Binge-Type Eating in Rats is Facilitated by Neuromedin U Receptor 2 in the Nucleus Accumbens and Ventral Tegmental Area

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. NMUR2 Immunohistochemistry
2.3. Binge Study Design
2.4. NAc and VTA Protein Extraction
2.5. NAc and VTA NMUR2 Protein Expression
2.6. Statistical Analysis
3. Results
3.1. NMUR2 is Expressed Presynaptically in the NAc and the VTA
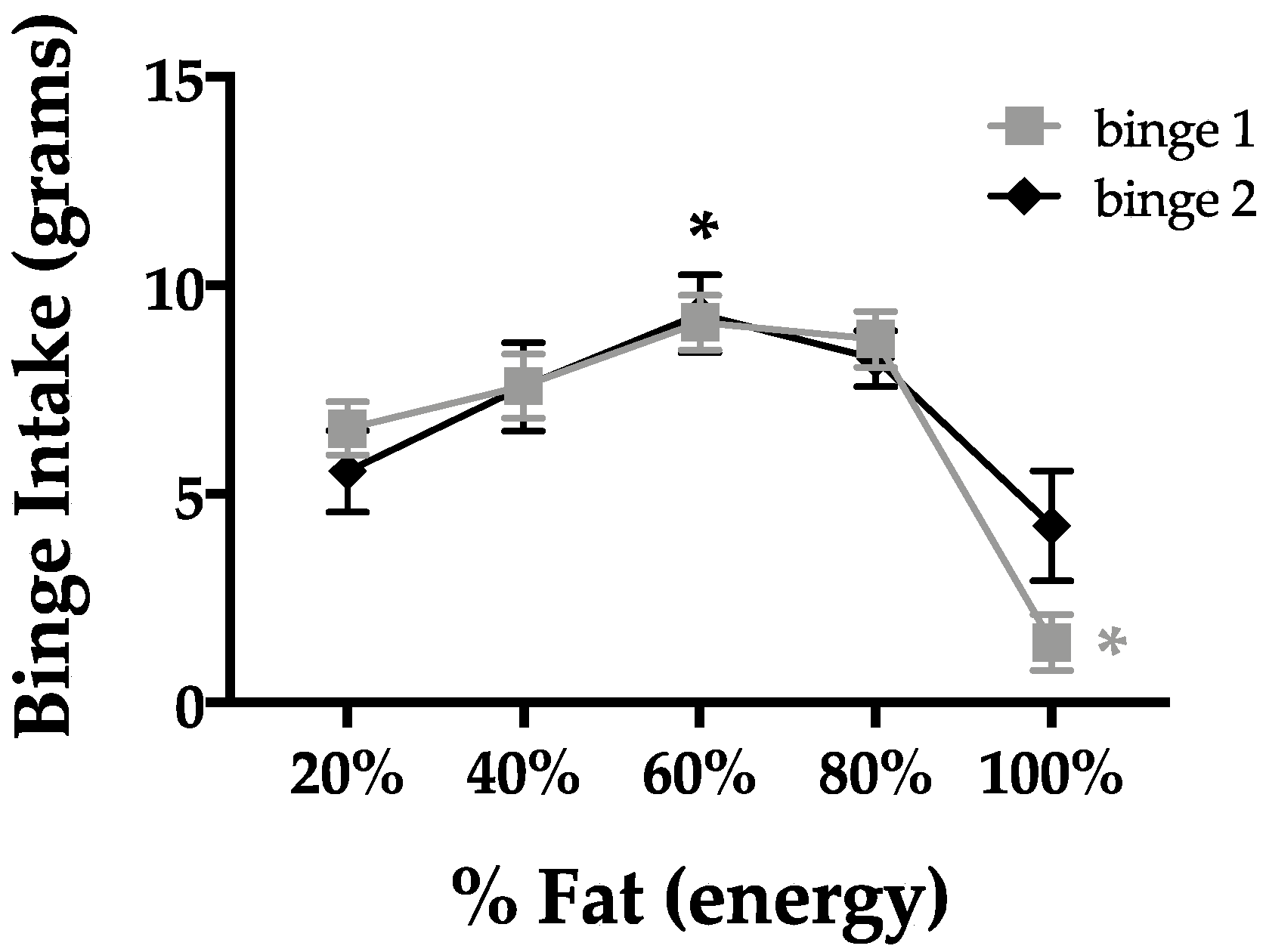
3.2. Fat Content and Binge-Type Eating
