The Morphological Features and Biology of a Relict and Endangered Woody Plant Species: Chamaedaphne calyculata (L.) Moench (Ericaceae)


Abstract
:1. Introduction
2. Materials and Methods
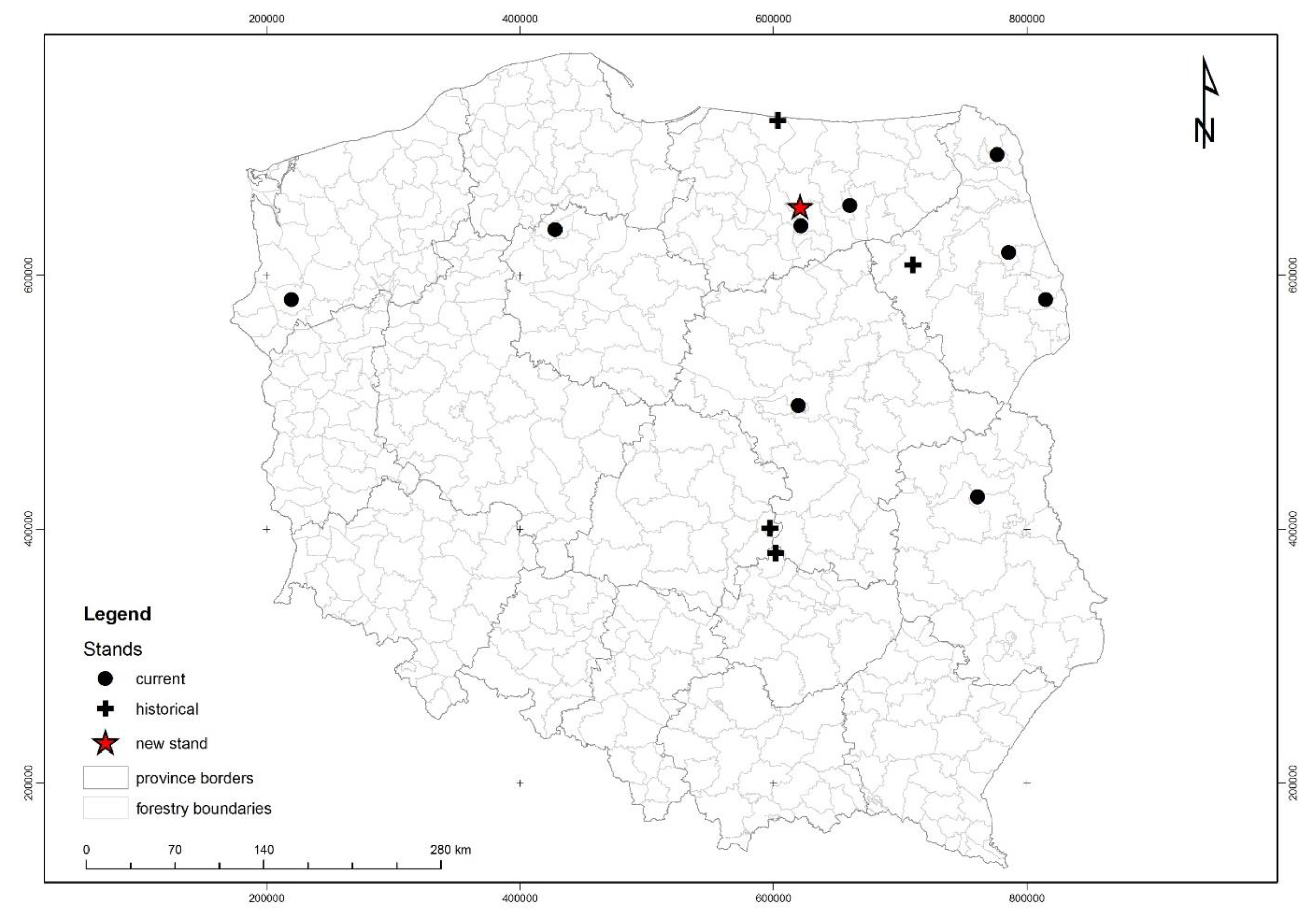
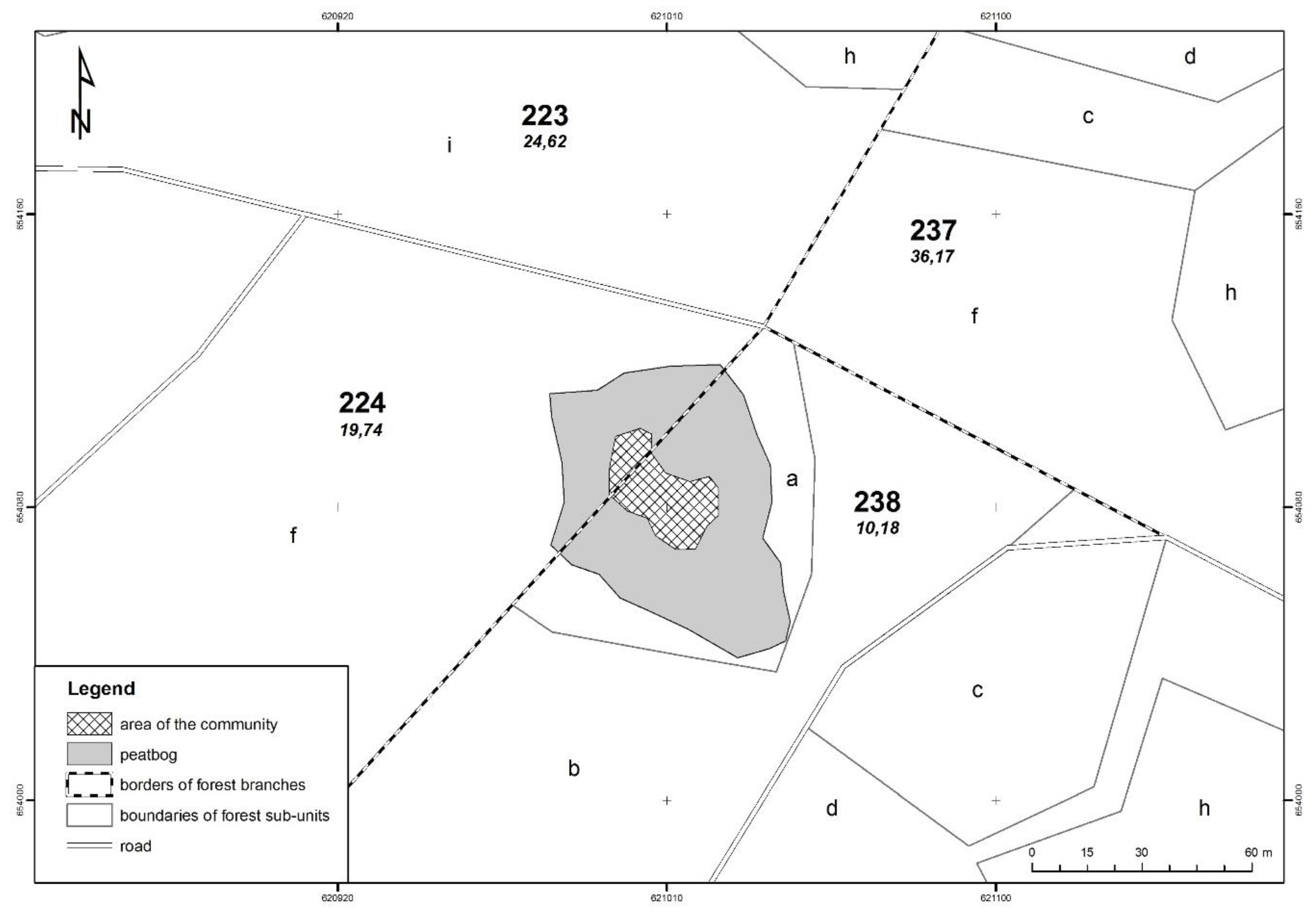
2.1. Study Area
2.2. Morphological Measurements of Leaves and Shoots
2.3. Observations of Flowering, Fruiting, and Seed Production
2.4. Germination Tests after Storage in Various Conditions
2.5. Statistical Analysis
3. Results
3.1. Description of the Vegetative Features and Morphology of Leaves in the Examined Population of C. calyculata
3.2. Description of the Generative Features in the Examined Population of C. calyculata
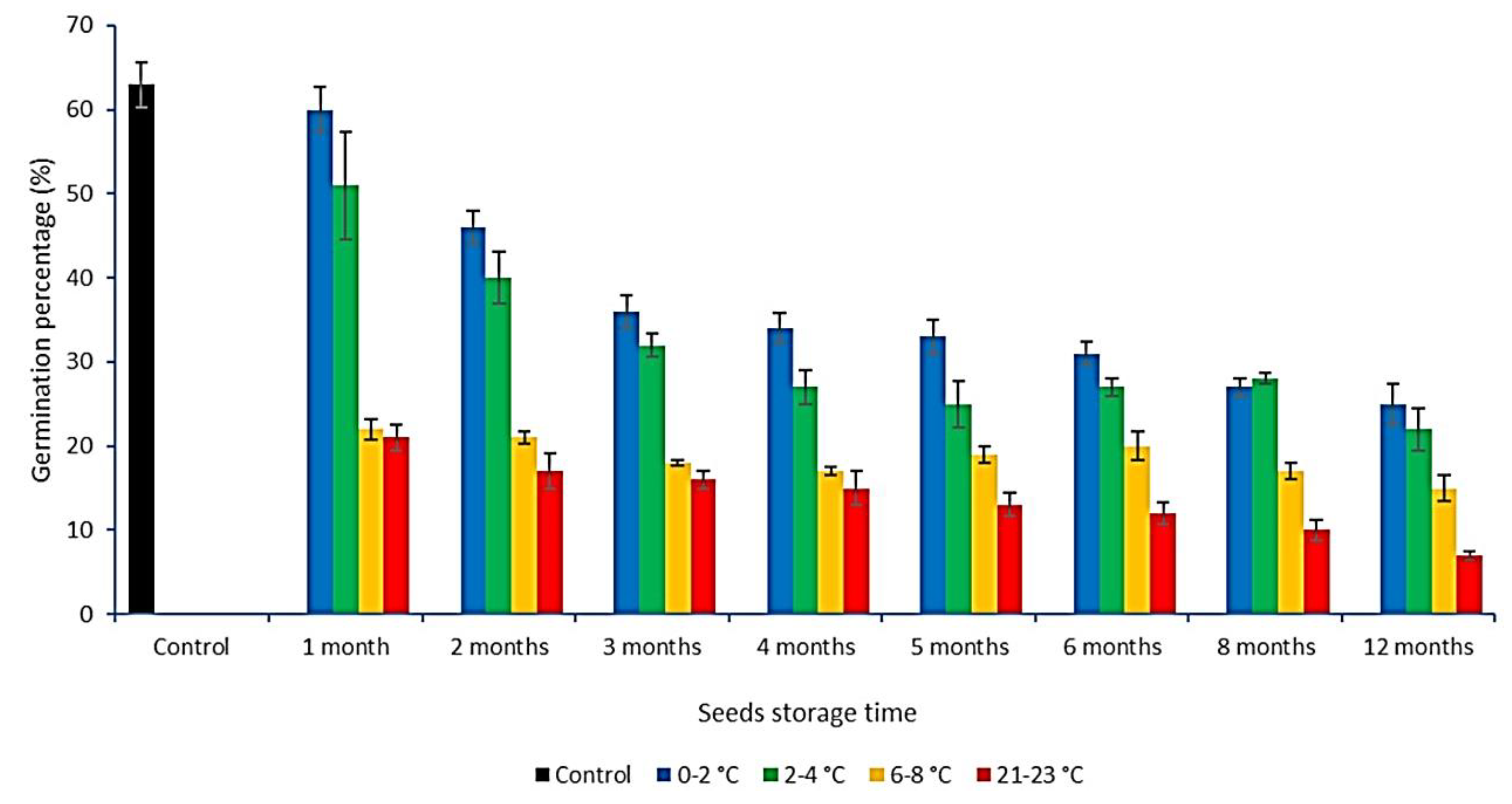
3.3. The Effect of Storage Time and Temperature on Seed Germination Process in the Examined Population of C. calyculata
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kaźmierczakowa, R.; Zarzycki, K.; Mirek, Z. Polish Red Data Book of Plants. Pteridophytes and Flowering Plants, 3rd ed.; Instytut Ochrony Przyrody PAN: Kraków, Poland, 2014. [Google Scholar]
- Kruszelnicki, J. Chamaedaphne calyculata (L.) Moench. In Polish Red Data Book of Plants. Pteridophytes and Flowering Plants, 3rd ed.; Kaźmierczakowa, R., Zarzycki, K., Mirek, Z., Eds.; Instytut Ochrony Przyrody PAN: Kraków, Poland, 2014; pp. 377–379. [Google Scholar]
- Wróblewska, A. Genetic Diversity and Spatial Genetic Structure of Chamaedaphne calyculata (Ericaceae) at the Western Periphery in Relation to its Main Continuous Range in Eurasia. Folia Geobot. 2014, 49, 193–208. [Google Scholar] [CrossRef]
- Wróblewska, A.; Mirski, P. From past to future: Impact of climate change on range shifts and genetic diversity patterns of circumboreal plants. Reg. Environ. Chang. 2018, 18, 409–424. [Google Scholar] [CrossRef]
