Divergent Changes in Plasma AGEs and sRAGE Isoforms Following an Overnight Fast in T1DM


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Clinical Procedures
2.3. Quantification of Circulating Protein Oxidation, and Glycation Free Adducts via LC-MS/MS
2.4. Quantification of Circulating sRAGE Isoforms
2.5. Quantification of Circulating Metabolic Substrates and Hormones
2.6. Data Analysis/Statistics
3. Results
3.1. Baseline and Metabolic Changes Following an Overnight Fast
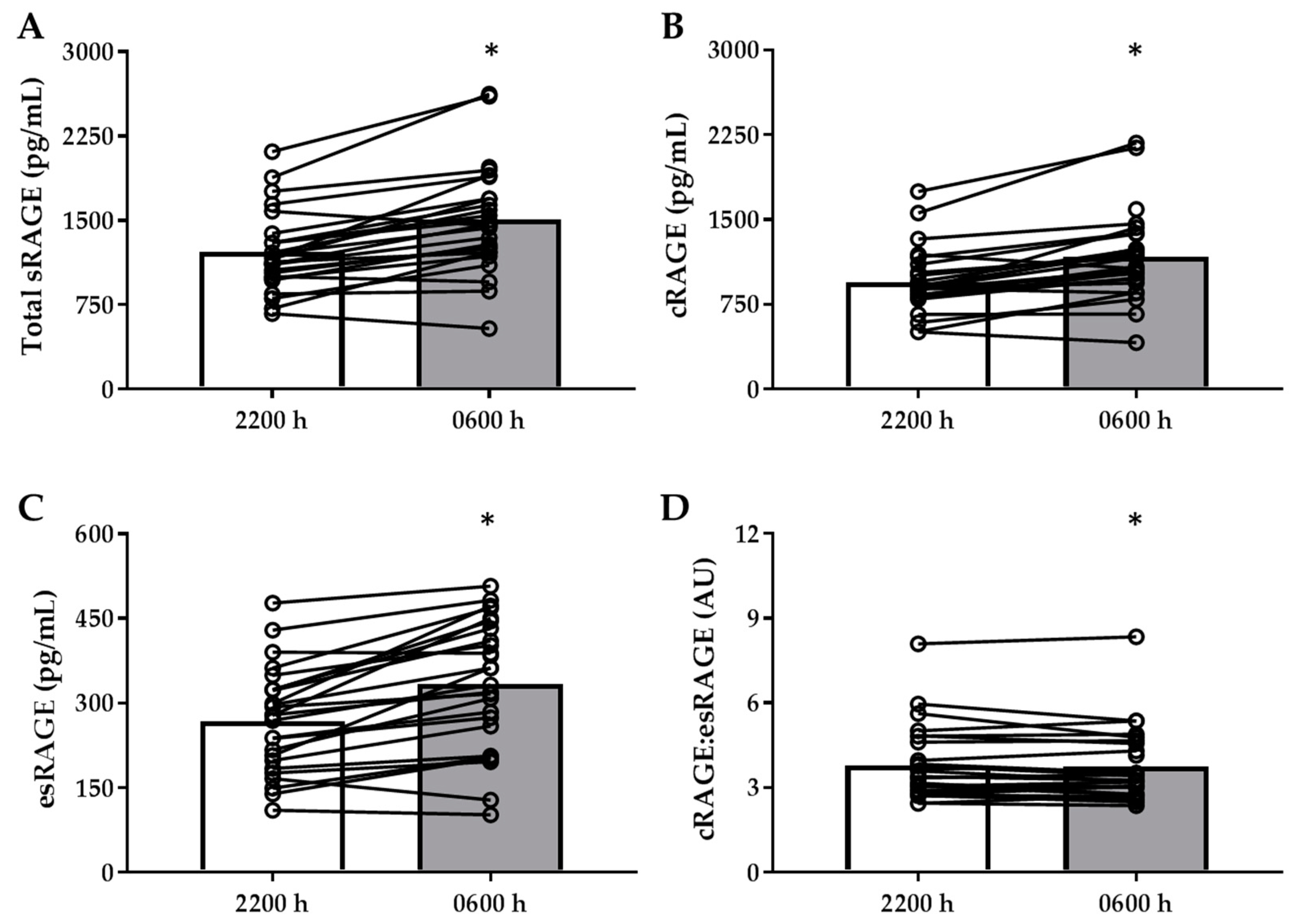
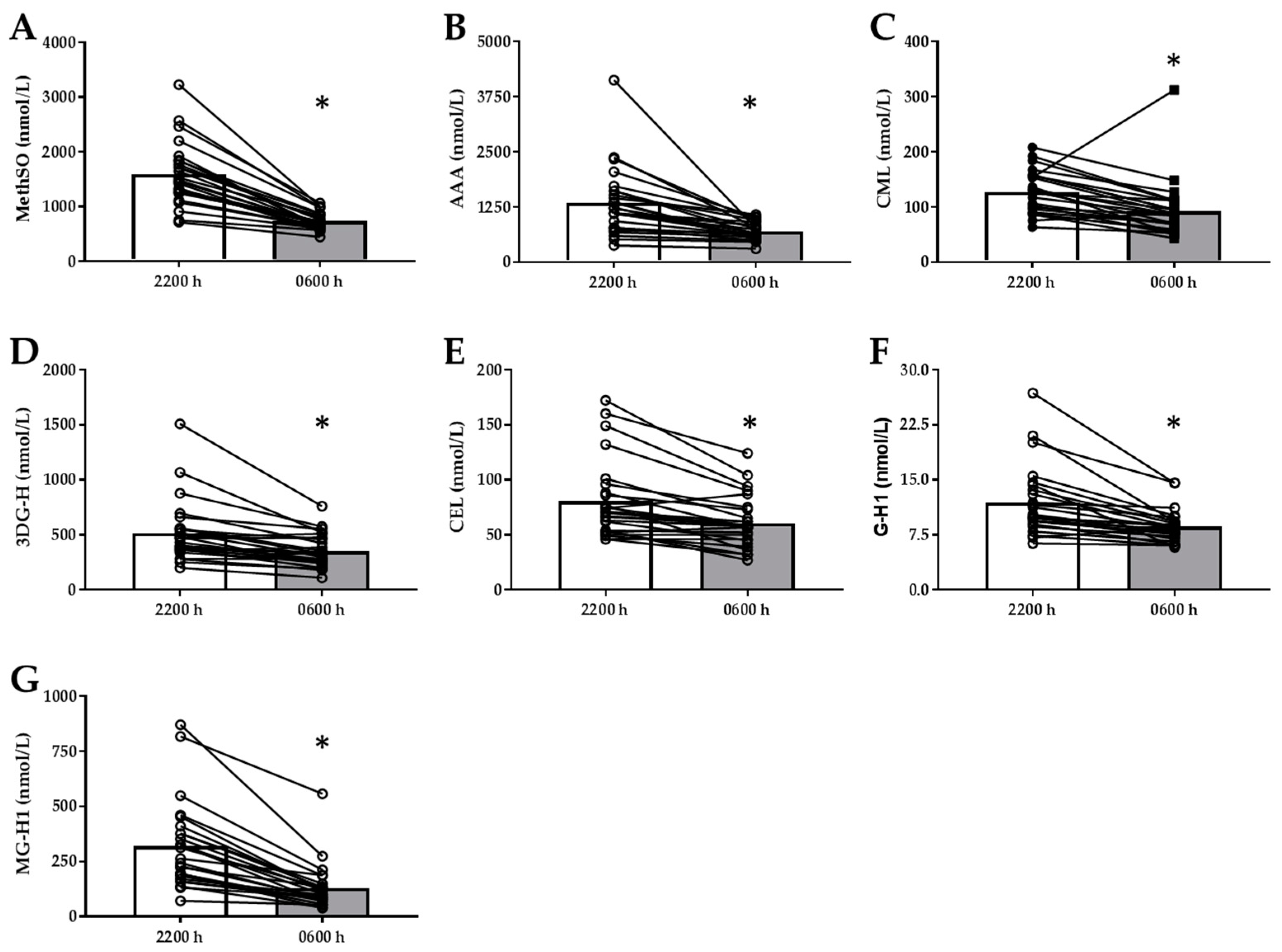
3.2. Divergent Changes Between sRAGE Isoforms, AGEs/Oxidative Stress Markers with Overnight Fast
3.3. Diurnal Changes in eGFRCystatin-C are Related to Changes in MG-H1 but Does Not Affect the Relationships between MG-H1 and sRAGE Isoforms
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Monnier, V.M. Toward a Maillard Reaction Theory of Aging. Prog. Clin. Biol. Res. 1989, 304, 1–22. [Google Scholar] [PubMed]
- Ahmed, N. Advanced glycation endproducts--role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Groener, J.B.; Oikonomou, D.; Cheko, R.; Kender, Z.; Zemva, J.; Kihm, L.; Muckenthaler, M.; Peters, V.; Fleming, T.; Kopf, S.; et al. Methylglyoxal and Advanced Glycation End Products in Patients with Diabetes—What We Know so Far and the Missing Links. Exp. Clin. Endocrinol. Diabetes 2017. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Babaei-Jadidi, R.; Howell, S.K.; Beisswenger, P.J.; Thornalley, P.J. Degradation products of proteins damaged by glycation, oxidation and nitration in clinical type 1 diabetes. Diabetologia 2005, 48, 1590–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.; Babaei-Jadidi, R.; Howell, S.K.; Thornalley, P.J.; Beisswenger, P.J. Glycated and oxidized protein degradation products are indicators of fasting and postprandial hyperglycemia in diabetes. Diabetes Care 2005, 28, 2465–2471. [Google Scholar] [CrossRef] [PubMed]
- Maessen, D.E.; Hanssen, N.M.; Scheijen, J.L.; van der Kallen, C.J.; van Greevenbroek, M.M.; Stehouwer, C.D.; Schalkwijk, C.G. Post-glucose load plasma alpha-dicarbonyl concentrations are increased in individuals with impaired glucose metabolism and type 2 diabetes: The CODAM study. Diabetes Care 2015, 38, 913–920. [Google Scholar] [CrossRef]
- Beisswenger, P.J.; Howell, S.K.; Russell, G.B.; Miller, M.E.; Rich, S.S.; Mauer, M. Early progression of diabetic nephropathy correlates with methylglyoxal-derived advanced glycation end products. Diabetes Care 2013, 36, 3234–3239. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Strokov, I.A.; Nosikov, V.V.; Savel’yeva, E.L.; Sitnikov, V.F.; Yegorov, Y.E.; Lankin, V.Z. The role of oxidative stress in diabetic neuropathy: Generation of free radical species in the glycation reaction and gene polymorphisms encoding antioxidant enzymes to genetic susceptibility to diabetic neuropathy in population of type i diabetic patients. Cell Biochem. Biophys. 2015, 71, 1425–1443. [Google Scholar] [CrossRef]
