
Endocannabinoid System in the Airways

Department of Pharmacology, Faculty of Pharmacy, Hacettepe University, Ankara 06100, Turkey
Molecules 2019, 24(24), 4626; https://doi.org/10.3390/molecules24244626
Submission received: 13 November 2019
/
Revised: 12 December 2019
/
Accepted: 15 December 2019
/
Published: 17 December 2019
(This article belongs to the Special Issue Research on the Cannabinoid Receptors Pharmacology)
Abstract
:Cannabinoids and the mammalian endocannabinoid system is an important research area of interest and attracted many researchers because of their widespread biological effects. The significant immune-modulatory role of cannabinoids has suggested their therapeutic use in several inflammatory conditions. Airways are prone to environmental irritants and stimulants, and increased inflammation is an important process in most of the respiratory diseases. Therefore, the main strategies for treating airway diseases are suppression of inflammation and producing bronchodilation. The ability of cannabinoids to induce bronchodilation and modify inflammation indicates their importance for airway physiology and pathologies. In this review, the contribution of cannabinoids and the endocannabinoid system in the airways are discussed, and the existing data for their therapeutic use in airway diseases are presented.
1. Cannabinoids and the Endocannabinoid System
The plant cannabis sativa, produces a variety of biologically active cannabinoids and related compounds. ∆9-tetrahydrocannabinol (THC) is the predominant molecule with 1–10% by weight. It was first extracted in 1964 by Gaoni and Mechoulum and is now known to be the primary molecule responsible for most of the biological effects of the cannabis plant [1]. Thereafter, in 1992, N-arachidonylethanolamine “anandamide” (AEA) was isolated from swine brains, which was the first endogenous cannabinoid related substance isolated from a mammal [2]. Related substances were also isolated from gastrointestinal tissues [3,4]. In the following years, multiple endogenous cannabinoid molecules like 2-arachidonylglycerol (2-AG), noladin ether, virodhamine, and oleoyl ethanolamine were identified and these substances were then named as “endocannabinoids”, which are derivatives of arachidonic acid conjugated with ethanolamine or glycerol. Endocannabinoids, their receptors, and metabolic pathways form the “Endocannabinoid System”, a term which was first used by Di Marzo and Fontana [5].
Among the wide variety of endocannabinoids, AEA and 2-AG have attracted a significant number of researchers in the endocannabinoid area. AEA is synthesized by the enzymes N-acyl-phosphatidylethanolamine phospholipase D (NAPE-PLD), α/β-hydrolase-4 (Abh4), and phospholipase-C (PLC)-catalyzed cleavage of NAPE to phosphoanandamide and then to AEA [6,7]. It is mainly catabolized by fatty acid amide hydrolase (FAAH), the enzyme which also catabolizes the non-cannabinoid fatty acids [8]. In addition to FAAH, N-acylethanolamine acid amidase (NAAA) has also been identified as another hydrolase for AEA [9]. 2-AG is synthesized by phospholipase C (PLC) and diacyl-glycerol-lipase (DAGL), and catabolized mainly by monoacylglycerol-lipase (MAGL) [10]. Although MAGL is the predominant enzyme for 2-AG metabolism, α/β-hydrolase-6 (Abh6) and α/β-hydrolase-12 (Abh12) also contribute to a minor degree. FAAH has a negligible effect in 2-AG catabolism [7]. In macrophages, it was also shown that carboxylesterase-1 (CES1) and palmitoyl protein thioesterase-1 (PPT1) could hydrolyze 2-AG [11,12]. Nevertheless, cyclooxygenase-2 (COX-2) is involved in the oxidation of both 2-AG and AEA [13]. COX-2 oxidizes AEA to generate prostamides, like prostaglandin H2 ethanolamide (PGH2-EA) and prostaglandin H2 (PGH2), respectively [14], and 2-AG to prostaglandin H2 glycerol (PGH2-G) [14,15].
Cannabinoids exert their biological effects mainly through two 7-transmembrane (TM) receptors, namely cannabinoid receptor CB1 and CB2 [16]. However, they can also bind to other targets like transient receptor potential vanilloid receptor1 (TRPV1) [7,17], orphan receptors G protein-coupled receptor-55 (GPR55) [18,19], GPR18 [20], GPR110 [21], GPR119 [22], and peroxisome proliferator-activated receptors (PPARs) (mostly PPARα) [22]. The cannabinoid receptors are G-protein coupled receptors (GPCRs) (Gi/o) and human CB1/CB2 receptors share 44% overall homology [16,23]. Since mice have been used in several cannabinoid related studies, it is also important to note that human and mouse CB1 receptors share 96% [24], and CB2 receptors share 82% homology [25], where mouse CB1 and CB2 receptors share 66% homology [25]. Both CB1 and CB2 receptors are negatively coupled to adenylyl cyclase and stimulate mitogen-activated protein kinase (MAPK). Furthermore, they also activate K+ channels and inhibit Ca++ channels, both of which result in the inhibition of transmitter release via the activation of Gβ/γ subunit [26,27,28].
CB1 receptors are mainly located in the nervous system, nerve terminals and a wide range of tissues including adipose tissue, liver, gastrointestinal tract, whereas CB2 receptors are expressed in peripheral tissues, primarily in immune cells, immune-related organs and tissues like tonsils, spleen, thymus and bone marrow [7,29,30]. However, similar to CB1, CB2 receptor expression has also been demonstrated in various tissues and cells such as the brain, spinal cord [31,32], lung, testes [23,33], osteoblasts, osteocytes, and osteoclasts [34].
Although most of the research has been focused on the central nervous system effects of cannabinoids, they have various biological effects in the immune, cardiovascular, gastrointestinal, and respiratory system [35]. These effects had attracted the interest in the clinical use of cannabinoids. However, most of the cannabinoid related drugs have been withdrawn from the market due to unacceptable central side effects, which are mainly mediated by CB1 receptors. Today “Sativex®” is the only cannabinoid-containing product, which consists of ∆9-tetrahydrocannabinol (THC) and cannabidiol, and have been approved for the spasticity of multiple sclerosis.
Cannabinoids have a significant contribution to the immune modulation [36], which suggests their importance in pathophysiological conditions and inflammatory diseases. The alteration of endocannabinoid levels by inflammation was observed in several studies performed in the brain, liver, coronary artery, and colon [37,38,39,40,41]. Furthermore, endocannabinoids and activation of the cannabinoid system have also been demonstrated to inhibit inflammatory responses in hepatitis [42], pulmonary inflammation [43], inflammatory pain [44], sepsis [45] and colitis [46] by reducing the recruitment of inflammatory cells and increasing the production of anti-inflammatory cytokines. Earlier studies have indicated that cannabinoids can inhibit antibody production [47,48]. THC has been shown to suppress Th1 and enhance Th2 cytokines, and AEA has been demonstrated to decrease T and B cell proliferation [49,50]. However, the immune-modulatory effects of cannabinoids can also lead to the down-regulation of immune responses, which can cause the augmentation of infections and tumorigenesis [51,52,53].
The intense involvement of cannabinoids in immunomodulation also suggested the role of CB2 receptors as they are highly expressed in the immune system. CB2 receptors are involved in B-cell differentiation [54], macrophage migration [55], and antigen processing [56]. CB2 receptors also contribute to mast cell activation by generating nitric oxide (NO) and prostaglandin-E2 (PGE2) [57]. Besides CB2 receptors, the existence of functional CB1 receptors was also shown in mast cells [58], suggesting both receptors are also important in allergic diseases [59,60]. On the other hand, when used in higher concentrations, the effects of cannabinoids on inflammation can occur through cannabinoid receptor-independent mechanisms such as PPARs [61,62].
2. The Role of Endocannabinoid System in the Airways
It is known that marijuana smoking can cause cellular damage in the lungs. However, there are some differences in the airways of marijuana and tobacco smokers [63,64]. Gong et al. have shown that both marijuana and tobacco smokers have goblet and basal cell hyperplasia with a tendency of higher hyperplasia in the marijuana group [65]. There was also cellular disorganization in more than 50% of marijuana smokers. In a study performed in primates, smoking marijuana was shown to induce bronchiolitis, alveolar cell hyperplasia, and fibrosis in greater incidence when compared to that of the cigarette group [66]. Therefore, it can be concluded that the plant-derived cannabinoids may have significant effects in the airways.
