Adverse Effects of Classical Swine Fever Virus LOM Vaccine and Jeju LOM Strains in Pregnant Sows and Specific Pathogen-Free Pigs

Abstract
:1. Introduction
2. Results
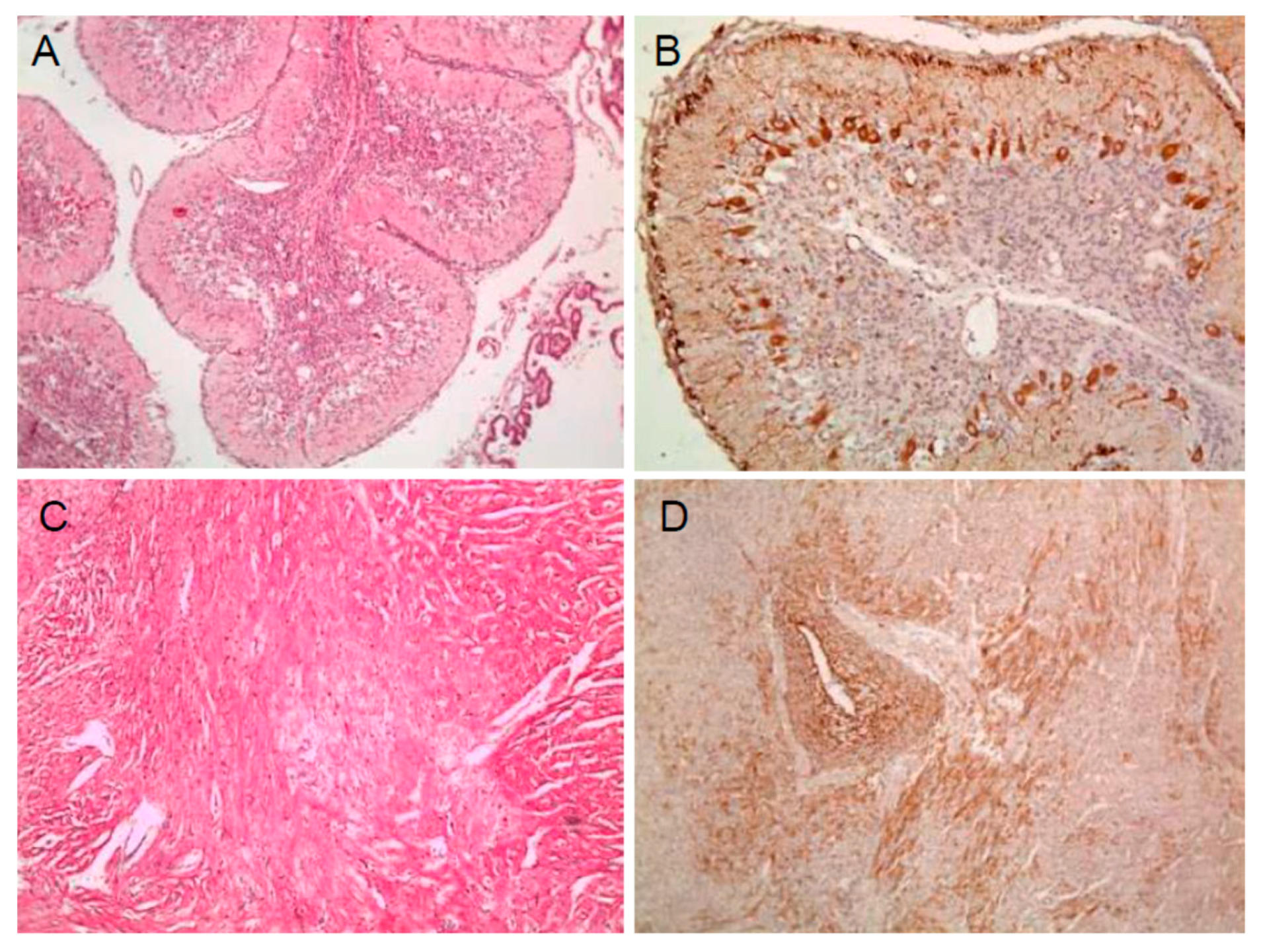
2.1. Viral Antigen Detection in Pregnant Pigs 3 Weeks Post Inoculation
2.2. Pathogenicity in Pregnant Pigs and Their Piglets
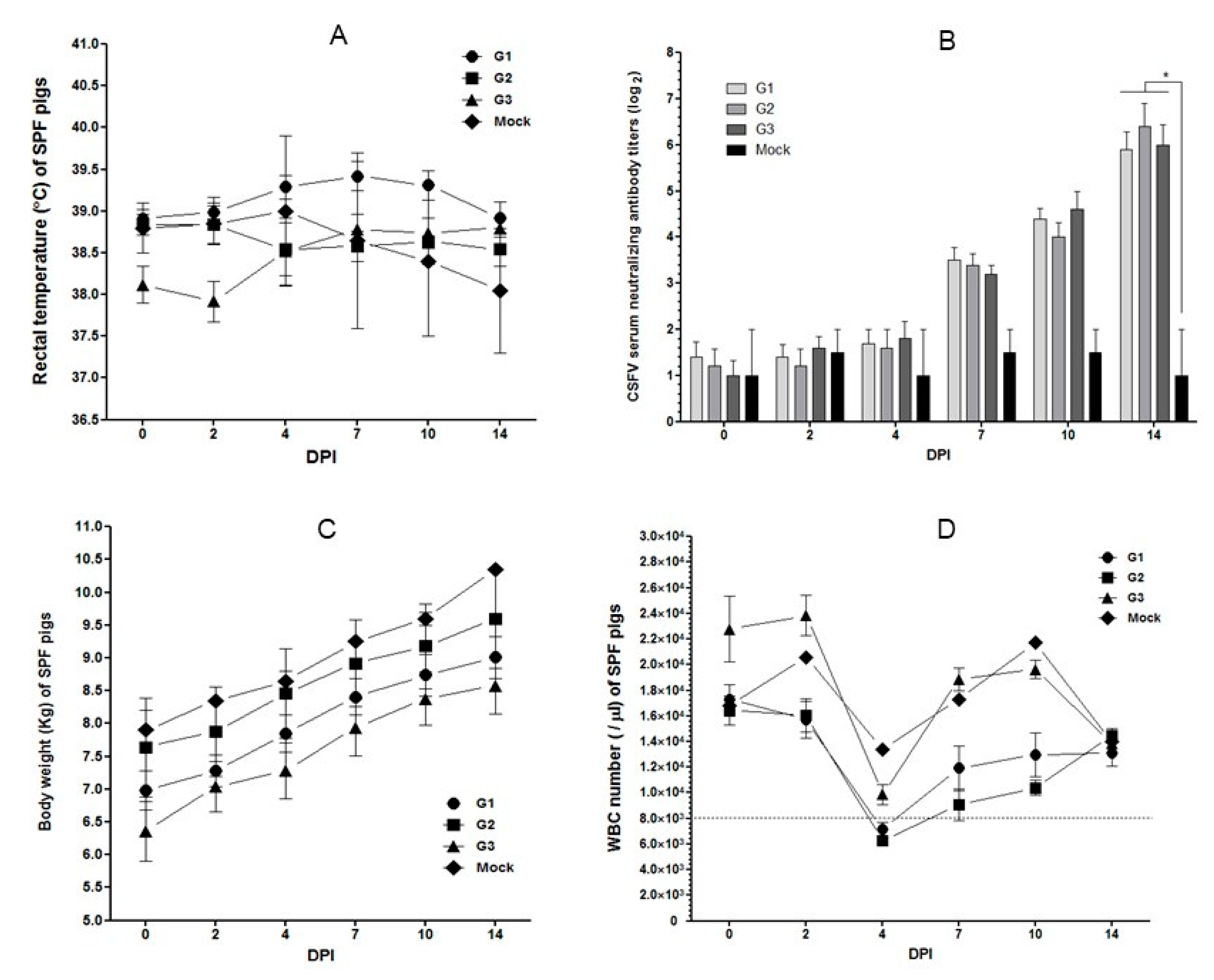
2.3. Pathogenicity in SPF Pigs
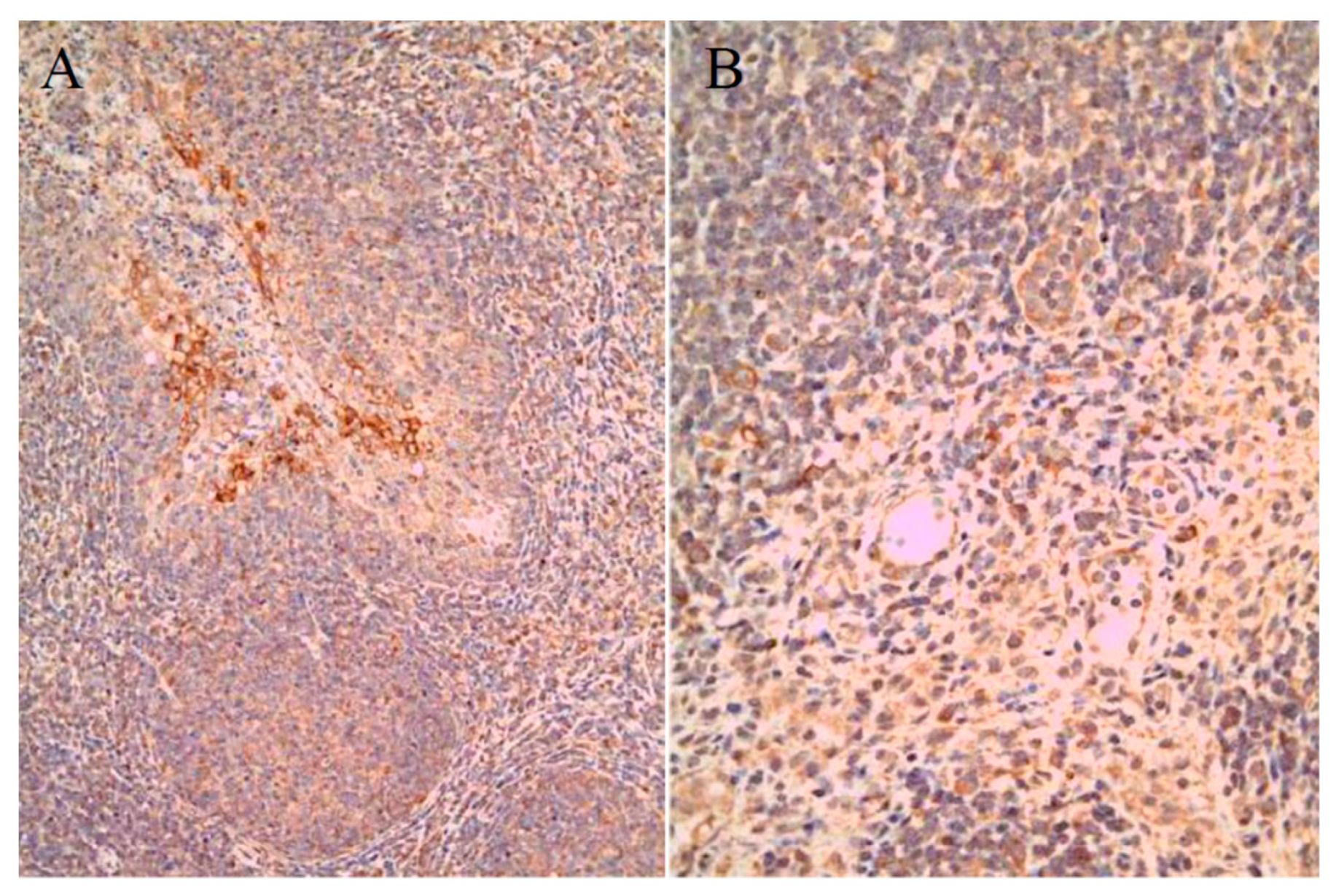
2.4. Histopathogenic Lesions and Virus Shedding in SPF Pigs
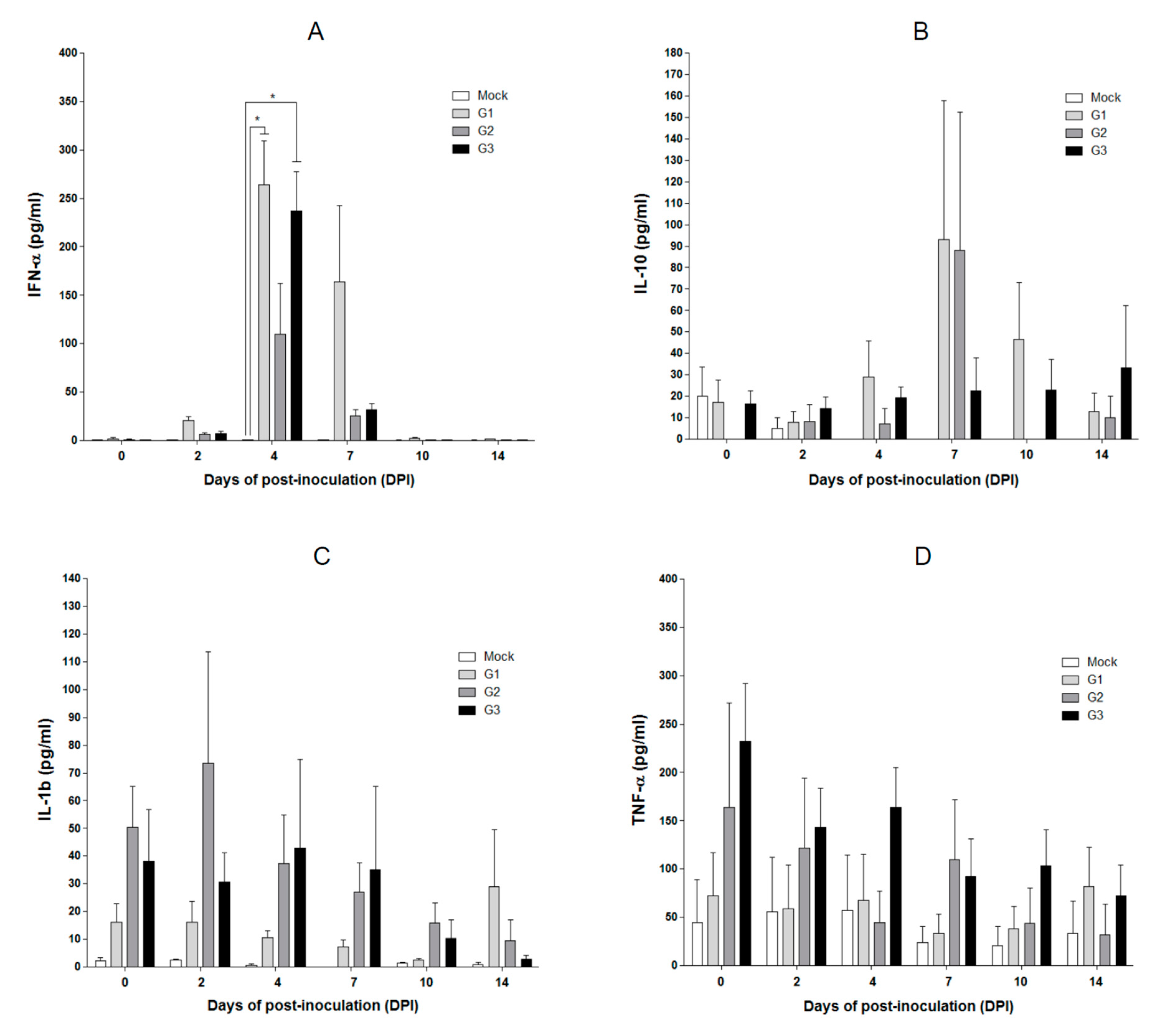
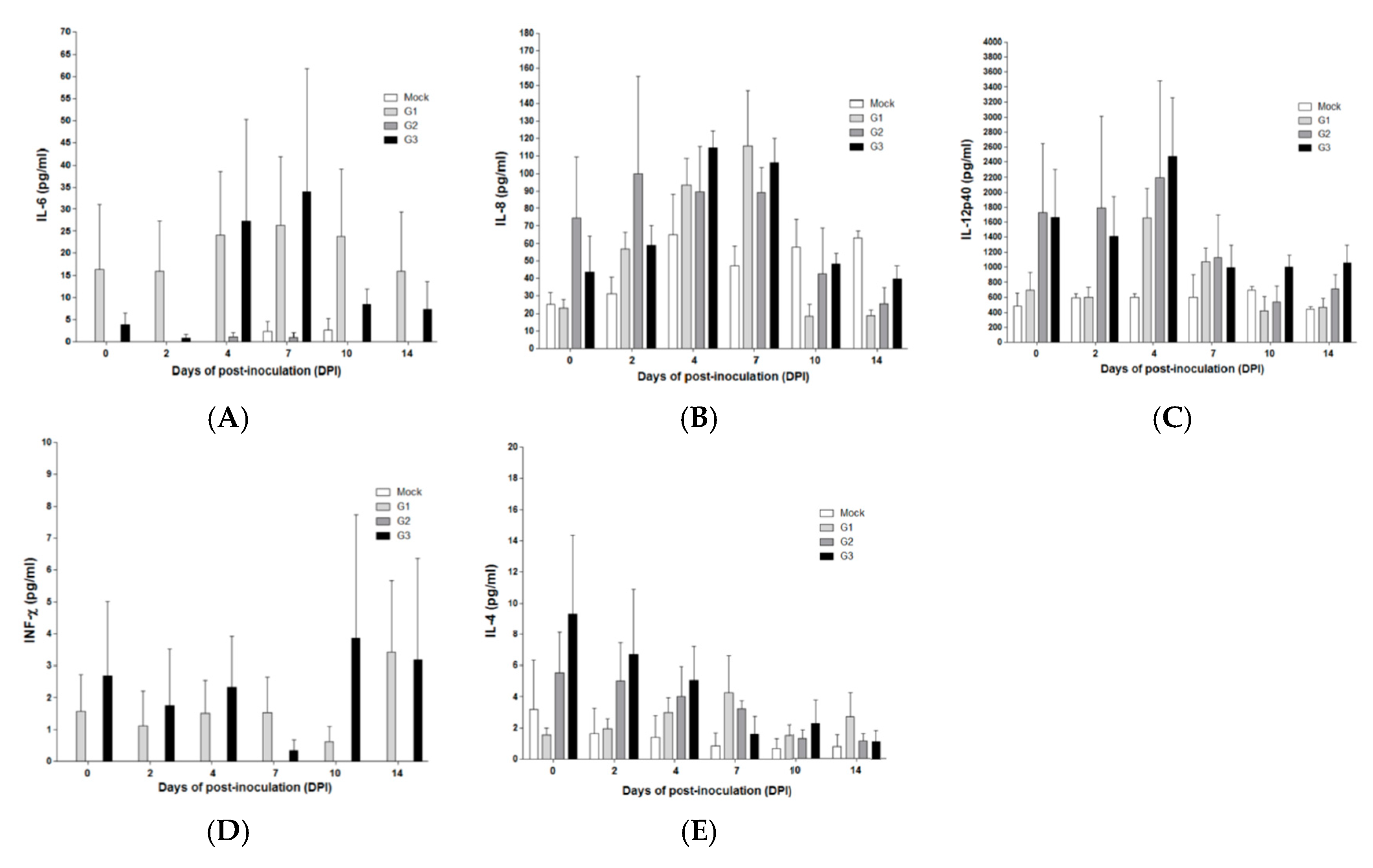
2.5. Cytokine Concentrations in SPF Pigs Inoculated with the MLV-LOM and Jeju LOM Strains
3. Discussion
4. Materials and Methods
4.1. Strains, Neutralization Antibody, IHC, and qRT-PCR
4.2. Pregnant Pigs Inoculated with MLV-LOM and Jeju LOM Strains
4.3. SPF Pigs Inoculated with the MLV-LOM and Jeju LOM Strains
4.4. Multiplex Immunoassay for SPF Pigs
4.5. Statistical Analysis
Author Contributions
Funding
Ethical Approval and Consent to Participate
Conflicts of Interest
References
- Thiel, H.J.; Collett, M.S.; Gould, E.A.; Heinz, F.X.; Houghton, M.; Meyers, G.; Purcell, R.H.; Rice, C.M. Family Flaviridae. In Virus Taxonomy: Eighth Report of the International Committee on Taxonomy of Viruses; Fauquet, C.M., Mayo, M., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Academic Press: San Diego, CA, USA, 2004; pp. 981–998. [Google Scholar]
- Meyer, H.; Liess, B.; Frey, H.R.; Hermanns, W.; Trautwein, G. Experimental transplacental transmission of hog cholera virus in pigs. IV. Virological and serological studies in newborn piglets. Zentralbl. Veterinarmed. B 1981, 28, 659–668. [Google Scholar] [CrossRef]
