HER2-Specific Pseudomonas Exotoxin A PE25 Based Fusions: Influence of Targeting Domain on Target Binding, Toxicity, and In Vivo Biodistribution

 ,
,  , , ,
, , ,  , ,
, ,
Abstract
:
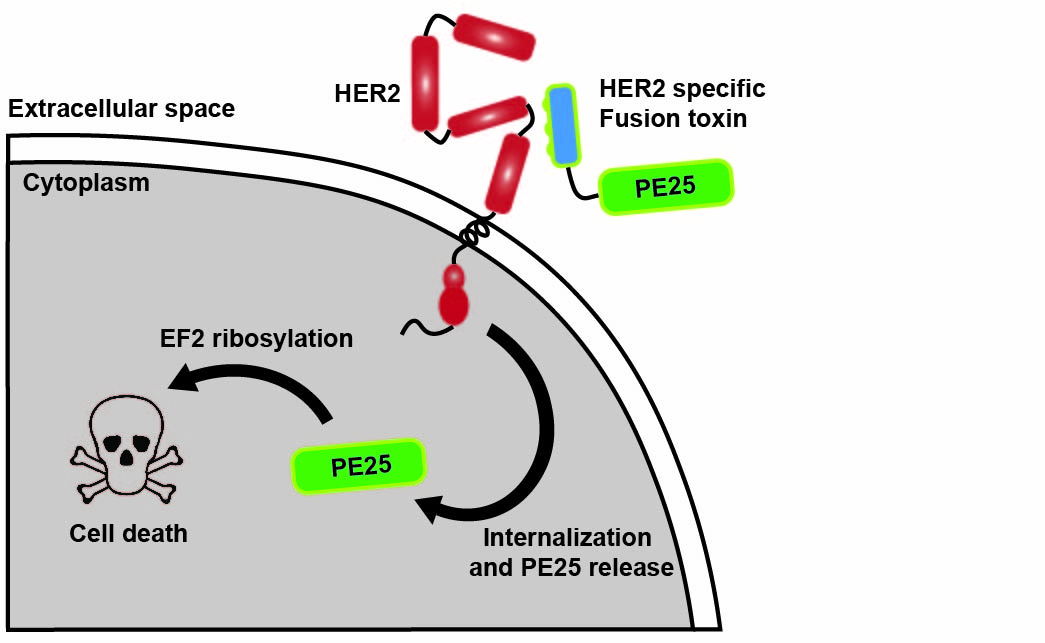
1. Introduction
2. Materials and Methods
2.1. General
2.2. Gene Construction
2.3. Protein Expression and Purification
2.4. Biosensor Analysis
2.5. Cell Culture
2.6. Flow Cytometry Analysis
2.7. Radiolabeling with 99mTc and 111In
2.8. Binding Studies Using a Ligandtracer Instrument
2.9. Cell Binding Specificity Test
2.10. In Vitro Cytotoxicity Analysis
2.11. Biodistribution Experiments
3. Results
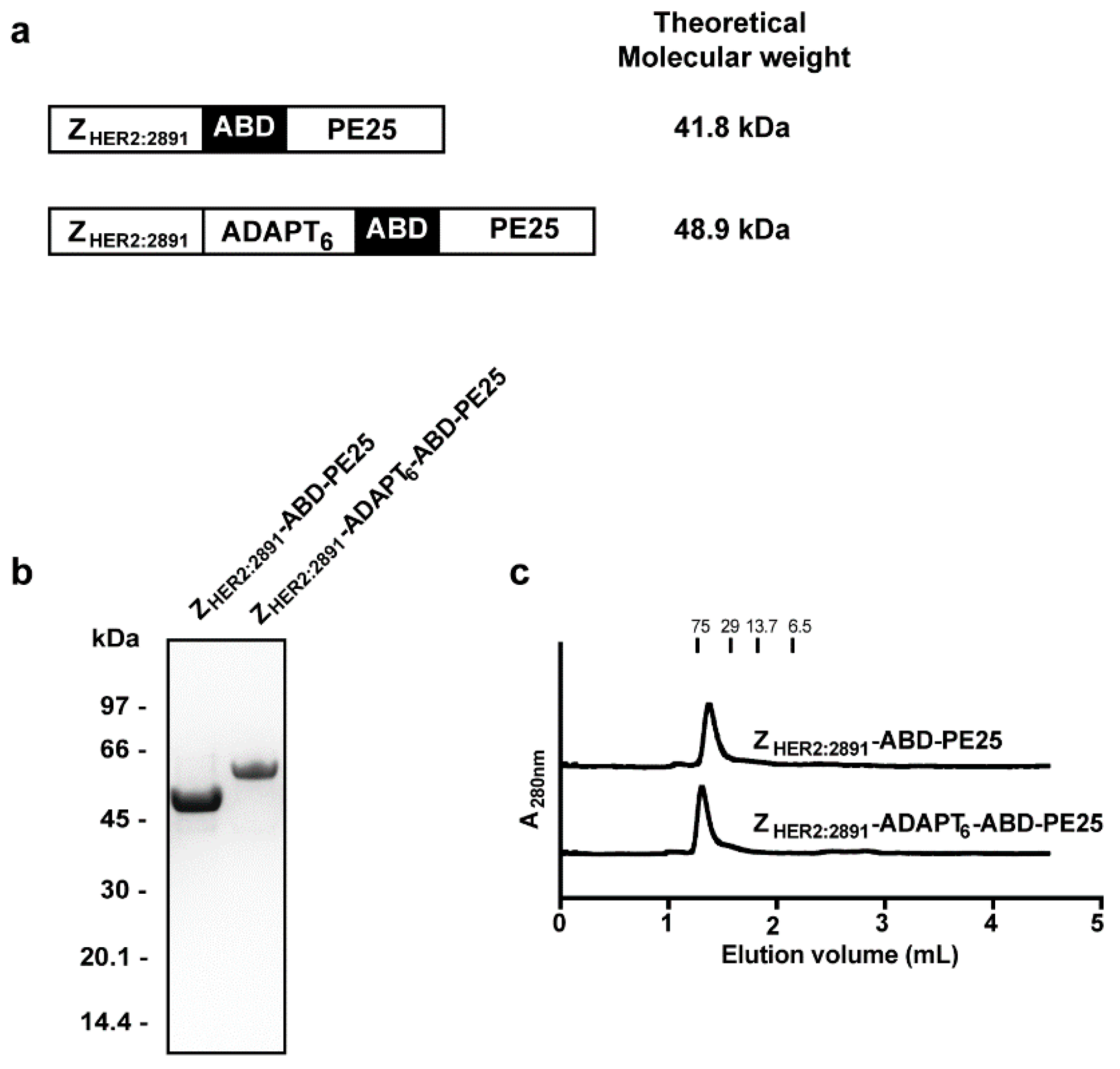
3.1. Construct Design, Protein Expression, and Purification
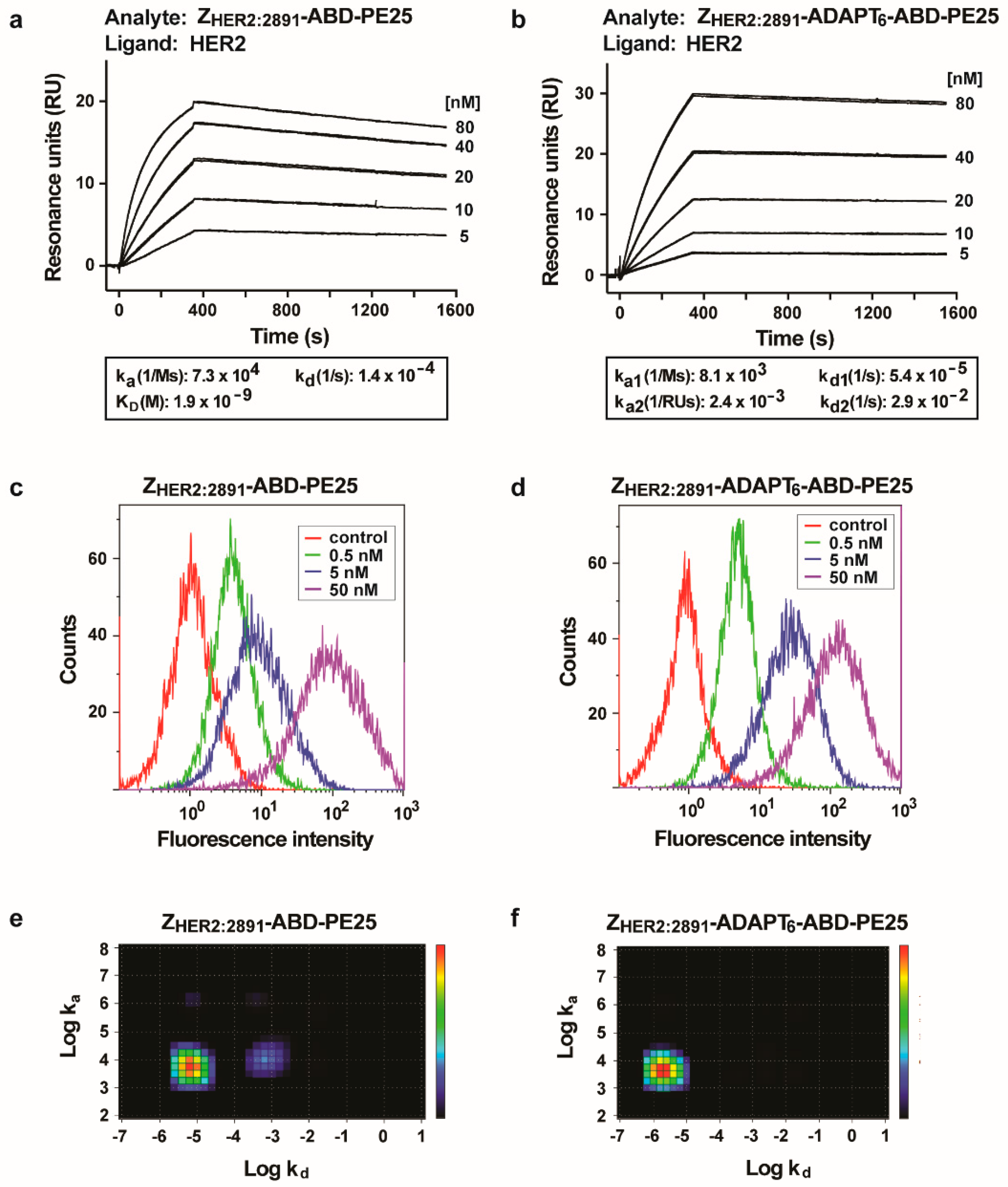
3.2. Interaction of the Fusion Toxins with HER2
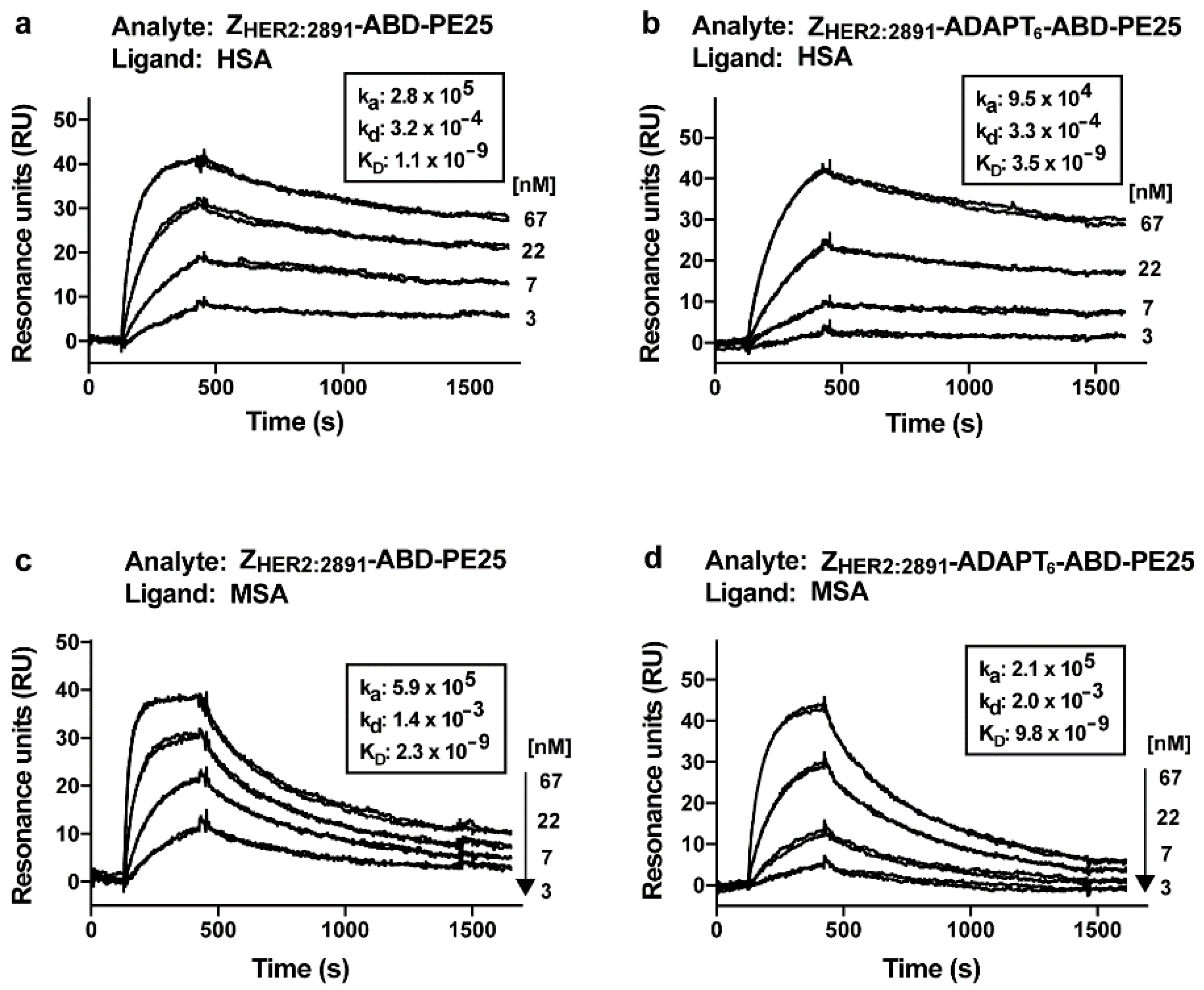
3.3. Analysis of Affinity to Serum Albumins
3.4. Cytotoxicity Analysis
3.5. Biodistribution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Emde, A.; Köstler, W.J.; Yarden, Y. Therapeutic strategies and mechanisms of tumorigenesis of HER2-overexpressing breast cancer. Crit. Rev. Oncol. Hematol. 2012, 84, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Schwaederle, M.; Arguello, D.; Millis, S.Z.; Gatalica, Z.; Kurzrock, R. HER2 expression status in diverse cancers: Review of results from 37,992 patients. Cancer Metastasis Rev. 2015, 34, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Citri, A.; Yarden, Y. EGF-ERBB signalling: Towards the systems level. Nat. Rev. Mol. Cell Biol. 2006, 7, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human breast cancer: Correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.L.; Cobleigh, M.A.; Tripathy, D.; Gutheil, J.C.; Harris, L.N.; Fehrenbacher, L.; Slamon, D.J.; Murphy, M.; Novotny, W.F.; Burchmore, M.; et al. Efficacy and Safety of Trastuzumab as a Single Agent in First-Lin Treatment of HER2-Overexpressing Metastatic Breast Cancer. J. Clin. Oncol. 2003, 20, 719–726. [Google Scholar] [CrossRef] [PubMed]
