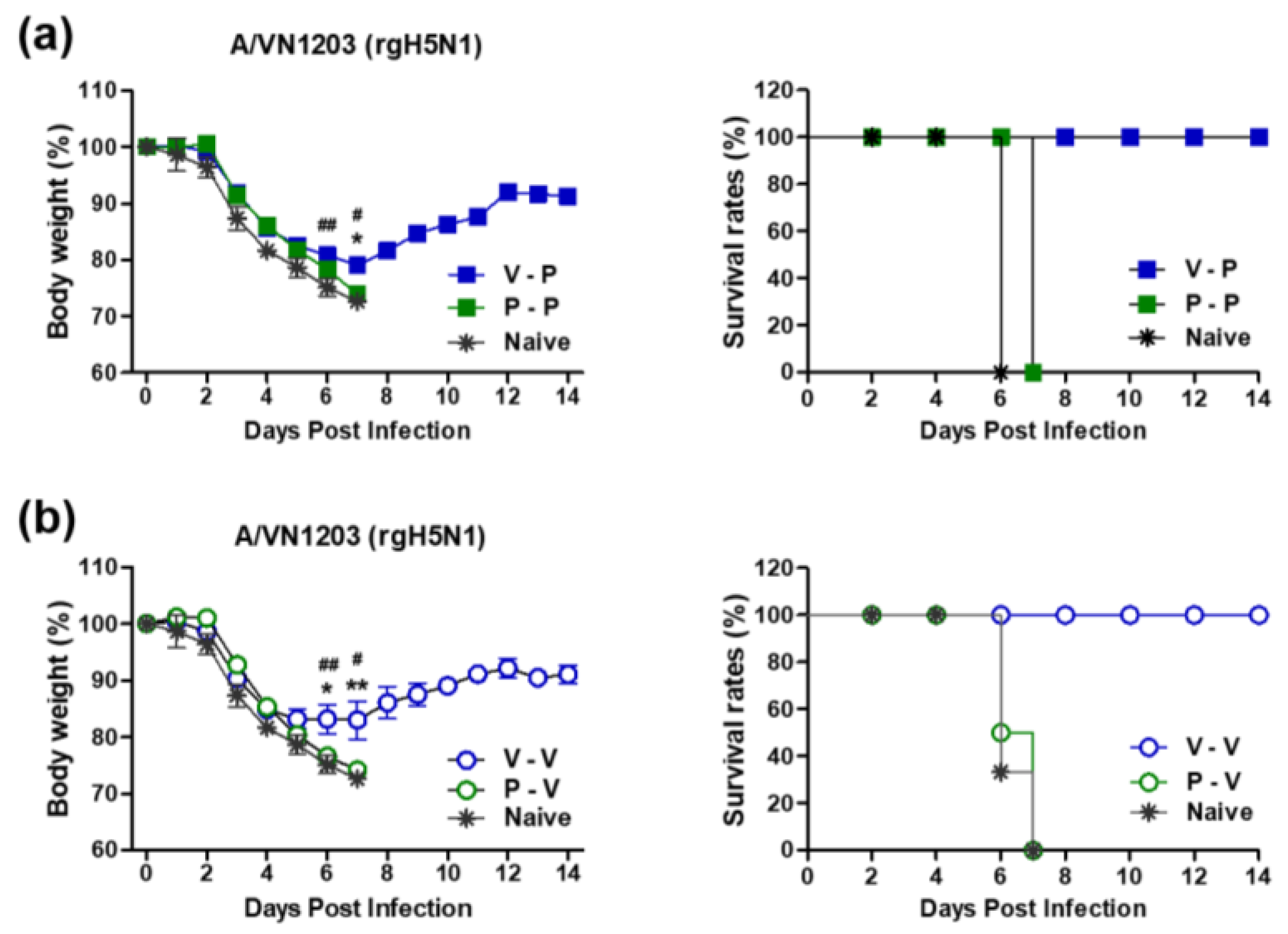
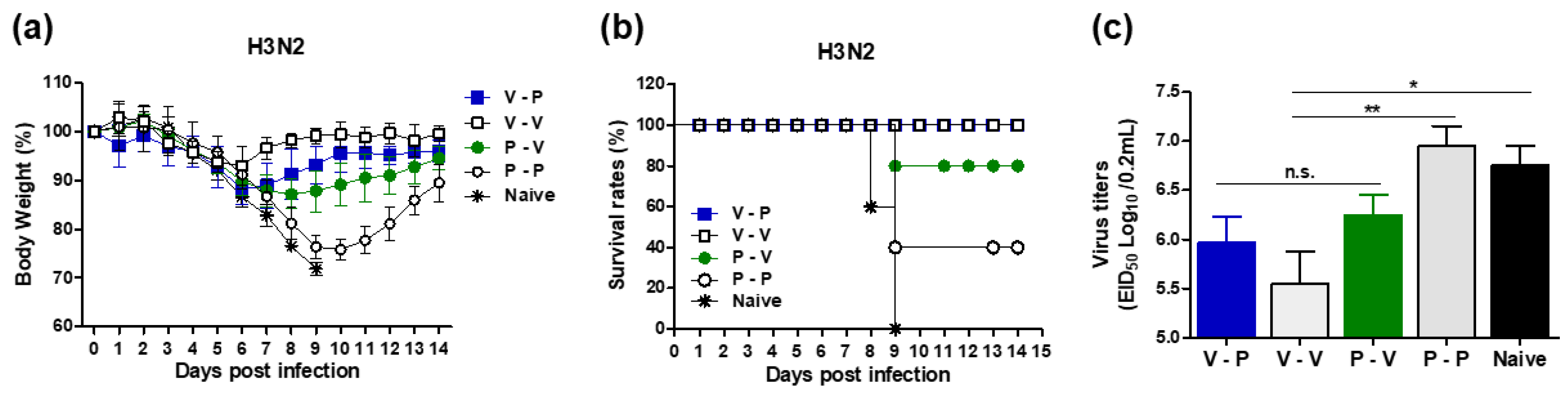
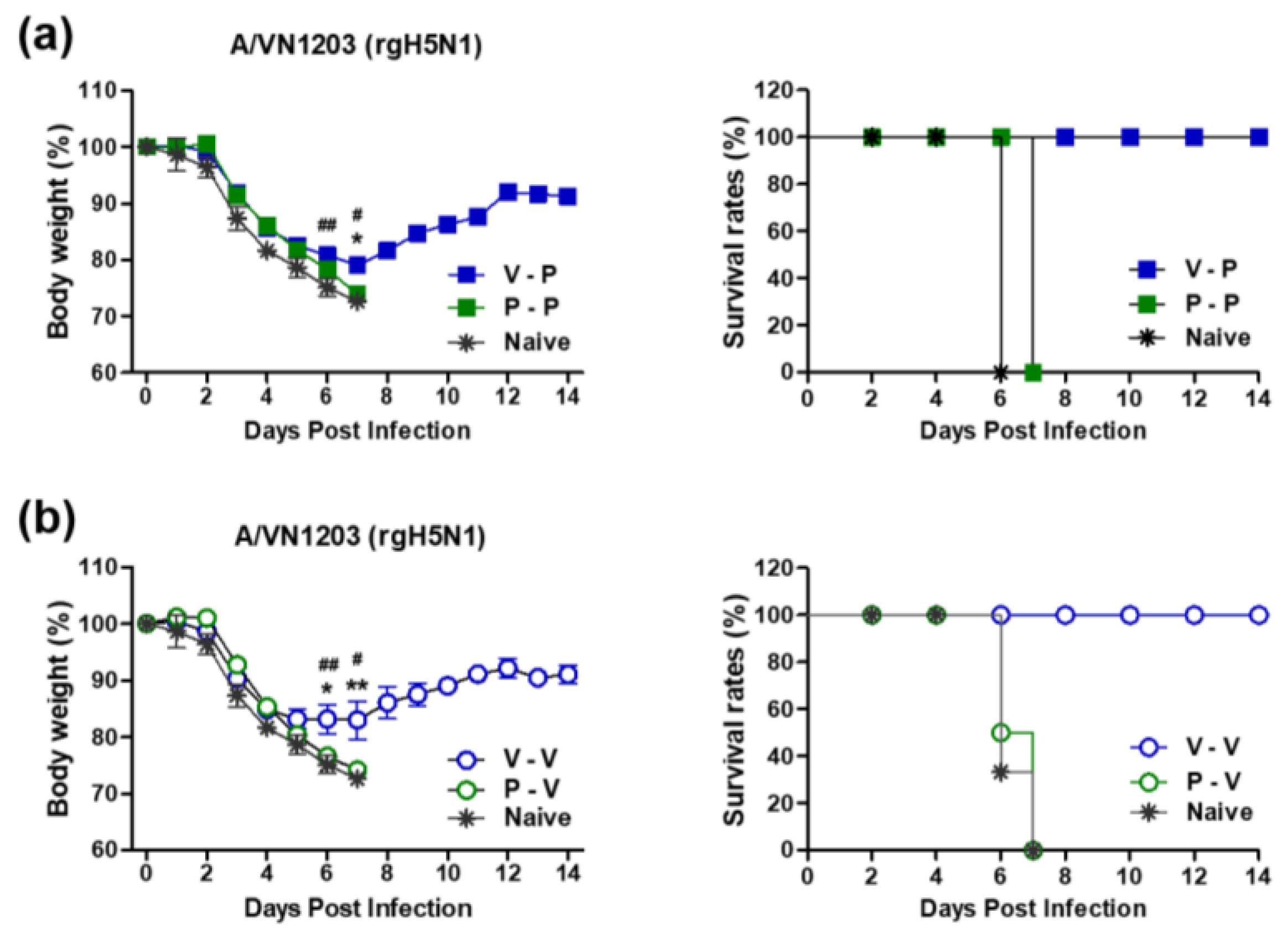
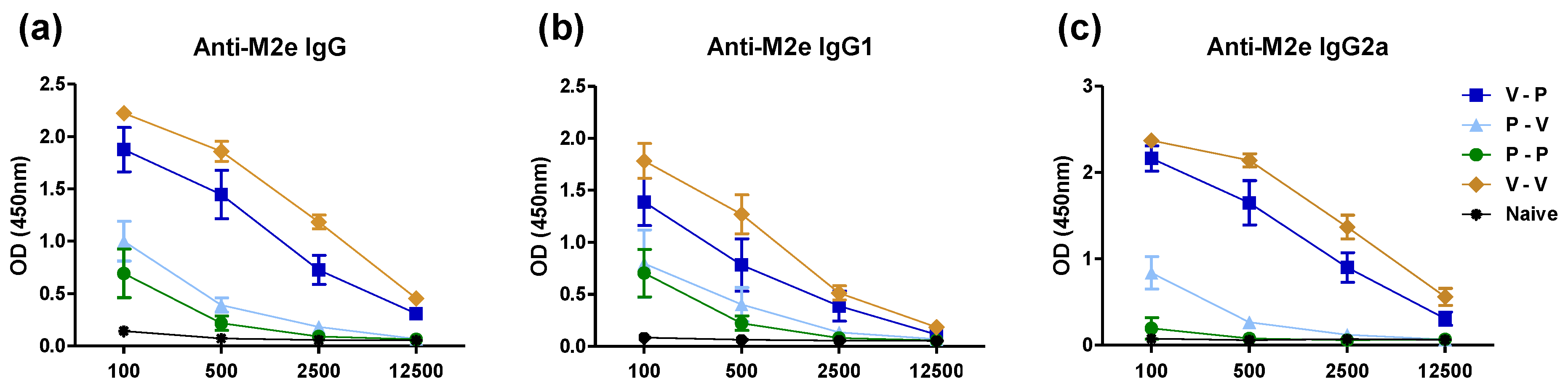
3.2. Heterologous VLP Prime and Protein Boost Improves Protection Compared to Protein-Only Immunizations
3.3. The 5xM2e Immune Sera from VLP-Primed but Not Protein-Primed Mice Confer Protection
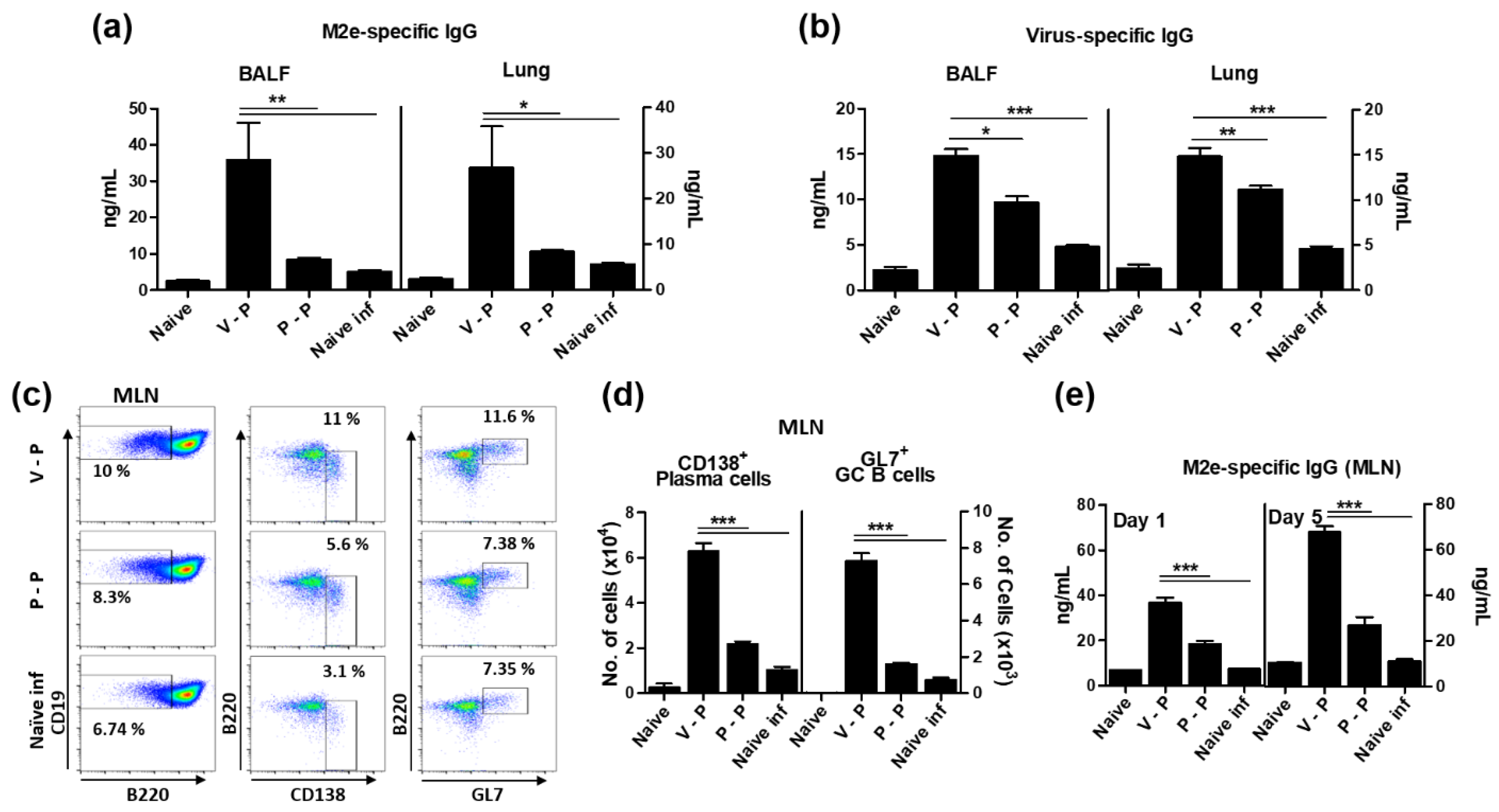
3.4. The 5xM2e VLP Is Superior to 5xM2e Protein in Inducing B Cell Responses
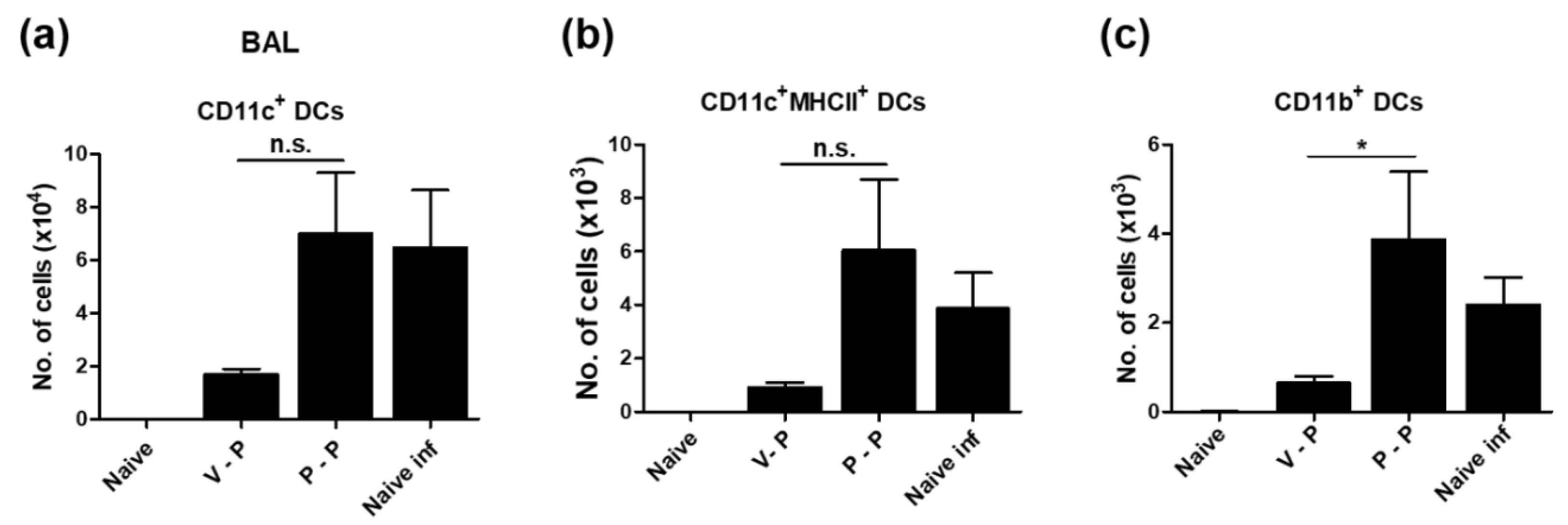
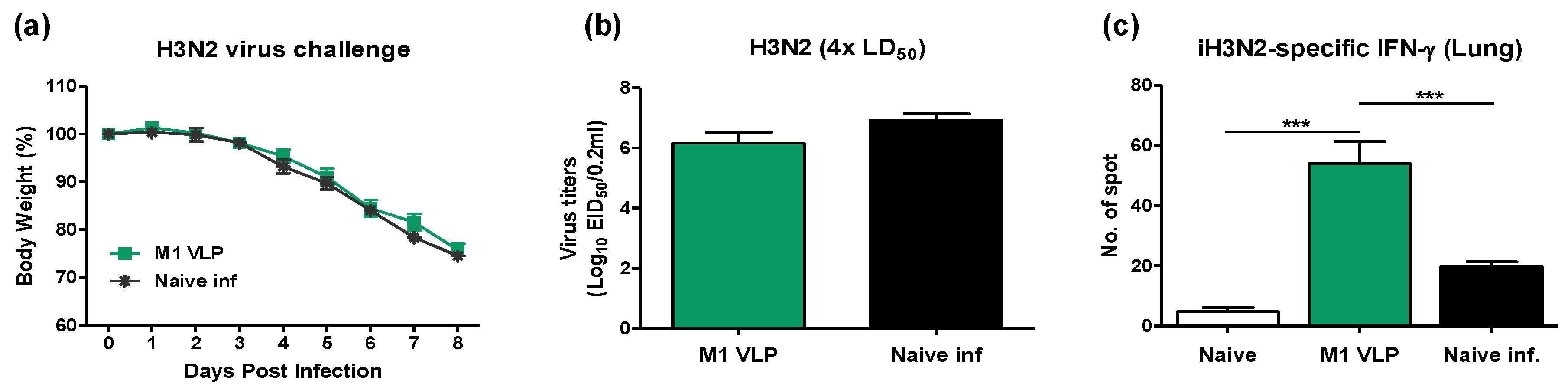
3.5. VLP-Primed Mice Prevent Excessive Dendritic Cell Recruitment upon Virus Infection
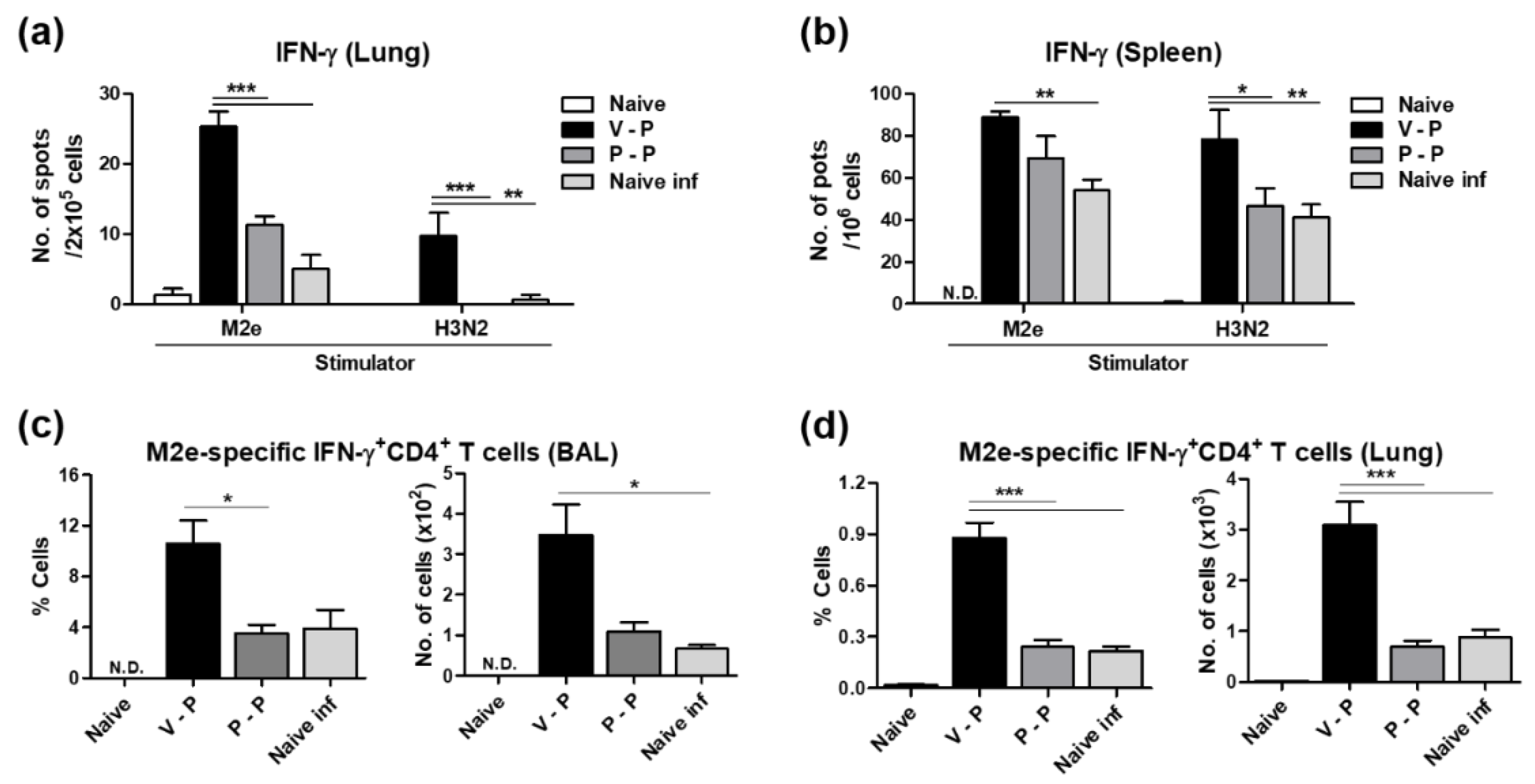
3.6. The 5xM2e VLP Platform Is More Effective in Inducing Effector CD4 T Cell Immune Responses than Protein
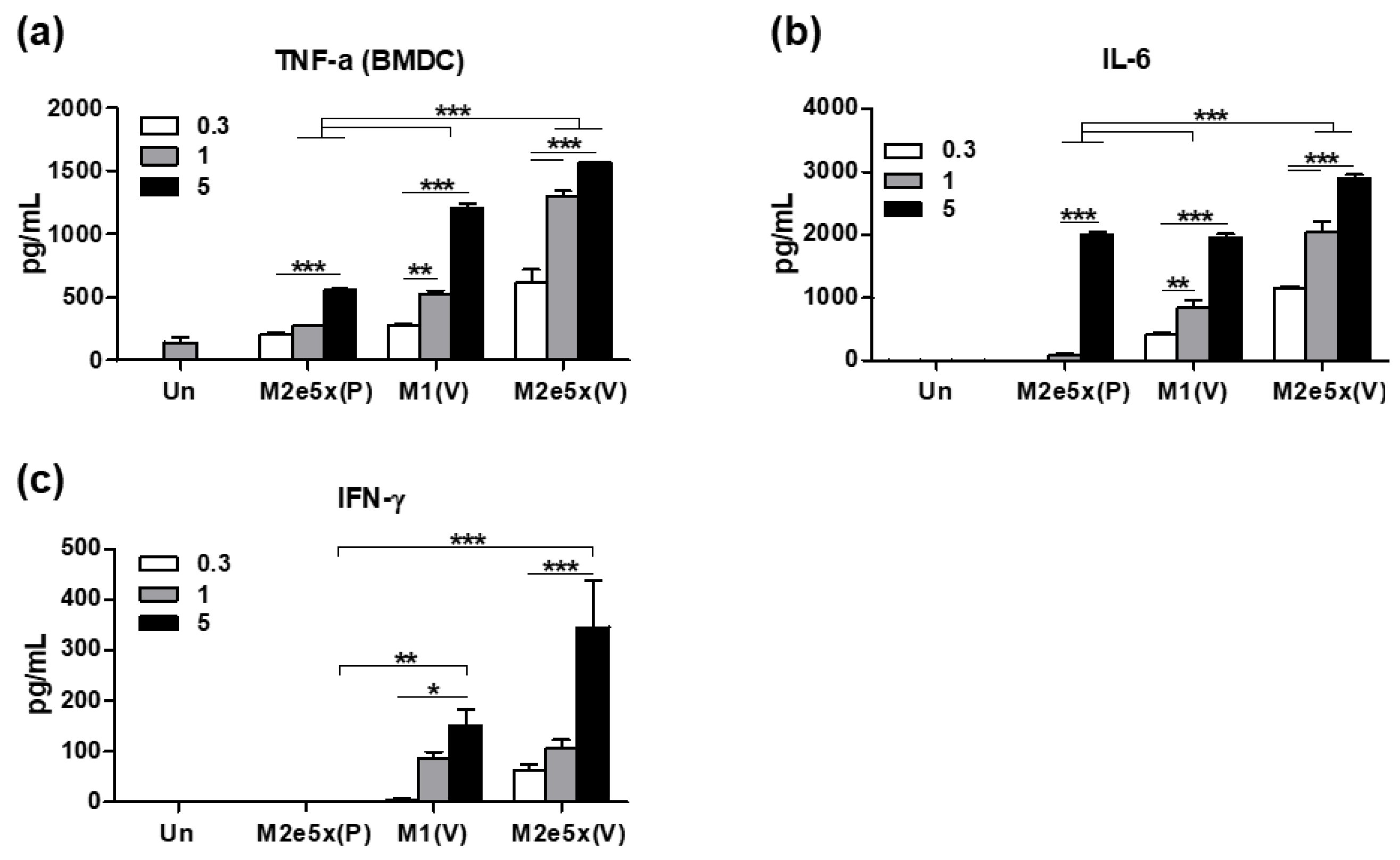
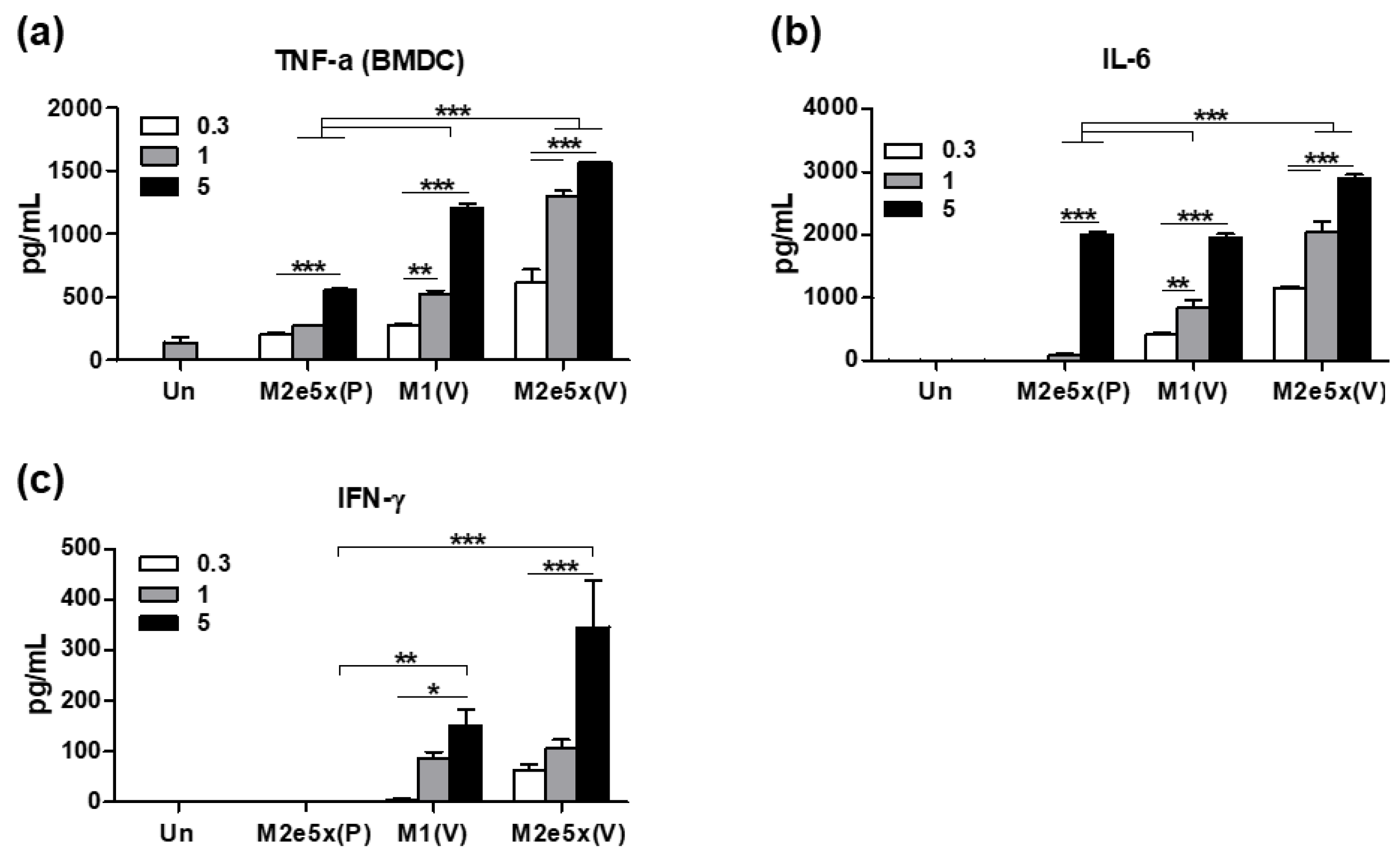
3.7. The 5xM2e VLP Is More Effective in Stimulating DCs In Vitro to Secrete Inflammatory Cytokines than 5xM2e Proteins
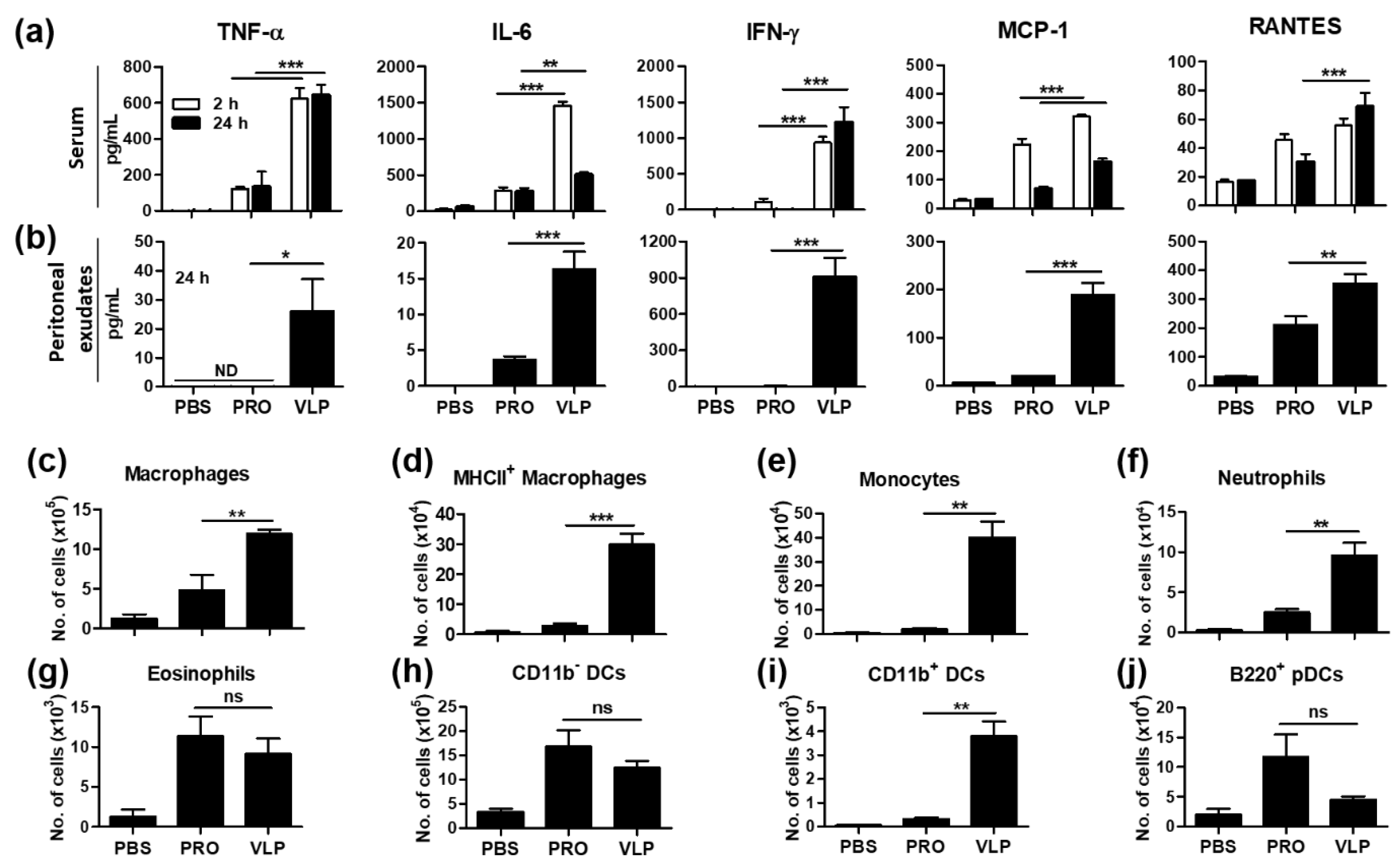
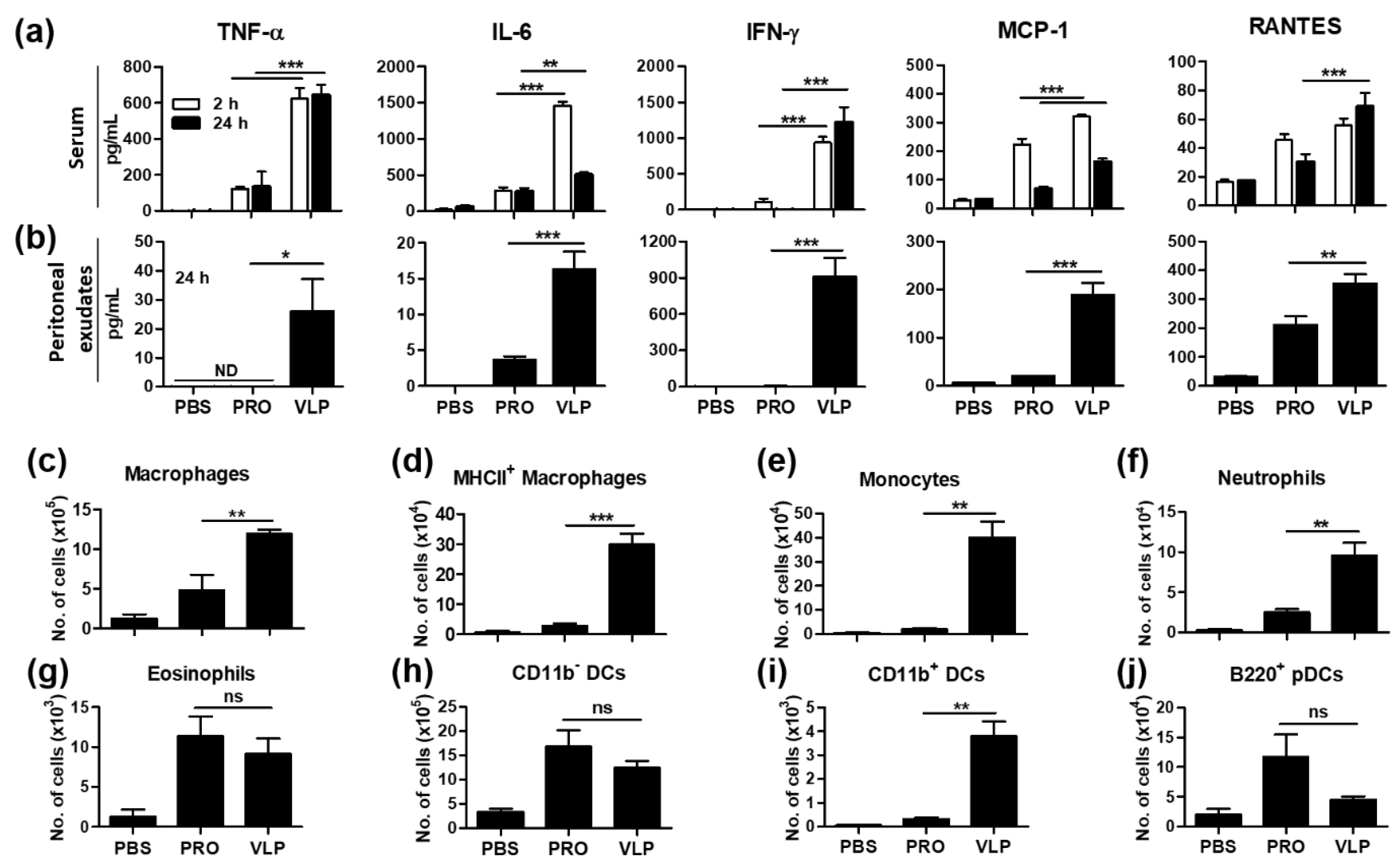
3.8. VLP Is Effective in the Acute Induction of Cytokines and Recruiting Innate Immune Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

References
- Liu, W.; Zou, P.; Ding, J.; Lu, Y.; Chen, Y.H. Sequence comparison between the extracellular domain of M2 protein human and avian influenza A virus provides new information for bivalent influenza vaccine design. Microbes Infect. 2005, 7, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Reed, C.; Blair, P.J.; Katz, J.M.; Hancock, K. Serum antibody response to matrix protein 2 following natural infection with 2009 pandemic influenza A(H1N1) virus in humans. J. Infect. Dis. 2014, 209, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, M.; Mozdzanowska, K.; Zharikova, D.; Hoff, H.; Wunner, W.; Couch, R.B.; Gerhard, W. Influenza A virus infection engenders a poor antibody response against the ectodomain of matrix protein 2. Virol. J. 2006, 3, 102. [Google Scholar] [CrossRef] [PubMed]
