Leucine Supplementation Does Not Attenuate Skeletal Muscle Loss during Leg Immobilization in Healthy, Young Men

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Subjects
2.2. Pretesting/Screening
2.3. Experimental Outline
2.4. Measurements
2.5. Limb Immobilization Protocol
2.6. Leucine Supplementation
2.7. Dietary Intake and Physical Activity Records
2.8. Blood Analyses
2.9. Muscle Analyses
2.10. Statistics
3. Results
3.1. Subjects
3.2. Dietary Intake
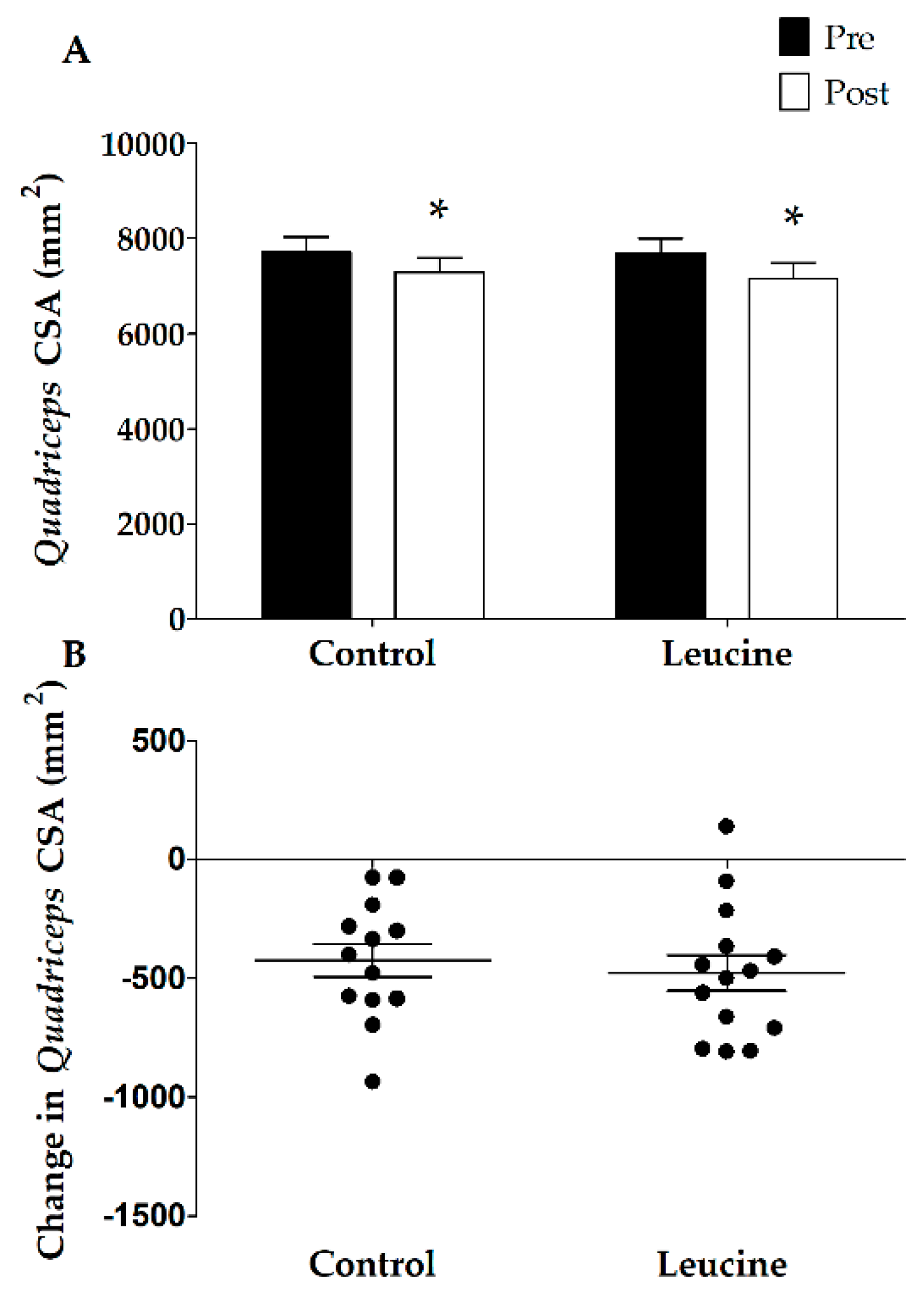
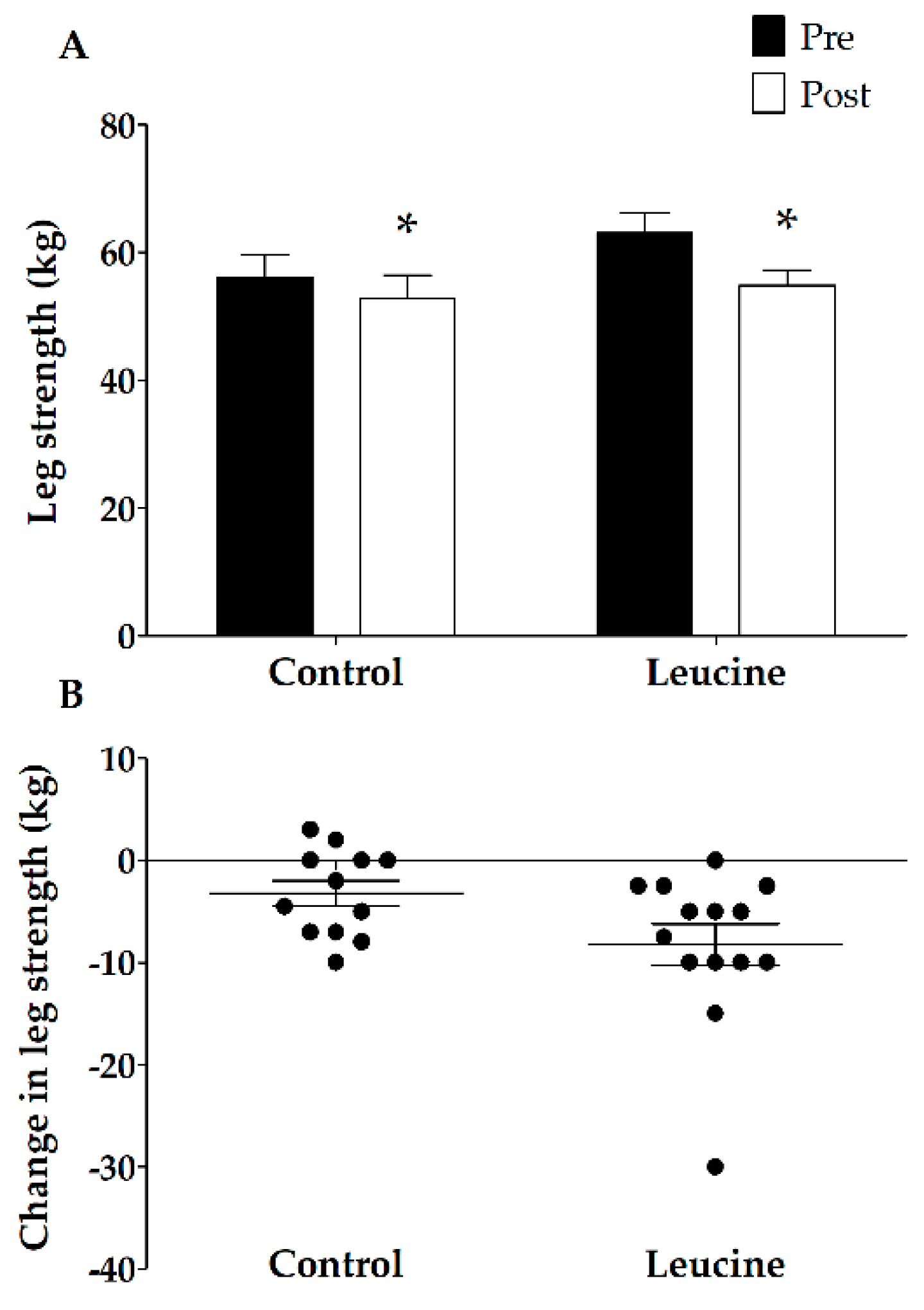
3.3. Muscle Mass and Strength
3.4. Plasma Analyses
3.5. Muscle Fiber Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 1-RM | one-repetition maximum |
| BSA | bovine serum albumin |
| CON | control group |
| CSA | cross-sectional area |
| CT-scan | computed tomography-scan |
| EDTA | ethylenediaminetetraacetic acid |
| FT | fiber typing |
| LEU | leucine group |
| MHC | myosin heavy chain |
| RDA | Recommended Dietary Allowance |
| TBDMS | tert-butyl dimethylsilyl |
References
- Evans, W.J. What is sarcopenia? J. Gerontol. A Biol. Sci. Med. Sci. 1995, 50, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; van Loon, L.J. Nutritional strategies to attenuate muscle disuse atrophy. Nutr. Rev. 2013, 71, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Paddon-Jones, D.; Campbell, W.W.; Jacques, P.F.; Kritchevsky, S.B.; Moore, L.L.; Rodriguez, N.R.; van Loon, L.J. Protein and healthy aging. Am. J. Clin. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; Dirks, M.L.; van Loon, L.J. Skeletal muscle atrophy during short-term disuse: Implications for age-related sarcopenia. Ageing Res. Rev. 2013, 12, 898–906. [Google Scholar] [CrossRef] [PubMed]
- English, K.L.; Paddon-Jones, D. Protecting muscle mass and function in older adults during bed rest. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Hvid, L.; Aagaard, P.; Justesen, L.; Bayer, M.L.; Andersen, J.L.; Ortenblad, N.; Kjaer, M.; Suetta, C. Effects of aging on muscle mechanical function and muscle fiber morphology during short-term immobilization and subsequent retraining. J. Appl. Physiol. 2010, 109, 1628–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hvid, L.G.; Suetta, C.; Nielsen, J.H.; Jensen, M.M.; Frandsen, U.; Ortenblad, N.; Kjaer, M.; Aagaard, P. Aging impairs the recovery in mechanical muscle function following 4 days of disuse. Exp. Gerontol. 2014, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Urso, M.L.; Clarkson, P.M.; Price, T.B. Immobilization effects in young and older adults. Eur. J. Appl. Physiol. 