- Beisswenger, P.J.; Makita, Z.; Curphey, T.J.; Moore, L.L.; Jean, S.; Brinck-Johnsen, T.; Bucala, R.; Vlassara, H. Formation of immunochemical advanced glycosylation end products precedes and correlates with early manifestations of renal and retinal disease in diabetes. Diabetes 1995, 44, 824–829. [Google Scholar] [CrossRef]
- Chawla, D.; Bansal, S.; Banerjee, B.D.; Madhu, S.V.; Kalra, O.P.; Tripathi, A.K. Role of advanced glycation end product (AGE)-induced receptor (RAGE) expression in diabetic vascular complications. Microvasc. Res. 2014, 95, 1–6. [Google Scholar] [CrossRef]
- Gugliucci, A.; Menini, T. The axis AGE-RAGE-soluble RAGE and oxidative stress in chronic kidney disease. Adv. Exp. Med. Biol. 2014, 824, 191–208. [Google Scholar] [CrossRef]
- Haus, J.M.; Carrithers, J.A.; Trappe, S.W.; Trappe, T.A. Collagen, cross-linking, and advanced glycation end products in aging human skeletal muscle. J. Appl. Physiol. 2007, 103, 2068–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mey, J.T.; Blackburn, B.K.; Miranda, E.R.; Chaves, A.B.; Briller, J.; Bonini, M.G.; Haus, J.M. Dicarbonyl stress and glyoxalase enzyme system regulation in human skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R181–R190. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Thornalley, P.J. Methylglyoxal, glyoxalase 1 and the dicarbonyl proteome. Amino Acids 2012, 42, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M.; Hori, O.; Brett, J.; Yan, S.D.; Wautier, J.L.; Stern, D. Cellular receptors for advanced glycation end products. Implications for induction of oxidant stress and cellular dysfunction in the pathogenesis of vascular lesions. Arterioscler. Thromb. 1994, 14, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Neeper, M.; Schmidt, A.M.; Brett, J.; Yan, S.D.; Wang, F.; Pan, Y.C.; Elliston, K.; Stern, D.; Shaw, A. Cloning and expression of a cell surface receptor for advanced glycosylation end products of proteins. J. Biol. Chem. 1992, 267, 14998–15004. [Google Scholar]
- Xue, J.; Rai, V.; Singer, D.; Chabierski, S.; Xie, J.; Reverdatto, S.; Burz, D.S.; Schmidt, A.M.; Hoffmann, R.; Shekhtman, A. Advanced glycation end product recognition by the receptor for AGEs. Structure 2011, 19, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Ray, R.; Singer, D.; Bohme, D.; Burz, D.S.; Rai, V.; Hoffmann, R.; Shekhtman, A. The receptor for advanced glycation end products (RAGE) specifically recognizes methylglyoxal-derived AGEs. Biochemistry 2014, 53, 3327–3335. [Google Scholar] [CrossRef]