- Polakowski, B. Chamaedaphne cayculata (L.) Moench in Poland. Zeszyty Naukowe Wyższej Szkoły Rolniczej w Olsztynie 1957, 2, 3–17. [Google Scholar]
- Matuszkiewicz, W. Guide for the Determination of Polish Plant Communities; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2014. [Google Scholar]
- Szkudlarz, P. The morphological and anatomical structure of dry fruits in the family Ericaceae. Biol. Bull. Pozn. 1999, 36, 27–41. [Google Scholar]
- Szkudlarz, P. The morphological and anatomical structure of seeds in the family Ericaceae. Biol. Bull. Pozn. 2001, 38, 113–132. [Google Scholar]
- Malinowska, H.; Janyszek, M.; Szczepaniak, M. Propagation of leather leaf Chamaedaphne calyculata (L.) Moench from seeds and shoot cuttings. Dendrobiology 2004, 52, 33–37. [Google Scholar]
- Densmore, R.V. Effect of day length on germination of seeds collected in Alaska. Am. J. Bot. 1997, 84, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Kloss, M. Plant succession on Chamaedaphne calyculata (L.) Moench mire in the Kampinos Forest. Pol. Ecol. Stud. 1996, 22, 129–139. [Google Scholar]
- Kloss, M. Chamedafne północna ginący gatunek borów bagiennych i mszarów torfowcowych. (Chamaedaphne calyculata, a dying species of bog forests and sphagnous peat bogs). Las Polski 1999, 10, 16–17. (In Polish) [Google Scholar]
- Dziedzic, J.; Dynowski, P.; Źróbek-Sokolnik, A. Chamaedaphne calyculata—A new location in the Warmia region (N-E Poland). Chrońmy Przyr. Ojcz. 2012, 68, 392–395. [Google Scholar]
- Szumarski, W. From laboratory to forest. Natura – Przyroda Warmii i Mazur 2009, 3, 24. [Google Scholar]