2006, 96, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Hvid, L.G.; Suetta, C.; Aagaard, P.; Kjaer, M.; Frandsen, U.; Ortenblad, N. Four days of muscle disuse impairs single fiber contractile function in young and old healthy men. Exp. Gerontol. 2013, 48, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Dirks, M.L.; Wall, B.T.; Nilwik, R.; Weerts, D.H.; Verdijk, L.B.; van Loon, L.J. Skeletal muscle disuse atrophy is not attenuated by dietary protein supplementation in healthy older men. J. Nutr. 2014, 144, 1196–1203. [Google Scholar] [CrossRef] [PubMed]
- Kortebein, P.; Ferrando, A.; Lombeida, J.; Wolfe, R.; Evans, W.J. Effect of 10 days of bed rest on skeletal muscle in healthy older adults. JAMA 2007, 297, 1772–1774. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Timmerman, K.L.; Markofski, M.M.; Walker, D.K.; Dickinson, J.M.; Jamaluddin, M.; Brasier, A.R.; Rasmussen, B.B.; Volpi, E. Short-term bed rest increases tlr4 and il-6 expression in skeletal muscle of older adults. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R216–R223. [Google Scholar] [CrossRef] [PubMed]
- Deutz, N.E.; Pereira, S.L.; Hays, N.P.; Oliver, J.S.; Edens, N.K.; Evans, C.M.; Wolfe, R.R. Effect of beta-hydroxy-beta-methylbutyrate (hmb) on lean body mass during 10 days of bed rest in older adults. Clin. Nutr. 2013, 32, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Suetta, C.; Frandsen, U.; Mackey, A.L.; Jensen, L.; Hvid, L.G.; Bayer, M.L.; Petersson, S.J.; Schroder, H.D.; Andersen, J.L.; Aagaard, P.; et al. Ageing is associated with diminished muscle re-growth and myogenic precursor cell expansion early after immobility-induced atrophy in human skeletal muscle. J. Physiol. 2013, 591, 3789–3804. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, D.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2005, 19, 422–424. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Selby, A.; Rankin, D.; Patel, R.; Atherton, P.; Hildebrandt, W.; Williams, J.; Smith, K.; Seynnes, O.; Hiscock, N.; et al. Age-related differences in the dose-response relationship of muscle protein synthesis to resistance exercise in young and old men. J. Physiol. 2009, 587, 211–217. [Google Scholar] [CrossRef] [PubMed]
- De Boer, M.D.; Selby, A.; Atherton, P.; Smith, K.; Seynnes, O.R.; Maganaris, C.N.; Maffulli, N.; Movin, T.; Narici, M.V.; Rennie, M.J. The temporal responses of protein synthesis, gene expression and cell signalling in human quadriceps muscle and patellar tendon to disuse. J. Physiol. 