- Tanaka, N.; Yonekura, H.; Yamagishi, S.; Fujimori, H.; Yamamoto, Y.; Yamamoto, H. The receptor for advanced glycation end products is induced by the glycation products themselves and tumor necrosis factor-alpha through nuclear factor-kappa B, and by 17beta-estradiol through Sp-1 in human vascular endothelial cells. J. Biol. Chem. 2000, 275, 25781–25790. [Google Scholar] [CrossRef]
- Wautier, M.P.; Chappey, O.; Corda, S.; Stern, D.M.; Schmidt, A.M.; Wautier, J.L. Activation of NADPH oxidase by AGE links oxidant stress to altered gene expression via RAGE. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E685–E694. [Google Scholar] [CrossRef]
- Raucci, A.; Cugusi, S.; Antonelli, A.; Barabino, S.M.; Monti, L.; Bierhaus, A.; Reiss, K.; Saftig, P.; Bianchi, M.E. A soluble form of the receptor for advanced glycation endproducts (RAGE) is produced by proteolytic cleavage of the membrane-bound form by the sheddase a disintegrin and metalloprotease 10 (ADAM10). FASEB J. 2008, 22, 3716–3727. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.I.; Carter, A.M.; Harja, E.; Kalea, A.Z.; Arriero, M.; Yang, H.; Grant, P.J.; Schmidt, A.M. Identification, classification, and expression of RAGE gene splice variants. FASEB J. 2008, 22, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Corbett, G.T.; Gonzalez, F.J.; Pahan, K. Activation of peroxisome proliferator-activated receptor alpha stimulates ADAM10-mediated proteolysis of APP. Proc. Natl. Acad. Sci. USA 2015, 112, 8445–8450. [Google Scholar] [CrossRef] [PubMed]
- Park, L.; Raman, K.G.; Lee, K.J.; Lu, Y.; Ferran, L.J.; Chow, W.S.; Stern, D.; Schmidt, A.M. Suppression of accelerated diabetic atherosclerosis by the soluble receptor for advanced glycation endproducts. Nat. Med. 1998, 4, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Cassese, A.; Esposito, I.; Fiory, F.; Barbagallo, A.P.; Paturzo, F.; Mirra, P.; Ulianich, L.; Giacco, F.; Iadicicco, C.; Lombardi, A.; et al. In skeletal muscle advanced glycation end products (AGEs) inhibit insulin action and induce the formation of multimolecular complexes including the receptor for AGEs. J. Biol. Chem. 2008, 283, 36088–36099. [Google Scholar] [CrossRef] [PubMed]
- Agalou, S.; Ahmed, N.; Thornalley, P.J.; Dawnay, A. Advanced glycation end product free adducts are cleared by dialysis. Ann. NY Acad. Sci. 2005, 1043, 734–739. [Google Scholar] [CrossRef]
- Nin, J.W.; Ferreira, I.; Schalkwijk, C.G.; Prins, M.H.; Chaturvedi, N.; Fuller, J.H.; Stehouwer, C.D.; Group, E.P.C.S. Levels of soluble receptor for AGE are cross-sectionally associated with cardiovascular disease in type 1 diabetes, and this association is partially mediated by endothelial and renal dysfunction and by low-grade inflammation: The EURODIAB Prospective Complications Study. Diabetologia 2009, 52, 705–714. [Google Scholar] [CrossRef]