- von Minckwitz, G.; Procter, M.; de Azambuja, E.; Zardavas, D.; Benyunes, M.; Viale, G.; Suter, T.; Arahmani, A.; Rouchet, N.; Clark, E.; et al. Adjuvant Pertuzumab and Trastuzumab in Early HER2-Positive Breast Cancer. N. Engl. J. Med. 2017, 377, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Miles, D.; Gianni, L.; Krop, I.E.; Welslau, M.; Baselga, J.; Pegram, M.; Oh, D.-Y.; Diéras, V.; Guardino, E.; et al. Trastuzumab emtansine for HER2-positive advanced breast cancer. N. Engl. J. Med. 2012, 367, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keam, S.J. Trastuzumab Deruxtecan: First Approval. Drugs 2020, 80, 501–508. [Google Scholar] [CrossRef]
- Mazor, R.; Onda, M.; Pastan, I. Immunogenicity of therapeutic recombinant immunotoxins. Immunol. Rev. 2016, 270, 152–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreitman, R.J.; Dearden, C.; Zinzani, P.L.; Delgado, J.; Karlin, L.; Robak, T.; Gladstone, D.E.; le Coutre, P.; Dietrich, S.; Gotic, M.; et al. Moxetumomab pasudotox in relapsed/refractory hairy cell leukemia. Leukemia 2018, 32, 1768–1777. [Google Scholar] [CrossRef]
- Liu, W.; Onda, M.; Lee, B.; Kreitman, R.J.; Hassan, R.; Xiang, L.; Pastan, I. Recombinant immunotoxin engineered for low immunogenicity and antigenicity by identifying and silencing human B-cell epitopes. Proc. Natl. Acad. Sci. USA 2012, 109, 11782–11787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onda, M.; Beers, R.; Xiang, L.; Nagata, S.; Wang, Q.-C.; Pastan, I. An immunotoxin with greatly reduced immunogenicity by identification and removal of B cell epitopes. Proc. Natl. Acad. Sci. USA 2008, 105, 11311–11316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onda, M.; Beers, R.; Xiang, L.; Lee, B.; Weldon, J.E.; Kreitman, R.J.; Pastan, I. Recombinant immunotoxin against B-cell malignancies with no immunogenicity in mice by removal of B-cell epitopes. Proc. Natl. Acad. Sci. USA 