2007, 585, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, A.A.; Lane, H.W.; Stuart, C.A.; Davis-Street, J.; Wolfe, R.R. Prolonged bed rest decreases skeletal muscle and whole body protein synthesis. Am. J. Physiol. 1996, 270, E627–E633. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, A.A.; Tipton, K.D.; Bamman, M.M.; Wolfe, R.R. Resistance exercise maintains skeletal muscle protein synthesis during bed rest. J. Appl. Physiol. 1997, 82, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.N.; Halliday, D.; Morrison, W.L.; Stoward, P.J.; Hornsby, G.A.; Watt, P.W.; Murdoch, G.; Rennie, M.J. Decrease in human quadriceps muscle protein turnover consequent upon leg immobilization. Clin. Sci. 1987, 72, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Glover, E.I.; Phillips, S.M.; Oates, B.R.; Tang, J.E.; Tarnopolsky, M.A.; Selby, A.; Smith, K.; Rennie, M.J. Immobilization induces anabolic resistance in human myofibrillar protein synthesis with low and high dose amino acid infusion. J. Physiol. 2008, 586, 6049–6061. [Google Scholar] [CrossRef] [PubMed]
- Biolo, G.; Ciocchi, B.; Lebenstedt, M.; Barazzoni, R.; Zanetti, M.; Platen, P.; Heer, M.; Guarnieri, G. Short-term bed rest impairs amino acid-induced protein anabolism in humans. J. Physiol. 2004, 558, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Biolo, G.; Ciocchi, B.; Lebenstedt, M.; Heer, M.; Guarnieri, G. Sensitivity of whole body protein synthesis to amino acid administration during short-term bed rest. J. Gravit. Physiol. 2002, 9, P197–P198. [Google Scholar] [PubMed]
- Wall, B.T.; Snijders, T.; Senden, J.M.; Ottenbros, C.L.; Gijsen, A.P.; Verdijk, L.B.; van Loon, L.J. Disuse impairs the muscle protein synthetic response to protein ingestion in healthy men. J. Clin. Endocrinol. Metab. 2013, 98, 4872–4881. [Google Scholar] [CrossRef] [PubMed]
- Abadi, A.; Glover, E.I.; Isfort, R.J.; Raha, S.; Safdar, A.; Yasuda, N.; Kaczor, J.J.; Melov, S.; Hubbard, A.; Qu, X.; et al. Limb immobilization induces a coordinate down-regulation of mitochondrial and other metabolic pathways in men and women. PLoS ONE 2009, 4, e6518. [Google Scholar] [CrossRef] [PubMed]
- Glover, E.I.; Yasuda, N.; Tarnopolsky, M.A.; Abadi, A.; Phillips, S.M. Little change in markers of protein breakdown and oxidative stress in humans in immobilization-induced skeletal muscle atrophy. Appl. Physiol. Nutr. Metab. 2010, 35, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, T.; Osterlund, T.; Flanagan, J.N.; von Walden, F.; Trappe, T.A.; Linnehan, R.M.; Tesch, P.A. Effects of 3 days unloading on molecular regulators of muscle size in humans. J. Appl. Physiol. 2010, 109, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.D.; Gurkin, B.E.; Matin, S.; Wolfe, R.R. Nonessential amino acids are not necessary to stimulate net muscle protein synthesis in healthy volunteers. J. Nutr. Biochem. 1999, 10, 89–95. [Google Scholar] [CrossRef]