- von Benten, K.; Trautwein, G.; Richter-Reichhelm, H.B.; Liess, B.; Frey, H.R. Experimental transplacental transmission of hog cholera virus in pigs. III. Histopathological findings in the fetus. Zentralbl. Veterinarmed. B 1980, 27, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Stewart, W.C.; Carbrey, E.A.; Kresse, J.I. Transplacental hog cholera infection in susceptible sows. Am. J. Vet. Res. 1973, 34, 637–640. [Google Scholar] [PubMed]
- Richter-Reichhelm, H.B.; Trautwein, G.; von Benten, K.; Liess, B.; Frey, H.R. Experimental transplacental transmission of hog cholera virus in pigs. II. Immunopathological findings in the fetus. Zentralbl. Veterinarmed. B 1980, 27, 243–252. [Google Scholar] [CrossRef]
- Hermanns, W.; Trautwein, G.; Meyer, H.; Liess, B. Experimental transplacental transmission of hog cholera virus in pigs. V. Immunopathological findings in newborn pigs. Zentralbl. Veterinarmed. B 1981, 28, 669–683. [Google Scholar] [CrossRef]
- Frey, H.R.; Liess, B.; Richter-Reichhelm, H.B.; von Benten, K.; Trautwein, G. Experimental transplacental transmission of hog cholera virus in pigs. I. Virological and serological studies. Zentralbl. Veterinarmed. B 1980, 27, 154–164. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Romanini, S.; Salguero, F.J.; Nunez, A.; Bautista, M.J.; Jover, A.; Gomez-Villamos, J.C. Apoptosis of thymocytes related to cytokine expression in experimental classical swine fever. J. Comp. Pathol. 2002, 127, 239–248. [Google Scholar] [CrossRef]
- Summerfield, A.; Alves, M.; Ruggli, N.; de Bruin, M.G.; McCullough, K.C. High IFN alpha responses associated with depletion of lymphocytes and natural IFN producing cells during classical swine fever. J. Interferon Cytokine Res. 2006, 26, 248–255. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Núñez, A.; Salguero, F.J.; Pedrera, M.; Fernández de Marco, M.; Gómez-Villamandos, J.C. Lymphocyte apoptosis and thrombocytopenia in spleen during classical swine fever: role of macrophages and cytokines. Vet. Pathol. 2005, 42, 477–488. [Google Scholar] [CrossRef]