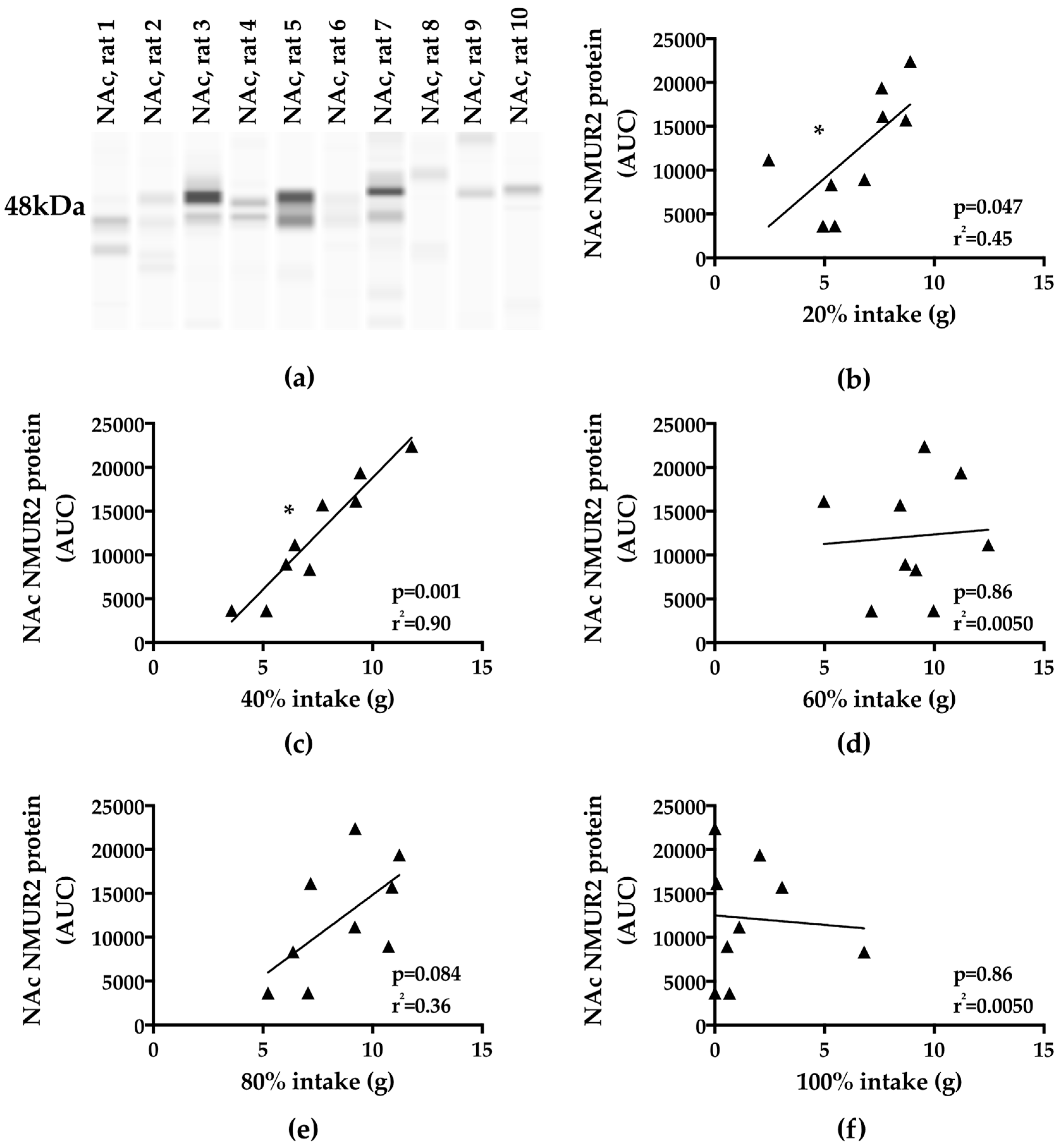
3.3. Binge-Type Intake and NAc NMUR2 Protein Expression
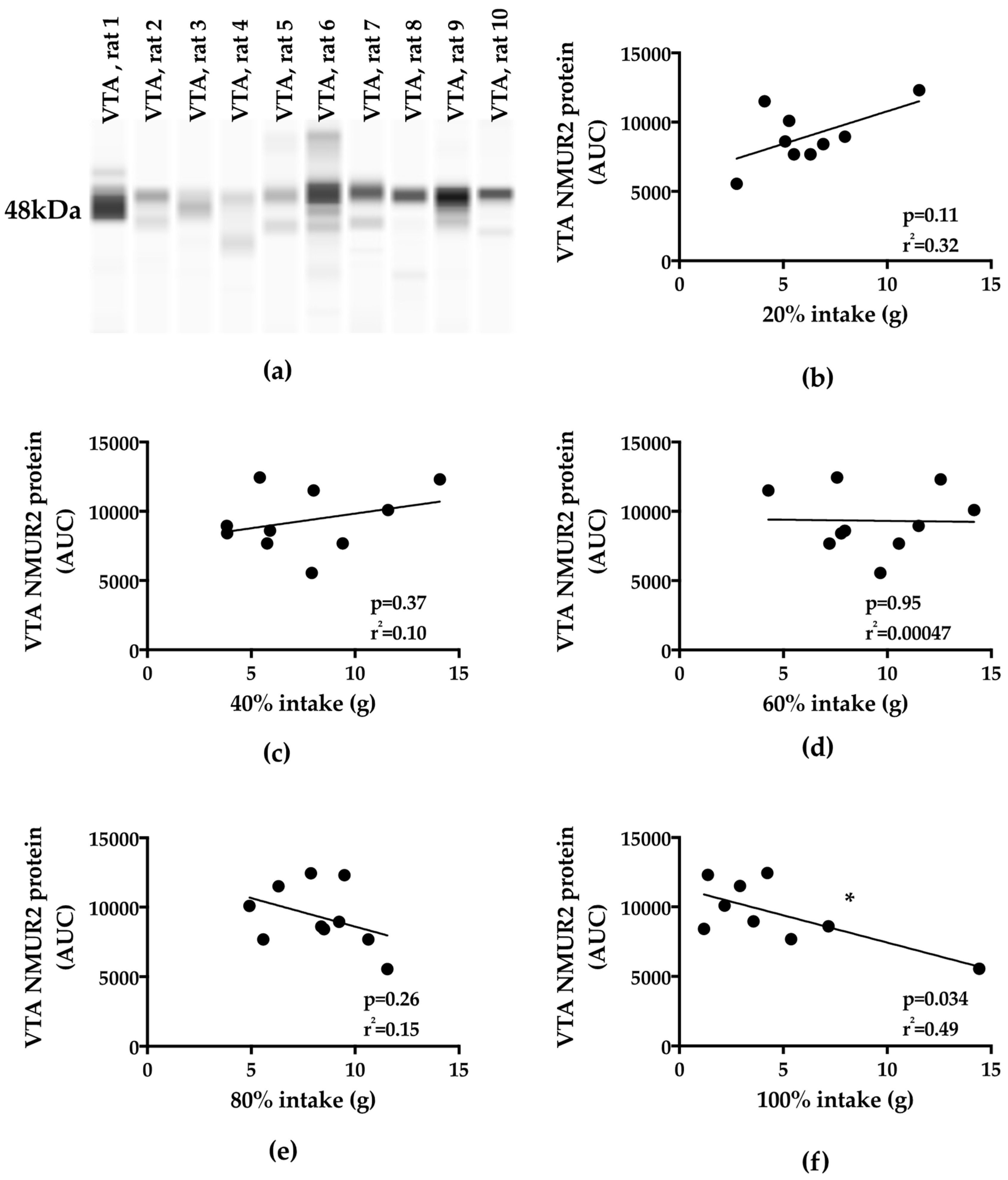
3.4. Binge-Type Intake and VTA NMUR2 Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kessler, R.C.; Berglund, P.A.; Chiu, W.T.; Deitz, A.C.; Hudson, J.I.; Shahly, V.; Aguilar-Gaxiola, S.; Alonso, J.; Angermeyer, M.C.; Benjet, C.; et al. The prevalence and correlates of binge eating disorder in the world health organization world mental health surveys. Biol. Psychiatry 2013, 73, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Guerdjikova, A.I.; Mori, N.; Casuto, L.S.; McElroy, S.L. Binge eating disorder. Psychiatr. Clin. N. Am. 2017, 40, 255–266. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Feeding and Eating Disorders; American Psychiatric Publishing: Washington, DC, USA, 2013. [Google Scholar]
- Eating Disorders and Obesity a Comprehensive Handbook, 3rd ed.; The Guilford Press: New York, NY, USA, 2017.