2011, 108, 5742–5747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazor, R.; Vassall, A.N.; Eberle, J.A.; Beers, R.; Weldon, J.E.; Venzon, D.J.; Tsang, K.Y.; Benhar, I.; Pastan, I. Identification and elimination of an immunodominant T-cell epitope in recombinant immunotoxins based on Pseudomonas exotoxin A. Proc. Natl. Acad. Sci. USA 2012, 109, E3597–E3603. [Google Scholar] [CrossRef] [Green Version]
- Weldon, J.E.; Skarzynski, M.; Therres, J.A.; Ostovitz, J.R.; Zhou, H.; Kreitman, R.J.; Pastan, I. Designing the Furin-Cleavable Linker in Recombinant Immunotoxins Based on Pseudomonas Exotoxin, A. Bioconjug. Chem. 2015, 26, 1120–1128. [Google Scholar] [CrossRef]
- Mazor, R.; Onda, M.; Park, D.; Addissie, S.; Xiang, L. Dual B- and T-cell de-immunization of recombinant immunotoxin targeting mesothelin with high cytotoxic activity. Oncotarget 2016, 7, 29916–29926. [Google Scholar] [CrossRef] [Green Version]
- Bera, T.K.; Onda, M.; Kreitman, R.J.; Pastan, I. An improved recombinant Fab-immunotoxin targeting CD22 expressing malignancies. Leuk. Res. 2014, 38, 1224–1229. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.; Nordberg, E.; Höidén-Guthenberg, I.; Brismar, H.; Adams, G.P.; Nilsson, F.Y.; Carlsson, J.; Ståhl, S. Phage display selection of Affibody molecules with specific binding to the extracellular domain of the epidermal growth factor receptor. Protein Eng. Des. Sel. 2007, 20, 189–199. [Google Scholar] [CrossRef]
- Orlova, A.; Magnusson, M.; Eriksson, T.L.J.; Nilsson, M.; Larsson, B.; Höiden-Guthenberg, I.; Widström, C.; Carlsson, J.; Tolmachev, V.; Ståhl, S.; et al. Tumor imaging using a picomolar affinity HER2 binding Affibody molecule. Cancer Res. 2006, 66, 4339–4348. [Google Scholar] [CrossRef] [Green Version]