- Moennig, V.; Floegel-Niesmann, G.; Greiser-Wilke, I. Clinical signs and epidemiology of classical swine fever: A review of new knowledge. Vet. J. 2003, 165, 11–20. [Google Scholar] [CrossRef]
- Petrov, A.; Blohm, U.; Beer, M.; Pietschmann, J.; Blome, S. Comparative analyses of host responses upon infection with moderately virulent classical swine fever virus in domestic pigs and wild boar. Virol. J. 2014, 11, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaden, V.; Ziegler, U.; Lange, E.; Dedek, J. Classical swine fever virus: Clinical, virological, serological and hematological findings after infection of domestic pigs and wild boars with the field isolate “Spante” originating from wild boar. Berl. Munch. Tierarztl. Wochenschr. 2000, 113, 412–416. [Google Scholar] [PubMed]
- Lim, S.I.; Song, J.Y.; Kim, J.; Hyun, B.H.; Kim, H.Y.; Cho, I.S.; Kim, B.; Woo, G.H.; Lee, J.B.; An, D.J. Safety of classical swine fever virus vaccine strain LOM in pregnant sows and their offspring. Vaccine 2016, 34, 2021–2026. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.I.; Jeoung, H.Y.; Kim, B.; Song, J.Y.; Kim, J.; Kim, H.Y.; Cho, I.S.; Woo, G.H.; Lee, J.B.; An, D.J. Impact of porcine reproductive and respiratory syndrome virus and porcine circovirus-2 infection on the potency of the classical swine fever vaccine (LOM strain). Vet. Microbiol. 2016, 193, 36–41. [Google Scholar] [CrossRef]
- Kim, B.; Song, J.Y.; Tark, D.S.; Lim, S.I.; Choi, E.J.; Park, C.K.; Lee, B.Y.; Wee, S.H.; Bae, Y.C.; Lee, O.S.; et al. Feed comtaminated with classical swine fever vaccine virus (LOM strain) can induce antibodies to the virus in pigs. Vet Rec. 2008, 162, 12–17. [Google Scholar] [CrossRef]
- Choe, S.; Kim, J.H.; Kim, K.S.; Song, S.; Kang, W.C.; Kim, H.J.; Park, G.N.; Cha, R.M.; Cho, I.S.; Hyun, B.H.; et al. Impact of a Live Attenuated Classical Swine Fever Virus Introduced to Jeju Island, a CSF-Free Area. Pathogens 2019, 8, 206. [Google Scholar] [CrossRef] [Green Version]