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Publishing: Washington, DC, USA, 2013. [Google Scholar]
- De Zwaan, M. Binge eating disorder and obesity. Int. J. Obes. Relat. Metab. Disord. 2001, 25, S51–S55. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C. ‘Liking’ and ‘wanting’ food rewards: Brain substrates and roles in eating disorders. Physiol. Behav. 2009, 97, 537–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, K.C.; Ho, C.Y.; Richard, J.M.; DiFeliceantonio, A.G. The tempted brain eats: Pleasure and desire circuits in obesity and eating disorders. Brain Res. 2010, 1350, 43–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkow, N.D.; Wang, G.J.; Tomasi, D.; Baler, R.D. The addictive dimensionality of obesity. Biol. Psychiatry 2013, 73, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Rancourt, D.; McCullough, M.B. Overlap in eating disorders and obesity in adolescence. Curr. Diab. Rep. 2015, 15, 78. [Google Scholar] [CrossRef]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of obesity among adults and youth: United States, 2015–2016. NCHS Data Brief. 2017, 219, 1–8. [Google Scholar]
- National Institutes of Health, National Heart, Lung, and Blood Institute. Managing Overweight and Obesity in Adults, Systematic Evidence Review from the Obesity Expert Panel. Available online: https://www.nhlbi.nih.gov/sites/default/files/media/docs/obesity-evidence-review.pdf (accessed on 1 December 2018).
- Drewnowski, A.; Almiron-Roig, E. Human perceptions and preferences for fat-rich foods. In Fat Detection: Taste, Texture, and Post Ingestive Effects; Montmayeur, J., le Coutre, J., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2010. [Google Scholar]
- Rippin, H.L.; Hutchinson, J.; Jewell, J.; Breda, J.J.; Cade, J.E. Portion size of energy-dense foods among french and uk adults by bmi status. Nutrients 2018. [Google Scholar] [CrossRef]
- Lyons, J.; Walton, J.; Flynn, A. Larger food portion sizes are associated with both positive and negative markers of dietary quality in irish adults. Nutrients 2018, 10, 1929. [Google Scholar] [CrossRef]
- Carstairs, S.A.; Caton, S.J.; Blundell-Birtill, P.; Rolls, B.J.; Hetherington, M.M.; Cecil, J.E. Can reduced intake associated with downsizing a high energy dense meal item be offset by increased vegetable variety in 3–5-year-old children? Nutrients 2018. [Google Scholar] [CrossRef] [PubMed]
- Reale, S.; Kearney, C.M.; Hetherington, M.M.; Croden, F.; Cecil, J.E.; Carstairs, S.A.; Rolls, B.J.; Caton, S.J. The feasibility and acceptability of two methods of snack portion control in United Kingdom (UK) preschool children: Reduction and replacement. Nutrients 2018. [Google Scholar] [CrossRef] [PubMed]
- Schebendach, J.; Broft, A.; Foltin, R.W.; Walsh, B.T. Can the reinforcing value of food be measured in bulimia nervosa? Appetite 2013, 62, 70–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkow, N.D.; Wang, G.J.; Tomasi, D.; Baler, R.D. Obesity and addiction: Neurobiological overlaps. Obes. Rev. 2013, 14, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wise, R.A. How can drug addiction help us understand obesity? Nat. Neurosci. 2005, 8, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.; Carter, J.C. Compulsive overeating as an addiction disorder. A review of theory and evidence. Appetite 2009, 53, 1–8. [Google Scholar] [CrossRef]