- Bakker, S.F.; Tushuizen, M.E.; Gozutok, E.; Ciftci, A.; Gelderman, K.A.; Mulder, C.J.; Simsek, S. Advanced glycation end products (AGEs) and the soluble receptor for AGE (sRAGE) in patients with type 1 diabetes and coeliac disease. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Skrha, J., Jr.; Kalousova, M.; Svarcova, J.; Muravska, A.; Kvasnicka, J.; Landova, L.; Zima, T.; Skrha, J. Relationship of soluble RAGE and RAGE ligands HMGB1 and EN-RAGE to endothelial dysfunction in type 1 and type 2 diabetes mellitus. Exp. Clin. Endocrinol. Diabetes 2012, 120, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.C.; Soderlund, J.; Lehto, M.; Makinen, V.P.; Moran, J.L.; Cooper, M.E.; Forsblom, C.; Groop, P.H.; FinnDiane Study, G. Soluble receptor for AGE (RAGE) is a novel independent predictor of all-cause and cardiovascular mortality in type 1 diabetes. Diabetologia 2011, 54, 2669–2677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katakami, N.; Matsuhisa, M.; Kaneto, H.; Matsuoka, T.A.; Sakamoto, K.; Nakatani, Y.; Ohtoshi, K.; Hayaishi-Okano, R.; Kosugi, K.; Hori, M.; et al. Decreased endogenous secretory advanced glycation end product receptor in type 1 diabetic patients: Its possible association with diabetic vascular complications. Diabetes Care 2005, 28, 2716–2721. [Google Scholar] [CrossRef] [PubMed]
- Katakami, N.; Matsuhisa, M.; Kaneto, H.; Matsuoka, T.A.; Sakamoto, K.; Yasuda, T.; Yamasaki, Y. Endogenous secretory RAGE but not soluble RAGE is associated with carotid atherosclerosis in type 1 diabetes patients. Diab. Vasc. Dis. Res. 2008, 5, 190–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, R.; Horak, K.; Lee, K.E.; Danforth, L.; Cruickshanks, K.J.; Tsai, M.Y.; Gangnon, R.E.; Klein, B.E.K. The Relationship of Serum Soluble Receptor for Advanced Glycation End Products (sRAGE) and Carboxymethyl Lysine (CML) to the Incidence of Diabetic Nephropathy in Persons with Type 1 Diabetes. Diabetes Care 2017, 40, e117–e119. [Google Scholar] [CrossRef] [PubMed]
- Heier, M.; Margeirsdottir, H.D.; Gaarder, M.; Stensaeth, K.H.; Brunborg, C.; Torjesen, P.A.; Seljeflot, I.; Hanssen, K.F.; Dahl-Jorgensen, K. Soluble RAGE and atherosclerosis in youth with type 1 diabetes: A 5-year follow-up study. Cardiovasc. diabetol. 2015, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Hurtado del Pozo, C.; Rosario, R.; Zou, Y.S.; Ananthakrishnan, R.; Xu, X.; Patel, P.R.; Benoit, V.M.; Yan, S.F.; Li, H.; et al. RAGE regulates the metabolic and inflammatory response to high-fat feeding in mice. Diabetes 2014, 63, 1948–1965. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.R.; Shin, H.K.; Park, S.Y.; Kim, H.Y.; Lee, W.S.; Rhim, B.Y.; Hong, K.W.; Kim, C.D. Cilostazol suppresses beta-amyloid production by activating a disintegrin and metalloproteinase 10 via the upregulation of SIRT1-coupled retinoic acid receptor-beta. J. Neurosci. Res. 2014, 92, 1581–1590. [Google Scholar] [CrossRef]
- Metz, V.V.; Kojro, E.; Rat, D.; Postina, R. Induction of RAGE shedding by activation of G protein-coupled receptors. PLoS ONE 2012, 7, e41823. [Google Scholar] [CrossRef]
