A Review on Gut Remediation of Selected Environmental Contaminants: Possible Roles of Probiotics and Gut Microbiota
by
 , ,
, ,
Pengya Feng
1,2,† ,
,
Ze Ye
1,2,†,
Apurva Kakade
2,
Amanpreet Kaur Virk
2,
Xiangkai Li
1,2 and
Pu Liu
1,2,* 1
Gansu Key Laboratory of Biomonitoring and Bioremediation for Environmental Pollution, School of Life Science, Lanzhou University, Tianshuinanlu #222, Lanzhou 730000, Gansu, China
2
Ministry of Education Key Laboratory of Cell Activities and Stress Adaptations, School of Life Science, Lanzhou University, Tianshuinanlu #222, Lanzhou 730000, Gansu, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Nutrients 2019, 11(1), 22; https://doi.org/10.3390/nu11010022
Submission received: 6 November 2018
/
Revised: 9 December 2018
/
Accepted: 17 December 2018
/
Published: 21 December 2018
(This article belongs to the Special Issue Diet as Means for studying gut-related Inflammation)
Abstract
:Various environmental contaminants including heavy metals, pesticides and antibiotics can contaminate food and water, leading to adverse effects on human health, such as inflammation, oxidative stress and intestinal disorder. Therefore, remediation of the toxicity of foodborne contaminants in human has become a primary concern. Some probiotic bacteria, mainly Lactobacilli, have received a great attention due to their ability to reduce the toxicity of several contaminants. For instance, Lactobacilli can reduce the accumulation and toxicity of selective heavy metals and pesticides in animal tissues by inhibiting intestinal absorption of contaminants and enhancing intestinal barrier function. Probiotics have also shown to decrease the risk of antibiotic-associated diarrhea possibly via competing and producing antagonistic compounds against pathogenic bacteria. Furthermore, probiotics can improve immune function by enhancing the gut microbiota mediated anti-inflammation. Thus, these probiotic bacteria are promising candidates for protecting body against foodborne contaminants-induced toxicity. Study on the mechanism of these beneficial bacterial strains during remediation processes and particularly their interaction with host gut microbiota is an active field of research. This review summarizes the current understanding of the remediation mechanisms of some probiotics and the combined effects of probiotics and gut microbiota on remediation of foodborne contaminants in vivo.
1. Introduction
The anthropogenic activities, rapid industrialization and urbanization have resulted in generation of hazardous toxic pollutants and consequent contamination of soil and water resources. For example, antibiotics (ABs) from medical waste, livestock manure and aquatic breeding have resulted in surface water contamination [1]. In addition, the large area of soil is contaminated by heavy metals (HMs) depositions and pesticides spraying [2]. It is reported that about 2.5 million hectares of soil area in Europe alone is a victim of pollution [3]. In China, mining has resulted in severe HMs contamination of 2.88 × 106 ha of land, with an additional mean area of 46,700 ha polluted annually [4]. HMs and pesticides can accumulate in agricultural products grown in the contaminated soil [5,6]. Hence, these environmental contaminants are readily transmitted into human body through water and diet, exerting negative effects on human health, such as inflammation [7,8], oxidative stress [9,10,11] and intestinal disorder [12,13,14]. Remediation of contaminants originated from environment has thus become a primary concern worldwide.
Many strategies have been developed for the remediation of soil and water contamination over the past decades, including physicochemical, microbial and phytoremediation methods. Phyto- and microbial remediation is regarded as useful approach with minimal site disruption [15], and eliminates the requirement for soil excavation and transport [16]. However, the total area that can be repaired in these traditional ways is far smaller than the total area of contamination. Thus, human exposure to contaminants is currently inevitable, and alternative methods are needed to protect not only the environment but also human against environmental contaminants.
Probiotics and live microbes that exert benefit on human health when supplemented in sufficient amounts [17], are considered as a promising tool for protection against foodborne contaminants. Evidences have shown that Lactobacilli can alleviate acute and chronic cadmium (Cd) toxicity [18,19], protect organisms against pesticides toxicity [20,21], reduce the risk of antibiotics associated diarrhea (AAD), and meantime rebalance the gut microbiota (GM) [22]. GM comprises about 3.8×1013 microorganisms inhabiting in the gastrointestinal tract (GIT), and the majority of these species belong to six bacterial phyla: Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, Fusobacteria, and Verrucomicrobia [23,24]. GM has been known to play significant role in many physiological functions, such as regulating immunity [25] and metabolism [26], and also in the bioavailability and toxicity of various contaminants [27,28]. Mounting evidences have suggested that GM can be modified by probiotics and contribute to detoxication of environmental contaminants [29]. Nevertheless, the possible roles of GM and probiotics in mediating the remediation of foodborne contaminants in mammals have not received much attention, especially how the GM modified by probiotics interacts with contaminants and how these interactions are relevant to host health. The aim of this study was to summarize the impact of environmental contaminants including HMs, pesticides and ABs on GM and host physiology, with focus on the potential mechanisms of probiotics during bioremediation processes. It could help to gain a better comprehension of the remediation mechanism of probiotics and provide new perspectives for future applications with probiotics.
2. Effect of HMs, Pesticides and ABs on the Composition and Function of GM
HMs, pesticides and ABs currently used in diverse industrial and agricultural sectors and in daily life are leading to the spread of contaminants into the environment, therefore increasing health related problems worldwide. During the last few decades, as more roles of GM are revealed, investigators have paid more attention on the impact of contaminants on GM mostly by analyzing stool microbiome in rodents, poultry, and aquatics using high-throughput sequencing techniques. These studies have demonstrated that the GM imbalance is often correlated with the occurrence of disorders of energy metabolism, nutrient absorption, and immune system [30,31].
2.1. HMs
HMs, such as Cd, chromium (Cr), arsenic (As), lead (Pb), nickel (Ni) are hard to be remediated and they exert high level of toxicity on animals and humans. About 40–60% of ingested metals are absorbed across the intestinal barrier [32,33], causing oxidative stress, inflammation, tissues damages, and gastrointestinal disorders [34].
HMs cause marked alterations in the composition of the GM (Table 1). First, a decrease in richness as well as the diversity of GM, is often observed after exposure to HMs [35,36]. Second, the ratio of Bacteroidetes to Firmicutes at phylum level is usually increased upon HMs exposure, which was thought to be associated with loss of body weight [37]. Recent studies revealed that exposure to Cd have contributed to profound effects on the microbiome in the intestinal tract of mice [38,39]. The ratio of Firmicutes to Bacteroidetes decreased significantly in mice treated with low (10 or 20 mg/kg) and high concentrations (100 mg/kg) of Cd [14,30]. Similar changes were also observed in mice exposed to As (10 mg/L) and Cr (VI) (100 mg/L) [29,40]. In Cr (VI)-treated mice, the proportion of Bacteroidetes and Tenericutes increased, and the proportion of Firmicutes declined, significantly. The exception to the tendency of alteration is Pb, where the ratio of Bacteroidetes and Firmicutes decreased [41]. Third, the influence of HMs on GM is usually dose-dependent. Higher concentration of Cd treatment posed a greater impact on intestinal flora than lower concentration [38].
The compositional and functional alterations of the GM are often linked to the intestinal and overall physical health of the host. Generally, the population of beneficial bacteria related to host physiology and biosynthesis was decreased and the number of pathogenic bacterial species correlated with the inflammation and oxidative stress was increased [29,42]. In Cd-treated mice, the abundance of beneficial bacteria such as Bifidobacteri and Lactobacilli was decreased significantly [38], whereas the relative abundance of harmful bacteria, Clostridiales, Prevotella and S24-7 was increased; and Cr (VI) induced the decrease of the relative abundance of Lachnospiraceae in mice [29]. Prevotella were associated with the transferable and colitogenic activity in a colitis mouse model [43,44]. Clostridiales affect the production of short chain fatty acids (SCFAs) [45,46]. SCFAs, mainly acetate, propionate and butyrate, are the ligands of G-protein-coupled receptor 41 (GPR41), GPR43, GRP109A, and also play important roles in the colonic epithelial cells maintenance, gluconeogenesis energy supply, and gut immunity [47,48]. Lachnospiraceae produce butyrate, with great impact in energy supply and the function of enterocytes [49]. Similarly in mice treated with Pb, the abundance of Lactococcus, Enterorhabdus, and Caulobacterales was decreased, and the abundance of Desulfovibrionaceae, Barnesiella and Clostridium was increased at the family level, which may be a contributing factor for the increase in weight and other diseases [41]. Other host phenotypes associated with the alteration of gut microbial communities include gut barrier impairment [50], increased oxidative stress in hepatocyte, hepatic inflammation and damage [29], rise in the levels of lipopolysaccharide in the serum, energy metabolism dysregulation [30], and even adult adiposity [51].
2.2. Pesticides
Pesticides are widely applied in agriculture to resist insects, weeds, and plant pathogens to promote plant growth. The crops are exposed to pesticides, which can readily get into human GIT through daily diet. Low levels of pesticides exposure can give rise to long-lasting adverse effects on skin, endocrine, and especially nervous system by inducing generation of free radicals that might cause lipid peroxidation, DNA damage, cell death and possible carcinogenic effects [52,53,54].
Many researchers have demonstrated the essential role of GM in the metabolism of pesticides in host. Some pesticides are known to be metabolized by the enzymes produced by GM. Organophosphate insecticide chlorpyrifos get metabolized into a more toxic molecule 3,5,6-trichloro-2-pyridinol via biotransformation by GM, resulting in biologically relevant and toxic consequences on host health. Whereas certain bacterial species, e.g. Pseudomonas spp. (ATCC700113), L. lactis, E. coli and L. fermentum present in GIT, are capable of utilizing 3,5,6-trichloro-2-pyridinol as their sole carbon and energy source [55,56,57]. In turn, the composition and function of GM are profoundly affected by long-term exposure to pesticides, correlated with various metabolic and immune diseases [28].