- Cabezon, O.; Colom-Cadena, A.; Munoz-Gonzalez, S.; Perez-Simo, M.; Bohorquez, J.A.; Rosell, R.; Marco, I.; Domingo, M.; Lavin, S.; Ganges, L. Post-natal persistent infection with classical swine fever virus in wild boar: A strategy for viral maintenance? Transbound. Emerg. Dis. 2017, 64, 651–655. [Google Scholar] [CrossRef]
- Munoz-Gonzalez, S.; Perez-Simo, M.; Munoz, M.; Bohorquez, J.A.; Rosell, R.; Summerfield, A.; Domingo, M.; Ruggli, N.; Ganges, L. Efficacy of a live attenuated vaccine in classical swine fever virus postnatally persistently infected pigs. Vet. Res. 2015, 46, 78. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Kim, Y.H.; Jin, Y.H.; Lee, O.S.; Choi, J.U.; Kim, C.G.; An, S.H.; Lee, J.J. Hog cholera live attenuated vaccine. III. Experiment of live attenuated LOM-Suri strain. KRVS (Livestock health) 1990, 32, 49–55. [Google Scholar]
- Nielsen, J.; Lohse, L.; Rasmussen, T.B.; Uttenthal, Å. Classical swine fever in 6-and 11-week-old pigs: haematological and immunological parameters are modulated in pigs with mild clinical disease. Vet. Immunol. Immunopathol. 2010, 138, 159–173. [Google Scholar] [CrossRef]
- Summerfield, A.; Hofmann, M.; McCullough, K.C. Immature granulocytic cells dominate the peripheral blood during classical swine fever and are targets for virus infection. Vet. Immunol. Immunopathol. 1998, 63, 289–301. [Google Scholar] [CrossRef]
- Susa, M.; König, M.; Saalmuller, A.; Reddehase, M.J.; Thiel, H.J. Pathogenesis of classical swine fever: B-lymphocyte deficiency caused by hog cholera virus. J. Virol. 1992, 66, 1171–1175. [Google Scholar] [PubMed]