The synthesis of endocannabinoids in the airways is established in various cell types (Figure 1). Although the data about the expression of CB1 and CB2 cannabinoid receptors in human airways are not clear, it is known that they are mostly expressed by the immune cells within the airways [67,68]. CB2 receptors are densely expressed on eosinophils [69,70,71] and monocytes, both of which also express CB1 receptors [33,72,73,74]. Eosinophil recruitment to the airways is an important process in the chronic inflammatory state of allergic asthma. The significant amount of CB2 receptors in human eosinophils indicates a critical response capacity of these cells to cannabinoids [75]. In this regard, 2-AG was shown to be a chemoattractant factor for human primary eosinophils [69,76,77]. Moreover, interleukin-5 (IL-5), which is an important mediator for eosinophil differentiation and priming, can enhance the effect of 2-AG [77]. Selective stimulation of CB2 receptors with synthetic ligands is also able to modify the activity of eosinophils to chemoattractants [78]. These studies indicate that endocannabinoids have a significant contribution to eosinophil recruitment. Human monocyte-derived dendritic cells (DCs) isolated from peripheral blood and murine bone marrow-derived DCs express both CB1 and CB2 receptors, which can synthesize AEA and 2-AG [49,79,80]. Studies have shown that the activation of DCs by cannabinoid ligands can inhibit the release of inflammatory cytokines [49] and can suppress the immune response by inducing apoptosis of these cells [79]. The interaction of DCs with T cells should also be considered for the effects of cannabinoids in the immune response. Do et al. suggested that inflammation-induced 2-AG production by DCs can affect cannabinoid receptors on T cells and switch the immune response from Th2 to Th1, and T cells can produce endocannabinoids which may affect their receptors on DCs. [80]. The expression of cannabinoid receptors and their function is more complicated in neutrophils. Detectable CB2 mRNA levels were shown in polymorphonuclear neutrophil cells [33]. However, Chouinard et al. have demonstrated that human neutrophils do not express a significant amount of CB2 receptor protein, although they are responsive to selective agonists [70,71,81].
2.1. Airway Reactivity
In 1973, Tashkin et al., have performed a study in healthy volunteers, in which they have investigated the effects of cannabinoids in the airways. Interestingly, marihuana inhalation and oral ingestion of THC had produced bronchodilation lasting for sixty minutes and six hours, respectively [82]. In the meantime, Vachon et al. have demonstrated a similar bronchodilatory effect by cannabinoids in healthy volunteers and have shown that the effect was dose-dependent [83]. Furthermore, in the doses that they produce bronchodilation, cannabinoids did not cause central respiratory depression.
These findings suggested the role of cannabinoids in asthma treatment. In two different studies, Tashkin et al., have shown that smoking marijuana or ingestion of THC by subjects with chronic, clinically stable, bronchial asthma of minimal or moderate severity, can produce bronchodilation [84,85]. A few years later, Hartley et al. have demonstrated that the bronchodilator effect of THC can be observed in concentrations that do not cause central or cardiovascular effects [86]. However, the mechanism for the bronchodilatory effect of cannabinoids was not known, and this effect did not appear to be related to β-adrenoceptor stimulation or cholinergic blockade [86].
The physiological significance of these studies and their therapeutic potential were complicated because some asthmatic patients had responded to these compounds with paradoxical bronchospasm [87,88]. In 2000, Calignano et al. have performed a study in rats and guinea pigs in order to investigate this controversy [89]. They have demonstrated that cannabinoids have bidirectional effects in the airways, depending on the airway tone. In their study, AEA had inhibited bronchial responsiveness to chemical irritation in rodents but caused bronchospasm when the constricting tone was removed. They have shown that both effects were mediated through peripheral CB1 cannabinoid receptors located on axon terminals of airway nerves [89].
In the isolated organ bath experiments, AEA and synthetic CB2 receptor agonist WIN 55,212-2 was shown to inhibit electrical field stimulation (EFS)-induced contractions of the rat tracheal rings by acting pre-junctionally [90]. This inhibitory effect was no surprise considering the inhibitory second messenger pathways mediating the effects of CB1 and CB2 receptors. In accordance with this data, cannabinoid agonists have been shown to inhibit acetylcholine release from cholinergic nerves via activation of CB2 [91]. However, this inhibition was not associated with a functional response.
Research on the effect of cannabinoids in the airways was mostly focused on airway inflammation after that time, and no new data was produced about their functional roles until the work by Grassin-Delyle et al. was published [92]. They have shown that pre-junctional CB1 receptor activation mediates the inhibition of cholinergic nerve mediated contractions in the human bronchus. Therefore, they have suggested this mechanism for the explanation of acute bronchodilation produced by marijuana smoking. Bozkurt et al. have also demonstrated that CB1 stimulation can inhibit the increased neuronal activity and nerve density in airway inflammation and can directly inhibit cholinergic contractions by a presynaptic mechanism, indicating a protective role of CB1 receptors in airway inflammation [93]. These findings indicate that cannabinoids can inhibit bronchoconstriction by a pre-junctional inhibition of neurotransmission.
2.2. Airway Diseases
Allergen challenge of asthmatic patients has been shown to result in increased AEA concentrations in their bronchoalveolar lavage (BAL) fluid [68]. This finding suggests the contribution of the endocannabinoid system in the pathophysiology of allergic asthma. It is also the first study to report the involvement of endocannabinoids in human asthma [68]. In that study, Zoerner et al. have demonstrated that allergen exposure increases AEA levels from about 5 pmol/L to 30 pmol/L in human BAL samples [68]. However, it cannot be concluded still, if the increased AEA concentrations is a cause or consequence of the pathophysiology of asthma in humans. As highlighted above, cannabinoids have significant anti-inflammatory effects; however, the endocannabinoid system can induce both pro-inflammatory and anti-inflammatory responses [94,95]. This controversy may be due to their heterogeneous targets other than cannabinoid receptors, like TRPV1, GPR119, GPR55, and PPARs [6]. However, in a study performed cardiomyocytes it has been shown that AEA concentrations should be around the micromolar range in order to induce tissue damage, which is very high when compared to that of normal tissue levels [95]. Therefore, as speculated by Zoerner et al., it can be concluded that the increase in AEA production could be a protective mechanism rather than a pathological component. AEA levels in the airways were reported as around 2 ng/mg tissue for mouse [96] and 0.3 pg/mL for rabbit lung [97], whereas 2-AG levels were about 20 ng/mg in the mouse lung [96].
Another cannabinoid related endogenous lipid mediator palmitoylethanolamide (PEA), has been shown to be decreased in the airways after allergen sensitization [98]. The concentration of PEA was reported to be around 6 pmol/mg in mouse harvested bronchi, which was reduced to 1 pmol/mg after ovalbumin sensitization [98]. PEA is co-released with AEA and behaves as a local autacoid down-regulator of mast cell activation and inflammation [98]. The supplementation of PEA has also been proposed to prevent the development of asthma-like features in the same study.
The plant-derived non-psychotropic cannabinoid cannabidiol, has immunosuppressive and anti-inflammatory effects [99]. In mice, cannabidiol has been shown to suppress lipopolysaccharide (LPS)-induced TNF-α production [100]. Ribeiro et al., have demonstrated in two different studies that cannabidiol can decrease inflammation and improve lung functions in LPS-induced acute lung injury in mice [101,102], which is further confirmed by Vuolo et al. [103]. The influence of cannabidiol on the antigen-induced contraction of guinea-pig airways has also been demonstrated [104]. However, it should also be noted that cannabidiol can cause drug interactions, hepatic abnormalities, diarrhea, fatigue, vomiting, and somnolence [105]. Therefore, these possible side effects and their severity should be considered for using cannabidiol as a therapeutic tool.
Studies have shown that endocannabinoids and cannabinoid CB1 receptors may have a significant inhibitory role in human mast cell degranulation and activation in the airway mucosa and skin, suggesting the contribution of the endocannabinoid system in the allergic diseases [59,60]. Martin-Fontecha et al. have shown that the expression of CB1 receptor has been upregulated in tonsils and peripheral blood of patients with allergic rhinitis, atopic dermatitis, and food allergy [106]. In accordance with these studies, the high expression of the CB1 receptor proteins were also demonstrated in B cells, T cells, pDCs, and mDCs of atopic donors [106]. Symptomatic allergic donors were also found to have higher expression of CB2 receptors on their eosinophils [78]. Frei et al. have shown that migratory responses of human and mouse eosinophils can be enhanced by selective activation of CB2 receptors through Gαq/MEK/ROCK (Gαq/ mitogen/extracellular signal-regulated kinase / rho-associated protein kinase) signaling [78].
The expression levels of cannabinoid receptors in the lungs were also shown to be affected by viral infections [107]. Tahamtan et al., have suggested in their studies that respiratory syncytial virus (RSV) infection of airways lead to an induction in CB1 receptor expression. However, in another study, they have also demonstrated the contribution of CB2 receptors for RSV infection in both mice and humans [108].
3. Targeting the Endocannabinoid System for the Treatment of Airway Diseases
Several pathological conditions have been associated with a change in the expression of cannabinoid receptors, altered endocannabinoid tissue concentrations, or a change in their metabolism. Therefore, different therapeutic strategies were considered by modifying the endocannabinoid system, such as targeting CB1/CB2 receptors or interfering with their metabolism [6].