Organophosphorus (OP) pesticide has been extensively applied since 1950s. Chlorpyrifos is a typical OP insecticide that can result in altered host metabolism, increased bacterial translocation, and alterations in GM compositions. For instance, chronic chlorpyrifos exposure in rats increased the abundance of opportunistic pathogens, and unfavorable metabolic-related strains, resulting in obese and diabetic phenotypes [58]. In addition, chlorpyrifos exposure affected the proliferation of subpopulations of some strains (Enterococcus spp., Bacteroides spp.) and increased bacterial translocation in spleen and liver of rats [59,60]. In the simulator of the human intestinal microbial ecosystem(SHIME) model, chlorpyrifos also had a great impact on the population of culturable bacteria, leading to an increase in Enterobacteria, Bacteroides spp., Clostridia count and decrease in Bifidobacterial count [60,61]. Similar experiments conducted in mice showed that the relative abundance of some key microbes was significantly altered under chlorpyrifos stress, with altered urine metabolites related to the metabolism of amino acids and energy, SCFAs, phenyl derivatives and bile acids [62]. Different bile acids can bind to different receptors and promote the absorption of dietary fats, regulate lipid and glucose metabolism, and shape the GM [63,64]. GM can transform bile acids and altered GM would influence the pool of bile acids and the host’s energy metabolism.
Malathion, diazinon and glyphosate are another three representatives of OP pesticide. In malathion-treated mice, gut microbiome development and quorum sensing were perturbed, with an increase in the relative abundance of bacterial genes associated to quorum sensing-related behaviors such as motility and pathogenicity [65]. Sex-specific impact on gut microbiome by diazinon was examined in a mouse model. Specifically, several bacterial genera, including Bacteroidaceae_Bacteroides, Burkholderiales_Other, Clostridiaceae_Other, and Erysipelotrichaceae_Coprobacillus, were only observed in male mice, while Lachnospiraceae_Butyrivibrio Lachnospiraceae_Shuttleworthia, and Staphylococcaceae_Staphylococcus were completely inhibited in males after diazinon exposure [66]. The effect of glyphosate on poultry microbiota was evidenced by the elevated resistance of pathogenic bacteria including Salmonella entritidis, Salmonella gallinarum, Salmonella typhimurium, Clostridium perfringens and Clostridium botulinum, and increased susceptibility of most of the beneficial bacteria such as Enterococcus faecalis, Enterococcus faecium, Bacillus badius, Bifidobacterium adolescentis and Lactobacillus spp. [67].
Organochloric pesticide (OCP), another type of common pesticide, interferes with intestinal flora, lipid metabolism, and tissue and body weight in animals. In mice, OCP induced increased abundance of Firmicutes and Proteobacteria, and decreased abundance of Bacteroidetes, Verrucomicrobia, and Actinobacteria. Meantime, the expression of genes involved in bile acid reabsorption by the terminal ileum was down-regulated, and compensatory expression of genes in synthesis of bile acids was up-regulated in the liver [68]. When permethrin was administered through diet in rat, it caused reduction in abundance of Bacteroides-Prevotella-Porphyromonas species and increase in the abundance of Enterobacteriaceae and Lactobacillus in fecal microbiota; altered SCFAs levels were registered over a 4-month period [69]. Pentachlorophenol exposure in gold fish led to an increased in the Bacteroidetes abundance and a decrease in the ratio of Firmicutes to Bacteroidetes in the gut, which played crucial roles in the reduction of body weight. Bacteroides genus within the Bacteroidetes phylum was significantly correlated with pentachlorophenol exposure dosage and duration [70].
Imidazole is widely used to inhibit fungus in agriculture. Recent studies revealed that GM dysbiosis induced by imidazole exposure is often associated with hepatic metabolism disorder and hepatic toxicity. When imazalil was orally given in zebrafish and mice, the abundance of Bacteroidetes was decreased, and Firmicutes increased in the gut at phylum level. In mice at the genus level, the abundance of Lactobacillus and Bifidobacterium decreased while those of Deltaproteobacteria and Desulfovibrio increased in response to imazalil exposure. In addition, the transcription of genes such as Aco, Cpt1, Acc1, Srebp1a and Fas, related to glycolysis and lipid metabolism was significantly decreased in the mouse liver [71,72]. In the mice that were exposed to carbendazim, the amounts of Bacteroidetes in the feces, and richness and diversity of GM in the cecum decreased significantly after the 5-day exposure. Analysis of operational taxonomic units (OTU) indicated that a total of 361 out of 3271 identified OTUs were significantly changed [31].
2.3. ABs
Abs are widely used in stockbreeding, veterinary and human medicines [73,74]. Part of the ingested ABs by humans and animals can enter the environment through feces or urine [75]. Large quantity of ABs was detected in the ecosystem [76,77]. Hence, humans are readily exposed to antibiotic contamination passively in addition to medical route. The side effects of ABs range from relative mild ones, such as allergy, asthma, and diarrhea to severe ones, e.g., death [78].
ABs administration has been correlated with changes in the population structure of microbiome, which might be linked to a multitude of diseases. In particular, AAD and Clostridium difficile infections can be common following ABs treatment [79,80]. It has been previously shown that the microbial diversity was significantly reduced after treatment with ampicillin, streptomycin and clindamycin in the cecal and large intestine contents of mice. The Bacteroidetes population was drastically reduced, which never fully recovered following cessation of treatment, and the outgrowth of two dominant genus, Stenotrophomonas and Xanthamonas [81]. The predominant genus Stenotrophomonas is noteworthy since this highly antibiotic resistant bacterium is also a potential emerging opportunistic pathogen [82]. Treatment with clindamycin and ampicillin made the patients susceptible to Clostridium difficile infection and decreased Clostridium scindens count, which is a secondary modulator of bile acid metabolism [83]. A number of recent studies revealed that the abundance of Proteobacteria phylum in microbiota was significantly increased as a consequence of antibiotic administration [84,85,86]. Proteobacteria encompass a wide variety of pathogens, such as Escherichia, Vibrio, Salmonella, Helicobacter, Yersinia, Legionellales and others. E. coli is responsible for a vast majority of Escherichia-related pathogenesis, and other members of this genus have also been implicated in human diseases [87,88]. Salmonella species are known intracellular pathogens and certain serotypes are responsible for illness [89]. Altogether these findings suggest that altered structure of intestinal microbiota is related to the pathogenesis of diseases.
ABs can affect the colonization resistance of host. Treatment with cefoperazone [90], tigecycline [79], clindamycin [80], or clindamycin in combination with a five-antibiotic cocktail in C57BL/6 mice had decreased the colonization resistance, as a result of a decrease in Lachnospiraceae and Barnesiella and an increase in Lactobacillaceae and Enterobacteriaceae. These results were largely consistent with human studies [91,92].
Effect of ABs on GM can be persistent. Fouhy et al. (2012) [84] evaluated the short-term recovery of the GM following parenteral ampicillin and gentamicin treatment for infant within 48 hours of birth. It was shown that the abundance of Proteobacteria remained significantly higher and the number of different Bifidobacterium species was reduced in the infants after 8 weeks of treatment with ABs. It is, thus, obvious that the use of certain ABs in early life can significantly affect the evolution of the infant GM. Another study investigated the short and long-term effects of macrolides on 2–7 year old children (N = 142), and found depletion of Actinobacteria, increased abundance of Bacteroidetes and Proteobacteria and increased macrolide resistance, which can persist for over 6 months. Additionally, it was mentioned that the use of macrolides in early life increased the risk of asthma and weight gain [85]. A study in mice reported that Bacteroidetes was drastically reduced following treatment with the antibiotic mixture of ampicillin, streptomycin, and clindamycin and never fully recovered after cessation of ABs treatment [81].
The literature regarding the role of altered GM in the development of ABs-related side effects, however, is scarce. The current understanding is that oral intake of ABs lead to disturbance of composition and more importantly the metabolism of GM, which might correlate with disrupted physiology of the host. Study in mice treated with combinative ABs of penicillin, vancomycin and chlortetracycline revealed significant alterations of microbial structure, and altered regulation of hepatic metabolism of lipids and cholesterol, as well as increase of the copies of key genes involved in the metabolism of SCFAs synthesis in fecal and cecal samples [93]. Metagenomic analysis in mice receiving early-life therapeutic-dose pulsed tylosin showed that tylosin intervention decreased the modules involved in glycolysis, gluconeogenesis and tRNA biosynthesis and increased the modules involved in citric acid cycle and nucleoside and amino acid biosynthesis [94]. A study in piglets treated with a mixture of ampicillin, gentamicin and metronidazole also indicated that altered GM was associated with increased metabolism of aromatic amino acids and decreased expression of neurotransmitter in hypothalamus [95].
3. Probiotics as a Potential Tool in Contaminants Remediation
Increasing evidence demonstrated that oral supplementation of probiotics is one of the effective strategies for protection against foodborne contaminants-induced toxicity. In general, probiotics applied in toxicant remediation are selected based upon their safety and viability during passage through the GIT [97] and importantly their capacity of contaminants adsorption [98]. The probiotic effects on the hosts are usually assessed by monitoring the individual growth, measuring the amount of pollutant and related biomarkers in tissues, and analyzing the compositional and functional changes of stool microbiota using 16S rRNA sequencing in murine and other models. Nonetheless, the interaction between probiotics and the GM is still poorly understood.
3.1. Role of Probiotics in HMs Remediation In Vivo
The protective effects of probiotics against HMs toxicity have been extensively studied. Supplementation of single probiotic or a combination of probiotics in mammalians has shown positive results in alleviating the toxicity of HMs including Cd, Hg, Cr, As and Pb (Table 2).