- Bensaude, E.; Turner, J.L.; Wakeley, P.R.; Sweetman, D.A.; Pardieu, C.; Drew, T.W.; Wileman, T.; Powell, P.P. Classical swine fever virus induces proinflammatory cytokines and tissue factor expression and inhibits apoptosis and interferon synthesis during the establishment of long-term infection of porcine vascular endothelial cells. J. Gen. Virol. 2004, 85, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.P.; Everett, H.E.; Johns, H.L.; Haines, F.J.; La Rocca, S.A.; Khatri, M.; Wright, I.K.; Drew, T.W.; Crooke, H.R. Characterisation of virus-specific peripheral blood cell cytokine responses following vaccination or infection with classical swine fever viruses. Vet. Microbiol. 2010, 142, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Jamin, A.; Gorin, S.; Cariolet, R.; Le Potier, M.F.; Kuntz-Simon, G. Classical swine fever virus induces activation of plasmacytoid and conventional dendritic cells in tonsil, blood, and spleen of infected pigs. Vet. Res. 2008, 39, 7. [Google Scholar] [CrossRef] [Green Version]
- Suradhat, S.; Sada, W.; Buranapraditkun, S.; Damrongwatanapokin, S. The kinetics of cytokine production and CD25 expression by porcine lymphocyte subpopulations following exposure to classical swine fever virus (CSFV). Vet. Immunol. Immunopathol. 2005, 106, 197–208. [Google Scholar] [CrossRef]
- Wang, J.; Sun, Y.; Meng, X.Y.; Li, L.F.; Li, Y.; Luo, Y.; Wang, W.; Yu, S.; Yin, C.; Li, S.; et al. Comprehensive evaluation of the host responses to infection with differentially virulent classical swine fever virus strains in pigs. Virus Res. 2018, 255, 68–76. [Google Scholar] [CrossRef]
- Choi, C.; Hwang, K.K.; Chae, C. Classical swine fever virus induces tumor necrosis factor-alpha and lymphocyte apoptosis. Arch. Virol. 2004, 149, 875–889. [Google Scholar] [CrossRef]
- Knoetig, S.M.; Summerfield, A.; Spagnuolo-Weaver, M.; McCullough, K.C. Immunopathogenesis of classical swine fever: role of monocytic cells. Immunology 1999, 97, 359–366. [Google Scholar] [CrossRef]