- Źróbek-Sokolnik, A.; Ślipiko, A.; Kucewicz, M.; Milewicz, M.; Hołdyński, C. Micropropagation of Chamaedaphne calyculata (L.) Moench by direct shoot organogenesis. Pol. J. Nat. Sci. 2011, 26, 207–215. [Google Scholar]
- Źróbek-Sokolnik, A.; Dynowski, P.; Hołdyński, C. In vitro callus and shoot organogenesis from leaf and stem explants of Chamaedaphne calyculata. Dendrobiology 2014, 72, 85–92. [Google Scholar] [CrossRef]
- Aparicio, A. Seed germination of Erica andevalensis Cabezudo and Rivera (Ericaceae), an endangered edaphic endemic in southwestern Spain. Seed Sci. Technol. 1995, 23, 705–713. [Google Scholar]
- Aparicio, A.; García-Martín, F. The reproductive biology and breeding system of Erica andevalensis Cabezudo & Rivera (Ericaceae), an endangered edaphic endemic of southwestern Spain. Implications for its conservation. Flora 1996, 191, 345–351. [Google Scholar] [CrossRef]
- Cantos, M.; Juana, L.; Garcia, J.L.; Garcia-Linan, M.; Dominguez, M.A.; Troncoso, A. The use of in vitro culture to improve the propagation of Rhododendron ponticum subsp. beaticum. Cent. Eur. J. Biol. 2007, 2, 297–306. [Google Scholar] [CrossRef]
- Moerman, D. Native American Ethnobotany; Timber Press: Portland, OR, USA, 1998. [Google Scholar]
- Shelyuto, V.L.; Glyzin, B.I. Guaiaverin from Chamaedafne calyculata. Chem. Nat. Compd. 1975, 4, 515. [Google Scholar]
- Gorkova, A.S.; Fursa, N.S. Working-out and validation of the method of phenolic glycosides quantitative determination in Chamaedaphne leaves. Bull. Voronezh State Univ. Ser. Chem. Biol. Pharm. 2015, 3, 107–111. [Google Scholar]
- Fursa, N.S.; Buzuk, G.N.; Kuzmichova, N.A.; Gorkova, A.S.; Konovalova, Y.A. Study of phenolic compounds of Chamaedaphne calyculata leaves and flowers by HPLC method. Bull. Pharm. 2016, 2, 51–55. [Google Scholar]