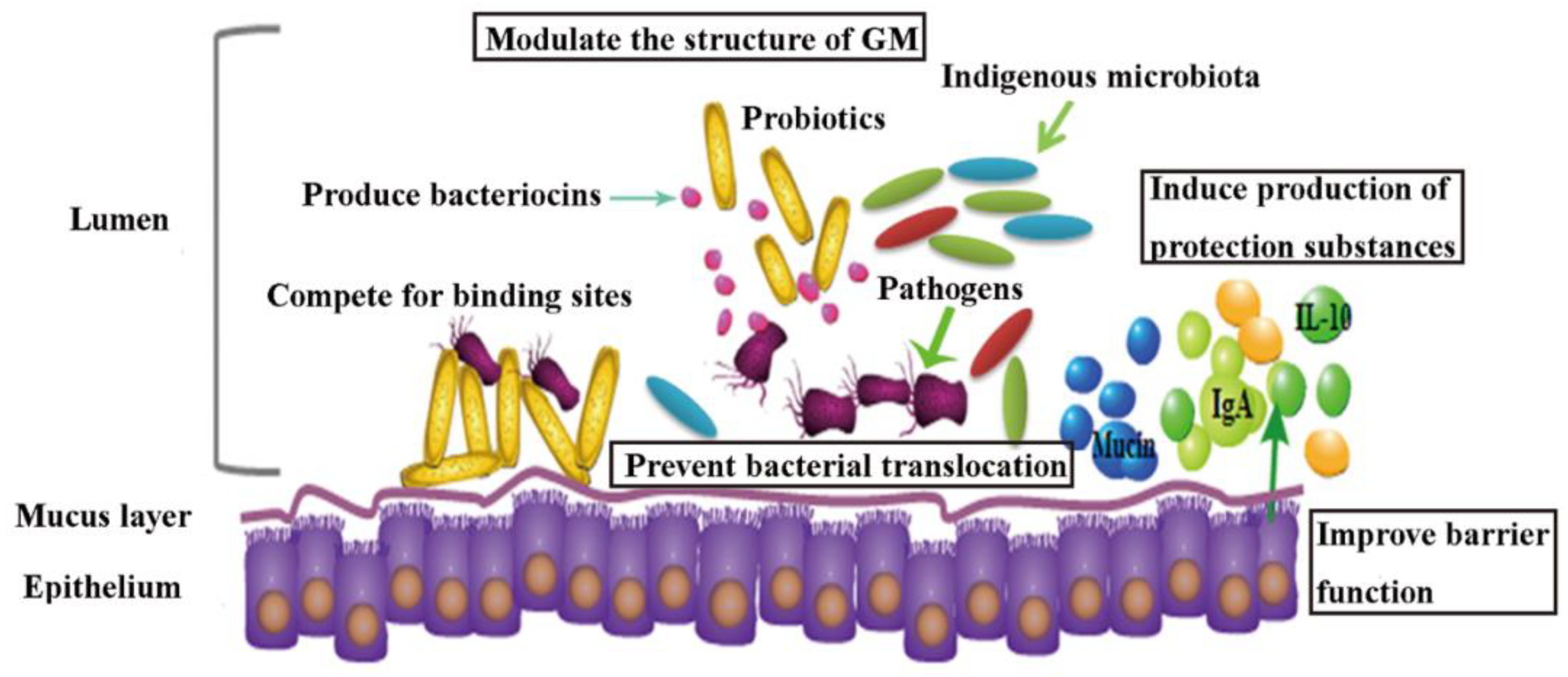
Probiotics utilized to reduce the toxicity of HMs are generally Lactobacilli, as they have excellent binding capacity for HMs, evidently lowering the availability of HMs for the host [99]. It has also been speculated that living probiotic strain L. plantarum CCFM8610 might competitively inhibit the intestinal absorption of Cd by increasing the dissolution and uptake of divalent essential elements like Ca, Mg, and Fe [18]. Probiotic strains can also promote gastrointestinal peristalsis, hence the excretion of HMs in feces is facilitated [18]. Furthermore, probiotic strains can limit the entrance of HMs by enhancing intestinal barrier function and regulating tight junction of epithelium of small intestine. Administration of L. plantarum CCFM8610 reversed all of the reductions of mRNA expression of tight-junction proteins (ZO-1, ZO-2, occludin, and claudin-1) caused by Cd exposure, decreased intestinal permeability and reduced Cd leakage into systemic circulation [50]. Preventing systemic absorption of HMs by probiotics thus leads to alleviation of oxidative stress in various tissues and consequent mitigation of tissue damages as reported [100,101,102]. For example, co-treatment of L. plantarum CCFM8610 and Cd cause a decreased production of metallothionein and downregulation expression of genes in the mitogen-activated protein kinases (MAPK) pathways in the liver [19]. Metallothionein has a high affinity for divalent cations [103] and the MAPK pathway is associated with reactive oxygen species production [19]. More recently, evidences have suggested that probiotics play a role in restoring the altered composition and function of GM induced by HMs. L. reuteri DSM17938 intervention contributed to restoring intestinal homeostasis in patient with low-Ni diets and the increase of lactic acid bacteria (LAB) biodiversity [104]. In the case of reducing Cr (VI) toxicity in mice, the crucial role of probiotic strain L. plantarum TW1-1 in maintaining GM homeostasis and enhancing Cr (VI)-reduction ability of intestinal bacteria was underscored [29]. The proposed protective mechanisms of probiotics on HMs remediation are shown in Figure 1.
To date, almost all studies on the efficacy of probiotics were carried out in animals, the only case reported in human was that of L. rhamnosus GR-1 (LGR-1)-supplemented yogurt which protected against the absorption of As and Hg in pregnant women and children [105]. Moreover, the effect of HM bioremediation by probiotics is strain-dependent and specific. Although the strain LGR-1 was effective in reducing Hg and As absorption, it could not significantly reduce the blood levels of Pb and Cd in populations, indicating the need for specific probiotics or cocktails of probiotics for protection against different types of HMs.
3.2. Probiotics’ Role in Pesticides Remediation In Vivo
Expensive drugs have been developed and long-time therapies have been employed to fight against damages caused by pesticides [106]. More economic alternatives are hence needed to reduce the adverse effects of pesticides. Mounting evidences have highlighted probiotics in mitigating the adverse effects of pesticides (Table 2), and their protective mechanisms are summarized as below. First, Lactobacilli protect against pesticides-induced oxidative stress and downstream cellular damage. A few researches have shown that supplementation with L. plantarum BJ0021 can decrease oxidative stress and MDA concentration in liver and kidney induced by endosulfan [107]. Another study showed L. casei ATCC334 could decrease DNA damage in rats exposed to a carcinogen 1, 2-dimethylhydrazine [54]. Second, probiotics maintain the integrity of intestinal barrier and reduce the absorption of pesticides [108]. It was found that L. plantarum MB452 enhanced the expression of tight junction proteins occludin, ZO-1, ZO-2, and cingulin in the Caco-2 intestinal cell-line [109]. Probiotics L. rhamnosus strain GG (LGG) and LGR-1 reduced the absorption of parathion or CP in a Caco-2 Transwell model [21]. Third, recent studies found that a few probiotics, mainly Lactobacillus from dairy products and wheat, were capable of degrading OCP enzymatically with phosphohydrolase [98,110]. Fourth, Lactobacilli stimulate host’s own immunity and detoxification mechanisms to resist pesticides and pathogen invasion. In the study using pattern insects, L. casei was found stimulating phase-II detoxification system and rescued malathion-induced physiological impairments in Caenorhabditis elegans [20]. Probiotic L. plantarum ATCC14917 has shown to stimulate immunity, and lower the pathogenic microorganism (Serratia marcescens) infections in fruit flies exposed to imidacloprid [111].
3.3. Probiotic Intervention in AAD Patients and Animal Models
There are a significant number of studies demonstrating the benefits of probiotics in reducing the occurrence of AAD, allergy, lactose intolerance, reduction of cholesterol etc. [112,113]. Patients receiving ABs for treatments are prone to suffer from gastrointestinal disturbances result from damage of the GI mucosal cells and disruption of the gut ecological balance. Probiotics replenish the natural GIT with nonpathogenic bacteria, and are considered as living drugs that help with ABs-associated diseases, without affecting the efficacy of ABs.
There are many favorable outcomes of probiotics in reducing the risk of AAD in adults and children based on extensive meta-analyses, and only a few studies using probiotics in patients undergoing antibiotic failed to acquire significant effect [114,115]. In a trial study with 246 children, co-treatment of Saccharomyces boulardii and ABs has been reported to lower the risk of diarrhea from 20.9% to 8.8% [116]. The updated results of meta-analysis, based on 10 RCTs, also showed that S. boulardii effectively prevented AAD in patients, with decrease of risk from 17.4% to 8.2% in adults [117]. The efficacy of LGG for preventing AAD in children and adults has also been evaluated. Treatment with LGG reduced the risk of AAD in patients receiving ABs from 22.4% to 12.3% [118]. The hospital patients who were administered with bio- yogurt containing a combined dose of L. acidophilus, L. delbrueckii, subspecies bulgaricus, and S. thermophiles showed reduced risk from 24% to 12% of AAD [119]. Furthermore, the effects of single and combinative probiotics on preventing AAD were compared. A meta-analysis in 2006 has reported that most of the RCTs used combinations of Lactobacillus species which were effective against diarrhea and the relative risk of AAD is 0.43, but there is no high-quality evidence for a single probiotic strain except for S. boulardii [120].
The efficacy of probiotic strains in reducing the risks of AAD in humans have been evidenced, however, the underlying mechanism of these probiotic strains is less well understood. By reviewing recent literatures (Table 2), probiotics have been proposed to be effective in alleviating ABs-associated diseases through multiple routes (Figure 2): (1) mediating the structure of gut microbial community [81,121,122] by promoting beneficial bacteria and suppressing opportunistic pathogens. A cocktail of L. rhamnosus A 191, L. acidophilus, B. breve and B. longum significantly caused suppression of gut opportunistic pathogens Enterobacteriaceae and promotion of Firmicutes following ABs treatment in mice [81]. In another study, it was confirmed that probiotic cocktail of four Lactobacillus species JUP-Y4 treatment decreased the levels of Desulfovibrionales, and promoted the levels of Akkermansis [122]. High abundance of Desulfovibrionales were related with Crohn’s disease [123] and human infections [124,125], and Akkermansis are biomarkers of intestinal health [126] and inversely linked with the severity of Crohn’s disease and ulcerative colitis [127]. Two probiotics Phaeobacter inhibens S4Sm and Bacillus pumilus RI06-95Sm in black molly, have been shown to colonize in intestine and reverse mortality caused by streptomycin by inhibiting Vibrio anguillarum [121], which are known opportunistic pathogens in fish [128] and are thought to be “r-strategists” capable of rapid growth and virulence in disturbed microbial communities [129,130]. (2) Improving immune function of host by enhancing anti-inflammation [131,132,133,134]. Shi et al. (2017) used two Lactobacillus cocktails (LacA and LacB, each contains four strains) to restore the cefixime-induced GM disturbance in mice, and alleviate intestinal inflammation possibly due to beneficial SCFAs production [134]. A probiotic compound of Streptococcus thermophiles, B. breve, B. longum, etc., also reportedly restored the expression of anti-inflammatory cytokine IL-10 completely without affecting pro-inflammatory mediators in mice following broad-spectrum antibiotic treatment. At the meantime, adaptive immunity was also restored, with increase of CD4+, CD8+, and B220+ cell numbers in the intestinal lamina propria [132]. Separate studies demonstrated that S. boulardii can up-regulate antitoxin A secretory IgA expression in animal models of AAD [135,136]. (3) Enhancing intestinal barrier function. A probiotic cocktail JUP-Y4 modulated ampicillin induced gut barrier dysfunction and GM disturbance in mice. Increased expression of intestinal epithelial tight-junction proteins, and reduced inflammatory cytokines in the ileum and the colon following JUP-Y4 use contributed to caecum tumefaction attenuation and a decrease in gut permeability [122]. Probiotics have also been shown to increase epithelium mucins production, which is a critical element of the epithelium barrier [137,138]. Probiotics also assist in producing antagonistic activity like bacteriocins against pathogenic bacteria, and inhibiting bacterial translocation by competing for receptors or adhesion to endothelial cells [139,140,141].