- Kronqvist, N.; Malm, M.; Göstring, L.; Gunneriusson, E.; Nilsson, M.; Höidén-Guthenberg, I.; Gedda, L.; Frejd, F.Y.; Sthl, S.; Löfblom, J. Combining phage and staphylococcal surface display for generation of ErbB3-specific Affibody molecules. Protein Eng. Des. Sel. 2011, 24, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lundberg, E.; Vernet, E.; Larsson, B.; Höiden-Guthenberg, I.; Gräslund, T. Selection of affibody molecules to the ligand-binding site of the insulin-like growth factor-1 receptor. Biotechnol. Appl. Biochem. 2010, 55, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Honarvar, H.; Garousi, J.; Gunneriusson, E.; Höidén-Guthenberg, I.; Altai, M.; Widström, C.; Tolmachev, V.; Frejd, F.Y. Imaging of CAIX-expressing xenografts in vivo using 99mTc-HEHEHE-ZCAIX:1 Affibody molecule. Int. J. Oncol. 2015, 46, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Ståhl, S.; Gräslund, T.; Eriksson Karlström, A.; Frejd, F.Y.; Nygren, P.Å.; Löfblom, J. Affibody Molecules in Biotechnological and Medical Applications. Trends Biotechnol. 2017, 35, 691–712. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Seijsing, J.; Frejd, F.; Tolmachev, V.; Gräslund, T. Target-specific cytotoxic effects on HER2-expressing cells by the tripartite fusion toxin ZHER2:2891-ABD-PE38X8, including a targeting affibody molecule and a half-life extension domain. Int. J. Oncol. 2015, 2, 601–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sörensen, J.; Velikyan, I.; Sandberg, D.; Wennborg, A.; Feldwisch, J.; Tolmachev, V.; Orlova, A.; Sandström, M.; Lubberink, M.; Olofsson, H.; et al. Measuring HER2-receptor expression in metastatic breast cancer using [68Ga]ABY-025 Affibody PET/CT. Theranostics 2016, 6, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Nilvebrant, J.; Åstrand, M.; Georgieva-Kotseva, M.; Björnmalm, M.; Löfblom, J.; Hober, S. Engineering of bispecific affinity proteins with high affinity for ERBB2 and adaptable binding to albumin. PLoS ONE 2014, 9, e103094. [Google Scholar] [CrossRef]
- Nilvebrant, J.; Åstrand, M.; Löfblom, J.; Hober, S. Development and characterization of small bispecific albumin-binding domains with high affinity for ErbB3. Cell. Mol. Life Sci. 2013, 70, 3973–3985. [Google Scholar] [CrossRef]