- Roggerio, A.; Strunz, C.M.C.; Pacanaro, A.P.; Leal, D.P.; Takada, J.Y.; Avakian, S.D.; Mansur, A.P. Gene Expression of Sirtuin-1 and Endogenous Secretory Receptor for Advanced Glycation End Products in Healthy and Slightly Overweight Subjects after Caloric Restriction and Resveratrol Administration. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Miranda, E.R.; Fuller, K.N.Z.; Perkins, R.K.; Kroeger, C.M.; Trepanowski, J.F.; Varady, K.A.; Haus, J.M. Endogenous secretory RAGE increases with improvements in body composition and is associated with markers of adipocyte health. Nutr. Metab. Cardiovasc. Diseases 2018, 28, 1155–1165. [Google Scholar] [CrossRef]
- Farabi, S.S.; Carley, D.W.; Quinn, L. Glucose variations and activity are strongly coupled in sleep and wake in young adults with type 1 diabetes. Biol. Res. Nurs. 2017, 19, 249–257. [Google Scholar] [CrossRef]
- Beisswenger, P.J.; Howell, S.K.; Russell, G.; Miller, M.E.; Rich, S.S.; Mauer, M. Detection of diabetic nephropathy from advanced glycation endproducts (AGEs) differs in plasma and urine, and is dependent on the method of preparation. Amino Acids 2014, 46, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Fuller, K.N.Z.; Miranda, E.R.; Thyfault, J.P.; Morris, J.K.; Haus, J.M. Metabolic derangements contribute to reduced srage isoforms in subjects with Alzheimer’s disease. Mediat. Inflamm. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Miranda, E.R.; Somal, V.S.; Mey, J.T.; Blackburn, B.K.; Wang, E.; Farabi, S.S.; Karstoft, K.; Fealy, C.E.; Kashyap, S.R.; Kirwan, J.P.; et al. Circulating Soluble RAGE Isoforms are Attenuated in Obese, Impaired Glucose Tolerant Individuals and are Associated with the Development of Type 2 Diabetes. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E631–E640. [Google Scholar] [CrossRef] [PubMed]
- Fuller, K.N.Z.; Valentine, R.J.; Miranda, E.R.; Kumar, P.; Prabhakar, B.S.; Haus, J.M. A single high-fat meal alters human soluble RAGE profiles and PBMC RAGE expression with no effect of prior aerobic exercise. Physiol. Rep. 2018, 6, e13811. [Google Scholar] [CrossRef] [PubMed]
- Macisaac, R.J.; Tsalamandris, C.; Thomas, M.C.; Premaratne, E.; Panagiotopoulos, S.; Smith, T.J.; Poon, A.; Jenkins, M.A.; Ratnaike, S.I.; Power, D.A.; et al. Estimating glomerular filtration rate in diabetes: A comparison of cystatin-C- and creatinine-based methods. Diabetologia 2006, 49, 1686–1689. [Google Scholar] [CrossRef] [PubMed]
- Chudleigh, R.A.; Ollerton, R.L.; Dunseath, G.; Peter, R.; Harvey, J.N.; Luzio, S.; Owens, D.R. Use of cystatin C-based estimations of glomerular filtration rate in patients with type 2 diabetes. Diabetologia 2009, 52, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Farabi, S.S.; Carley, D.W.; Akasheh, R.T.; Quinn, L. Tumor necrosis factor alpha increases following sleep in young adults with type 1 diabetes. Acta. Diabetol. 2016, 53, 1049–1051. [Google Scholar] [CrossRef]