4. Conclusions and Future Perspectives
The foodborne contaminants, such as HMs, pesticides and ABs, cause harmful effects on animal and human health. GM is a major player in the remediation of these contaminants. Both contaminants-induced toxicity and impaired structure and metabolic activity of GM have significant impacts on target organs, causing tissue damage and other disease. Dietary supplementation with probiotics appears to be a promising adjunct intervention for effectively reducing the damage caused by foodborne contaminants and re-balancing the GM of humans and animals under a constant threat of pollutants.
The understanding of host-GM interactions must be further developed using a series of techniques such as metagenomics, metatranscriptomics and metabonomics, to provide meaningful insights into the mechanisms of GM, and to clarify the causal relationship between GM and GM-associated symptoms. Additionally, this work needs to be extended to human studies, as majority of research on contaminants remediation using probiotics comes from animal models, rather than humans. Meanwhile, almost all of current studies on the GM and contaminants solely rely on stool microbiota, which is part of the GM and may yield limited conclusions [146]. Hence gut mucosal sampling should also be considered in future studies. Furthermore, the colonization of probiotics in human may vary from person to person, depending on factors such as the composition of individual community, the composition of the colonizers, and intrinsic host factor [147]. Thus, in future applications with probiotics in human, personalized probiotic regimen based on the consumer at different contexts must be considered.
Author Contributions
P.L. and X.L. reviewed the manuscript; P.F. and Z.Y. wrote the manuscript; A.K. and A.K.V. prepared the figures and tables.
Acknowledgments
This work was supported by Fundamental Research Funds for the Central Universities grant (No: lzujbky-2017-br01) and Gansu province major science and technology projects (No: 17ZD2WA017). National Natural Science Foundation Grant (No: 31870082).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Watkinson, A.J.; Murby, E.J.; Kolpin, D.W.; Costanzo, S.D. The occurrence of antibiotics in an urban watershed: From wastewater to drinking water. Sci. Total Environ. 2009, 407, 2711–2723. [Google Scholar] [CrossRef]
- Pimentel, D. Amounts of pesticides reaching target pests: Environmental impacts and ethics. J. Agric. Environ. Ethics 1995, 8, 17–29. [Google Scholar] [CrossRef]
- Panagos, P.; Van Liedekerke, M.; Yigini, Y.; Montanarella, L. Contaminated Sites in Europe: Review of the Current Situation Based on Data Collected through a European Network. Int. J. Environ. Res. Public Health 2013, 2013, 158764. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Calvelo Pereira, R.; Camps-Arbestain, M.; Rodríguez Garrido, B.; Macías, F.; Monterroso, C. Behaviour of α-, β-, γ-, and δ-hexachlorocyclohexane in the soil–plant system of a contaminated site. Environ. Pollut. 2006, 144, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Sridhara Chary, N.; Kamala, C.T.; Samuel Suman Raj, D. Assessing risk of heavy metals from consuming food grown on sewage irrigated soils and food chain transfer. Ecotoxicol. Environ. Saf. 2008, 69, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Madani, F.Z.; Hafida, M.; Merzouk, S.A.; Loukidi, B.; Taouli, K.; Narce, M. Hemostatic, inflammatory, and oxidative markers in pesticide user farmers. Biomarkers 2016, 21, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Knoop, K.A.; McDonald, K.G.; Kulkarni, D.H.; Newberry, R.D. Antibiotics promote inflammation through the translocation of native commensal colonic bacteria. Gut 2016, 65, 1100–1109. [Google Scholar] [CrossRef]
- Narayana, K. An aminoglycoside antibiotic gentamycin induces oxidative stress, reduces antioxidant reserve and impairs spermatogenesis in rats. Toxicol. Sci. 2008, 33, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic Metals and Oxidative Stress Part I: Mechanisms Involved in Metal-induced Oxidative Damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef]
- Koner, B.C.; Banerjee, B.D.; Ray, A. Organochlorine pesticide-induced oxidative stress and immune suppression in rats. Indian J. Exp. Biol. 1998, 36, 395–398. [Google Scholar] [PubMed]
- Sekirov, I.; Tam, N.M.; Jogova, M.; Robertson, M.L.; Li, Y.; Lupp, C.; Finlay, B.B. Antibiotic-Induced Perturbations of the Intestinal Microbiota Alter Host Susceptibility to Enteric Infection. Infect. Immun. 2008, 76, 4726–4736. [Google Scholar] [CrossRef] [Green Version]
- Poet, T.S.; Wu, H.; Kousba, A.A.; Timchalk, C. In Vitro Rat Hepatic and Intestinal Metabolism of the Organophosphate Pesticides Chlorpyrifos and Diazinon. Toxicol. Sci. 2003, 72, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breton, J.; Massart, S.; Vandamme, P.; De Brandt, E.; Pot, B.; Foligné, B. Ecotoxicology inside the gut: Impact of heavy metals on the mouse microbiome. BMC Pharmacol. Toxicol. 2013, 14, 62. [Google Scholar] [CrossRef] [PubMed]
- Gavrilescu, M. Removal of heavy metals from the environment by biosorption. Eng. Life Sci. 2004, 4, 219–232. [Google Scholar] [CrossRef]
- Tangahu, B.V.; Sheikh Abdullah, S.R.; Basri, H.; Idris, M.; Anuar, N.; Mukhlisin, M. A Review on Heavy Metals (As, Pb, and Hg) Uptake by Plants through Phytoremediation. Int. J. Chem. Eng. 2011, 2011, 939161. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Prebiotics and probiotics; modifying and mining the microbiota. Pharmacol. Res. 2010, 61, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Wang, G.; Zhao, J.; Liu, X.; Tian, F.; Zhang, H.; Chen, W. Protective effects of Lactobacillus plantarum CCFM8610 against acute cadmium toxicity in mice. Appl. Environ. Microbiol. 2013, 79, 1508–1515. [Google Scholar] [CrossRef]
- Zhai, Q.; Wang, G.; Zhao, J.; Liu, X.; Narbad, A.; Chen, Y.Q.; Zhang, H.; Tian, F.; Chen, W. Protective Effects of Lactobacillus plantarum CCFM8610 against Chronic Cadmium Toxicity in Mice Indicate Routes of Protection besides Intestinal Sequestration. Appl. Environ. Microbiol. 2014, 80, 4063–4071. [Google Scholar] [CrossRef]
- Kamaladevi, A.; Ganguli, A.; Balamurugan, K. Lactobacillus casei stimulates phase-II detoxification system and rescues malathion-induced physiological impairments in Caenorhabditis elegans. Comp. Biochem. Physiol. C Pharmacol. Toxicol. 2016, 179, 19–28. [Google Scholar] [CrossRef]
- Trinder, M.; McDowell, T.W.; Daisley, B.A.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Probiotic Lactobacillus rhamnosus reduces organophosphate pesticide absorption and toxicity to Drosophila melanogaster. Appl. Environ. Microbiol. 2016, 82, 6204–6213. [Google Scholar] [CrossRef]
- Hempel, S.; Newberry, S.J.; Maher, A.R.; Wang, Z.; Miles, J.N.; Shanman, R.; Johnsen, B.; Shekelle, P.G. Probiotics for the prevention and treatment of antibiotic-associated diarrhea: A systematic review and meta-analysis. JAMA 2012, 307, 1959–1969. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the Human Intestinal Microbial Flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [Green Version]
- Breton, J.; Daniel, C.; Dewulf, J.; Pothion, S.; Froux, N.; Sauty, M.; Thomas, P.; Pot, B.; Foligné, B. Gut microbiota limits heavy metals burden caused by chronic oral exposure. Toxicol. Lett. 2013, 222, 132–138. [Google Scholar] [CrossRef]
- Claus, S.P.; Guillou, H.; Ellero-Simatos, S. The gut microbiota: A major player in the toxicity of environmental pollutants? npj Biofilms Microbiomes 2016, 2, 16003. [Google Scholar] [CrossRef]
- Wu, G.; Xiao, X.; Feng, P.; Xie, F.; Yu, Z.; Yuan, W.; Liu, P.; Li, X. Gut remediation: A potential approach to reducing chromium accumulation using Lactobacillus plantarum TW1-1. Sci. Rep. 2017, 7, 15000. [Google Scholar] [CrossRef]
- Zhang, S.; Jin, Y.; Zeng, Z.; Liu, Z.; Fu, Z. Subchronic exposure of mice to cadmium perturbs their hepatic energy metabolism and gut microbiome. Chem. Res. Toxicol. 2015, 28, 2000–2009. [Google Scholar] [CrossRef]
- Jin, Y.; Zeng, Z.; Wu, Y.; Zhang, S.; Fu, Z. Oral Exposure of Mice to Carbendazim Induces Hepatic Lipid Metabolism Disorder and Gut Microbiota Dysbiosis. Toxicol. Sci. 2015, 147, 116–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, J.L.; Kang, H.K.; Spivey, G. Arsenic levels in human blood, urine, and hair in response to exposure via drinking water. Environ. Res. 1979, 20, 24–32. [Google Scholar] [CrossRef]
- Wester, R.C.; Maibach, H.I.; Sedik, L.; Melendres, J.; Dizio, S.; Wade, M. In Vitro Percutaneous Absorption of Cadmium from Water and Soil into Human Skin. Toxicol. Sci. 1992, 19, 1–5. [Google Scholar] [CrossRef]
- Loubières, Y.; Lassence, A.D.; Bernier, M.; Vieillard-Baron, A.; Schmitt, J.-M.; Page, B.; Jardin, F. Acute, Fatal, Oral Chromic Acid Poisoning. J. Toxicol. Clin. Toxicol. 1999, 37, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Lu, L.; Jin, C.; Wang, S.; Zhou, J.; Ni, Y.; Fu, Z.; Jin, Y. Effects of short term lead exposure on gut microbiota and hepatic metabolism in adult zebrafish. Comp. Biochem. Physiol. C Pharmacol. Toxicol. 2018, 209, 1–8. [Google Scholar] [CrossRef]