- Garousi, J.; Lindbo, S.; Nilvebrant, J.; Astrand, M.; Buijs, J.; Sandström, M.; Honarvar, H.; Orlova, A.; Tolmachev, V.; Hober, S. ADAPT, a novel scaffold protein-based probe for radionuclide imaging of molecular targets that are expressed in disseminated cancers. Cancer Res. 2015, 75, 4364–4371. [Google Scholar] [CrossRef] [Green Version]
- Garousi, J.; Lindbo, S.; Borin, J.; von Witting, E.; Vorobyeva, A.; Oroujeni, M.; Mitran, B.; Orlova, A.; Buijs, J.; Tolmachev, V.; et al. Comparative evaluation of dimeric and monomeric forms of ADAPT scaffold protein for targeting of HER2-expressing tumours. Eur. J. Pharm. Biopharm. 2019, 134, 37–48. [Google Scholar] [CrossRef]
- Tolmachev, V.; Bragina, O.; von Witting, E.; Garousi, J.; Zelchan, R.; Sinilkin, I.; Medvedeva, A.; Doroshenko, A.; Vorobyeva, A.; Lindbo, S.; et al. First-in-humans Evaluation of [99mTc]-ADAPT6, a Novel Scaffold Protein for Visualization of HER2 Expression. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, S166. [Google Scholar]
- Liu, H.; Lindbo, S.; Ding, H.; Altai, M.; Garousi, J.; Orlova, A.; Tolmachev, V.; Hober, S.; Gräslund, T. Potent and specific fusion toxins consisting of a HER2--binding, ABD--derived affinity protein, fused to truncated versions of Pseudomonas exotoxin A. Int. J. Oncol. 2019, 55, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Tolmachev, V.; Orlova, A.; Pehrson, R.; Galli, J.; Baastrup, B.; Andersson, K.; Sandström, M.; Rosik, D.; Carlsson, J.; Lundqvist, H.; et al. Radionuclide therapy of HER2-positive microxenografts using a 177Lu-labeled HER2-specific Affibody molecule. Cancer Res. 2007, 67, 2773–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roopenian, D.C.; Akilesh, S. FcRn: The neonatal Fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, A.; Dogan, J.; Herne, N.; Abrahmsén, L.; Nygren, P.