- Tang, Q.; Guo, K.; Kang, K.; Zhang, Y.; He, L.; Wang, J. Classical swine fever virus NS2 protein promotes interleukin-8 expression and inhibits MG132-induced apoptosis. Virus Genes 2011, 42, 355–362. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Inoculum Strain | No. of Pregnant Sow | Day of Inoculation Post-Pregnancy | Day of Autopsy Post-Pregnancy | Antigen Detection in Organs from Pregnant Sows | Antigen Positive Fetuses/Total Fetuses (%) | |||
|---|---|---|---|---|---|---|---|---|---|
| Tonsil | Heart | Lymph Node | Other Organs * | ||||||
| G1 | JJ16LOM-YJK08 | 37-5952 | 66 | 87 | + | - | - | - | 17/20 (85) |
| G2 | JJ16LOM-YYM02 | 51-2104 | 64 | 85 | - | + | - | - | 9/14 (64.2) |
| G3 | 16LOM-KM00 | LI-5534 | 67 | 88 | + | - | - | - | 7/9 (77.7) |
| Group | Inoculum Strain | No. of Sow | * Day of Inoculation Post-Pregnancy (Days) | Total Period of Pregnancy (Days) | No. Viral Antigen Positive/No. of Offspring | ** Positive Antigen/Test No. (%) | Crown-Rump (cm) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Total | Mummified | Stillborn | Live | |||||||
| G1 | JJ16LOM-YJK08 | 102 | 65 | 113 | 8 | 6/6 | 2/2 | - | 8/8 (100) | 19–28.5 |
| 34-5053 | 66 | 114 | 15 | 0/3 | 11/11 | *** 1/1 | 12/15 (80) | 5–28 | ||
| G2 | JJ16LOM-YYM02 | Ll-6061 | 65 | 115 | 14 | 5/5 | 6/6 | 0/3 | 11/14 (78.5) | 6.5–27.5 |
| 62-0093 | 65 | 114 | 18 | 3/6 | 9/9 | 0/3 | 12/18 (66.6) | 17–30 | ||
| G3 | 16LOM-KM00 | 55-4424 | 65 | 114 | 7 | 7/7 | - | - | 7/7 (100) | 11.5–17.5 |
| 45-0502 | 64 | 113 | 6 | 5/5 | - | 0/1 | 5/6 (83.3) | 16.5–28 | ||
| G4 | Control | 720 | 65 | 114 | 17 | - | 0/2 | 0/15 | 0/17 (0) | 25–30 |
| Group | Inoculum Strain | No. of SPF Pigs | Sample | No. of CSFV RNA Positive Pigs/No. of Pigs Tested | |||||
|---|---|---|---|---|---|---|---|---|---|
| * DPI0 | DPI2 | DPI4 | DPI7 | DPI10 | DPI14 | ||||