- Dichter, G.S.; Damiano, C.A.; Allen, J.A. Reward circuitry dysfunction in psychiatric and neurodevelopmental disorders and genetic syndromes: Animal models and clinical findings. J. Neurodev. Disord. 2012, 4, 19. [Google Scholar]
- Witt, A.A.; Lowe, M.R. Hedonic hunger and binge eating among women with eating disorders. Int. J. Eat. Disord. 2014, 47, 273–280. [Google Scholar] [CrossRef]
- Satta, V.; Scherma, M.; Piscitelli, F.; Usai, P.; Castelli, M.P.; Bisogno, T.; Fratta, W.; Fadda, P. Limited access to a high fat diet alters endocannabinoid tone in female rats. Front. Neurosci. 2018, 12, 40. [Google Scholar] [CrossRef]
- Corwin, R.L.; Wojnicki, F.H.; Zimmer, D.J.; Babbs, R.K.; McGrath, L.E.; Olivos, D.R.; Mietlicki-Baase, E.G.; Hayes, M.R. Binge-type eating disrupts dopaminergic and gabaergic signaling in the prefrontal cortex and ventral tegmental area. Obesity 2016, 24, 2118–2125. [Google Scholar] [CrossRef]
- Hommel, J.D.; Trinko, R.; Sears, R.M.; Georgescu, D.; Liu, Z.W.; Gao, X.B.; Thurmon, J.J.; Marinelli, M.; DiLeone, R.J. Leptin receptor signaling in midbrain dopamine neurons regulates feeding. Neuron 2006, 51, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Corwin, R.L.; Wojnicki, F.H. Binge eating in rats with limited access to vegetable shortening. Curr. Protoc. Neurosci. 2006. [Google Scholar] [CrossRef]
- Wojnicki, F.H.; Babbs, R.K.; Corwin, R.L. Reinforcing efficacy of fat, as assessed by progressive ratio responding, depends upon availability not amount consumed. Physiol. Behav. 2010, 100, 316–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corwin, R.L.; Avena, N.M.; Boggiano, M.M. Feeding and reward: Perspectives from three rat models of binge eating. Physiol. Behav. 2011, 104, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corwin, R.L.; Babbs, R.K. Rodent models of binge eating: Are they models of addiction? ILAR J. 2012, 53, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Babbs, R.K.; Wojnicki, F.H.; Corwin, R.L. Assessing binge eating. An analysis of data previously collected in bingeing rats. Appetite 2012, 59, 478–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzon, C.R.; Johnson, S.B.; McCue, D.L.; Li, D.; Green, T.A.; Hommel, J.D. Neuromedin u receptor 2 knockdown in the paraventricular nucleus modifies behavioral responses to obesogenic high-fat food and leads to increased body weight. Neuroscience 2014, 258, 270–279. [Google Scholar] [CrossRef]
- Price, A.E.; Stutz, S.J.; Hommel, J.D.; Anastasio, N.C.; Cunningham, K.A. Anterior insula activity regulates the associated behaviors of high fat food binge intake and cue reactivity in male rats. Appetite 2018, 133, 231–239. [Google Scholar] [CrossRef]
- Price, A.E.; Anastasio, N.C.; Stutz, S.J.; Hommel, J.D.; Cunningham, K.A. Serotonin 5-ht2c receptor activation suppresses binge intake and the reinforcing and motivational properties of high-fat food. Front. Pharmacol. 2018, 9, 821. [Google Scholar] [CrossRef]
- Kasper, J.M.; Johnson, S.B.; Hommel, J.D. Fat preference: A novel model of eating behavior in rats. J. Vis. Exp. 2014, e51575. [Google Scholar] [CrossRef]