- Bernelin-Cottet, C.; Deloizy, C.; Stanek, O.; Barc, C.; Bouguyon, E.; Urien, C.; Boulesteix, O.; Pezant, J.; Richard, C.A.; Moudjou, M.; et al. A Universal Influenza Vaccine Can Lead to Disease Exacerbation or Viral Control Depending on Delivery Strategies. Front. Immunol. 2016, 7, 641. [Google Scholar] [CrossRef] [PubMed]
- Herve, P.L.; Raliou, M.; Bourdieu, C.; Dubuquoy, C.; Petit-Camurdan, A.; Bertho, N.; Eleouet, J.F.; Chevalier, C.; Riffault, S. A novel subnucleocapsid nanoplatform for mucosal vaccination against influenza virus that targets the ectodomain of matrix protein 2. J. Virol. 2014, 88, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Neirynck, S.; Deroo, T.; Saelens, X.; Vanlandschoot, P.; Jou, W.M.; Fiers, W. A universal influenza A vaccine based on the extracellular domain of the M2 protein. Nat. Med. 1999, 5, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Jegerlehner, A.; Schmitz, N.; Storni, T.; Bachmann, M.F. Influenza A vaccine based on the extracellular domain of M2: Weak protection mediated via antibody-dependent NK cell activity. J. Immunol. 2004, 172, 5598–5605. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Liang, X.; Horton, M.S.; Perry, H.C.; Citron, M.P.; Heidecker, G.J.; Fu, T.M.; Joyce, J.; Przysiecki, C.T.; Keller, P.M.; et al. Preclinical study of influenza virus A M2 peptide conjugate vaccines in mice, ferrets, and rhesus monkeys. Vaccine 2004, 22, 2993–3003. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.M.; Grimm, K.M.; Citron, M.P.; Freed, D.C.; Fan, J.; Keller, P.M.; Shiver, J.W.; Liang, X.; Joyce, J.G. Comparative immunogenicity evaluations of influenza A virus M2 peptide as recombinant virus like particle or conjugate vaccines in mice and monkeys. Vaccine 2009, 27, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, R.M.; Przysiecki, C.T.; Liang, X.; Garsky, V.M.; Fan, J.; Wang, B.; Troutman, R.; Rippeon, Y.; Flanagan, E.; Shiver, J.; et al. Pharmaceutical and immunological evaluation of human papillomavirus viruslike particle as an antigen carrier. J. Pharm. Sci. 2006, 95, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, S.M.; Zhao, Z.S.; Lo, C.Y.; Misplon, J.A.; Liu, T.; Ye, Z.; Hogan, R.J.; Wu, Z.; Benton, K.A.; Tumpey, T.M.; et al. Matrix protein 2 vaccination and protection against influenza viruses, including subtype H5N1. Emerg. Infect. Dis. 2007, 13, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Ernst, W.A.; Kim, H.J.; Tumpey, T.M.; Jansen, A.D.; Tai, W.; Cramer, D.V.; Adler-Moore, J.P.; Fujii, G. Protection against H1, H5, H6 and H9 influenza A infection with liposomal matrix 2 epitope vaccines. Vaccine 2006, 24, 5158–5168. [Google Scholar] [CrossRef] [PubMed]
- Eliasson, D.G.; Bakkouri, K.E.; Schon, K.; Ramne, A.; Festjens, E.; Lowenadler, B.; Fiers, W.; Saelens, X.; Lycke, N. CTA1-M2e-DD: A novel mucosal adjuvant targeted influenza vaccine. Vaccine 2008, 26, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Bessa, J.; Schmitz, N.; Hinton, H.J.; Schwarz, K.; Jegerlehner, A.; Bachmann, M.F. Efficient induction of mucosal and systemic immune responses by virus-like particles administered intranasally: Implications for vaccine design. Eur. J. Immunol. 2008, 38, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Turley, C.B.; Rupp, R.E.; Johnson, C.; Taylor, D.N.; Wolfson, J.; Tussey, L.; Kavita, U.; Stanberry, L.; Shaw, A. Safety and immunogenicity of a recombinant M2e-flagellin influenza vaccine (STF2.4xM2e) in healthy adults. Vaccine 2011, 29, 5145–5152. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yuan, X.Y.; Li, J.; Chen, Y.H. The co-administration of CpG-ODN influenced protective activity of influenza M2e vaccine. Vaccine 2009, 27, 4320–4324. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Peng, Z.; Liu, Z.; Lu, Y.; Ding, J.; Chen, Y.H. High epitope density in a single recombinant protein molecule of the extracellular domain of influenza A virus M2 protein significantly enhances protective immunity. Vaccine 2004, 23, 366–371. [Google Scholar] [CrossRef] [PubMed]