Å. Engineering of a femtomolar affinity binding protein to human serum albumin. Protein Eng. Des. Sel. 2008, 21, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Altai, M.; Liu, H.; Ding, H.; Mitran, B.; Edqvist, P.-H.; Tolmachev, V.; Orlova, A.; Gräslund, T. Affibody-derived drug conjugates: Potent cytotoxic molecules for treatment of HER2 over-expressing tumors. J. Control. Release 2018, 288, 84–95. [Google Scholar] [CrossRef]
- Björke, H.; Andersson, K. Measuring the affinity of a radioligand with its receptor using a rotating cell dish with in situ reference area. Appl. Radiat. Isot. 2006, 64, 32–37. [Google Scholar] [CrossRef]
- Pastan, I.; Beers, R.; Bera, T.K. Recombinant Immunotoxins in the Treatment of Cancer. Methods Mol. Biol. 2004, 248, 503–518. [Google Scholar]
- Feldwisch, J.; Tolmachev, V.; Lendel, C.; Herne, N.; Sjöberg, A.; Larsson, B.; Rosik, D.; Lindqvist, E.; Fant, G.; Höidén-Guthenberg, I.; et al. Design of an optimized scaffold for affibody molecules. J. Mol. Biol. 2010, 398, 232–247. [Google Scholar] [CrossRef]
- Ekerljung, L.; Lindborg, M.; Gedda, L.; Frejd, F.Y.; Carlsson, J.; Lennartsson, J. Dimeric HER2-specific affibody molecules inhibit proliferation of the SKBR-3 breast cancer cell line. Biochem. Biophys. Res. Commun. 2008, 377, 489–494. [Google Scholar] [CrossRef]
- Ekerljung, L.; Steffen, A.C.; Carlsson, J.; Lennartsson, J. Effects of HER2-binding affibody molecules on intracellular signaling pathways. Tumor Biol. 2006, 27, 201–210. [Google Scholar] [CrossRef]
- Tamaskovic, R.; Schwill, M.; Nagy-Davidescu, G.; Jost, C.; Schaefer, D.C.; Verdurmen, W.P.R.; Schaefer, J.V.; Honegger, A.; Plückthun, A. Intermolecular biparatopic trapping of ErbB2 prevents compensatory activation of PI3K/AKT via RAS-p110 crosstalk. Nat. Commun. 2016, 7, 11672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaghy, H. Effects of antibody, drug and linker on the preclinical and clinical toxicities of antibody-drug conjugates. MAbs 2016, 8, 659–671. [Google Scholar] [CrossRef]