- Sarkany, Z.; Ikonen, T.P.; Ferreira-da-Silva, F.; Saraiva, M.J.; Svergun, D.; Damas, A.M. Solution structure of the soluble receptor for advanced glycation end products (sRAGE). J. Biol. Chem. 2011, 286, 37525–37534. [Google Scholar] [CrossRef]
- Koska, J.; Saremi, A.; Howell, S.; Bahn, G.; De Courten, B.; Ginsberg, H.; Beisswenger, P.J.; Reaven, P.D.; Investigators, V. Advanced glycation end products, oxidation products, and incident cardiovascular events in patients with type 2 diabetes. Diabetes Care 2018, 41, 570–576. [Google Scholar] [CrossRef]
- Liu, X.Y.; Li, H.L.; Su, J.B.; Ding, F.H.; Zhao, J.J.; Chai, F.; Li, Y.X.; Cui, S.C.; Sun, F.Y.; Wu, Z.Y.; et al. Regulation of RAGE splicing by hnRNP A1 and Tra2beta-1 and its potential role in AD pathogenesis. J. Neurochem. 2015, 133, 187–198. [Google Scholar] [CrossRef]
- Pihlajamaki, J.; Lerin, C.; Itkonen, P.; Boes, T.; Floss, T.; Schroeder, J.; Dearie, F.; Crunkhorn, S.; Burak, F.; Jimenez-Chillaron, J.C.; et al. Expression of the splicing factor gene SFRS10 is reduced in human obesity and contributes to enhanced lipogenesis. Cell Metab. 2011, 14, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Reverdatto, S.; Frolov, A.; Hoffmann, R.; Burz, D.S.; Shekhtman, A. Structural basis for pattern recognition by the receptor for advanced glycation end products (RAGE). J. Biol. Chem. 2008, 283, 27255–27269. [Google Scholar] [CrossRef] [PubMed]
- Beisswenger, P.J.; Howell, S.K.; O’Dell, R.M.; Wood, M.E.; Touchette, A.D.; Szwergold, B.S. alpha-Dicarbonyls increase in the postprandial period and reflect the degree of hyperglycemia. Diabetes Care 2001, 24, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.K.; Wang, Y.; Shiu, S.W.; Wong, Y.; Betteridge, D.J.; Tan, K.C. Effect of insulin on the soluble receptor for advanced glycation end products (RAGE). Diabet. Med. 2013, 30, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Milutinovic, P.S.; Englert, J.M.; Crum, L.T.; Mason, N.S.; Ramsgaard, L.; Enghild, J.J.; Sparvero, L.J.; Lotze, M.T.; Oury, T.D. Clearance kinetics and matrix binding partners of the receptor for advanced glycation end products. PLoS ONE 2014, 9, e88259. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, T.; Bierhaus, A.; Al-Fakhri, N.; Schneider, D.; Witte, S.; Linn, T.; Nagashima, M.; Morser, J.; Arnold, B.; Preissner, K.T.; et al. The pattern recognition receptor (RAGE) is a counterreceptor for leukocyte integrins: A novel pathway for inflammatory cell recruitment. J. Exp. Med. 2003, 198, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Skrha, J., Jr.; Soupal, J.; Loni Ekali, G.; Prazny, M.; Kalousova, M.; Kvasnicka, J.; Landova, L.; Zima, T.; Skrha, J. Skin autofluorescence relates to soluble receptor for advanced glycation end-products and albuminuria in diabetes mellitus. J. Diabetes Res. 2013, 2013, 650694. [Google Scholar] [CrossRef]