- Klimko, M.; Kosińska, E.; Szkudlarz, P. Morphometry of the leaves of Chamaedaphne calyculata (L.) Moench. Roczniki Akademii Rolniczej w Poznaniu Botanika 2000, 3, 143–154. [Google Scholar]
- Klimko, M.; Szkudlarz, P. Morphological variability of the leaves of Chamaedaphne calyculata (L.) Moench. Ecol. Quest. 2002, 2, 9–15. [Google Scholar]
- Ranal, M.A.; Santana, D.G.; Ferreira, W.R.; Mendes-Rodrigues, C. Calculating germination measurements and organizing spreadsheets. Rev. Brasil. Bot. 2009, 32, 849–855. [Google Scholar] [CrossRef]
- Nasr, S.M.H.; Savadkoohi, S.K.; Ahmadi, E. Effect of different seed treatments on dormancy breaking and germination in three species in arid and semi-arid lands. For. Sci. Pract. 2013, 15, 130–136. [Google Scholar] [CrossRef]
- Tylżanowski, T. Chamedafne północna—Chamaedaphne calyculata in Bory Tucholskie. Chrońmy Przyr. Ojcz. 1975, 31, 51–52. [Google Scholar]
- Reader, R.J. Bog ericad flowers: Self-compatibility and relative attractiveness to bees. Can. J. Bot. 1977, 55, 2279–2287. [Google Scholar] [CrossRef]
- Les, D.H. Aquatic Dicotyledons of North America: Ecology, Life History and Systematics; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2017. [Google Scholar]
- Imbert, E. Ecological consequences and ontogeny of seed heteromorphism. Perspect. Plant Ecol. Evol. Syst. 2002, 5/1, 13–36. [Google Scholar] [CrossRef]
- Deno, N.C. Seed Germination: Theory and Practice, 2nd ed.; University Park, Pennsylvania State University: State College, PA, USA, 1993. [Google Scholar]
- Deno, N.C. First Supplement to the Second Edition of Seed Germination: Theory and Practice; University Park, Pennsylvania State University: State College, PA, USA, 1996. [Google Scholar]