- De Filette, M.; Martens, W.; Roose, K.; Deroo, T.; Vervalle, F.; Bentahir, M.; Vandekerckhove, J.; Fiers, W.; Saelens, X. An influenza A vaccine based on tetrameric ectodomain of matrix protein 2. J. Biol. Chem. 2008, 283, 11382–11387. [Google Scholar] [CrossRef] [PubMed]
- Fiers, W.; De Filette, M.; Birkett, A.; Neirynck, S.; Min Jou, W. A “universal” human influenza A vaccine. Virus Res. 2004, 103, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Galarza, J.M.; Latham, T.; Cupo, A. Virus-like particle (VLP) vaccine conferred complete protection against a lethal influenza virus challenge. Viral Immunol. 2005, 18, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Pushko, P.; Tumpey, T.M.; Bu, F.; Knell, J.; Robinson, R.; Smith, G. Influenza virus-like particles comprised of the HA, NA, and M1 proteins of H9N2 influenza virus induce protective immune responses in BALB/c mice. Vaccine 2005, 23, 5751–5759. [Google Scholar] [CrossRef] [PubMed]
- Quan, F.S.; Huang, C.; Compans, R.W.; Kang, S.M. Virus-like particle vaccine induces protective immunity against homologous and heterologous strains of influenza virus. J. Virol. 2007, 81, 3514–3524. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.C.; Song, J.M.; Eunju, O.; Kwon, Y.M.; Lee, Y.J.; Compans, R.W.; Kang, S.M. Virus-like particles containing multiple M2 extracellular domains confer improved cross-protection against various subtypes of influenza virus. Mol. Ther. J. Am. Soc. Gene Ther. 2013, 21, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Kim, M.C.; Lee, Y.T.; Hwang, H.S.; Lee, J.; Kim, C.; Kang, S.M. Cross Protection against Influenza A Virus by Yeast-Expressed Heterologous Tandem Repeat M2 Extracellular Proteins. PLoS ONE 2015, 10, e0137822. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.A.; Singh, M. Acceptable levels of endotoxin in vaccine formulations during preclinical research. J. Pharm. Sci. 2011, 100, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Quan, F.S.; Compans, R.W.; Nguyen, H.H.; Kang, S.M. Induction of heterosubtypic immunity to influenza virus by intranasal immunization. J. Virol. 2008, 82, 1350–1359. [Google Scholar] [CrossRef] [PubMed]