- Howard, A.D.; Wang, R.; Pong, S.S.; Mellin, T.N.; Strack, A.; Guan, X.M.; Zeng, Z.; Williams, D.L.; Feighner, S.D.; Nunes, C.N.; et al. Identification of receptors for neuromedin u and its role in feeding. Nature 2000, 406, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Qiao, X.; Crona, J.H.; Behan, J.; Wang, S.; Laz, T.; Bayne, M.; Gustafson, E.L.; Monsma, F.J.; Hedrick, J.A. Identification of a novel neuromedin u receptor subtype expressed in the central nervous system. J. Biol. Chem. 2000, 275, 39482–39486. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, M.; Moriya, T.; Kawamata, Y.; Ohkubo, S.; Fujii, R.; Matsui, H.; Shintani, Y.; Fukusumi, S.; Habata, Y.; Hinuma, S.; et al. Identification and functional characterization of a novel subtype of neuromedin u receptor. J. Biol. Chem. 2000, 275, 29528–29532. [Google Scholar] [CrossRef] [PubMed]
- Raddatz, R.; Wilson, A.E.; Artymyshyn, R.; Bonini, J.A.; Borowsky, B.; Boteju, L.W.; Zhou, S.; Kouranova, E.V.; Nagorny, R.; Guevarra, M.S.; et al. Identification and characterization of two neuromedin u receptors differentially expressed in peripheral tissues and the central nervous system. J. Biol. Chem. 2000, 275, 32452–32459. [Google Scholar] [CrossRef] [PubMed]
- Brighton, P.J.; Szekeres, P.G.; Willars, G.B. Neuromedin u and its receptors: Structure, function, and physiological roles. Pharmacol. Rev. 2004, 56, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Egecioglu, E.; Ploj, K.; Xu, X.; Bjursell, M.; Salomé, N.; Andersson, N.; Ohlsson, C.; Taube, M.; Hansson, C.; Bohlooly-Y, M.; et al. Central nmu signaling in body weight and energy balance regulation: Evidence from nmur2 deletion and chronic central nmu treatment in mice. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E708–E716. [Google Scholar] [CrossRef] [PubMed]
- McCue, D.L.; Kasper, J.M.; Hommel, J.D. Regulation of motivation for food by neuromedin u in the paraventricular nucleus and the dorsal raphe nucleus. Int. J. Obes. 2017, 41, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Gartlon, J.; Szekeres, P.; Pullen, M.; Sarau, H.M.; Aiyar, N.; Shabon, U.; Michalovich, D.; Steplewski, K.; Ellis, C.; Elshourbagy, N.; et al. Localisation of nmu1r and nmu2r in human and rat central nervous system and effects of neuromedin-u following central administration in rats. Psychopharmacology 2004, 177, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kasper, J.; Smith, A.; Hommel, J. Cocaine-evoked locomotor activity correlates with the expression of neuromedin-u receptor 2 in the nucleus accumbens. Front. Behav. Neurosci. 2018. In press. [Google Scholar] [CrossRef]
- Zander, M.E.; De Young, K.P. Individual differences in negative affect and weekly variability in binge eating frequency. Int. J. Eat. Disord. 2014, 47, 296–301. [Google Scholar] [CrossRef]