- Deno, N.C. Second Supplement to Seed Germination: Theory and Practice; University Park, Pennsylvania State University: State College, PA, USA, 1998. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press—Elsevier: San Diego, CA, USA, 2014. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average | Min. | Max. | SD | CV | Average CV | ||
|---|---|---|---|---|---|---|---|
| Vegetative features of shoot | Height (cm) | 55.64 | 21.00 | 97.00 | 18.30 | 32.89% | 62.68% |
| Maximum length of increments | 13.13 | 0.00 | 33.50 | 8.03 | 61.14% | ||
| Average length of increments | 9.42 | 0.00 | 19.00 | 4.87 | 51.72% | ||
| Number of branches | 3.36 | 0.00 | 9.00 | 2.18 | 64.97% | ||
| Leaf morphological features | Length of leaf blade | 29.79 | 18.33 | 42.12 | 4.99 | 16.74% | 20.51% |
| Leaf blade width | 10.13 | 6.74 | 12.93 | 1.43 | 14.10% | ||
| Length of petiole | 1.75 | 0.44 | 2.80 | 0.61 | 35.05% | ||
| Leaf blade length-to-width ratio | 2.94 | 2.37 | 3.66 | 0.31 | 10.55% | ||
| Half of the angle at the leaf blade base | 48.72 | 25.27 | 69.27 | 9.32 | 19.14% | ||
| Half of the angle at the leaf blade apex | 61.20 | 29.58 | 88.12 | 12.01 | 19.63% | ||
| Leaf blade area (mm2) | 231.25 | 101.13 | 369.00 | 65.64 | 28.38% | ||
| Generative features of shoot | Number of flower buds | 13.72 | 0.00 | 61.00 | 15.33 | 111.73% | 119.55% |
| Number of flowers | 9.93 | 0.00 | 60.00 | 11.97 | 120.57% | ||
| Number of produced fruits | 4.72 | 0.00 | 25.00 | 5.96 | 126.37% | ||
| Seeds | Number of seeds in fruit | 59.90 | 43.00 | 68 | 8.49 | 15.19% | 15.19% |
| Features of the seed germination process parameters | GP | 26.34 | 6.00 | 70.00 | 13.93 | 52.87% | 37.23% |
| T | 4.69 | 2.00 | 12.00 | 1.77 | 38.00% | ||