| G1 | JJ16LOM-YJK08 | 10 | Blood | 0/10 | 0/10 | 0/10 | ** 6/10 | 8/10 | 3/10 |
| Nasal | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | |||
| Rectal | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | |||
| G2 | JJ16LOM-YJK08-F | 5 | Blood | 0/5 | 0/5 | 0/5 | 4/5 | 5/5 | 2/5 |
| Nasal | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | |||
| Rectal | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | |||
| G3 | 16LOM-KM00 | 5 | Blood | 0/5 | 0/5 | 0/5 | 0/5 | 2/5 | 0/5 |
| Nasal | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | |||
| Rectal | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | |||
| G4 | Control | 2 | Blood | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 |
| Nasal | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | |||
| Rectal | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | |||
| Group | Inoculum Strain | No. of SPF Pigs | Autopsy Day (DPI) | Antigen in Organs * of SPF Pigs (Copy Number (Log 10) by qRT-PCR/Score by IHC) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| To | Sp | Lu | Ki | He | Li | Bl | Il | ML | SL | IL | Br | ||||
| G1 | JJ16LOM-YJK08 | 1 | 14 | 2.1/+ | 1.8/− | - | - | - | - | - | 1.9/− | - | 2.2/+ | - | - |
| 2 | 14 | 2.5/++ | 2.0/− | - | - | - | - | - | 1.4/− | 1.6/− | 1.8/− | - | - | ||
| 3 | 14 | 2.0/++ | 1.8/+ | - | - | - | - | - | 1.8/+ | - | 2.4/+ | - | - | ||
| 4 | 14 | 2.3/+ | - | - | - | - | - | - | 1.7/− | - | 1.8/− | - | - | ||
| 5 | 14 | 2.6/− | - | - | - | - | - | - | - | - | - | - | - | ||
| 6 | 21 | - | - | - | - | - | - | - | - | - | - | - | - | ||
| 7 | 21 | 1.8/+ | - | - | - | - | - | - | - | - | - | - | - | ||
| 8 | 21 | - | - | - | - | - | - | - | - | - | - | - | - | ||
| 9 | 21 | 1.6/+ | - | - | - | - | - | - | 2.2/− | - | - | - | - | ||
| 10 | 21 | 2.7/+ | - | - | - | - | - | 2.0/V | - | 1.6/− | - | - | - | ||