- Kasper, J.M.; McCue, D.L.; Milton, A.J.; Szwed, A.; Sampson, C.M.; Huang, M.; Carlton, S.; Meltzer, H.Y.; Cunningham, K.A.; Hommel, J.D. Gamma-aminobutyric acidergic projections from the dorsal raphe to the nucleus accumbens are regulated by neuromedin u. Biol. Psychiatry 2016, 80, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Anastasio, N.C.; Stutz, S.J.; Fink, L.H.; Swinford-Jackson, S.E.; Sears, R.M.; DiLeone, R.J.; Rice, K.C.; Moeller, F.G.; Cunningham, K.A. Serotonin (5-ht) 5-ht2a receptor (5-ht2ar):5-ht2cr imbalance in medial prefrontal cortex associates with motor impulsivity. ACS Chem. Neurosci. 2015, 6, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Breukel, A.I.; Besselsen, E.; Ghijsen, W.E. Synaptosomes. A model system to study release of multiple classes of neurotransmitters. Methods Mol. Biol. 1997, 72, 33–47. [Google Scholar] [PubMed]
- Fink, L.H.; Anastasio, N.C.; Fox, R.G.; Rice, K.C.; Moeller, F.G.; Cunningham, K.A. Individual differences in impulsive action reflect variation in the cortical serotonin 5-ht2a receptor system. Neuropsychopharmacology 2015, 40, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Drewnowski, A.; Grinker, J.A.; Hirsch, J. Obesity and flavor perception: Multidimensional scaling of soft drinks. Appetite 1982, 3, 361–368. [Google Scholar] [CrossRef]
- Drewnowski, A.; Greenwood, M.R. Cream and sugar: Human preferences for high-fat foods. Physiol. Behav. 1983, 30, 629–633. [Google Scholar] [CrossRef]
- Drewnowski, A. Taste preferences and food intake. Annu. Rev. Nutr. 1997, 17, 237–253. [Google Scholar] [CrossRef]
- Yanovski, S.Z.; Leet, M.; Yanovski, J.A.; Flood, M.; Gold, P.W.; Kissileff, H.R.; Walsh, B.T. Food selection and intake of obese women with binge-eating disorder. Am. J. Clin. Nutr. 1992, 56, 975–980. [Google Scholar] [CrossRef]
- Sampson, C.M.; Kasper, J.M.; Felsing, D.E.; Raval, S.R.; Ye, N.; Wang, P.; Patrikeev, I.; Rytting, E.; Zhou, J.; Allen, J.A.; et al. Small-molecule neuromedin u receptor 2 agonists suppress food intake and decrease visceral fat in animal models. Pharmacol. Res. Perspect. 2018, 6, e00425. [Google Scholar] [CrossRef]
- Kaisho, T.; Nagai, H.; Asakawa, T.; Suzuki, N.; Fujita, H.; Matsumiya, K.; Nishizawa, N.; Kanematsu-Yamaki, Y.; Dote, K.; Sakamoto, J.I.; et al. Effects of peripheral administration of a neuromedin u receptor 2-selective agonist on food intake and body weight in obese mice. Int. J. Obes. 2017, 41, 1790–1797. [Google Scholar] [CrossRef]
- Kanematsu-Yamaki, Y.; Nishizawa, N.; Kaisho, T.; Nagai, H.; Mochida, T.; Asakawa, T.; Inooka, H.; Dote, K.; Fujita, H.; Matsumiya, K.; et al. Potent body weight-lowering effect of a neuromedin u receptor 2-selective pegylated peptide. J. Med. Chem. 2017, 60, 6089–6097. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, A.E.; Kasper, J.M.; Ara 13; Anastasio, N.C.; Hommel, J.D. Binge-Type Eating in Rats is Facilitated by Neuromedin U Receptor 2 in the Nucleus Accumbens and Ventral Tegmental Area. Nutrients 2019, 11, 327. https://doi.org/10.3390/nu11020327
Smith AE, Kasper JM, Ara 13, Anastasio NC, Hommel JD. Binge-Type Eating in Rats is Facilitated by Neuromedin U Receptor 2 in the Nucleus Accumbens and Ventral Tegmental Area. Nutrients. 2019; 11(2):327. https://doi.org/10.3390/nu11020327
Chicago/Turabian StyleSmith, Ashley E., James M. Kasper, Ara 13, Noelle C. Anastasio, and Jonathan D. Hommel. 2019. "Binge-Type Eating in Rats is Facilitated by Neuromedin U Receptor 2 in the Nucleus Accumbens and Ventral Tegmental Area" Nutrients 11, no. 2: 327. https://doi.org/10.3390/nu11020327