Due to the immunomodulatory effect of cannabinoids, a significant variety of studies were focused on their possible therapeutic potential on inflammatory diseases like asthma [43,67,68,92,93,104,109,110,111,112,113,114]. The molecular mechanisms mediating the effects of cannabinoids in allergic airway responses mainly depend on their effects on immune cells and the related release of cytokines [115]. In mice, treatment with the plant-derived cannabinoids, cannabinol and THC, was able to inhibit the expression of critical T cell cytokines and inflammatory response in ovalbumin-induced experimental allergic airway inflammation [116]. Using the ovalbumin model, Braun et al., have demonstrated that THC can inhibit cell proliferation, suppress cytokine and chemokine production, and stimulate regulatory T cells [112]. However, they have also suggested that these effects are probably mediated by cannabinoid receptor-independent mechanisms [112].
Giannini et al. demonstrated that the non-selective cannabinoid receptor agonist CP-55,940 can prevent allergen-induced bronchospasm, and reduce cough and leukocyte recruitment in the lung [67]. This effect could be blocked by both CB1 and CB2 receptor blockade, suggesting that both receptors were involved in this effect [67]. In guinea pig airways, the synthetic cannabinoid ligand WIN-55212-2 has demonstrated a reduction in airway neurogenic inflammation in vivo [91]. This was due to an inhibition of C-fiber nerve activity and mediated by the activation of CB2 receptors. WIN-55,212-2 was also shown to inhibit airway plasma extravasation and bronchoconstriction induced by intra-esophageal HCl instillation in guinea-pigs, an effect which was also mediated through CB2 activation [113]. CB2 receptor modulation of airway sensory nerve activity and related cough reflex have been highlighted in several different studies [110,117]. These findings suggest that CB2 receptors can contribute to both neurogenic airway inflammation and hyperreactivity as well. This may also be related to a species difference since studies underscoring the CB2 receptor-mediated reactivity changes in the airways are mostly performed in guinea pigs. However, the crucial role of CB2 activation in the regulation of pulmonary natural killer (NK) cell function has also been demonstrated in mice [118]. Therefore, blocking the CB2 receptors in allergic airway inflammation may modulate NK cell response during airway inflammation. The plant-derived cannabinoid, cannabidiol, was also shown to inhibit allergen-induced contraction of airway smooth muscle, and reduce antigen-induced airway obstruction in guinea pigs [104]. This effect seems to be partly mediated by the inhibition of mast cell degranulation.
In the experimental studies performed in mice, selective CB1 receptor agonists have been shown to inhibit inflammation-induced hyperreactivity in an in vitro model of nerve growth factor (NGF)-induced neurogenic inflammation [93]. The inhibitory effect of CB1 receptors was demonstrated using two different synthetic selective CB1 agonists ACEA and ACPA. In the same study, these selective ligands were also able to inhibit nerve mediated cholinergic contractions, further confirming the inhibitory role of CB1 receptors in airway reactivity [93]. Moreover, in a model of experimental non-atopic asthma in mice, in vivo intranasal treatment with the CB1 selective agonist, ACEA was shown to prevent airway hyperreactivity [111].
Inhaled AEA pre-treatment of guinea pigs has shown to prevent leukotriene-D4 aerosol-induced bronchospasm [114], an effect suggesting the role of endocannabinoid targeted therapy in airway diseases. AEA has also been shown to reduce trans-epithelial resistance in airway cells, which indicates an increase in barrier permeability [119]. However, this effect seems to be mediated by the metabolism of AEA to one or more LOX and COX metabolites rather than CB1 and CB2 receptor-dependent mechanisms. Increased AEA levels in asthmatic patients may contribute to increased permeability of the epithelium through degradation to arachidonic acid metabolites. Therefore, preventing AEA hydrolysis in the airways may help to prevent epithelial permeability in asthma [119]. A strategy for this and to increase AEA tissue levels is to inhibit the activity of the FAAH enzyme, which is responsible for the hydrolysis of AEA and other related amidated signaling lipids, such as PEA, N-oleoylethanolamide (OEA) and linoleoylethanolamide (LEA). Inhibition of FAAH has been shown to produce antitussive effects in guinea pigs [120]. This effect was suggested to be mediated by elevated fatty amino acids, which act on cannabinoid (CB2) receptors on vagal sensory nerves [120]. Therefore, increasing the endocannabinoid levels by FAAH inhibition can be a promising strategy as a new treatment option for antitussive therapy.
The effect of endocannabinoids was studied in the LPS-induced experimental acute lung injury (ALI) model in mice [121]. It is well known that ALI may occur due to sepsis, pneumonia, acid aspiration, toxic inhalation, etc. In their study, Costola-de-Souza et al. showed that treatment with the MAGL inhibitor, JZL184 attenuated the pathological changes of ALI by increasing 2-AG levels in the lungs. However, in their study, neither CB1 nor CB2 receptor antagonists were able to fully block the effect of JZL184, suggesting the involvement of other mechanisms such as stimulation of non-cannabinoid receptors or an increase in non-cannabinoid products like prostaglandins. Abohalaka et al. studied the effects of FAAH or MAGL inhibitor treatments in LPS-induced airway inflammation and airway hyperreactivity [109]. These inhibitors were applied either systemically (i.p. route) or locally (i.n.) before LPS administration to mice. Both FAAH inhibitor and MAGL inhibitor treatments were found to be effective in preventing airway hyperreactivity, whether they were applied via the i.p. or i.n. route. The treatments were also able to prevent the histopathological changes in the lungs, except the local i.n. application of the FAAH inhibitor URB597 [109]. These effects were possibly mediated by the increased AEA and 2-AG levels in the lungs (unpublished data). These data suggest that increasing the concentration of endocannabinoids in the airways by the inhibition of primary endocannabinoid degrading enzymes FAAH and MAGL can prevent airway hyperreactivity and airway inflammation. However, more studies should be performed in order to clarify the effects of FAAH and MAGL inhibitors in the airways since different fatty amino acids are also substrates for these enzymes.
The endocannabinoid system also contributes to pulmonary fibrosis, which is a life-threatening disease. Endocannabinoids are shown to promote the progression of fibrosis in liver [122,123,124], kidney [125,126], heart [127], and skin [128]. Studies in mice suggest that CB1 receptors are associated with radiation-induced pulmonary fibrosis [129]. Cinar et al. performed a detailed study in order to identify the contribution of CB1 receptors in lung fibrosis, and have shown that both the genetic deletion of CB1 receptors or their pharmacological inhibition with a CB1 antagonist can attenuate lung inflammation and fibrosis, and hence increase animal survival in a mouse model of radiation-induced pulmonary fibrosis [130]. This finding indicates a different pattern than other studies in which cannabinoid CB1 receptor stimulation has been raised as a therapeutic approach, as discussed above. This controversy may be due to the models used to induce fibrosis and related pathophysiology of fibrosis, which is different from acute or allergic airway inflammation. However, although CB1 activation can prevent airway hyperreactivity and inflammation, CB2 receptor antagonism has also been suggested as a therapeutic strategy for allergic diseases [78,118].
4. Conclusions
The studies about the contribution of the endocannabinoid system in the airways indicate the importance of both CB1 and CB2 receptors (Table 1). Among these two receptors, CB1 subtype is more likely to be involved in the functional reactivity of the airways as its stimulation can inhibit the contraction of airway smooth muscle. This effect seems to be mediated by the inhibition of acetylcholine release from cholinergic nerves, rather than a direct effect on the smooth muscle itself. Unlike CB1 receptors, CB2 receptors are likely to be involved in the mechanisms for neurogenic inflammation, probably acting through the sensory nerves (Figure 1). The contribution of both receptors in the immune modulation of airways is well established, as discussed above. The data of the present literature indicate a significant contribution of CB2 receptors in allergic diseases, which can be considered for the treatment of allergic asthma. However, the possible involvement of CB1 receptors should not be excluded, since they are expressed and functional almost in every immune cell. Therefore, appropriate cannabinoid receptor ligands may be rational candidates for the treatment of airway diseases because of their anti-inflammatory and bronchodilatory effects.