- Fries, L.F.; Smith, G.E.; Glenn, G.M. A recombinant viruslike particle influenza A (H7N9) vaccine. N. Engl. J. Med. 2013, 369, 2564–2566. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Macias, C.; Ferat-Osorio, E.; Tenorio-Calvo, A.; Isibasi, A.; Talavera, J.; Arteaga-Ruiz, O.; Arriaga-Pizano, L.; Hickman, S.P.; Allende, M.; Lenhard, K.; et al. Safety and immunogenicity of a virus-like particle pandemic influenza A (H1N1) 2009 vaccine in a blinded, randomized, placebo-controlled trial of adults in Mexico. Vaccine 2011, 29, 7826–7834. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Wu, J.; Verma, N.; Verma, S.; Raghunandan, R.; Manischewitz, J.; King, L.R.; Kpamegan, E.; Pincus, S.; Smith, G.; et al. H5N1 virus-like particle vaccine elicits cross-reactive neutralizing antibodies that preferentially bind to the oligomeric form of influenza virus hemagglutinin in humans. J. Virol. 2011, 85, 10945–10954. [Google Scholar] [CrossRef] [PubMed]
- Song, J.M.; Van Rooijen, N.; Bozja, J.; Compans, R.W.; Kang, S.M. Vaccination inducing broad and improved cross protection against multiple subtypes of influenza A virus. Proc. Natl. Acad. Sci. USA 2011, 108, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Black, R.A.; Rota, P.A.; Gorodkova, N.; Klenk, H.D.; Kendal, A.P. Antibody response to the M2 protein of influenza A virus expressed in insect cells. J. Gen. Virol. 1993, 74 (Pt 1), 143–146. [Google Scholar] [CrossRef]
- Lee, Y.N.; Lee, Y.T.; Kim, M.C.; Gewirtz, A.T.; Kang, S.M. A Novel Vaccination Strategy Mediating the Induction of Lung-Resident Memory CD8 T Cells Confers Heterosubtypic Immunity against Future Pandemic Influenza Virus. J. Immunol. 2016, 196, 2637–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.N.; Kim, M.C.; Lee, Y.T.; Hwang, H.S.; Cho, M.K.; Lee, J.S.; Ko, E.J.; Kwon, Y.M.; Kang, S.M. AS04-adjuvanted virus-like particles containing multiple M2 extracellular domains of influenza virus confer improved protection. Vaccine 2014, 32, 4578–4585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennings, G.T.; Bachmann, M.F. The