- Zielinski, R.; Lyakhov, I.; Hassan, M.; Kuban, M.; Shafer-Weaver, K.; Gandjbakhche, A.; Capala, J. HER2-affitoxin: A potent therapeutic agent for the treatment of HER2-overexpressing tumors. Clin. Cancer Res. 2011, 17, 5071–5081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.; Cao, L.; Guo, W.; Liu, H.; Xu, H.; Fang, Q.; Hong, Z. HER2-targeted immunotoxins with low nonspecific toxicity and immunogenicity. Biochem. Biophys. Res. Commun. 2016, 475, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Lewis Phillips, G.D.; Li, G.; Dugger, D.L.; Crocker, L.M.; Parsons, K.L.; Mai, E.; Blättler, W.A.; Lambert, J.M.; Chari, R.V.J.; Lutz, R.J.; et al. Targeting HER2-positive breast cancer with trastuzumab-DM1, an antibody-cytotoxic drug conjugate. Cancer Res. 2008, 68, 9280–9290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eigenbrot, C.; Ultsch, M.; Dubnovitsky, A.; Abrahmsen, L.; Hard, T. Structural basis for high-affinity HER2 receptor binding by an engineered protein. Proc. Natl. Acad. Sci. USA 2010, 107, 15039–15044. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ZHER2:2891-ABD-PE25 | ADAPT6-ABD-PE25 e | ZHER2:2891-ADAPT6-ABD-PE25 | |
|---|---|---|---|
| At 4 h post-injection | |||
| Blood | 9.2 ± 1.0 c,d | 3.2 ± 0.1 | 4.6 ± 0.9 |
| Heart | 3.0 ± 0.5 c,d | 1.1 ± 0.2 | 1.7 ± 0.2 |
| Lung | 4.0 ± 0.5 c,d | 1.3 ± 0.1 d | 2.2 ± 0.5 |
| Salivary gland | 1.6 ± 0.4 c,d | 0.7 ± 0.2 | 0.8 ± 0.3 |
| Liver | 13.9 ± 2.2 c | 42.4 ± 6.8 d | 23.1 ± 3.8 |
| Spleen | 6.2 ± 2.0 c | 18.9 ± 4.2 d | 7.8 ± 2.0 |
| Pancreas | 0.9 ± 0.1 | 0.4 ± 0.1 | 1.0 ± 0.5 |
| Stomach | 1.0 ± 0.1 c | 0.5 ± 0.2 | 0.7 ± 0.1 |
| Kidney | 40.1 ± 4.8 c,d | 12.5 ± 0.6 d | 69.7 ± 8.0 |
| Colon | 1.3 ± 0.1 c | 0.6 ± 0.3 | 1.0 ± 0.3 |
| Skin | 1.5 ± 0.5 c | 0.6 ± 0.1 | 0.9 ± 0.2 |
| Muscle | 0.7 ± 0.1 c,d | 0.3 ± 0.1 | 0.4 ± 0.1 |
| Bone | 1.4 ± 0.1 d | 1.1 ± 0.3 | 0.9 ± 0.2 |
| GI tract b | 3.5 ± 0.5 | 6.9 ± 4.4 | 3.3 ± 1.1 |
| Carcass b | 17.8 ± 2.1 c,d | 3.5 ± 2.8 d | 13.0 ± 0.8 |