Altering the tissue levels of endocannabinoids as a therapeutic strategy is a complex issue. One reason for that is cannabinoids have other targets then CB receptors, which can cause unpredictable effects. The enzymes responsible for endocannabinoid metabolism also have other bioactive by-products, which have significant biological effects. Furthermore, in addition to FAAH or MAGL, other enzymes are also involved in endocannabinoid metabolism, such as COX-2, which leads to the production of various inflammatory prostaglandins. In this respect, the elevation of AEA or 2-AG tissue levels by FAAH or MAGL inhibition were shown to enhance their COX-2-mediated oxidation, and increase prostamide and PG-G signaling [131,132]. These factors make the therapeutic use of FAAH and MAGL inhibitors more complicated, and may partly account for the failure of some FAAH inhibitors like PF-04457845, BIA 10-2474, PF-06818883, V-158866 in clinical trials [6,133,134,135]. For instance, the reasons for the interruption of the clinical trial of BIA 10–2474 due to a death of a volunteer are still not fully established and thought to be related to the off-target effects of FAAH inhibitors [134,135]. However, elevated endocannabinoid levels were not reported to be associated with severe toxic effects on the central nervous system [134]. The specificity of the molecule itself or its metabolite(s) should be considered for the off-target effects, which can cause unpredictable adverse events in clinical trials. Therefore, more studies should be performed in order to clarify the contribution of by-products and other bioactive fatty acids in airway inflammation.
Funding
This research received no external funding.
Conflicts of Interest
The author declare no conflict of interest.
References
- Gaoni, Y.; Mechoulam, R. Isolation, Structure, and Partial Synthesis of an Active Constituent of Hashish. J. Am. Chem. Soc. 1964, 86, 1646–1647. [Google Scholar] [CrossRef]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science (NY) 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R.; et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharm. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Izzo, A.A.; Camilleri, M. Emerging role of cannabinoids in gastrointestinal and liver diseases: Basic and clinical aspects. Gut 2008, 57, 1140–1155. [Google Scholar] [CrossRef]
- Di Marzo, V.; Fontana, A. Anandamide, an endogenous cannabinomimetic eicosanoid: ’killing two birds with one stone’. Prostaglandinsleukot. Essent. Fat. Acids. 1995, 53, 1–11. [Google Scholar] [CrossRef]
- Di Marzo, V. New approaches and challenges to targeting the endocannabinoid system. Nat. Rev. Drug. Discov. 2018, 17, 623–639. [Google Scholar] [CrossRef]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharm. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Cravatt, B.F.; Giang, D.K.; Mayfield, S.P.; Boger, D.L.; Lerner, R.A.; Gilula, N.B. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 1996, 384, 83–87. [Google Scholar] [CrossRef]
- Sun, Y.X.; Tsuboi, K.; Zhao, L.Y.; Okamoto, Y.; Lambert, D.M.; Ueda, N. Involvement of N-acylethanolamine-hydrolyzing acid amidase in the degradation of anandamide and other N-acylethanolamines in macrophages. Biochim. Biophys. Acta 2005, 1736, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Dinh, T.P.; Carpenter, D.; Leslie, F.M.; Freund, T.F.; Katona, I.; Sensi, S.L.; Kathuria, S.; Piomelli, D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc. Natl. Acad. Sci. USA 2002, 99, 10819–10824. [Google Scholar] [CrossRef] [Green Version]
- Szafran, B.N.; Lee, J.H.; Borazjani, A.; Morrison, P.; Zimmerman, G.; Andrzejewski, K.L.; Ross, M.K.; Kaplan, B.L.F. Characterization of Endocannabinoid-Metabolizing Enzymes in Human Peripheral Blood Mononuclear Cells under Inflammatory Conditions. Molecules 2018, 23, 3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Borazjani, A.; Matthews, A.T.; Mangum, L.C.; Edelmann, M.J.; Ross, M.K. Identification of palmitoyl protein thioesterase 1 in human THP1 monocytes and macrophages and characterization of unique biochemical activities for this enzyme. Biochemistry 2013, 52, 7559–7574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuo, W.; Leleu-Chavain, N.; Spencer, J.; Sansook, S.; Millet, R.; Chavatte, P. Therapeutic Potential of Fatty Acid Amide Hydrolase, Monoacylglycerol Lipase, and N-Acylethanolamine Acid Amidase Inhibitors. J. Med. Chem. 2017, 60, 4–46. [Google Scholar] [CrossRef] [PubMed]
- Piomelli, D.; Sasso, O. Peripheral gating of pain signals by endogenous lipid mediators. Nat. Neurosci. 2014, 17, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Rouzer, C.A.; Marnett, L.J. Endocannabinoid oxygenation by cyclooxygenases, lipoxygenases, and cytochromes P450: Cross-talk between the eicosanoid and endocannabinoid signaling pathways. Chem. Rev. 2011, 111, 5899–5921. [Google Scholar] [CrossRef]
- Devane, W.A.; Dysarz, F.A., 3rd; Johnson, M.R.; Melvin, L.S.; Howlett, A.C. Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharm. 1988, 34, 605–613. [Google Scholar]
- De Petrocellis, L.; Nabissi, M.; Santoni, G.; Ligresti, A. Actions and Regulation of Ionotropic Cannabinoid Receptors. Adv. Pharm. 2017, 80, 249–289. [Google Scholar]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.Y.; Lu, H.C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [Green Version]
- Ryberg, E.; Larsson, N.; Sjogren, S.; Hjorth, S.; Hermansson, N.O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharm. 2007, 152, 1092–1101. [Google Scholar] [CrossRef]
- McHugh, D.; Page, J.; Dunn, E.; Bradshaw, H.B. Delta(9) -Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharm. 2012, 165, 2414–2424. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Huang, B.X.; Kwon, H.; Rashid, M.A.; Kharebava, G.; Desai, A.; Patnaik, S.; Marugan, J.; Kim, H.Y. Orphan GPR110 (ADGRF1) targeted by N-docosahexaenoylethanolamine in development of neurons and cognitive function. Nat. Commun. 2016, 7, 13123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.J. Novel cannabinoid receptors. Br. J. Pharm. 2007, 152, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Onaivi, E.S.; Chaudhuri, G. Cloning and sequencing of a cDNA encoding the mouse brain-type cannabinoid receptor protein. Dna. Seq. 1995, 5, 385–388. [Google Scholar] [CrossRef]
- Shire, D.; Calandra, B.; Rinaldi-Carmona, M.; Oustric, D.; Pessegue, B.; Bonnin-Cabanne, O.; Le Fur, G.; Caput, D.; Ferrara, P. Molecular cloning, expression and function of the murine CB2 peripheral cannabinoid receptor. Biochim. Biophys. Acta 1996, 1307, 132–136. [Google Scholar] [CrossRef]
- Demuth, D.G.; Molleman, A. Cannabinoid signalling. Life Sci. 2006, 78, 549–563. [Google Scholar] [CrossRef]
- Howlett, A.C. Cannabinoid receptor signaling. Handb. Exp. Pharm. 2005, 168, 53–79. [Google Scholar]
- Rodriguez de Fonseca, F.; Del Arco, I.; Bermudez-Silva, F.J.; Bilbao, A.; Cippitelli, A.; Navarro, M. The endocannabinoid system: Physiology and pharmacology. Alcohol. Alcohol. (Oxf. Oxfs.) 2005, 40, 2–14. [Google Scholar] [CrossRef]
- Bouaboula, M.; Rinaldi, M.; Carayon, P.; Carillon, C.; Delpech, B.; Shire, D.; Le Fur, G.; Casellas, P. Cannabinoid-receptor expression in human leukocytes. Eur. J. Biochem. 1993, 214, 173–180. [Google Scholar] [CrossRef]
- Howlett, A.C. The cannabinoid receptors. Prostaglandins. Other. Lipid. Mediat. 2002, 68–69, 619–631. [Google Scholar] [CrossRef]