| G2 | JJ16LOM-YJK08-F | 1 | 21 | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 21 | 1.2/+ | 1.5/− | - | - | - | - | - | - | - | 2.0/− | 2.6/− | - | ||
| 3 | 21 | - | - | - | - | 1.8/− | - | - | - | - | - | - | - | ||
| 4 | 21 | 1.8/+ | - | - | - | - | - | - | - | - | 2.2/− | - | - | ||
| 5 | 21 | 2.3/+ | - | - | - | - | - | - | - | - | 1.9/− | - | - | ||
| G3 | 16LOM-KM00 | 1 | 14 | 1.8/+ | 1.9/− | - | - | - | - | - | 1.7/− | - | 1.6/- | - | - |
| 2 | 14 | 2.6/+ | 1.8/+ | - | - | - | - | - | 2.1/+ | 1.8/− | - | - | - | ||
| 3 | 14 | 2.4/+ | 1.3/+ | - | - | - | - | - | - | - | - | - | - | ||
| 4 | 14 | 2.1/++ | - | - | - | - | - | - | - | 1.5/− | - | - | - | ||
| 5 | 14 | 2.7/+ | - | - | - | - | - | - | - | - | - | - | - | ||
| G4 | Control | 1 | 14 | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 21 | - | - | - | - | - | - | - | - | - | - | - | - | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choe, S.; Kim, J.-H.; Kim, K.-S.; Song, S.; Cha, R.M.; Kang, W.-C.; Kim, H.-J.; Park, G.-N.; Shin, J.; Jo, H.-N.; et al. Adverse Effects of Classical Swine Fever Virus LOM Vaccine and Jeju LOM Strains in Pregnant Sows and Specific Pathogen-Free Pigs. Pathogens 2020, 9, 18. https://doi.org/10.3390/pathogens9010018
Choe S, Kim J-H, Kim K-S, Song S, Cha RM, Kang W-C, Kim H-J, Park G-N, Shin J, Jo H-N, et al. Adverse Effects of Classical Swine Fever Virus LOM Vaccine and Jeju LOM Strains in Pregnant Sows and Specific Pathogen-Free Pigs. Pathogens. 2020; 9(1):18. https://doi.org/10.3390/pathogens9010018
Chicago/Turabian StyleChoe, SeEun, Jae-Hoon Kim, Ki-Sun Kim, Sok Song, Ra Mi Cha, Wan-Choul Kang, Hyeun-Ju Kim, Gyu-Nam Park, Jihye Shin, Hyoung-Nam Jo, and et al. 2020. "Adverse Effects of Classical Swine Fever Virus LOM Vaccine and Jeju LOM Strains in Pregnant Sows and Specific Pathogen-Free Pigs" Pathogens 9, no. 1: 18. https://doi.org/10.3390/pathogens9010018