| T100 | 14.08 | 8.00 | 21.00 | 2.62 | 18.58% | ||
| MGT | 9.17 | 6.12 | 15.43 | 1.81 | 19.79% | ||
| MR | 0.11 | 0.06 | 0.16 | 0.02 | 18.57% | ||
| GI | 2.40 | 0.50 | 7.34 | 1.41 | 58.79% | ||
| GRI | 3.53 | 0.45 | 7.76 | 1.90 | 54.00% |
| Year | Total Number of Flower Buds | Min. and Max. Number of Flower Buds on a Single Plant | Average Number of flower Buds on a Single Plant | Total Number of Formed Flowers | Min. and Max. Number of Formed Flowers on a Single Plant | Average Number of Formed Flowers on a Single Plant |
|---|---|---|---|---|---|---|
| 2013 | 669 | 0–61 | 13 ± 1.98 | 449 | 0–34 | 9 ± 1.37 |
| 2014 | 703 | 0–61 | 14 ± 2.39 | 544 | 0–60 | 11 ± 2.02 |
| Average for total | 686 | 0–61 | 14 ± 1.53 | 496 | 0–47 | 10 ± 1.20 |
| Year | Total Number of Formed Fruits | Min. and Max. Number of Formed Fruits on a Single Plant | Average Number of Formed Fruits on a Single Plant |
|---|---|---|---|
| 2013 | 203 | 0–17 | 4 ± 0.70 |
| 2014 | 269 | 0–25 | 5 ± 0.98 |
| Average for total | 236 | 0–21 | 5 ± 0.60 |
| Seeds Storage Temp. (°C) | Seeds Storage Time (months) | T (day) | T100 (day) | MGT (day) | MR (day −1) | GI | GRI (% day −1) |
|---|---|---|---|---|---|---|---|
| control | 0 | 3 ± 0.24 | 15 ± 0.40 | 9.40 ± 0.31 | 0.11 ± 0.00 | 5.96 ± 0.40 | 7.40 ± 0.11 |
| 0–2 | |||||||
| 1 | 3 ± 0.24ab | 16 ± 0.60a | 9.75 ± 0.28a | 0.10 ± 0.01ab | 5.87 ± 0.40a | 7.40 ± 0.11a | |
| 2 | 3 ± 0.24ab | 15 ± 0.49a | 10.23 ± 0.55a | 0.10 ± 0.01a | 4.69 ± 0.43a | 5.64 ± 0.22bc | |
| 3 | 3 ± 0.40a | 14 ± 0.95a | 7.73 ± 0.48b | 0.13 ± 0.01bc | 2.78 ± 0.22b | 6.26 ± 0.61ab | |
| 4 | 3 ± 0.24ab | 13 ± 1.32a | 7.74 ± 0.58b | 0.13 ± 0.01c | 2.66 ± 0.31b | 5.40 ± 0.32bcd | |
| 5 | 4 ± 0.20ab | 14 ± 0.97a | 8.38 ± 0.40ab | 0.12 ± 0.01abc | 2.80 ± 0.28b | 4.73 ± 0.09cde | |