- Volpi, E.; Kobayashi, H.; Sheffield-Moore, M.; Mittendorfer, B.; Wolfe, R.R. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am. J. Clin. Nutr. 2003, 78, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C.; Anthony, T.G.; Layman, D.K. Leucine supplementation enhances skeletal muscle recovery in rats following exercise. J. Nutr. 1999, 129, 1102–1106. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C.; Reiter, A.K.; Anthony, T.G.; Crozier, S.J.; Lang, C.H.; MacLean, D.A.; Kimball, S.R.; Jefferson, L.S. Orally administered leucine enhances protein synthesis in skeletal muscle of diabetic rats in the absence of increases in 4e-bp1 or s6k1 phosphorylation. Diabetes 2002, 51, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Dardevet, D.; Sornet, C.; Bayle, G.; Prugnaud, J.; Pouyet, C.; Grizard, J. Postprandial stimulation of muscle protein synthesis in old rats can be restored by a leucine-supplemented meal. J. Nutr. 2002, 132, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Frexes-Steed, M.; Lacy, D.B.; Collins, J.; Abumrad, N.N. Role of leucine and other amino acids in regulating protein metabolism in vivo. Am. J. Physiol. 1992, 262, E925–E935. [Google Scholar] [CrossRef] [PubMed]
- Koopman, R.; Wagenmakers, A.J.; Manders, R.J.; Zorenc, A.H.; Senden, J.M.; Gorselink, M.; Keizer, H.A.; van Loon, L.J. Combined ingestion of protein and free leucine with carbohydrate increases postexercise muscle protein synthesis in vivo in male subjects. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E645–E653. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Balage, M.; Sornet, C.; Giraudet, C.; Pujos, E.; Grizard, J.; Mosoni, L.; Dardevet, D. Leucine supplementation improves muscle protein synthesis in elderly men independently of hyperaminoacidaemia. J. Physiol. 2006, 575, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Ham, D.J.; Caldow, M.K.; Lynch, G.S.; Koopman, R. Leucine as a treatment for muscle wasting: A critical review. Clin. Nutr. 2014, 33, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C.; Anthony, T.G.; Kimball, S.R.; Vary, T.C.; Jefferson, L.S. Orally administered leucine stimulates protein synthesis in skeletal muscle of postabsorptive rats in association with increased eif4f formation. J. Nutr. 2000, 130, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C.; Yoshizawa, F.; Anthony, T.G.; Vary, T.C.; Jefferson, L.S.; Kimball, S.R. Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive pathway. J. Nutr. 2000, 130, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Kimball, S.R.; Jefferson, L.S. Signaling pathways and molecular mechanisms through which branched-chain amino acids mediate translational control of protein synthesis. J. Nutr. 2006, 136, 227S–231S. [Google Scholar] [CrossRef] [PubMed]
- Magne, H.; Savary-Auzeloux, I.; Remond, D.; Dardevet, D. Nutritional strategies to counteract muscle atrophy caused by disuse and to improve recovery. Nutr. Res. Rev. 2013, 26, 149–165. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.J. Leucine as a pharmaconutrient in health and disease. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Leenders, M.; van Loon, L.J. Leucine as a pharmaconutrient to prevent and treat sarcopenia and type 2 diabetes. Nutr. Rev. 2011, 69, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Backx, E.M.P.; Hangelbroek, R.; Snijders, T.; Verscheijden, M.L.; Verdijk, L.B.; de Groot, L.; van Loon, L.J.C. Creatine loading does not preserve muscle mass or strength during leg immobilization in healthy, young males: A randomized controlled trial. Sports Med. 2017, 47, 1661–1671. [Google Scholar] [CrossRef] [PubMed]
- Dirks, M.L.; Wall, B.T.; Snijders, T.; Ottenbros, C.L.; Verdijk, L.B.; van Loon, L.J. Neuromuscular electrical stimulation prevents muscle disuse atrophy during leg immobilization in humans. Acta Physiol. 2014, 210, 628–641. [Google Scholar] [CrossRef] [PubMed]
- Verdijk, L.B.; van Loon, L.; Meijer, K.; Savelberg, H.H. One-repetition maximum strength test represents a valid means to assess leg strength in vivo in humans. J. Sports Sci. 2009, 27, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, J. Percutaneous needle biopsy of skeletal muscle in physiological and clinical research. Scand. J. Clin. Lab. Investig. 1975, 35, 609–616. [Google Scholar] [CrossRef]
- Wall, B.T.; Dirks, M.L.; Snijders, T.; Senden, J.M.; Dolmans, J.; van Loon, L.J. Substantial skeletal muscle loss occurs during only 5 days of disuse. Acta Physiol. 2014, 210, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; Hamer, H.M.; de Lange, A.; Kiskini, A.; Groen, B.B.; Senden, J.M.; Gijsen, A.P.; Verdijk, L.B.; van Loon, L.J. Leucine co-ingestion improves post-prandial muscle protein accretion in elderly men. Clin. Nutr. 2013, 32, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; Morton, J.P.; van Loon, L.J. Strategies to maintain skeletal muscle mass in the injured athlete: Nutritional considerations and exercise mimetics. Eur. J. Sport Sci. 2015, 15, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Verdijk, L.B.; Koopman, R.; Schaart, G.; Meijer, K.; Savelberg, H.H.; van Loon, L.J. Satellite cell content is specifically reduced in type ii skeletal muscle fibers in the elderly. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E151–E157. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.R.; Kuo, Y.F.; Graham, J.E.; Ottenbacher, K.J.; Ostir, G.V. Early ambulation and length of stay in older adults hospitalized for acute illness. Arch. Intern. Med. 2010, 170, 1942–1943. [Google Scholar] [CrossRef] [PubMed]
- Trappe, T.A.; Burd, N.A.; Louis, E.S.; Lee, G.A.; Trappe, S.W. Influence of concurrent exercise or nutrition countermeasures on thigh and calf muscle size and function during 60 days of bed rest in women. Acta Physiol. 2007, 191, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Brooks, N.; Cloutier, G.J.; Cadena, S.M.; Layne, J.E.; Nelsen, C.A.; Freed, A.M.; Roubenoff, R.; Castaneda-Sceppa, C. Resistance training and timed essential amino acids protect against the loss of muscle mass and strength during 28 days of bed rest and energy deficit. J. Appl. Physiol. 2008, 105, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Rejc, E.; di Prampero, P.E.; Lazzer, S.; Grassi, B.; Simunic, B.; Pisot, R.; Antonutto, G.; Narici, M. Maximal explosive power of the lower limbs before and after 35 days of bed rest under different diet energy intake. Eur. J. Appl. Physiol. 2015, 115, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Dickinson, J.M.; Fry, C.S.; Walker, D.K.; Gundermann, D.M.; Reidy, P.T.; Timmerman, K.L.; Markofski, M.M.; Paddon-Jones, D.; Rasmussen, B.B.; et al. Bed rest impairs skeletal muscle amino acid transporter expression, mtorc1 signaling, and protein synthesis in response to essential amino acids in older adults. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1113–E1122. [Google Scholar] [CrossRef] [PubMed]
- Suetta, C.; Frandsen, U.; Jensen, L.; Jensen, M.M.; Jespersen, J.G.; Hvid, L.G.; Bayer, M.; Petersson, S.J.; Schroder, H.D.; Andersen, J.L.; et al. Aging affects the transcriptional regulation of human skeletal muscle disuse atrophy. PLoS ONE 2012, 7, e51238. [Google Scholar] [CrossRef] [PubMed]
- Christensen, T.; Bendix, T.; Kehlet, H. Fatigue and cardiorespiratory function following abdominal surgery. Br. J. Surg. 1982, 69, 417–419. [Google Scholar] [CrossRef] [PubMed]
- Deitrick, J.E. The effect of immobilization on metabolic and physiological functions of normal men. Bull. N. Y. Acad. Med. 1948, 24, 364–375. [Google Scholar] [PubMed]