coming of age of virus-like particle vaccines. Biol. Chem. 2008, 389, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Chackerian, B. Virus-like particles: Flexible platforms for vaccine development. Expert Rev. Vaccines 2007, 6, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Huber, V.C.; McKeon, R.M.; Brackin, M.N.; Miller, L.A.; Keating, R.; Brown, S.A.; Makarova, N.; Perez, D.R.; Macdonald, G.H.; McCullers, J.A. Distinct contributions of vaccine-induced immunoglobulin G1 (IgG1) and IgG2a antibodies to protective immunity against influenza. Clin. Vaccine Immunol. 2006, 13, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Gessner, J.E.; Heiken, H.; Tamm, A.; Schmidt, R.E. The IgG Fc receptor family. Ann. Hematol. 1998, 76, 231–248. [Google Scholar] [CrossRef] [PubMed]
- El Bakkouri, K.; Descamps, F.; De Filette, M.; Smet, A.; Festjens, E.; Birkett, A.; Van Rooijen, N.; Verbeek, S.; Fiers, W.; Saelens, X. Universal vaccine based on ectodomain of matrix protein 2 of influenza A: Fc receptors and alveolar macrophages mediate protection. J. Immunol. 2011, 186, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Hinton, H.J.; Jegerlehner, A.; Bachmann, M.F. Pattern recognition by B cells: The role of antigen repetitiveness versus Toll-like receptors. Curr. Top. Microbiol. Immunol. 2008, 319, 1–15. [Google Scholar] [PubMed]
- Song, J.M.; Hossain, J.; Yoo, D.G.; Lipatov, A.S.; Davis, C.T.; Quan, F.S.; Chen, L.M.; Hogan, R.J.; Donis, R.O.; Compans, R.W.; et al. Protective immunity against H5N1 influenza virus by a single dose vaccination with virus-like particles. Virology 2010, 405, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.J.; Bourgeois, M.; Quan, F.S.; Lipatov, A.S.; Song, J.M.; Chen, L.M.; Compans, R.W.; York, I.; Kang, S.M.; Donis, R.O. Virus-Like Particle Vaccine Containing Hemagglutinin Confers Protection against 2009 H1N1 Pandemic Influenza. Clin. Vaccine immunol. 2011, 18, 2010–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pone, E.J.; Xu, Z.; White, C.A.; Zan, H.; Casali, P. B cell TLRs and induction of immunoglobulin class-switch DNA recombination. Front. Biosci. 2012, 17, 2594–2615. [Google Scholar] [CrossRef]