- Gong, J.P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 receptors: Immunohistochemical localization in rat brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.-P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. FUNCTIONAL EXPRESSION OF BRAIN NEURONAL CB2 CANNABINOID RECEPTORS ARE INVOLVED IN THE EFFECTS OF DRUGS OF ABUSE AND IN DEPRESSION. Ann. NY Acad. Sci. 2008, 1139, 434–449. [Google Scholar] [CrossRef] [Green Version]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Ofek, O.; Karsak, M.; Leclerc, N.; Fogel, M.; Frenkel, B.; Wright, K.; Tam, J.; Attar-Namdar, M.; Kram, V.; Shohami, E.; et al. Peripheral cannabinoid receptor, CB2, regulates bone mass. Proc. Natl. Acad. Sci. USA 2006, 103, 696–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacher, P.; Kogan, N.M.; Mechoulam, R. Beyond THC and Endocannabinoids. Annu. Rev. Pharm. Toxicol. 2019, 60, 32.1–32.23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdyshev, E.V. Cannabinoid receptors and the regulation of immune response. Chem. Phys. Lipids 2000, 108, 169–190. [Google Scholar] [CrossRef]
- D’Argenio, G.; Valenti, M.; Scaglione, G.; Cosenza, V.; Sorrentini, I.; Di Marzo, V. Up-regulation of anandamide levels as an endogenous mechanism and a pharmacological strategy to limit colon inflammation. FASEB J. 2006, 20, 568–570. [Google Scholar] [CrossRef]
- Kurabayashi, M.; Takeyoshi, I.; Yoshinari, D.; Matsumoto, K.; Maruyama, I.; Morishita, Y. 2-Arachidonoylglycerol increases in ischemia-reperfusion injury of the rat liver. J. Invest. Surg. 2005, 18, 25–31. [Google Scholar] [CrossRef]
- Muthian, S.; Rademacher, D.J.; Roelke, C.T.; Gross, G.J.; Hillard, C.J. Anandamide content is increased and CB1 cannabinoid receptor blockade is protective during transient, focal cerebral ischemia. Neuroscience 2004, 129, 743–750. [Google Scholar] [CrossRef]
- Panikashvili, D.; Simeonidou, C.; Ben-Shabat, S.; Hanus, L.; Breuer, A.; Mechoulam, R.; Shohami, E. An endogenous cannabinoid (2-AG) is neuroprotective after brain injury. Nature 2001, 413, 527–531. [Google Scholar] [CrossRef]
- Sugamura, K.; Sugiyama, S.; Nozaki, T.; Matsuzawa, Y.; Izumiya, Y.; Miyata, K.; Nakayama, M.; Kaikita, K.; Obata, T.; Takeya, M.; et al. Activated endocannabinoid system in coronary artery disease and antiinflammatory effects of cannabinoid 1 receptor blockade on macrophages. Circulation 2009, 119, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, V.L.; Hegde, S.; Cravatt, B.F.; Hofseth, L.J.; Nagarkatti, M.; Nagarkatti, P.S. Attenuation of experimental autoimmune hepatitis by exogenous and endogenous cannabinoids: Involvement of regulatory T cells. Mol. Pharm. 2008, 74, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Berdyshev, E.; Boichot, E.; Corbel, M.; Germain, N.; Lagente, V. Effects of cannabinoid receptor ligands on LPS-induced pulmonary inflammation in mice. Life Sci. 1998, 63, PL125–PL129. [Google Scholar] [CrossRef]
- Comelli, F.; Giagnoni, G.; Bettoni, I.; Colleoni, M.; Costa, B. The inhibition of monoacylglycerol lipase by URB602 showed an anti-inflammatory and anti-nociceptive effect in a murine model of acute inflammation. Br. J. Pharm. 2007, 152, 787–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csoka, B.; Nemeth, Z.H.; Mukhopadhyay, P.; Spolarics, Z.; Rajesh, M.; Federici, S.; Deitch, E.A.; Batkai, S.; Pacher, P.; Hasko, G. CB2 cannabinoid receptors contribute to bacterial invasion and mortality in polymicrobial sepsis. PLoS ONE 2009, 4, e6409. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.A.; Kellermann, C.A.; Burnat, G.; Hahn, E.G.; Rau, T.; Konturek, P.C. Mice lacking cannabinoid CB1-, CB2-receptors or both receptors show increased susceptibility to trinitrobenzene sulfonic acid (TNBS)-induced colitis. J. Physiol. Pharm. 2010, 61, 89–97. [Google Scholar]
- Kaminski, N.E.; Koh, W.S.; Yang, K.H.; Lee, M.; Kessler, F.K. Suppression of the humoral immune response by cannabinoids is partially mediated through inhibition of adenylate cyclase by a pertussis toxin-sensitive G-protein coupled mechanism. Biochem. Pharm. 1994, 48, 1899–1908. [Google Scholar] [CrossRef]
- Titishov, N.; Mechoulam, R.; Zimmerman, A.M. Stereospecific effects of (-)- and (+)-7-hydroxy-delta-6-tetrahydrocannabinol-dimethylheptyl on the immune system of mice. Pharmacology 1989, 39, 337–349. [Google Scholar] [CrossRef]
- Lu, T.; Newton, C.; Perkins, I.; Friedman, H.; Klein, T.W. Role of cannabinoid receptors in Delta-9-tetrahydrocannabinol suppression of IL-12p40 in mouse bone marrow-derived dendritic cells infected with Legionella pneumophila. Eur. J. Pharmacol. 2006, 532, 170–177. [Google Scholar] [CrossRef]
- Klein, T.W.; Lane, B.; Newton, C.A.; Friedman, H. The cannabinoid system and cytokine network. Proc. Soc. Exp. Biol. Med. 2000, 225, 1–8. [Google Scholar] [CrossRef]
- Klein, T.W.; Newton, C.A.; Nakachi, N.; Friedman, H. Delta 9-tetrahydrocannabinol treatment suppresses immunity and early IFN-gamma, IL-12, and IL-12 receptor beta 2 responses to Legionella pneumophila infection. J. Immunol. (Baltimore, Md. 1950) 2000, 164, 6461–6466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKallip, R.J.; Nagarkatti, M.; Nagarkatti, P.S. Delta-9-tetrahydrocannabinol enhances breast cancer growth and metastasis by suppression of the antitumor immune response. J. Immunol. (Baltimore, Md. 1950) 2005, 174, 3281–3289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishkin, E.M.; Cabral, G.A. delta-9-Tetrahydrocannabinol decreases host resistance to herpes simplex virus type 2 vaginal infection in the B6C3F1 mouse. J. Gen. Virol. 1985, 66, 2539–2549. [Google Scholar] [CrossRef] [PubMed]
- Carayon, P.; Marchand, J.; Dussossoy, D.; Derocq, J.M.; Jbilo, O.; Bord, A.; Bouaboula, M.; Galiegue, S.; Mondiere, P.; Penarier, G.; et al. Modulation and functional involvement of CB2 peripheral cannabinoid receptors during B-cell differentiation. Blood 1998, 92, 3605–3615. [Google Scholar] [CrossRef]
- Sacerdote, P.; Massi, P.; Panerai, A.E.; Parolaro, D. In vivo and in vitro treatment with the synthetic cannabinoid CP55, 940 decreases the in vitro migration of macrophages in the rat: Involvement of both CB1 and CB2 receptors. J. Neuroimmunol. 2000, 109, 155–163. [Google Scholar] [CrossRef]
- McCoy, K.L.; Matveyeva, M.; Carlisle, S.J.; Cabral, G.A. Cannabinoid inhibition of the processing of intact lysozyme by macrophages: Evidence for CB2 receptor participation. J. Pharm. Exp. 1999, 289, 1620–1625. [Google Scholar]
- Vannacci, A.; Giannini, L.; Passani, M.B.; Di Felice, A.; Pierpaoli, S.; Zagli, G.; Fantappie, O.; Mazzanti, R.; Masini, E.; Mannaioni, P.F. The endocannabinoid 2-arachidonylglycerol decreases the immunological activation of Guinea pig mast cells: Involvement of nitric oxide and eicosanoids. J. Pharm. Exp. 2004, 311, 256–264. [Google Scholar] [CrossRef]
- Samson, M.T.; Small-Howard, A.; Shimoda, L.M.; Koblan-Huberson, M.; Stokes, A.J.; Turner, H. Differential roles of CB1 and CB2 cannabinoid receptors in mast cells. J. Immunol. (Baltimore, Md. 1950) 2003, 170, 4953–4962. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, K.; Biro, T.; Tsuruta, D.; Toth, B.I.; Kromminga, A.; Zakany, N.; Zimmer, A.; Funk, W.; Gibbs, B.F.; Zimmer, A.; et al. Endocannabinoids limit excessive mast cell maturation and activation in human skin. J. Allergy Clin. Immunol. 2012, 129, 726–738.e8. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, K.; Zakany, N.; Hundt, T.; Emelianov, V.; Tsuruta, D.; Schafer, C.; Kloepper, J.E.; Biro, T.; Paus, R. Cannabinoid receptor 1 controls human mucosal-type mast cell degranulation and maturation in situ. J. Allergy Clin. Immunol. 2013, 132, 182–193. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, S.E. Cannabinoids go nuclear: Evidence for activation of peroxisome proliferator-activated receptors. Br. J. Pharm. 2007, 152, 576–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Alexander, S.P.; Kendall, D.A.; Bennett, A.J. Cannabinoids and PPARalpha signalling. Biochem. Soc. Trans. 2006, 34, 1095–1097. [Google Scholar] [CrossRef] [PubMed]