- Stuart, C.A.; Shangraw, R.E.; Prince, M.J.; Peters, E.J.; Wolfe, R.R. Bed-rest-induced insulin resistance occurs primarily in muscle. Metab. Clin. Exp. 1988, 37, 802–806. [Google Scholar] [CrossRef]
- Haruna, Y.; Suzuki, Y.; Kawakubo, K.; Yanagibori, R.; Gunji, A. Decremental reset in basal metabolism during 20-days bed rest. Acta Physiol. Scand. Suppl. 1994, 616, 43–49. [Google Scholar] [PubMed]
- Wall, B.T.; Dirks, M.L.; Snijders, T.; van Dijk, J.W.; Fritsch, M.; Verdijk, L.B.; van Loon, L.J. Short-term muscle disuse lowers myofibrillar protein synthesis rates and induces anabolic resistance to protein ingestion. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E137–E147. [Google Scholar] [CrossRef] [PubMed]
- Pennings, B.; Groen, B.; de Lange, A.; Gijsen, A.P.; Zorenc, A.H.; Senden, J.M.; van Loon, L.J. Amino acid absorption and subsequent muscle protein accretion following graded intakes of whey protein in elderly men. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E992–E999. [Google Scholar] [CrossRef] [PubMed]
- Casperson, S.L.; Sheffield-Moore, M.; Hewlings, S.J.; Paddon-Jones, D. Leucine supplementation chronically improves muscle protein synthesis in older adults consuming the rda for protein. Clin. Nutr. 2012, 31, 512–519. [Google Scholar] [CrossRef] [PubMed]
- English, K.L.; Mettler, J.A.; Ellison, J.B.; Mamerow, M.M.; Arentson-Lantz, E.; Pattarini, J.M.; Ploutz-Snyder, R.; Sheffield-Moore, M.; Paddon-Jones, D. Leucine partially protects muscle mass and function during bed rest in middle-aged adults. Am. J. Clin. Nutr. 2016, 103, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, S.; Vanschoonbeek, K.; Verdijk, L.B.; Koopman, R.; Wodzig, W.K.; Dendale, P.; van Loon, L.J. Long-term leucine supplementation does not increase muscle mass or strength in healthy elderly men. Am. J. Clin. Nutr. 2009, 89, 1468–1475. [Google Scholar] [CrossRef] [PubMed]
- Leenders, M.; Verdijk, L.B.; van der Hoeven, L.; van Kranenburg, J.; Hartgens, F.; Wodzig, W.K.; Saris, W.H.; van Loon, L.J. Prolonged leucine supplementation does not augment muscle mass or affect glycemic control in elderly type 2 diabetic men. J. Nutr. 2011, 141, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Panel, E.N. Scientific opinion on dietary reference values for protein. EFSA J. 2012, 10, 2557. [Google Scholar]
- Tieland, M.; Borgonjen-Van den Berg, K.J.; van Loon, L.J.; de Groot, L.C. Dietary protein intake in community-dwelling, frail, and institutionalized elderly people: Scope for improvement. Eur. J. Nutr. 2012, 51, 173–179. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Control | Leucine | |