- Kang, S.M.; Yoo, D.G.; Kim, M.C.; Song, J.M.; Park, M.K.; Eunju, O.; Quan, F.S.; Akira, S.; Compans, R.W. MyD88 Plays an Essential Role in Inducing B Cells Capable of Differentiating into Antibody-Secreting Cells after Vaccination. J. Virol. 2011, 85, 11391–11400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingulli, E.; Funatake, C.; Jacovetty, E.L.; Zanetti, M. Cutting edge: Antigen presentation to CD8 T cells after influenza A virus infection. J. Immunol. 2009, 182, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Braciale, T.J. Respiratory dendritic cell subsets differ in their capacity to support the induction of virus-specific cytotoxic CD8+ T cell responses. PLoS ONE 2009, 4, e4204. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Tato, A.; Leon, B.; Lund, F.E.; Randall, T.D. Temporal changes in dendritic cell subsets, cross-priming and costimulation via CD70 control CD8(+) T cell responses to influenza. Nat. Immunol. 2010, 11, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Moltedo, B.; Li, W.; Yount, J.S.; Moran, T.M. Unique type I interferon responses determine the functional fate of migratory lung dendritic cells during influenza virus infection. PLoS Pathog. 2011, 7, e1002345. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Jandus, C.; Maurer, P.; Hammann-Haenni, A.; Schwarz, K.; Bachmann, M.F.; Speiser, D.E.; Romero, P. Virus-like particles induce robust human T-helper cell responses. Eur. J. Immunol. 2012, 42, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Yarilina, A.; Park-Min, K.H.; Antoniv, T.; Hu, X.; Ivashkiv, L.B. TNF activates an IRF1-dependent autocrine loop leading to sustained expression of chemokines and STAT1-dependent type I interferon-response genes. Nat. Immunol. 2008, 9, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Lauder, S.N.; Jones, E.; Smart, K.; Bloom, A.; Williams, A.S.; Hindley, J.P.; Ondondo, B.; Taylor, P.R.; Clement, M.; Fielding, C.; et al. Interleukin-6 limits influenza-induced inflammation and protects against fatal lung pathology. Eur. J. Immunol. 