- Prasad, K.; Dhar, I.; Caspar-Bell, G. Role of advanced glycation end products and its receptors in the pathogenesis of cigarette smoke-induced cardiovascular disease. Int. J. Angiol. 2015, 24, 75–80. [Google Scholar] [CrossRef]
- Ahmed, N.; Mirshekar-Syahkal, B.; Kennish, L.; Karachalias, N.; Babaei-Jadidi, R.; Thornalley, P.J. Assay of advanced glycation endproducts in selected beverages and food by liquid chromatography with tandem mass spectrometric detection. Mol. Nutr. Food Res. 2005, 49, 691–699. [Google Scholar] [CrossRef]
- Macias-Cervantes, M.H.; Rodriguez-Soto, J.M.; Uribarri, J.; Diaz-Cisneros, F.J.; Cai, W.; Garay-Sevilla, M.E. Effect of an advanced glycation end product-restricted diet and exercise on metabolic parameters in adult overweight men. Nutrition 2015, 31, 446–451. [Google Scholar] [CrossRef] [Green Version]
- Vlassara, H.; Cai, W.; Tripp, E.; Pyzik, R.; Yee, K.; Goldberg, L.; Tansman, L.; Chen, X.; Mani, V.; Fayad, Z.A.; et al. Oral AGE restriction ameliorates insulin resistance in obese individuals with the metabolic syndrome: A randomised controlled trial. Diabetologia 2016, 59, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Uribarri, J.; Cai, W.; Pyzik, R.; Goodman, S.; Chen, X.; Zhu, L.; Ramdas, M.; Striker, G.E.; Vlassara, H. Suppression of native defense mechanisms, SIRT1 and PPARgamma, by dietary glycoxidants precedes disease in adult humans; relevance to lifestyle-engendered chronic diseases. Amino Acids 2014, 46, 301–309. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| 22:00 | 06:00 | p | |
|---|---|---|---|
| Gender (M/F) | 14/12 | N/A | |
| Age (years) | 24 ± 1 | N/A | |
| Diabetes Duration (y) | 12 ± 1 | N/A | |
| Weight (kg) | 75.6 ± 2.5 | N/A | |
| BMI (kg/m2) | 26.2 ± 0.7 | N/A | |
| HbA1C (%) | 7.69 ± 0.27 | N/A | |
| Avg Overnight Glucose (mmol/L) | 7.27 ± 0.58 | N/A | |
| Glucose (mmol/L) | 8.46 ± 0.86 | 5.67 ± 0.59 | 0.003 |
| eGFRcystatin-C (mL ·min−1 ·1.73−2) | 109.7 ± 2.8 | 107.2 ± 5.2 | 0.011 |
| Insulin (pmol/L) | 113.4 ± 12.9 | 54.1 ± 3.5 | <0.001 |
| NEFA (mEq/L) | 0.32 ± 0.04 | 0.34 ± 0.04 | 0.459 |
| Total Cholesterol (mmol/L) | 4.65 ± 0.18 | 4.63 ± 0.20 | 0.621 |
| Triglycerides (mmol/L) | 1.10 ± 0.11 | 1.02 ± 0.12 | 0.073 |
| HDL (mmol/L) | 1.51 ± 0.09 | 1.42 ± 0.08 | 0.157 |
| LDL (mmol/L) | 2.52 ± 0.16 | 2.68 ± 0.16 | 0.397 |
| Non-HDL (mmol/L) | 3.03 ± 0.18 | 3.04 ± 0.18 | 0.791 |
| IL-6 (pg/mL) | 1.02 ± 0.11 | 0.83 ± 0.11 | 0.020 |
| TNF-α (pg/mL) | 0.95 ± 0.09 | 1.43 ± 0.13 | <0.001 |
| Cortisol (nmol/L) | 822.9 ± 3.8 | 1712.3 ± 24.7 | <0.001 |
| Glucagon (ng/L) | 15.4 ± 2.4 | 8.9 ± 1.2 | 0.002 |
| ∆Total sRAGE (pg/mL) | ∆cRAGE (pg/mL) | ∆esRAGE (pg/mL) | ∆cRAGE:esRAGE | |||||