|---|---|---|
| Age (years) | 23 ± 1 | 21 ± 1 |
| Body mass (kg) | 73.1 ± 3.2 | 73.5 ± 1.9 |
| Height (m) | 1.76 ± 0.03 | 1.80 ± 0.02 |
| BMI (kg·m−2) | 23.5 ± 0.8 | 22.7 ± 0.6 |
| 1 RM leg extension (kg) | 56 ± 4 | 63 ± 3 |
| Whole-thigh muscle CSA (mm2) | 14,184 ± 462 | 14,417 ± 537 |
| Quadriceps muscle CSA (mm2) | 7712 ± 324 | 7643 ± 317 |
| Control Group (n = 14) | Leucine Group (n = 8) | |||
|---|---|---|---|---|
| Pre-Immobilization | During Immobilization | Pre-Immobilization | During Immobilization | |
| Energy (MJ/day) | 7.6 ± 0.4 | 7.4 ± 0.6 | 10.0 ± 1 | 9.8 ± 1 |
| Carbohydrate (En%) | 54 ± 8 | 47 ± 3 | 52 ± 3 | 50 ± 3 |
| Fat (En%) | 31 ± 2 | 31 ± 3 | 31 ± 3 | 29 ± 3 |
| Protein (En%) | 19 ± 1 | 18 ± 1 | 16 ± 1 | 17 ± 2 |
| Protein (g/kg/day) | 1.2 ± 0.1 | 1.1 ± 0.1 | 1.3 ± 0.2 | 1.3 ± 0.2 |
| Fiber Type | Control Group (n = 13) | Leucine Group (n = 11) | |||
|---|---|---|---|---|---|
| Pre-Immobilization | Post-Immobilization | Pre-Immobilization | Post-Immobilization | ||
| Muscle fiber CSA (μm2) | I | 6034 ± 501 | 6620 ± 508 | 5781 ± 354 | 6196 ± 415 |
| II * | 7202 ± 640 | 7540 ± 587 | 6033 ± 482 | 6589 ± 509 | |
| Fiber type distribution (%) | I | 38 ± 4 | 33 ± 3 | 39 ± 5 | 41 ± 5 |
| II * | 62 ± 4 | 67 ± 3 | 61 ± 5 | 59 ± 5 | |
| Fiber type distribution (% area) | I | 34 ± 4 | 31 ± 3 | 38 ± 4 | 40 ± 5 |
| II * | 66 ± 4 | 69 ± 3 | 62 ± 4 | 60 ± 5 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Backx, E.M.P.; Horstman, A.M.H.; Marzuca-Nassr, G.N.; Van Kranenburg, J.; Smeets, J.S.; Fuchs, C.J.; Janssen, A.A.W.; De Groot, L.C.P.G.M.; Snijders, T.; Verdijk, L.B.; et al. Leucine Supplementation Does Not Attenuate Skeletal Muscle Loss during Leg Immobilization in Healthy, Young Men. Nutrients 2018, 10, 635. https://doi.org/10.3390/nu10050635
Backx EMP, Horstman AMH, Marzuca-Nassr GN, Van Kranenburg J, Smeets JS, Fuchs CJ, Janssen AAW, De Groot LCPGM, Snijders T, Verdijk LB, et al. Leucine Supplementation Does Not Attenuate Skeletal Muscle Loss during Leg Immobilization in Healthy, Young Men. Nutrients. 2018; 10(5):635. https://doi.org/10.3390/nu10050635
Chicago/Turabian StyleBackx, Evelien M.P., Astrid M.H. Horstman, Gabriel N. Marzuca-Nassr, Janneau Van Kranenburg, Joey S. Smeets, Cas J. Fuchs, Anniek A.W. Janssen, Lisette C.P.G.M. De Groot, Tim Snijders, Lex B. Verdijk, and et al. 2018. "Leucine Supplementation Does Not Attenuate Skeletal Muscle Loss during Leg Immobilization in Healthy, Young Men" Nutrients 10, no. 5: 635. https://doi.org/10.3390/nu10050635