2013, 43, 2613–2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddahri, F.; Denanglaire, S.; Bureau, F.; Spolski, R.; Leonard, W.J.; Leo, O.; Andris, F. Interleukin-6/STAT3 signaling regulates the ability of naive T cells to acquire B-cell help capacities. Blood 2009, 113, 2426–2433. [Google Scholar] [CrossRef] [PubMed]
- Le, Y.; Zhou, Y.; Iribarren, P.; Wang, J. Chemokines and chemokine receptors: Their manifold roles in homeostasis and disease. Cell. Mol. Immunol. 2004, 1, 95–104. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5xM2e Vaccine | M2e Sequences (2–20) | Challenge Virus | M2 |
|---|---|---|---|
| Human (2×) | SLLTEVETPIRNEWGSRSN (consensus) | H3N2 (A/Philippines) | Human M2 |
| Swine (1×) | SLLTEVETPTRSEWESRSS (A/California/4/2009) | rgH5N1 (A/PR8 backbone) | Human M2 |
| Avian I (1×) | SLLTEVETPTRNEWESRSS (most common) | ||
| Avian II (1×) | SLLTEVETLTRNGWGCRCS (second common H5, H9) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.-H.; Kwon, Y.-M.; Lee, Y.-T.; Kim, M.-C.; Hwang, H.S.; Ko, E.-J.; Lee, Y.; Choi, H.-J.; Kang, S.-M. Virus-Like Particles Are a Superior Platform for Presenting M2e Epitopes to Prime Humoral and Cellular Immunity against Influenza Virus. Vaccines 2018, 6, 66. https://doi.org/10.3390/vaccines6040066
Kim K-H, Kwon Y-M, Lee Y-T, Kim M-C, Hwang HS, Ko E-J, Lee Y, Choi H-J, Kang S-M. Virus-Like Particles Are a Superior Platform for Presenting M2e Epitopes to Prime Humoral and Cellular Immunity against Influenza Virus. Vaccines. 2018; 6(4):66. https://doi.org/10.3390/vaccines6040066
Chicago/Turabian StyleKim, Ki-Hye, Young-Man Kwon, Young-Tae Lee, Min-Chul Kim, Hye Suk Hwang, Eun-Ju Ko, Youri Lee, Hyo-Jick Choi, and Sang-Moo Kang. 2018. "Virus-Like Particles Are a Superior Platform for Presenting M2e Epitopes to Prime Humoral and Cellular Immunity against Influenza Virus" Vaccines 6, no. 4: 66. https://doi.org/10.3390/vaccines6040066