| 6 | 4 ± 0.24ab | 12 ± 1.58a | 7.39 ± 0.19b | 0.14 ± 0.00c | 2.27 ± 0.10b | 4.91 ± 0.20bcde | |
| 8 | 4 ± 0.37b | 12 ± 0.92a | 7.49 ± 0.13b | 0.13 ± 0.00c | 2.04 ± 0.10b | 4.14 ± 0.18de | |
| 12 | 4 ± 0.32ab | 13 ± 1.24a | 7.78 ± 0.50b | 0.13 ± 0.01bc | 1.94 ± 0.27b | 3.81 ± 0.42e | |
| 2–4 | |||||||
| 1 | 4 ± 0.49ab | 15 ± 1.60a | 9.61 ± 0.73a | 0.11 ± 0.01a | 5.05 ± 0.84a | 6.19 ± 0.59a | |
| 2 | 4 ± 0.24ab | 14 ± 1.30a | 9.56 ± 0.47a | 0.11 ± 0.01a | 3.91 ± 0.43ab | 5.13 ± 0.41ab | |
| 3 | 3 ± 0.40a | 15 ± 0.63a | 8.64 ± 0.40a | 0.12 ± 0.01a | 2.77 ± 0.19bc | 4.65 ± 0.38abc | |
| 4 | 4 ± 0.24ab | 15 ± 0.86a | 8.02 ± 0.47a | 0.13 ± 0.01a | 2.18 ± 0.29bc | 4.14 ± 0.33bc | |
| 5 | 4 ± 0.32ab | 14 ± 1.40a | 8.27 ± 0.56a | 0.12 ± 0.01a | 2.09 ± 0.28bc | 3.67 ± 0.50bc | |
| 6 | 4 ± 0.93ab | 13 ± 1.29a | 8.35 ± 0.91a | 0.12 ± 0.01a | 2.28 ± 0.28bc | 4.00 ± 0.42bc | |
| 8 | 4 ± 0.32ab | 12 ± 1.03a | 7.46 ± 0.15a | 0.13 ± 0.00a | 2.09 ± 0.06bc | 4.27 ± 0.09bc | |
| 12 | 5 ± 0.86b | 13 ± 1.05a | 8.48 ± 0.69a | 0.12 ± 0.01a | 1.95 ± 0.34c | 3.09 ± 0.39c | |
| 6–8 | |||||||
| 1 | 5 ± 0.20a | 16 ± 1.20a | 10.19 ± 0.88a | 0.10 ± 0.01a | 2.21 ± 0.24a | 2.52 ± 0.25ab | |
| 2 | 5 ± 0.86a | 15 ± 0.60a | 10.15 ± 0.48a | 0.10 ± 0.01a | 2.14 ± 0.09a | 2.48 ± 0.26ab | |
| 3 | 5 ± 0.37a | 13 ± 0.87a | 8.54 ± 0.27a | 0.12 ± 0.00a | 1.57 ± 0.06ab | 2.46 ± 0.14ab | |
| 4 | 4 ± 0.37a | 14 ± 1.16a | 8.28 ± 0.45a | 0.12 ± 0.01a | 1.43 ± 0.10b | 2.55 ± 0.19ab | |
| 5 | 4 ± 0.24a | 14 ± 1.10a | 8.84 ± 0.56a | 0.11 ± 0.01a | 1.71 ± 0.17ab | 2.55 ± 0.18ab | |
| 6 | 4 ± 0.55a | 12 ± 1.07a | 7.71 ± 0.33a | 0.13 ± 0.01a | 1.54 ± 0.15ab | 3.17 ± 0.36b | |
| 8 | 5 ± 0.37a | 12 ± 0.86a | 8.34 ± 0.35a | 0.12 ± 0.00a | 1.43 ± 0.09b | 2.32 ± 0.17ab | |