- Moir, D.; Rickert, W.S.; Levasseur, G.; Larose, Y.; Maertens, R.; White, P.; Desjardins, S. A comparison of mainstream and sidestream marijuana and tobacco cigarette smoke produced under two machine smoking conditions. Chem. Res. Toxicol. 2008, 21, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Owen, K.P.; Sutter, M.E.; Albertson, T.E. Marijuana: Respiratory tract effects. Clin. Rev. Allergy Immunol. 2014, 46, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Gong, H., Jr.; Fligiel, S.; Tashkin, D.P.; Barbers, R.G. Tracheobronchial changes in habitual, heavy smokers of marijuana with and without tobacco. Am. Rev. Respir. Dis. 1987, 136, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Fligiel, S.E.; Beals, T.F.; Tashkin, D.P.; Paule, M.G.; Scallet, A.C.; Ali, S.F.; Bailey, J.R.; Slikker, W., Jr. Marijuana exposure and pulmonary alterations in primates. Pharm. Biochem. Behav. 1991, 40, 637–642. [Google Scholar] [CrossRef]
- Giannini, L.; Nistri, S.; Mastroianni, R.; Cinci, L.; Vannacci, A.; Mariottini, C.; Passani, M.B.; Mannaioni, P.F.; Bani, D.; Masini, E. Activation of cannabinoid receptors prevents antigen-induced asthma-like reaction in guinea pigs. J. Cell Mol. Med. 2008, 12, 2381–2394. [Google Scholar] [CrossRef] [Green Version]
- Zoerner, A.A.; Stichtenoth, D.O.; Engeli, S.; Batkai, S.; Winkler, C.; Schaumann, F.; Janke, J.; Holz, O.; Krug, N.; Tsikas, D.; et al. Allergen challenge increases anandamide in bronchoalveolar fluid of patients with allergic asthma. Clin. Pharm. 2011, 90, 388–391. [Google Scholar] [CrossRef]
- Oka, S.; Ikeda, S.; Kishimoto, S.; Gokoh, M.; Yanagimoto, S.; Waku, K.; Sugiura, T. 2-arachidonoylglycerol, an endogenous cannabinoid receptor ligand, induces the migration of EoL-1 human eosinophilic leukemia cells and human peripheral blood eosinophils. J. Leukoc Biol. 2004, 76, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- Chouinard, F.; Lefebvre, J.S.; Navarro, P.; Bouchard, L.; Ferland, C.; Lalancette-Hebert, M.; Marsolais, D.; Laviolette, M.; Flamand, N. The endocannabinoid 2-arachidonoyl-glycerol activates human neutrophils: Critical role of its hydrolysis and de novo leukotriene B4 biosynthesis. J. Immunol. (Baltimore, Md. 1950) 2011, 186, 3188–3196. [Google Scholar] [CrossRef] [Green Version]
- Chouinard, F.; Turcotte, C.; Guan, X.; Larose, M.C.; Poirier, S.; Bouchard, L.; Provost, V.; Flamand, L.; Grandvaux, N.; Flamand, N. 2-Arachidonoyl-glycerol- and arachidonic acid-stimulated neutrophils release antimicrobial effectors against E. coli, S. aureus, HSV-1, and RSV. J. Leukoc Biol. 2013, 93, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montecucco, F.; Burger, F.; Mach, F.; Steffens, S. CB2 cannabinoid receptor agonist JWH-015 modulates human monocyte migration through defined intracellular signaling pathways. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1145–H1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.H.; Lim, S.; Ryu, J.; Lee, C.W.; Kim, Y.; Kang, J.H.; Kang, S.S.; Ahn, Y.K.; Park, C.S.; Kim, J.J. CB1 and CB2 cannabinoid receptors differentially regulate the production of reactive oxygen species by macrophages. Cardiovasc. Res. 2009, 84, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, J.T.; Harui, A.; Kiertscher, S.M.; Roth, J.D.; Roth, M.D. Differential expression of intracellular and extracellular CB(2) cannabinoid receptor protein by human peripheral blood leukocytes. J. Neuroimmune Pharm. 2013, 8, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, C.; Blanchet, M.R.; Laviolette, M.; Flamand, N. Impact of Cannabis, Cannabinoids, and Endocannabinoids in the Lungs. Front. Pharm. 2016, 7, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, S.; Oka, S.; Gokoh, M.; Sugiura, T. Chemotaxis of human peripheral blood eosinophils to 2-arachidonoylglycerol: Comparison with other eosinophil chemoattractants. Int. Arch. Allergy Immunol. 2006, 140, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Larose, M.C.; Turcotte, C.; Chouinard, F.; Ferland, C.; Martin, C.; Provost, V.; Laviolette, M.; Flamand, N. Mechanisms of human eosinophil migration induced by the combination of IL-5 and the endocannabinoid 2-arachidonoyl-glycerol. J. Allergy Clin. Immunol. 2014, 133, 1480–1482. [Google Scholar] [CrossRef]
- Frei, R.B.; Luschnig, P.; Parzmair, G.P.; Peinhaupt, M.; Schranz, S.; Fauland, A.; Wheelock, C.E.; Heinemann, A.; Sturm, E.M. Cannabinoid receptor 2 augments eosinophil responsiveness and aggravates allergen-induced pulmonary inflammation in mice. Allergy 2016, 71, 944–956. [Google Scholar] [CrossRef]
- Matias, I.; Pochard, P.; Orlando, P.; Salzet, M.; Pestel, J.; Di Marzo, V. Presence and regulation of the endocannabinoid system in human dendritic cells. Eur. J. Biochem. 2002, 269, 3771–3778. [Google Scholar] [CrossRef] [Green Version]
- Do, Y.; McKallip, R.J.; Nagarkatti, M.; Nagarkatti, P.S. Activation through cannabinoid receptors 1 and 2 on dendritic cells triggers NF-kappaB-dependent apoptosis: Novel role for endogenous and exogenous cannabinoids in immunoregulation. J. Immunol. (Baltimore, Md. 1950) 2004, 173, 2373–2382. [Google Scholar] [CrossRef] [Green Version]
- Kurihara, R.; Tohyama, Y.; Matsusaka, S.; Naruse, H.; Kinoshita, E.; Tsujioka, T.; Katsumata, Y.; Yamamura, H. Effects of peripheral cannabinoid receptor ligands on motility and polarization in neutrophil-like HL60 cells and human neutrophils. J. Biol. Chem. 2006, 281, 12908–12918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashkin, D.P.; Shapiro, B.J.; Frank, I.M. Acute pulmonary physiologic effects of smoked marijuana and oral (Delta)9 -tetrahydrocannabinol in healthy young men. N. Engl. J. Med. 1973, 289, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Vachon, L.; FitzGerald, M.X.; Solliday, N.H.; Gould, I.A.; Gaensler, E.A. Single-dose effects of marihuana smoke. Bronchial dynamics and respiratory-center sensitivity in normal subjects. N. Engl. J. Med. 1973, 288, 985–989. [Google Scholar] [CrossRef] [PubMed]
- Tashkin, D.P.; Shapiro, B.J.; Frank, I.M. Acute effects of smoked marijuana and oral delta9-tetrahydrocannabinol on specific airway conductance in asthmatic subjects. Am. Rev. Respir. Dis. 1974, 109, 420–428. [Google Scholar]
- Tashkin, D.P.; Shapiro, B.J.; Lee, Y.E.; Harper, C.E. Effects of smoked marijuana in experimentally induced asthma. Am. Rev. Respir. Dis. 1975, 112, 377–386. [Google Scholar]
- Hartley, J.P.; Nogrady, S.G.; Seaton, A. Bronchodilator effect of delta1-tetrahydrocannabinol. Br. J. Clin. Pharm. 1978, 5, 523–525. [Google Scholar] [CrossRef] [Green Version]
- Abboud, R.T.; Sanders, H.D. Effect of oral administration of delta-tetrahydrocannabinol on airway mechanics in normal and asthmatic subjects. Chest 1976, 70, 480–485. [Google Scholar] [CrossRef]
- Tashkin, D.P.; Reiss, S.; Shapiro, B.J.; Calvarese, B.; Olsen, J.L.; Lodge, J.W. Bronchial effects of aerosolized delta 9-tetrahydrocannabinol in healthy and asthmatic subjects. Am. Rev. Respir. Dis. 1977, 115, 57–65. [Google Scholar]
- Calignano, A.; Katona, I.; Desarnaud, F.; Giuffrida, A.; La Rana, G.; Mackie, K.; Freund, T.F.; Piomelli, D. Bidirectional control of airway responsiveness by endogenous cannabinoids. Nature 2000, 408, 96–101. [Google Scholar] [CrossRef]