- Zhai, Q.; Yu, L.; Li, T.; Zhu, J.; Zhang, C.; Zhao, J.; Zhang, H.; Chen, W. Effect of dietary probiotic supplementation on intestinal microbiota and physiological conditions of Nile tilapia (Oreochromis niloticus) under waterborne cadmium exposure. Antonie van Leeuwenhoek 2017, 110, 501–513. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Liu, K.; Shen, J. Exposing to Cadmium Stress Cause Profound Toxic Effect on Microbiota of the Mice Intestinal Tract. PLoS ONE 2014, 9, e85323. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, M.; Hassanzadeh, P.; Alaei, S. Cadmium chloride exhibits a profound toxic effect on bacterial microflora of the mice gastrointestinal tract. Hum. Exp. Toxicol. 2010, 30, 152–159. [Google Scholar] [CrossRef]
- Lu, K.; Abo, R.P.; Schlieper, K.A.; Graffam, M.E.; Levine, S.; Wishnok, J.S.; Swenberg, J.A.; Tannenbaum, S.R.; Fox, J.G. Arsenic exposure perturbs the gut microbiome and its metabolic profile in mice: An integrated metagenomics and metabolomics analysis. Environ. Health Perspect. 2014, 122, 284–291. [Google Scholar] [CrossRef]
- Wu, J.; Wen, X.W.; Faulk, C.; Boehnke, K.; Zhang, H.; Dolinoy, D.C.; Xi, C. Perinatal Lead Exposure Alters Gut Microbiota Composition and Results in Sex-specific Bodyweight Increases in Adult Mice. Toxicol. Sci. 2016, 151, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104, S1–S63. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, B.M.; Becker, A.; Ayiseh, R.B.; Hildebrand, F.; Raes, J.; Huys, G.; Vandenabeele, P. Gut microbiota affects sensitivity to acute DSS-induced colitis independently of host genotype. Inflamm. Bowel Dis. 2013, 19, 2560–2567. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Imhann, F.; Vich Vila, A.; Bonder, M.J.; Fu, J.; Gevers, D.; Visschedijk, M.C.; Spekhorst, L.M.; Alberts, R.; Franke, L.; van Dullemen, H.M.; et al. Interplay of host genetics and gut microbiota underlying the onset and clinical presentation of inflammatory bowel disease. Gut 2018, 67, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Cani, P.D. Interaction Between Obesity and the Gut Microbiota: Relevance in Nutrition. Annu. Rev. Nutr. 2011, 31, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Abdou, R.M.; Zhu, L.; Baker, R.D.; Baker, S.S. Gut Microbiota of Nonalcoholic Fatty Liver Disease. Dig. Dis. Sci. 2016, 61, 1268–1281. [Google Scholar] [CrossRef] [PubMed]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut Microbiota in Health and Disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Q.; Tian, F.; Zhao, J.; Zhang, H.; Narbad, A.; Chen, W. Oral administration of probiotics inhibits heavy metal cadmium absorption by protecting intestinal barrier. Appl. Environ. Microbiol. 2016, 82, 4429–4440. [Google Scholar] [CrossRef] [PubMed]
- Ba, Q.; Li, M.; Chen, P.; Huang, C.; Duan, X.; Lu, L.; Li, J.; Chu, R.; Xie, D.; Song, H.; et al. Sex-Dependent Effects of Cadmium Exposure in Early Life on Gut Microbiota and Fat Accumulation in Mice. Environ. Health Perspect. 2017, 125, 437–446. [Google Scholar] [CrossRef]
- Sarwar, M. The dangers of pesticides associated with public health and preventing of the risks. Int. J. Bioinform. Biomed. Eng. 2015, 1, 130–136. [Google Scholar]
- El-Gendy, K.S.; Aly, N.M.; Mahmoud, F.H.; Kenawy, A.; El-Sebae, A.K.H. The role of vitamin C as antioxidant in protection of oxidative stress induced by imidacloprid. Food Chem. Toxicol. 2010, 48, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Villarini, M.; Caldini, G.; Moretti, M.; Trotta, F.; Pasquini, R.; Cenci, G. Modulatory activity of a Lactobacillus casei strain on 1,2-dimethylhydrazine-induced genotoxicity in rats. Environ. Mol. Mutagen. 2008, 49, 192–199. [Google Scholar] [CrossRef]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Collins, S.L.; Sumarah, M.W.; Reid, G. Microbiota-mediated modulation of organophosphate insecticide toxicity by species-dependent lactobacilli interactions in a Drosophila melanogaster insect model. Appl. Environ. Microbiol. 2018, 84, e02820-17. [Google Scholar] [CrossRef] [PubMed]
- Harishankar, M.K.; Sasikala, C.; Ramya, M. Efficiency of the intestinal bacteria in the degradation of the toxic pesticide, chlorpyrifos. 3 Biotech 2013, 3, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Racke, K.D.; Bollag, J. Isolation and characterization of a chlorinated-pyridinol-degrading bacterium. Appl. Environ. Microbiol. 1997, 63, 4096–4098. [Google Scholar]
- Fang, B.; Li, J.W.; Zhang, M.; Ren, F.Z.; Pang, G.F. Chronic chlorpyrifos exposure elicits diet-specific effects on metabolism and the gut microbiome in rats. Food Chem. Toxicol. 2018, 111, 144–152. [Google Scholar] [CrossRef]
- Joly Condette, C.; Bach, V.; Mayeur, C.; Gay-Quéheillard, J.; Khorsi-Cauet, H. Chlorpyrifos Exposure During Perinatal Period Affects Intestinal Microbiota Associated With Delay of Maturation of Digestive Tract in Rats. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 30–40. [Google Scholar] [CrossRef]
- Joly, C.; Gay-Quéheillard, J.; Léké, A.; Chardon, K.; Delanaud, S.; Bach, V.; Khorsi-Cauet, H. Impact of chronic exposure to low doses of chlorpyrifos on the intestinal microbiota in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME®) and in the rat. Environ. Sci. Pollut. Res. 2013, 20, 2726–2734. [Google Scholar] [CrossRef]
- Reygner, J.; Joly Condette, C.; Bruneau, A.; Delanaud, S.; Rhazi, L.; Depeint, F.; Abdennebi-Najar, L.; Bach, V.; Mayeur, C.; Khorsi-Cauet, H. Changes in Composition and Function of Human Intestinal Microbiota Exposed to Chlorpyrifos in Oil as Assessed by the SHIME® Model. Int. J. Environ. Res. Public Health 2016, 13, 1088. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, Y.; Wang, G.; Han, R.; Xie, X. Effects of chlorpyrifos on the gut microbiome and urine metabolome in mouse (Mus musculus). Chemosphere 2016, 153, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hylemon, P.B.; Zhou, H.; Pandak, W.M.; Ren, S.; Gil, G.; Dent, P. Bile acids as regulatory molecules. J. Lipid Res. 2009, 50, 1509–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.; Chi, L.; Tu, P.; Bian, X.; Thomas, J.; Ru, H.; Lu, K. The organophosphate malathion disturbs gut microbiome development and the quorum-Sensing system. Toxicol. Lett. 2018, 283, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Bian, X.; Mahbub, R.; Lu, K. Sex-Specific Effects of Organophosphate Diazinon on the Gut Microbiome and Its Metabolic Functions. Environ. Health Perspect. 2017, 125, 198–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shehata, A.A.; Schrödl, W.; Aldin, A.A.; Hafez, H.M.; Krüger, M. The Effect of Glyphosate on Potential Pathogens and Beneficial Members of Poultry Microbiota In Vitro. Curr. Microbiol. 2013, 66, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Shao, W.; Zhang, C.; Xu, C.; Wang, Q.; Liu, H.; Sun, H.; Jiang, Z.; Gu, A. Organochloride pesticides modulated gut microbiota and influenced bile acid metabolism in mice. Environ. Pollut. 2017, 226, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Nasuti, C.; Coman, M.M.; Olek, R.A.; Fiorini, D.; Verdenelli, M.C.; Cecchini, C.; Silvi, S.; Fedeli, D.; Gabbianelli, R. Changes on fecal microbiota in rats exposed to permethrin during postnatal development. Environ. Sci. Pollut. Res. 2016, 23, 10930–10937. [Google Scholar] [CrossRef]
- Kan, H.; Zhao, F.; Zhang, X.-X.; Ren, H.; Gao, S. Correlations of Gut Microbial Community Shift with Hepatic Damage and Growth Inhibition of Carassius auratus Induced by Pentachlorophenol Exposure. Environ. Sci. Technol. 2015, 49, 11894–11902. [Google Scholar] [CrossRef]
- Jin, C.; Zeng, Z.; Fu, Z.; Jin, Y. Oral imazalil exposure induces gut microbiota dysbiosis and colonic inflammation in mice. Chemosphere 2016, 160, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Luo, T.; Zhu, Z.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z.; Jin, Y. Imazalil exposure induces gut microbiota dysbiosis and hepatic metabolism disorder in zebrafish. Comp. Biochem. Phys. C 2017, 202, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Brüssow, H. Growth promotion and gut microbiota: Insights from antibiotic use. Environ. Microbiol. 2015, 17, 2216–2227. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Liu, W. Occurrence, fate, and ecotoxicity of antibiotics in agro-ecosystems. A review. Agron. Sustain. Dev. 2012, 32, 309–327. [Google Scholar] [CrossRef]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B.A. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef]
- Dong, H.; Yuan, X.; Wang, W.; Qiang, Z. Occurrence and removal of antibiotics in ecological and conventional wastewater treatment processes: A field study. J. Environ. Manag. 2016, 178, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Qian, M.; Wu, H.; Wang, J.; Zhang, H.; Zhang, Z.; Zhang, Y.; Lin, H.; Ma, J. Occurrence of trace elements and antibiotics in manure-based fertilizers from the Zhejiang Province of China. Sci. Total Environ. 2016, 559, 174–181. [Google Scholar] [CrossRef]