| At 24 h post-injection | |||
| Blood | 2.6 ± 0.5 d | 2.0 ± 0.1 | 1.8 ± 0.3 |
| Heart | 1.6 ± 0.3 | 1.8 ± 0.2 d | 1.2 ± 0.1 |
| Lung | 1.8 ± 0.3 d | 1.7 ± 0.2 d | 1.2 ± 0.1 |
| Sal gland | 1.2 ± 0.3 d | 1.2 ± 0.1 d | 0.8 ± 0.1 |
| Liver | 8.7 ± 0.9 c,d | 21.9 ± 2.0 | 16.2 ± 1.7 |
| Spleen | 5.5 ± 1.1 c | 8.5 ± 1.7 d | 5.3 ± 1.3 |
| Pancreas | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.5 ± 0.1 |
| Stomach | 0.8 ± 0.1 | 0.7 ± 0.1 | 0.6 ± 0.1 |
| Kidney | 33.4 ± 4.4 c,d | 9.3 ± 1.3 d | 47.1 ± 5.2 |
| Colon | 0.5 ± 0.1 c | 0.9 ± 0.1 d | 0.5 ± 0.1 |
| Skin | 2.0 ± 0.4 c,d | 1.2 ± 0.1 | 1.3 ± 0.1 |
| Muscle | 0.5 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.0 |
| Bone | 1.6 ± 0.5 | 1.8 ± 0.3 | 0.9 ± 0.2 |
| GI tract b | 1.5 ± 0.1 c | 2.7 ± 0.7 d | 1.0 ± 0.1 |
| Carcass b | 16.4 ± 1.5 c,d | 13.3 ± 1.4 | 11.8 ± 1.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, H.; Altai, M.; Yin, W.; Lindbo, S.; Liu, H.; Garousi, J.; Xu, T.; Orlova, A.; Tolmachev, V.; Hober, S.; et al. HER2-Specific Pseudomonas Exotoxin A PE25 Based Fusions: Influence of Targeting Domain on Target Binding, Toxicity, and In Vivo Biodistribution. Pharmaceutics 2020, 12, 391. https://doi.org/10.3390/pharmaceutics12040391
Ding H, Altai M, Yin W, Lindbo S, Liu H, Garousi J, Xu T, Orlova A, Tolmachev V, Hober S, et al. HER2-Specific Pseudomonas Exotoxin A PE25 Based Fusions: Influence of Targeting Domain on Target Binding, Toxicity, and In Vivo Biodistribution. Pharmaceutics. 2020; 12(4):391. https://doi.org/10.3390/pharmaceutics12040391
Chicago/Turabian StyleDing, Haozhong, Mohamed Altai, Wen Yin, Sarah Lindbo, Hao Liu, Javad Garousi, Tianqi Xu, Anna Orlova, Vladimir Tolmachev, Sophia Hober, and et al. 2020. "HER2-Specific Pseudomonas Exotoxin A PE25 Based Fusions: Influence of Targeting Domain on Target Binding, Toxicity, and In Vivo Biodistribution" Pharmaceutics 12, no. 4: 391. https://doi.org/10.3390/pharmaceutics12040391