- Yousif, M.H.; Oriowo, M.A. Inhibitory effects of cannabinoid receptor ligands on electrically-evoked responses in rat isolated tracheal ring segments. Pharm. Res. 1999, 40, 415–421. [Google Scholar] [CrossRef]
- Spicuzza, L.; Haddad, E.-B.; Birrell, M.; Ling, A.; Clarke, D.; Venkatesan, P.; Barnes, P.J.; Belvisi, M.G. Characterization of the effects of cannabinoids on guinea-pig tracheal smooth muscle tone: Role in the modulation of acetylcholine release from parasympathetic nerves. Br. J. Pharm. 2000, 130, 1720–1726. [Google Scholar] [CrossRef] [PubMed]
- Grassin-Delyle, S.; Naline, E.; Buenestado, A.; Faisy, C.; Alvarez, J.C.; Salvator, H.; Abrial, C.; Advenier, C.; Zemoura, L.; Devillier, P. Cannabinoids inhibit cholinergic contraction in human airways through prejunctional CB1 receptors. Br. J. Pharm. 2014, 171, 2767–2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozkurt, T.E.; Larsson, O.; Adner, M. Stimulation of cannabinoid CB1 receptors prevents nerve-mediated airway hyperreactivity in NGF-induced inflammation in mouse airways. Eur. J. Pharm. 2016, 776, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Batkai, S.; Rajesh, M.; Mukhopadhyay, P.; Hasko, G.; Liaudet, L.; Cravatt, B.F.; Csiszar, A.; Ungvari, Z.; Pacher, P. Decreased age-related cardiac dysfunction, myocardial nitrative stress, inflammatory gene expression, and apoptosis in mice lacking fatty acid amide hydrolase. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H909–H918. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Horvath, B.; Rajesh, M.; Matsumoto, S.; Saito, K.; Batkai, S.; Patel, V.; Tanchian, G.; Gao, R.Y.; Cravatt, B.F.; et al. Fatty acid amide hydrolase is a key regulator of endocannabinoid-induced myocardial tissue injury. Free Radic. Biol. Med. 2011, 50, 179–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Yao, H.; Cheng, Y.; Gong, D.; Liao, X.; Wang, R. Effects of monoacylglycerol lipase inhibitor URB602 on lung ischemia-reperfusion injury in mice. Biochem. Biophys. Res. Commun. 2018, 506, 578–584. [Google Scholar] [CrossRef]
- Yin, H.; Li, X.; Xia, R.; Yi, M.; Cheng, Y.; Wu, Y.; Ke, B.; Wang, R. Posttreatment with the Fatty Acid Amide Hydrolase Inhibitor URB937 Ameliorates One-Lung Ventilation-Induced Lung Injury in a Rabbit Model. J. Surg. Res. 2019, 239, 83–91. [Google Scholar] [CrossRef]
- Roviezzo, F.; Rossi, A.; Caiazzo, E.; Orlando, P.; Riemma, M.A.; Iacono, V.M.; Guarino, A.; Ialenti, A.; Cicala, C.; Peritore, A.; et al. Palmitoylethanolamide Supplementation during Sensitization Prevents Airway Allergic Symptoms in the Mouse. Front. Pharm. 2017, 8, 857. [Google Scholar] [CrossRef] [Green Version]
- Mechoulam, R.; Parker, L.A.; Gallily, R. Cannabidiol: An overview of some pharmacological aspects. J. Clin. Pharm. 2002, 42, 11S–19S. [Google Scholar] [CrossRef]
- Carrier, E.J.; Auchampach, J.A.; Hillard, C.J. Inhibition of an equilibrative nucleoside transporter by cannabidiol: A mechanism of cannabinoid immunosuppression. Proc. Natl. Acad. Sci. USA 2006, 103, 7895–7900. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, A.; Almeida, V.I.; Costola-de-Souza, C.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Vitoretti, L.B.; Gimenes-Junior, J.A.; Akamine, A.T.; Crippa, J.A.; Tavares-de-Lima, W.; et al. Cannabidiol improves lung function and inflammation in mice submitted to LPS-induced acute lung injury. Immunopharmacol. Immunotoxicol. 2015, 37, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Vitoretti, L.B.; Mariano-Souza, D.P.; Quinteiro-Filho, W.M.; Akamine, A.T.; Almeida, V.I.; Quevedo, J.; Dal-Pizzol, F.; et al. Cannabidiol, a non-psychotropic plant-derived cannabinoid, decreases inflammation in a murine model of acute lung injury: Role for the adenosine A(2A) receptor. Eur. J. Pharmacol. 2012, 678, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Vuolo, F.; Abreu, S.C.; Michels, M.; Xisto, D.G.; Blanco, N.G.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Reis, C.; Bahl, M.; et al. Cannabidiol reduces airway inflammation and fibrosis in experimental allergic asthma. Eur. J. Pharmacol. 2019, 843, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Dudasova, A.; Keir, S.D.; Parsons, M.E.; Molleman, A.; Page, C.P. The effects of cannabidiol on the antigen-induced contraction of airways smooth muscle in the guinea-pig. Pulm. Pharm. 2013, 26, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Huestis, M.A.; Solimini, R.; Pichini, S.; Pacifici, R.; Carlier, J.; Busardo, F.P. Cannabidiol Adverse Effects and Toxicity. Curr. Neuropharmacol. 2019, 17, 974–989. [Google Scholar] [CrossRef] [PubMed]
- Martin-Fontecha, M.; Eiwegger, T.; Jartti, T.; Rueda-Zubiaurre, A.; Tiringer, K.; Stepanow, J.; Puhakka, T.; Ruckert, B.; Ortega-Gutierrez, S.; Lopez-Rodriguez, M.L.; et al. The expression of cannabinoid receptor 1 is significantly increased in atopic patients. J. Allergy Clin. Immunol. 2014, 133, 926–929.e2. [Google Scholar] [CrossRef]
- Tahamtan, A.; Tavakoli-Yaraki, M.; Shadab, A.; Rezaei, F.; Marashi, S.M.; Shokri, F.; Mokhatri-Azad, T.; Salimi, V. The Role of Cannabinoid Receptor 1 in the Immunopathology of Respiratory Syncytial Virus. Viral. Immunol. 2018, 31, 292–298. [Google Scholar] [CrossRef]
- Tahamtan, A.; Samieipoor, Y.; Nayeri, F.S.; Rahbarimanesh, A.A.; Izadi, A.; Rashidi-Nezhad, A.; Tavakoli-Yaraki, M.; Farahmand, M.; Bont, L.; Shokri, F.; et al. Effects of cannabinoid receptor type 2 in respiratory syncytial virus infection in human subjects and mice. Virulence 2018, 9, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Abohalaka, R.; Bozkurt, T.E.; Onder, S.C.; Sahin-Erdemli, I. The effects of fatty acid amide hydrolase and monoacylglycerol lipase inhibitors in lipopolysaccharide-induced airway inflammation and tracheal hyperreactivity in mice. Eur. Respir. J. 2018, 52, PA5249. [Google Scholar]
- Belvisi, M.G.; Patel, H.J.; Freund-Michel, V.; Hele, D.J.; Crispino, N.; Birrell, M.A. Inhibitory activity of the novel CB2 receptor agonist, GW833972A, on guinea-pig and human sensory nerve function in the airways. Br. J. Pharm. 2008, 155, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Bozkurt, T.E.; Kaya, Y.; Durlu-Kandilci, N.T.; Onder, S.; Sahin-Erdemli, I. The effect of cannabinoids on dinitrofluorobenzene-induced experimental asthma in mice. Respir. Physiol. Neurobiol. 2016, 231, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Braun, A.; Engel, T.; Aguilar-Pimentel, J.A.; Zimmer, A.; Jakob, T.; Behrendt, H.; Mempel, M. Beneficial effects of cannabinoids (CB) in a murine model of allergen-induced airway inflammation: Role of CB1/CB2 receptors. Immunobiology 2011, 216, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.Y.; D’Agostino, B.; Risse, P.A.; Marrocco, G.; Naline, E.; Zhang, Y.; Chen, H.Z.; Finance, O.; Rinaldi-Carmona, M.; Rossi, F.; et al. Cannabinoid CB(2) receptor activation prevents bronchoconstriction and airway oedema in a model of gastro-oesophageal reflux. Eur. J. Pharm. 2007, 573, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Stengel, P.W.; Cockerham, S.L.; Silbaugh, S.A. Inhaled anandamide reduces leukotriene D4-induced airway obstruction in guinea pigs. Eur. J. Pharm. 2007, 557, 66–68. [Google Scholar] [CrossRef]
- Gkoumassi, E.; Dekkers, B.G.J.; Dröge, M.J.; Elzinga, C.R.S.; Schmidt, M.; Meurs, H.; Zaagsma, J.; Nelemans, S.A. Virodhamine and CP55,940 modulate cAMP production and IL-8 release in human bronchial epithelial cells. Br. J. Pharm. 2007, 151, 1041–1048. [Google Scholar] [CrossRef] [Green Version]