- Demoly, P.; Benahmed, S.; Valembois, M.; Sahla, H.; Messaad, D.; Godard, P.; Michel, F.B.; Bousquet, J. Allergy to macrolide antibiotics. Review of the literature. Presse Med. 2000, 29, 321–326. [Google Scholar]
- Schubert, A.M.; Sinani, H.; Schloss, P.D. Antibiotic-induced alterations of the murine gut microbiota and subsequent effects on colonization resistance against Clostridium difficile. mBio 2015, 6, e00974-15. [Google Scholar] [CrossRef]
- Buffie, C.G.; Jarchum, I.; Equinda, M.; Lipuma, L.; Gobourne, A.; Viale, A.; Ubeda, C.; Xavier, J.; Pamer, E.G. Profound alterations of intestinal microbiota following a single dose of clindamycin results in sustained susceptibility to Clostridium difficile-induced colitis. Infect. Immun. 2012, 80, 62–73. [Google Scholar] [CrossRef]
- Grazul, H.; Kanda, L.L.; Gondek, D. Impact of probiotic supplements on microbiome diversity following antibiotic treatment of mice. Gut Microbes 2016, 7, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed]
- Buffie, C.G.; Bucci, V.; Stein, R.R.; McKenney, P.T.; Ling, L.; Gobourne, A.; No, D.; Liu, H.; Kinnebrew, M.; Viale, A.; et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2014, 517, 205–208. [Google Scholar] [CrossRef]
- Fouhy, F.; Guinane, C.M.; Hussey, S.; Wall, R.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C.; et al. High-Throughput Sequencing Reveals the Incomplete, Short-Term Recovery of Infant Gut Microbiota following Parenteral Antibiotic Treatment with Ampicillin and Gentamicin. Antimicrob. Agents Chemother. 2012, 56, 5811–5820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korpela, K.; Salonen, A.; Virta, L.J.; Kekkonen, R.A.; Forslund, K.; Bork, P.; De Vos, W.M. Intestinal microbiome is related to lifetime antibiotic use in Finnish pre-school children. Nat. Commun. 2016, 7, 10410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rea, M.C.; Dobson, A.; O’Sullivan, O.; Crispie, F.; Fouhy, F.; Cotter, P.D.; Shanahan, F.; Kiely, B.; Hill, C.; Ross, R.P. Effect of broad-and narrow-spectrum antimicrobials on Clostridium difficile and microbial diversity in a model of the distal colon. Proc. Natl. Acad. Sci. USA 2011, 108, 4639–4644. [Google Scholar] [CrossRef] [PubMed]
- Pien, F.D.; Shrum, S.; Swenson, J.M.; Hill, B.C.; Thornsberry, C.; Farmer, J.J. Colonization of human wounds by Escherichia vulneris and Escherichia hermannii. J. Clin. Microbiol. 1985, 22, 283–285. [Google Scholar]
- Chaudhury, A.; Nath, G.; Tikoo, A.; Sanyal, S.C. Enteropathogenicity and Antimicrobial Susceptibility of New Escherichia Spp. J. Diarrhoeal Dis. Res. 1999, 17, 85–87. [Google Scholar]
- Jantsch, J.; Chikkaballi, D.; Hensel, M. Cellular aspects of immunity to intracellular Salmonella enterica. Immunol. Rev. 2011, 240, 185–195. [Google Scholar] [CrossRef]
- Reeves, A.E.; Theriot, C.M.; Bergin, I.L.; Huffnagle, G.B.; Schloss, P.D.; Young, V.B. The interplay between microbiome dynamics and pathogen dynamics in a murine model of Clostridium difficile Infection. Gut Microbes 2011, 2, 145–158. [Google Scholar] [CrossRef]
- Schubert, A.M.; Rogers, M.A.; Ring, C.; Mogle, J.; Petrosino, J.P.; Young, V.B.; Aronoff, D.M.; Schloss, P.D. Microbiome data distinguish patients with Clostridium difficile infection and non-C. difficile-associated diarrhea from healthy controls. mBio 2014, 5, e01021-14. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C.; Stephens, D.A.; Loo, V.G.; Edens, T.J.; Behr, M.A.; Dewar, K.; Manges, A.R. Reductions in intestinal Clostridiales precede the development of nosocomial Clostridium difficile infection. Microbiome 2013, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Yamanishi, S.; Cox, L.; Methe, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobel, Y.R.; Cox, L.M.; Kirigin, F.F.; Bokulich, N.A.; Yamanishi, S.; Teitler, I.; Chung, J.; Sohn, J.; Barber, C.M.; Goldfarb, D.S.; et al. Metabolic and metagenomic outcomes from early-life pulsed antibiotic treatment. Nat. Commun. 2015, 6, 7486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, K.; Pi, Y.; Mu, C.-L.; Peng, Y.; Huang, Z.; Zhu, W.-Y. Antibiotics-induced modulation of large intestinal microbiota altered aromatic amino acid profile and expression of neurotransmitters in the hypothalamus of piglets. J. Neurochem. 2018, 146, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, Q.; Huan, F.; Qu, J.; Liu, W.; Gu, A.; Wang, Y.; Jiang, Z. Changes in Gut Microbiota May Be Early Signs of Liver Toxicity Induced by Epoxiconazole in Rats. Chemotherapy 2014, 60, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Tuomola, E.; Crittenden, R.; Playne, M.; Isolauri, E.; Salminen, S. Quality assurance criteria for probiotic bacteria. Am. J. Clin. Nutr. 2001, 73, 393s–398s. [Google Scholar] [CrossRef] [PubMed]
- Chiocchetti, G.M.; Jadán-Piedra, C.; Monedero, V.; Zúñiga, M.; Vélez, D.; Devesa, V. Use of lactic acid bacteria and yeasts to reduce exposure to chemical food contaminants and toxicity. Crit. Rev. Food Sci. Nutr. 2018, 1–12. [Google Scholar] [CrossRef]
- Monachese, M.; Burton, J.P.; Reid, G. Bioremediation and Tolerance of Humans to Heavy Metals through Microbial Processes: A Potential Role for Probiotics? Appl. Environ. Microbiol. 2012, 78, 6397–6404. [Google Scholar] [CrossRef]
- Raghuvanshi, R.; Chaudhari, A.; Kumar, G.N. Amelioration of cadmium- and mercury-induced liver and kidney damage in rats by genetically engineered probiotic Escherichia coli Nissle 1917 producing pyrroloquinoline quinone with oral supplementation of citric acid. Nutrition 2016, 32, 1285–1294. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Khatun, S.; Maity, M.; Jana, S.; Perveen, H.; Dash, M.; Dey, A.; Jana, L.R.; Maity, P.P. Association of Vitamin B12, Lactate Dehydrogenase, and Regulation of NF-κB in the Mitigation of Sodium Arsenite-Induced ROS Generation in Uterine Tissue by Commercially Available Probiotics. Probiotics Antimicrob. Proteins 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ojekunle, O.; Banwo, K.; Sanni, A.I. In vitro and in vivo evaluation of Weissella cibaria and Lactobacillus plantarum for their protective effect against cadmium and lead toxicities. Lett. Appl. Microbiol. 2017, 64, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, M.; Nordberg, G.F. Toxicological aspects of metallothionein. Cell. Mol. Biol. (Noisy-le-Grand France) 2000, 46, 451–463. [Google Scholar]
- Randazzo, C.L.; Pino, A.; Ricciardi, L.; Romano, C.; Comito, D.; Arena, E.; Saitta, S.; Caggia, C. Probiotic supplementation in systemic nickel allergy syndrome patients: Study of its effects on lactic acid bacteria population and on clinical symptoms. J. Appl. Microbiol. 2014, 118, 202–211. [Google Scholar] [CrossRef]
- Bisanz, J.E.; Enos, M.K.; Mwanga, J.R.; Changalucha, J.; Burton, J.P.; Gloor, G.B.; Reid, G. Randomized open-label pilot study of the influence of probiotics and the gut microbiome on toxic metal levels in Tanzanian pregnant women and school children. mBio 2014, 5, e01580-14. [Google Scholar] [CrossRef] [PubMed]
- Masson, P. Evolution of and perspectives on therapeutic approaches to nerve agent poisoning. Toxicol. Lett. 2011, 206, 5–13. [Google Scholar] [CrossRef]
- Bouhafs, L.; Moudilou, E.N.; Exbrayat, J.M.; Lahouel, M.; Idoui, T. Protective effects of probiotic Lactobacillus plantarum BJ0021 on liver and kidney oxidative stress and apoptosis induced by endosulfan in pregnant rats. Ren. Fail. 2015, 37, 1370–1378. [Google Scholar] [CrossRef]
- Cao, L.; Yang, X.J.; Li, Z.J.; Sun, F.F.; Wu, X.H.; Yao, J.H. Reduced lesions in chickens with Clostridium perfringens-induced necrotic enteritis by Lactobacillus fermentum 1.202912. Poult. Sci. 2012, 91, 3065–3071. [Google Scholar] [CrossRef]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Park, Z.; McCann, M.J.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef]
- Russell, R.J.; Scott, C.; Jackson, C.J.; Pandey, R.; Pandey, G.; Taylor, M.C.; Coppin, C.W.; Liu, J.-W.; Oakeshott, J.G. The evolution of new enzyme function: Lessons from xenobiotic metabolizing bacteria versus insecticide-resistant insects. Evol. Appl. 2011, 4, 225–248. [Google Scholar] [CrossRef]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Welle, H.; Dube, J.S.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Neonicotinoid-induced pathogen susceptibility is mitigated by Lactobacillus plantarum immune stimulation in a Drosophila melanogaster model. Sci. Rep. 2017, 7, 2703. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, I.; Sybesma, W.; Phothirath, P.; Ananta, E.; Mercenier, A. Application of probiotics in food products—Challenges and new approaches. Curr. Opin Biotechnol. 2010, 21, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Rijkers, G.T.; Bengmark, S.; Enck, P.; Haller, D.; Herz, U.; Kalliomaki, M.; Kudo, S.; Lenoir-Wijnkoop, I.; Mercenier, A.; Myllyluoma, E.; et al. Guidance for Substantiating the Evidence for Beneficial Effects of Probiotics: Current Status and Recommendations for Future Research. J. Nutr. 2010, 140, 671S–676S. [Google Scholar] [CrossRef] [Green Version]