|---|---|---|---|---|---|---|---|---|
| Corr. | p | Corr. | p | Corr. | p | Corr. | p | |
| ∆MethSO (nmol/L) | −0.007 | 0.973 | −0.006 | 0.978 | −0.011 | 0.960 | −0.105 | 0.617 |
| ∆AAA (nmol/L) | 0.121 | 0.565 | 0.088 | 0.674 | 0.124 | 0.556 | −0.153 | 0.465 |
| ∆CML (nmol/L) | −0.112 | 0.594 | −0.162 | 0.438 | −0.071 | 0.737 | −0.169 | 0.419 |
| ∆3DG−H (nmol/L) | −0.041 | 0.847 | −0.084 | 0.690 | 0.121 | 0.565 | −0.353 | 0.083 |
| ∆CEL (nmol/L) | −0.235 | 0.259 | −0.274 | 0.185 | −0.195 | 0.351 | −0.145 | 0.489 |
| ∆G−H1 (nmol/L) | −0.295 | 0.153 | −0.274 | 0.185 | −0.474 | 0.017 | 0.112 | 0.596 |
| ∆ MG −H1 (nmol/L) | − 0.505 | 0.010 | −0.493 | 0.012 | − 0.589 | 0.002 | 0.103 | 0.624 |
| Dependent Variable: ΔMG-H1 (nmol/L) | ||||
|---|---|---|---|---|
| Adjusted R2 | Standardized β | 95% CI | p Value | |
| Model 1 | 0.587 | - | - | 0.0003 |
| ΔTotal sRAGE (pg/mL) | - | −0.517 | −0.492, −0.093 | 0.007 |
| Δ eGFRCystatin-C (mL ·min−1 ·1.73−2) | - | −0.294 | −7.43, 0.202 | 0.062 |
| ΔGlucose (mmol/L) | - | 0.242 | −1.50, 0.940 | 0.144 |
| ΔGlucagon (ng/L) | - | −0.083 | −6.23, 3.854 | 0.620 |
| ΔInsulin (pmol/L) | - | 0.132 | −3.20, 7.57 | 0.402 |
| ΔCortisol (nmol/L) | - | −0.288 | −1.88, 0.105 | 0.076 |
| Model 2 | 0.587 | - | - | 0.002 |
| ΔcRAGE (pg/mL) | - | −0.511 | −0.613, −0.116 | 0.007 |
| Δ eGFRCystatin-C (mL ·min−1 ·1.73−2) | - | −0.309 | −7.59, 0.006 | 0.050 |
| ΔGlucose (mmol/L) | - | 0.221 | −0.187, 0.909 | 0.182 |
| ΔGlucagon (ng/L) | - | −0.090 | −6.33, 3.74 | 0.593 |
| ΔInsulin (pmol/L) | - | 0.138 | −3.01, 7.64 | 0.380 |
| ΔCortisol (nmol/L) | - | −0.302 | −1.91, 0.056 | 0.063 |
| Model 3 | 0.502 | - | - | 0.006 |
| ΔesRAGE (pg/mL) | - | −0.415 | −1.87, −0.073 | 0.036 |
| Δ eGFRCystatin-C (mL·min−1 ·1.73−2) | - | −0.284 | −7.74, 0.770 | 0.102 |
| ΔGlucose (mmol/L) | - | 0.318 | −0.75, 1.11 | 0.083 |
| ΔGlucagon (ng/L) | - | −0.143 | −7.53, 3.39 | 0.433 |
| ΔInsulin (pmol/L) | - | 0.186 | −2.75, 8.89 | 0.279 |
| ΔCortisol (nmol/L) | - | −0.286 | −1.98, 0.226 | 0.111 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda, E.R.; Fuller, K.N.Z.; Perkins, R.K.; Beisswenger, P.J.; Farabi, S.S.; Quinn, L.; Haus, J.M. Divergent Changes in Plasma AGEs and sRAGE Isoforms Following an Overnight Fast in T1DM. Nutrients 2019, 11, 386. https://doi.org/10.3390/nu11020386
Miranda ER, Fuller KNZ, Perkins RK, Beisswenger PJ, Farabi SS, Quinn L, Haus JM. Divergent Changes in Plasma AGEs and sRAGE Isoforms Following an Overnight Fast in T1DM. Nutrients. 2019; 11(2):386. https://doi.org/10.3390/nu11020386
Chicago/Turabian StyleMiranda, Edwin R., Kelly N. Z. Fuller, Ryan K. Perkins, Paul J. Beisswenger, Sarah S. Farabi, Lauretta Quinn, and Jacob M. Haus. 2019. "Divergent Changes in Plasma AGEs and sRAGE Isoforms Following an Overnight Fast in T1DM" Nutrients 11, no. 2: 386. https://doi.org/10.3390/nu11020386