| 12 | 5 ± 0.73a | 13 ± 0.95a | 8.86 ± 0.35a | 0.12 ± 0.01a | 1.34 ± 0.18b | 1.99 ± 0.24a | |
| 21–23 | |||||||
| 1 | 7 ± 0.55a | 17 ± 0.20a | 11.35 ± 0.36a | 0.09 ± 0.00a | 2.35 ± 0.14a | 2.07 ± 0.24a | |
| 2 | 8 ± 1.20a | 15 ± 0.37a | 11.12 ± 0.62a | 0.09 ± 0.00a | 1.87 ± 0.17ab | 1.73 ± 0.32ab | |
| 3 | 6 ± 0.37a | 14 ± 0.95a | 9.80 ± 0.82a | 0.10 ± 0.01a | 1.57 ± 0.16abc | 1.88 ± 0.22ab | |
| 4 | 6 ± 0.75a | 16 ± 1.61a | 10.93 ± 1.02a | 0.09 ± 0.01a | 1.60 ± 0.25abc | 1.58 ± 0.30abc | |
| 5 | 7 ± 0.73a | 16 ± 1.17a | 11.92 ± 0.99a | 0.09 ± 0.01a | 1.54 ± 0.21abc | 1.19 ± 0.18abc | |
| 6 | 6 ± 0.75a | 14 ± 1.96a | 10.40 ± 1.30a | 0.10 ± 0.01a | 1.35 ± 0.29bc | 1.31 ± 0.09abc | |
| 8 | 6 ± 0.51a | 13 ± 1.46a | 10.19 ± 0.90a | 0.10 ± 0.01a | 1.09 ± 0.20bc | 1.10 ± 0.10bc | |
| 12 | 7 ± 1.41a | 14 ± 1.24a | 11.42 ± 0.96a | 0.09 ± 0.01a | 0.78 ± 0.10c | 0.63 ± 0.05c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Źróbek-Sokolnik, A.; Dynowski, P.; Fenyk, M.A. The Morphological Features and Biology of a Relict and Endangered Woody Plant Species: Chamaedaphne calyculata (L.) Moench (Ericaceae). Plants 2019, 8, 129. https://doi.org/10.3390/plants8050129
Źróbek-Sokolnik A, Dynowski P, Fenyk MA. The Morphological Features and Biology of a Relict and Endangered Woody Plant Species: Chamaedaphne calyculata (L.) Moench (Ericaceae). Plants. 2019; 8(5):129. https://doi.org/10.3390/plants8050129
Chicago/Turabian StyleŹróbek-Sokolnik, Anna, Piotr Dynowski, and Mieczysława Aldona Fenyk. 2019. "The Morphological Features and Biology of a Relict and Endangered Woody Plant Species: Chamaedaphne calyculata (L.) Moench (Ericaceae)" Plants 8, no. 5: 129. https://doi.org/10.3390/plants8050129