- Conway, S.; Hart, A.; Clark, A.; Harvey, I. Does eating yogurt prevent antibiotic-associated diarrhoea?: A placebo-controlled randomised controlled trial in general practice. Br. J. Gen. Pract. 2007, 57, 953–959. [Google Scholar] [CrossRef]
- Allen, S.J.; Wareham, K.; Wang, D.; Bradley, C.; Hutchings, H.; Harris, W.; Dhar, A.; Brown, H.; Foden, A.; Gravenor, M.B. Lactobacilli and bifidobacteria in the prevention of antibiotic-associated diarrhoea and Clostridium difficile diarrhoea in older inpatients (PLACIDE): A randomised, double-blind, placebo-controlled, multicentre trial. Lancet 2013, 382, 1249–1257. [Google Scholar] [CrossRef]
- Kotowska, M.; Albrecht, P.; Szajewska, H. Saccharomyces boulardii in the prevention of antibiotic-associated diarrhoea in children: A randomized double-blind placebo-controlled trial. Aliment. Pharmacol. Ther. 2005, 21, 583–590. [Google Scholar] [CrossRef]
- McFarland, L.V. Systematic review and meta-analysis of Saccharomyces boulardii in adult patients. World J. Gastroenterol. 2010, 16, 2202–2222. [Google Scholar] [CrossRef]
- Szajewska, H.; Kołodziej, M. Systematic review with meta-analysis: Lactobacillus rhamnosus GG in the prevention of antibiotic-associated diarrhoea in children and adults. Aliment. Pharmacol. Ther. 2015, 42, 1149–1157. [Google Scholar] [CrossRef]
- Beniwal, R.S.; Arena, V.C.; Thomas, L.; Narla, S.; Imperiale, T.F.; Chaudhry, R.A.; Ahmad, U.A. A Randomized Trial of Yogurt for Prevention of Antibiotic-Associated Diarrhea. Dig. Dis. Sci. 2003, 48, 2077–2082. [Google Scholar] [CrossRef]
- McFarland, L.V. Meta-Analysis of Probiotics for the Prevention of Antibiotic Associated Diarrhea and the Treatment of Clostridium difficile Disease. Am. J. Gastroenterol. 2006, 101, 812–822. [Google Scholar] [CrossRef]
- Schmidt, V.; Gomez-Chiarri, M.; Roy, C.; Smith, K.; Amaral-Zettler, L. Subtle Microbiome Manipulation Using Probiotics Reduces Antibiotic-Associated Mortality in Fish. mSystems 2017, 2, e00133-17. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhao, X.; Zhao, J.; Zhang, H.; Zhai, Q.; Narbad, A.; Chen, W. A mixture of Lactobacillus species isolated from traditional fermented foods promote recovery from antibiotic-induced intestinal disruption in mice. J. Appl. Microbiol. 2018, 124, 842–854. [Google Scholar] [CrossRef] [PubMed]
- Chiodini, R.J.; Dowd, S.E.; Chamberlin, W.M.; Galandiuk, S.; Davis, B.; Glassing, A. Microbial Population Differentials between Mucosal and Submucosal Intestinal Tissues in Advanced Crohn’s Disease of the Ileum. PLoS ONE 2015, 10, e0134382. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Loening-Baucke, V.; Vaneechoutte, M.; Doerffel, Y. Active Crohn’s disease and ulcerative colitis can be specifically diagnosed and monitored based on the biostructure of the fecal flora. Inflamm. Bowel Dis. 2008, 14, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Day, A.S.; Huinao, K.D.; Leach, S.T.; Lemberg, D.A.; Dowd, S.E.; Mitchell, H.M. Microbial dysbiosis in pediatric patients with Crohn’s disease. J. Clin. Microbiol. 2012, 50, 3258–3265. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Dörffel, Y.; Loening-Baucke, V.; Theissig, F.; Rückert, J.C.; Ismail, M.; Rau, W.A.; Gaschler, D.; Weizenegger, M.; Kühn, S.; et al. Acute appendicitis is characterised by local invasion with Fusobacterium nucleatum/necrophorum. Gut 2011, 60, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Png, C.W.; Lindén, S.K.; Gilshenan, K.S.; Zoetendal, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.A.; Florin, T.H.J. Mucolytic Bacteria With Increased Prevalence in IBD Mucosa Augment In Vitro Utilization of Mucin by Other Bacteria. Am. J. Gastroenterol. 2010, 105, 2420–2428. [Google Scholar] [CrossRef]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef]
- Schmidt, V.T.; Reveillaud, J.; Zettler, E.; Mincer, T.J.; Murphy, L.; Amaral-Zettler, L.A. Oligotyping reveals community level habitat selection within the genus Vibrio. Front. Microbiol. 2014, 5, 563. [Google Scholar] [CrossRef]
- De Schryver, P.; Vadstein, O. Ecological theory as a foundation to control pathogenic invasion in aquaculture. ISME J. 2014, 8, 2360–2368. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Ai, C.; Wang, G.; Liu, X.; Tian, F.; Zhao, J.; Zhang, H.; Chen, Y.Q.; Chen, W. Oral application of lactic acid bacteria following treatment with antibiotics inhibits allergic airway inflammation. J. Appl. Microbiol. 2015, 119, 809–817. [Google Scholar] [CrossRef]
- Ekmekciu, I.; von Klitzing, E.; Fiebiger, U.; Neumann, C.; Bacher, P.; Scheffold, A.; Bereswill, S.; Heimesaat, M.M. The probiotic compound VSL#3 modulates mucosal, peripheral, and systemic immunity following murine broad-spectrum antibiotic treatment. Front. Cell. Infect. Microbiol. 2017, 7, 167. [Google Scholar] [CrossRef]
- Jijon, H.; Backer, J.; Diaz, H.; Yeung, H.; Thiel, D.; McKaigney, C.; De Simone, C.; Madsen, K. DNA from probiotic bacteria modulates murine and human epithelial and immune function. Gastroenterology 2004, 126, 1358–1373. [Google Scholar] [CrossRef]
- Shi, Y.; Zhai, Q.; Li, D.; Mao, B.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Restoration of cefixime-induced gut microbiota changes by Lactobacillus cocktails and fructooligosaccharides in a mouse model. Microbiol. Res. 2017, 200, 14–24. [Google Scholar] [CrossRef]
- Buts, J.-P.; Keyser, N.D.; Raedemaeker, L.D. Saccharomyces boulardii Enhances Rat Intestinal Enzyme Expression by Endoluminal Release of Polyamines. Pediatr. Res. 1994, 36, 522–527. [Google Scholar] [CrossRef]
- Qamar, A.; Aboudola, S.; Warny, M.; Michetti, P.; Pothoulakis, C.; LaMont, J.T.; Kelly, C.P. Saccharomyces boulardii stimulates intestinal immunoglobulin a immune response to clostridium difficiletoxin a in mice. Infect. Immun. 2001, 69, 2762–2765. [Google Scholar] [CrossRef]
- Mack, D.R.; Michail, S.; Wei, S.; McDougall, L.; Hollingsworth, M.A. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. 1999, 276, G941–G950. [Google Scholar] [CrossRef]
- Willemsen, L.E.M.; Koetsier, M.A.; van Deventer, S.J.H.; van Tol, E.A.F. Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E1 and E2 production by intestinal myofibroblasts. Gut 2003, 52, 1442–1447. [Google Scholar] [CrossRef] [Green Version]
- Gopal, P.K.; Prasad, J.; Smart, J.; Gill, H.S. In vitro adherence properties of Lactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and their antagonistic activity against an enterotoxigenic Escherichia coli. Int. J. Food. Microbiol. 2001, 67, 207–216. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef] [Green Version]
- Balcázar, J.L.; Vendrell, D.; de Blas, I.; Ruiz-Zarzuela, I.; Gironés, O.; Múzquiz, J.L. In vitro competitive adhesion and production of antagonistic compounds by lactic acid bacteria against fish pathogens. Vet. Microbiol. 2007, 122, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jama, A.M.; Mitić-Ćulafić, D.; Kolarević, S.; Đurašević, S.F.; Knežević-Vukčević, J. Protective effect of probiotic bacteria against cadmium-induced genotoxicity in rat hepatocytes in vivo and in vitro. Arch. Biol. Sci. 2012, 64, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Djurasevic, S.; Jama, A.; Jasnic, N.; Vujovic, P.; Jovanovic, M.; Mitic-Culafic, D.; Knezevic-Vukcevic, J.; Cakic-Milosevic, M.; Ilijevic, K.; Djordjevic, J. The protective effects of probiotic bacteria on cadmium toxicity in rats. J. Med. Food 2016, 20, 189–196. [Google Scholar] [CrossRef]
- Majlesi, M.; Shekarforoush, S.S.; Ghaisari, H.R.; Nazifi, S.; Sajedianfard, J.; Eskandari, M.H. Effect of probiotic Bacillus coagulans and Lactobacillus plantarum on alleviation of mercury toxicity in rat. Probiotics Antimicrob. Proteins 2017, 9, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Jin, D.; Yu, S.; Etareri Evivie, S.; Muhammad, Z.; Huo, G.; Liu, F. In Vitro and In Vivo Evaluation of Lactobacillus delbrueckii subsp. bulgaricus KLDS1.0207 for the Alleviative Effect on Lead Toxicity. Nutrients 2017, 9, 845. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.; et al. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell 2018, 174, 1388.e21–1405.e21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Chow, K.-H.; Fleming, E.; Oh, J. Selective colonization ability of human fecal microbes in different mouse gut environments. ISME J. 2018. [Google Scholar] [CrossRef]
Figure 1.
Proposed protective mechanisms of probiotics on HMs remediation in vivo.

Figure 2.
Schematic representation of proposed mechanisms of probiotic action on antibiotics associated diarrhea (AAD).
Figure 2.
Schematic representation of proposed mechanisms of probiotic action on antibiotics associated diarrhea (AAD).


{kind=link}
{kind=link}
Table 1.
Recent studies on the effects of foodborne contaminants on hosts and GM.
| Type | References | Models | Pollutants and Dosage | Outcomes | Main Conclusion on GM |
|---|---|---|---|---|---|
| HMs | [30] | Mice | Cd at 10 mg/L for 10 weeks | Hepatic inflammation, energy metabolism dysregulation | Firmicutes↓, Bacteroidetes↑, γ- Proteobacteria↓ |
| [41] | Mice | Pb at 32 ppm for 2 weeks | Bodyweight ↑ | Firmicutes/Bacteroidetes↑, Desulfovibrionaceae↑, Barnesiella↑, Clostridium XIVb↑, Lactococcus↓, Enterorhabdus↓, Caulobacterales↓ | |
| [40] | Mice | As at 10 ppm for 4 weeks | Perturbed lipid metabolites, indole-containing metabolites, isoflavone metabolites, and bile acid metabolites | Firmicutes↓, Bacteroidetes↑ | |
| [29] | Mice | Cr (VI) at 2 mM for 7 weeks | Oxidative stress↑, liver damage, GM disturbance | Bacteroidetes↑, Tenericutes↑, Firmicutes↓, Paraprevotellaceae↑, S24-7↑, Lachnospiraceae↓ | |
| Pesticides | [58,60] | Rats | Chlorpyrifos at 0.3 or 3.0 mg/kg bodyweight/day for 9 weeks | Obese and diabetic phenotypes↑, bacterial translocation↑ | Sutterella↑, Candidatus arthromitus↓, Olsenella↑ Clostridium sensu stricto 1↑, Amphibacillus↑, Enterorhabdus↑, Alloprevotella↑ |
| [65] | Mice | Malathion at 2 mg/L in drinking water (∼0.6 mg/kg bodyweight/ day) for 13 weeks | Motility and pathogenicity↑ | Corynebacterium↑, S24-7↑, Planococcaceae↓, Christenseneellaceae↓, Clostridium↑, Lachnospiraceae_Other↓, Anaerostipes↓, Blautia↓, Dorea↓, Roseburia↓, Mogibacteriaceae↑, Akkermansia↓, | |
| [66] | Mice | Diazinon at 4 mg/L for 13 weeks | Taurine level↑, glycine acetyltransferase and threonine dehydrogenase↓ in male mice | Bacteroidaceae_Bacteroides↑, Burkholderiales_Other↑, Clostridiaceae_Other↑, Erysipelotrichaceae_Coprobacillus↑, Lachnospiraceae_Butyrivibrio↓, Lachnospiraceae_Shuttleworthia↓, Staphylococcaceae_Staphylococcus ↓ | |
| [68] | Mice | p,p’-dichlorodiphenyldichloroethylene and β-hexachlorocyclohexane at 1 and 10 mg/kg body weight/day, for 8 weeks, respectively | Bile acid reabsorption in the terminal ileum and compensatory↓, bile acid and hydrophobicity↑, the genes expression on synthesis of bile acids in the liver↑ | Firmicutes↑, Proteobacteria↑, Bacteroidetes↓, Verrucomicrobia↓, Actinobacteria↓ | |
| [70] | Gold Fish | Pentachlorophenol at 0, 10, 50, and 100 μg/L for 28 days | Body weight and liver weight↓, oxidative stress↑, liver damage↑ | Bacteroidetes↑, Firmicutes↓, Bacteroides↑, Chryseobacterium↓, Microbacterium↓, Arthrobacter↓, Legionella↓ | |
| ABs | [72] | Zebrafish | Imazalil at 100 and 1000 μg/L for 1, 7 and 21 days | Glucokinase↓, hexokinase 1↓, pyruvate kinase↓, cytosolic Phosphoenol pyruvate carboxykinase (Pepckc) in liver ↓ | Bacteroidetes↓, Firmicutes↑ |
| [71] | Mice | Imazalil at 25, 50 or 100 mg/kg body weight daily for 4 weeks | Genes related to glycolysis and lipid metabolism↓ | Lactobacillus↓, Bifidobacterium↓ Deltaproteobacteria↑, Desulfovibrio↑ | |
| [96] | Rats | Epoxiconazole at 4 and 100 mg/kg body weight/day for 90 days | Weight of the liver and kidney↑, total bilirubin and cholinesterase in serum↓, blood glucose↑ | Firmicutes↓, Bacteroidetes↑, Proteobacteria↑, Lactobacillaceae↓, Bacteroidaceae↑, Enterobacteriaceae↑, Lachnospiraceae↑ | |
| [81] | Mice | The mixture of ampicillin, streptomycin, and clindamycin at 1 mg/mL for 2–4 week | The ceca size↑, a deeper shade of brown in ceca | Microbial diversity↓, Bacteroidetes↓, Stenotrophomonas↑, Xanthamonas↑ | |
| [95] | Piglets | The mixture of ampicillin, gentamicin, and metronidazole at 150, 4, and 30 mg/kg/day, respectively, for 25 days | Neurotransmitters in blood and hypothalamus↓, amino acids in feces, blood and hypothalamus↓ | Microbial diversity in feces↓, Firmicutes↑, Actinobacteria↑, Streptococcus↑, Lactobacillus↑, Bifidobacterium↑, Blautia↑, Klebsiella↑, Euryarchaeota↓, Spirochaetes↓, Tenericutes↓, Ruminococcus↓, Clostridium↓, unclassified Clostridiales↓, Christensenella↓, Methanobrevibacter↓, Prevotella↓ |
↑: Increase of relative abundance of the species or the severity of the outcomes; ↓: Decrease of relative abundance of the species or the severity of the outcomes.
Table 2.
Recent studies on the protective effects of probiotics against foodborne contaminants toxicity.
Table 2.
Recent studies on the protective effects of probiotics against foodborne contaminants toxicity.
| Type | References | Models | Contaminants Dosage | Supplementation Dosage | Main Conclusion |
|---|---|---|---|---|---|
| HMs | [142,143] | Rats | Cd | CdCl2 at 70 ppm, the mixture of L. acidophilus Rosell-52, L. rhamnosus Rosell-11 and B. longum Rosell-175 (5 × 108 CFU/g food) for 5 weeks | Marked decrease genotoxicity and the toxicity to lactobacilli, promoted Cd excretion in feces; decreased Cd in body; relieved liver and kidney damage, increased the number of L. acidophilus in feces |
| [144] | Rats | Hg | A total of 0.5 mL HgCl2 at 20 μg/mL and 1ml B. coagulans and L. plantarum CNR273 (109 CFU/mL) daily for 48 days | Marked increase Hg excretion in feces; reduce Hg levels in liver and kidney; prevent oxidative stress; reduce liver and kidney damage; increase the number of fecal LAB and the total bacteria counts | |
| [145] | Mice | Pb | A total of 2 mg (CH3COO)2Pb·3H2O in 0.4 mL plain water, L. bulgaricus KLDS1.0207 1 × 1010 (high dose), 1 × 109 (medial dose) and 1 × 108 (low dose) CFU/mL in 0.4 mL skim milk | Lower mortality rates, increased Pb excretion in feces, decreased tissue Pb enrichment, improved the antioxidant in the liver and kidney, and relieved renal pathological damage | |
| [101] | Rats | As | NaAsO2 at 1.0 mg/100 g body weight, the mixture of L. acidophilus, L. rhamnosus, B. longum, and S. boulardii at 0.25 mg/100 g body weight for 16 days | Reduction of oxidative stress, inflammation in uterine, protection against mutagenic uterine DNA-breakage, necrosis, ovarian-uterine tissue damages | |
| [29] | Mice | Cr (VI) | A total of 1mM K2Cr2O7 in drinking water, L. plantarum TW1-1 (1 × 109 CFU/once every other day) for 7 weeks | Promoted Cr excretion in feces, reduced Cr accumulation in tissues; decreased oxidative stress and damage in liver; partially restored the GM community | |
| Pesticides | [107] | Rats | Endosulfan | Endosulfan at 4 mg/kg bodyweight from the 6th to 20th day of gestation, L. plantarum BJ0021 0.1 mL per os and one hour before the administration endosulfan | Significantly reduced the cholesterol level and marked depletion of hepatic enzymes, decreased the number of apoptotic nuclei in kidney |
| [20] | Caenorhabditis elegans | Malathion | Exposure to malathion at 300 mM for 4 h at 20 °C after administration L. casei liquid cultures of 0.1 OD at 600 nm for 4 h | Reproduction protection with increase of rate of egg laying and brood size, and rescued locomotion of C. elegans | |
| [21] | Drosophila melanogaster | Chloropyrifos parathion | Co-exposure 10 μM chloropyrifos parathion and 100 μL L. rhamnosus GG (109 CFU) for 12 days | Prolonged overall survival and decreased early deaths | |
| ABs | [81] | Mice | Different ABs | Ampicillin, Streptomycin, and Clindamycin at 1 mg/mL, A cocktails of L. rhamnosus A191, L. acidophilus, B. breve, B. longum (4 × 109/mL) at 0.1 mL/mouse for 2 weeks | Lead a rise in microbial diversity; small increase in Firmicutes, increase in Enterobacteriaceae, and a bloom of Anaerotruncus, decrease in Xanthamonas |
| [121] | Fish | Streptomycin sulfate | A total of 200 g/mL of streptomycin sulfate daily for 13 days, 1 × 105 CFU/mL P. inhibens S4Sm and B. pumilus RI06-95Sm daily for 5 days following ABs treatment | Probiotics can colonize fish microbiome, decrease mortality in fish with subtle GM changes | |
| [122] | Mice | Ampicillin | Ampicillin (500 mg/kg) twice-daily for 14 days, a cocktail of L. plantarum, L. casei, L. rhamnosus and L. helveticus (2 × 109 CFU/0.2 mL/dose) for 4 weeks | Restore diversity of GM, decrease Firmicutes, reduce Desulfovibrionales, Dorea, Ruminococcus, Clostridia and Helicobacter, enrich Akkermansia, Alistipes and Porphyromonadaceae |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Feng, P.; Ye, Z.; Kakade, A.; Virk, A.K.; Li, X.; Liu, P. A Review on Gut Remediation of Selected Environmental Contaminants: Possible Roles of Probiotics and Gut Microbiota. Nutrients 2019, 11, 22. https://doi.org/10.3390/nu11010022
AMA Style
Feng P, Ye Z, Kakade A, Virk AK, Li X, Liu P. A Review on Gut Remediation of Selected Environmental Contaminants: Possible Roles of Probiotics and Gut Microbiota. Nutrients. 2019; 11(1):22. https://doi.org/10.3390/nu11010022
Chicago/Turabian StyleFeng, Pengya, Ze Ye, Apurva Kakade, Amanpreet Kaur Virk, Xiangkai Li, and Pu Liu. 2019. "A Review on Gut Remediation of Selected Environmental Contaminants: Possible Roles of Probiotics and Gut Microbiota" Nutrients 11, no. 1: 22. https://doi.org/10.3390/nu11010022
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.