- Jan, T.-R.; Farraj, A.K.; Harkema, J.R.; Kaminski, N.E. Attenuation of the ovalbumin-induced allergic airway response by cannabinoid treatment in A/J mice. Toxicol. Appl. Pharm. 2003, 188, 24–35. [Google Scholar] [CrossRef]
- Patel, H.J.; Birrell, M.A.; Crispino, N.; Hele, D.J.; Venkatesan, P.; Barnes, P.J.; Yacoub, M.H.; Belvisi, M.G. Inhibition of guinea-pig and human sensory nerve activity and the cough reflex in guinea-pigs by cannabinoid (CB2) receptor activation. Br. J. Pharm. 2003, 140, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Ferrini, M.E.; Hong, S.; Stierle, A.; Stierle, D.; Stella, N.; Roberts, K.; Jaffar, Z. CB2 receptors regulate natural killer cells that limit allergic airway inflammation in a murine model of asthma. Allergy 2017, 72, 937–947. [Google Scholar] [CrossRef]
- Shang, V.C.; O’Sullivan, S.E.; Kendall, D.A.; Roberts, R.E. The endogenous cannabinoid anandamide increases human airway epithelial cell permeability through an arachidonic acid metabolite. Pharm. Res. 2016, 105, 152–163. [Google Scholar] [CrossRef]
- Wortley, M.A.; Adcock, J.J.; Dubuis, E.D.; Maher, S.A.; Bonvini, S.J.; Delescluse, I.; Kinloch, R.; McMurray, G.; Perros-Huguet, C.; Papakosta, M.; et al. Targeting fatty acid amide hydrolase as a therapeutic strategy for antitussive therapy. Eur. Respir. J. 2017, 50, 1700782. [Google Scholar] [CrossRef] [Green Version]
- Costola-de-Souza, C.; Ribeiro, A.; Ferraz-de-Paula, V.; Calefi, A.S.; Aloia, T.P.; Gimenes-Junior, J.A.; de Almeida, V.I.; Pinheiro, M.L.; Palermo-Neto, J. Monoacylglycerol lipase (MAGL) inhibition attenuates acute lung injury in mice. PLoS ONE 2013, 8, e77706. [Google Scholar] [CrossRef] [PubMed]
- Cinar, R.; Iyer, M.R.; Liu, Z.; Cao, Z.; Jourdan, T.; Erdelyi, K.; Godlewski, G.; Szanda, G.; Liu, J.; Park, J.K.; et al. Hybrid inhibitor of peripheral cannabinoid-1 receptors and inducible nitric oxide synthase mitigates liver fibrosis. JCI Insight 2016, 1, e87336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patsenker, E.; Stoll, M.; Millonig, G.; Agaimy, A.; Wissniowski, T.; Schneider, V.; Mueller, S.; Brenneisen, R.; Seitz, H.K.; Ocker, M.; et al. Cannabinoid receptor type I modulates alcohol-induced liver fibrosis. Mol. Med. 2011, 17, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Trebicka, J.; Racz, I.; Siegmund, S.V.; Cara, E.; Granzow, M.; Schierwagen, R.; Klein, S.; Wojtalla, A.; Hennenberg, M.; Huss, S.; et al. Role of cannabinoid receptors in alcoholic hepatic injury: Steatosis and fibrogenesis are increased in CB2 receptor-deficient mice and decreased in CB1 receptor knockouts. Liver. Int. 2011, 31, 860–870. [Google Scholar] [CrossRef]
- Lecru, L.; Desterke, C.; Grassin-Delyle, S.; Chatziantoniou, C.; Vandermeersch, S.; Devocelle, A.; Vernochet, A.; Ivanovski, N.; Ledent, C.; Ferlicot, S.; et al. Cannabinoid receptor 1 is a major mediator of renal fibrosis. Kidney Int. 2015, 88, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.L.; Hsu, Y.C.; Lee, P.H.; Lei, C.C.; Wang, J.Y.; Huang, Y.T.; Wang, S.Y.; Wang, F.S. Cannabinoid receptor 1 disturbance of PPARgamma2 augments hyperglycemia induction of mesangial inflammation and fibrosis in renal glomeruli. J. Mol. Med. (Berl.) 2014, 92, 779–792. [Google Scholar] [CrossRef]
- Slavic, S.; Lauer, D.; Sommerfeld, M.; Kemnitz, U.R.; Grzesiak, A.; Trappiel, M.; Thone-Reineke, C.; Baulmann, J.; Paulis, L.; Kappert, K.; et al. Cannabinoid receptor 1 inhibition improves cardiac function and remodelling after myocardial infarction and in experimental metabolic syndrome. J. Mol. Med. (Berl.) 2013, 91, 811–823. [Google Scholar] [CrossRef]
- Lazzerini, P.E.; Natale, M.; Gianchecchi, E.; Capecchi, P.L.; Montilli, C.; Zimbone, S.; Castrichini, M.; Balistreri, E.; Ricci, G.; Selvi, E.; et al. Adenosine A2A receptor activation stimulates collagen production in sclerodermic dermal fibroblasts either directly and through a cross-talk with the cannabinoid system. J. Mol. Med. (Berl.) 2012, 90, 331–342. [Google Scholar] [CrossRef]
- Bronova, I.; Smith, B.; Aydogan, B.; Weichselbaum, R.R.; Vemuri, K.; Erdelyi, K.; Makriyannis, A.; Pacher, P.; Berdyshev, E.V. Protection from Radiation-Induced Pulmonary Fibrosis by Peripheral Targeting of Cannabinoid Receptor-1. Am. J. Respir. Cell Mol. Biol. 2015, 53, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Cinar, R.; Gochuico, B.R.; Iyer, M.R.; Jourdan, T.; Yokoyama, T.; Park, J.K.; Coffey, N.J.; Pri-Chen, H.; Szanda, G.; Liu, Z.; et al. Cannabinoid CB1 receptor overactivity contributes to the pathogenesis of idiopathic pulmonary fibrosis. JCI Insight 2017, 2, e92281. [Google Scholar] [CrossRef]
- Valdeolivas, S.; Pazos, M.R.; Bisogno, T.; Piscitelli, F.; Iannotti, F.A.; Allara, M.; Sagredo, O.; Di Marzo, V.; Fernandez-Ruiz, J. The inhibition of 2-arachidonoyl-glycerol (2-AG) biosynthesis, rather than enhancing striatal damage, protects striatal neurons from malonate-induced death: A potential role of cyclooxygenase-2-dependent metabolism of 2-AG. Cell Death Dis. 2013, 4, e862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, A.; Ni, J.; Ling, K.H.; Acheampong, A.; Tang-Liu, D.D.; Burk, R.; Cravatt, B.F.; Woodward, D. Formation of prostamides from anandamide in FAAH knockout mice analyzed by HPLC with tandem mass spectrometry. J. Lipid. Res. 2004, 45, 757–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huggins, J.P.; Smart, T.S.; Langman, S.; Taylor, L.; Young, T. An efficient randomised, placebo-controlled clinical trial with the irreversible fatty acid amide hydrolase-1 inhibitor PF-04457845, which modulates endocannabinoids but fails to induce effective analgesia in patients with pain due to osteoarthritis of the knee. Pain 2012, 153, 1837–1846. [Google Scholar] [PubMed]
- Kerbrat, A.; Ferre, J.C.; Fillatre, P.; Ronziere, T.; Vannier, S.; Carsin-Nicol, B.; Lavoue, S.; Verin, M.; Gauvrit, J.Y.; Le Tulzo, Y.; et al. Acute Neurologic Disorder from an Inhibitor of Fatty Acid Amide Hydrolase. N. Engl. J. Med. 2016, 375, 1717–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Esbroeck, A.C.M.; Janssen, A.P.A.; Cognetta, A.B., 3rd; Ogasawara, D.; Shpak, G.; van der Kroeg, M.; Kantae, V.; Baggelaar, M.P.; de Vrij, F.M.S.; Deng, H.; et al. Activity-based protein profiling reveals off-target proteins of the FAAH inhibitor BIA 10-2474. Science (NY) 2017, 356, 1084–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staiano, R.I.; Loffredo, S.; Borriello, F.; Iannotti, F.A.; Piscitelli, F.; Orlando, P.; Secondo, A.; Granata, F.; Lepore, M.T.; Fiorelli, A.; et al. Human lung-resident macrophages express CB1 and CB2 receptors whose activation inhibits the release of angiogenic and lymphangiogenic factors. J. Leukoc. Biol. 2016, 99, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, S.H.; Reichenbach, N.L.; Fan, S.; Rom, S.; Merkel, S.F.; Wang, X.; Ho, W.Z.; Persidsky, Y. Attenuation of HIV-1 replication in macrophages by cannabinoid receptor 2 agonists. J. Leukoc. Biol. 2013, 93, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Small-Howard, A.L.; Shimoda, L.M.; Adra, C.N.; Turner, H. Anti-inflammatory potential of CB1-mediated cAMP elevation in mast cells. Biochem. J. 2005, 388, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Sanchez Lopez, A.J.; Roman-Vega, L.; Ramil Tojeiro, E.; Giuffrida, A.; Garcia-Merino, A. Regulation of cannabinoid receptor gene expression and endocannabinoid levels in lymphocyte subsets by interferon-beta: A longitudinal study in multiple sclerosis patients. Clin. Exp. Immunol. 2015, 179, 119–127. [Google Scholar] [CrossRef] [Green Version]
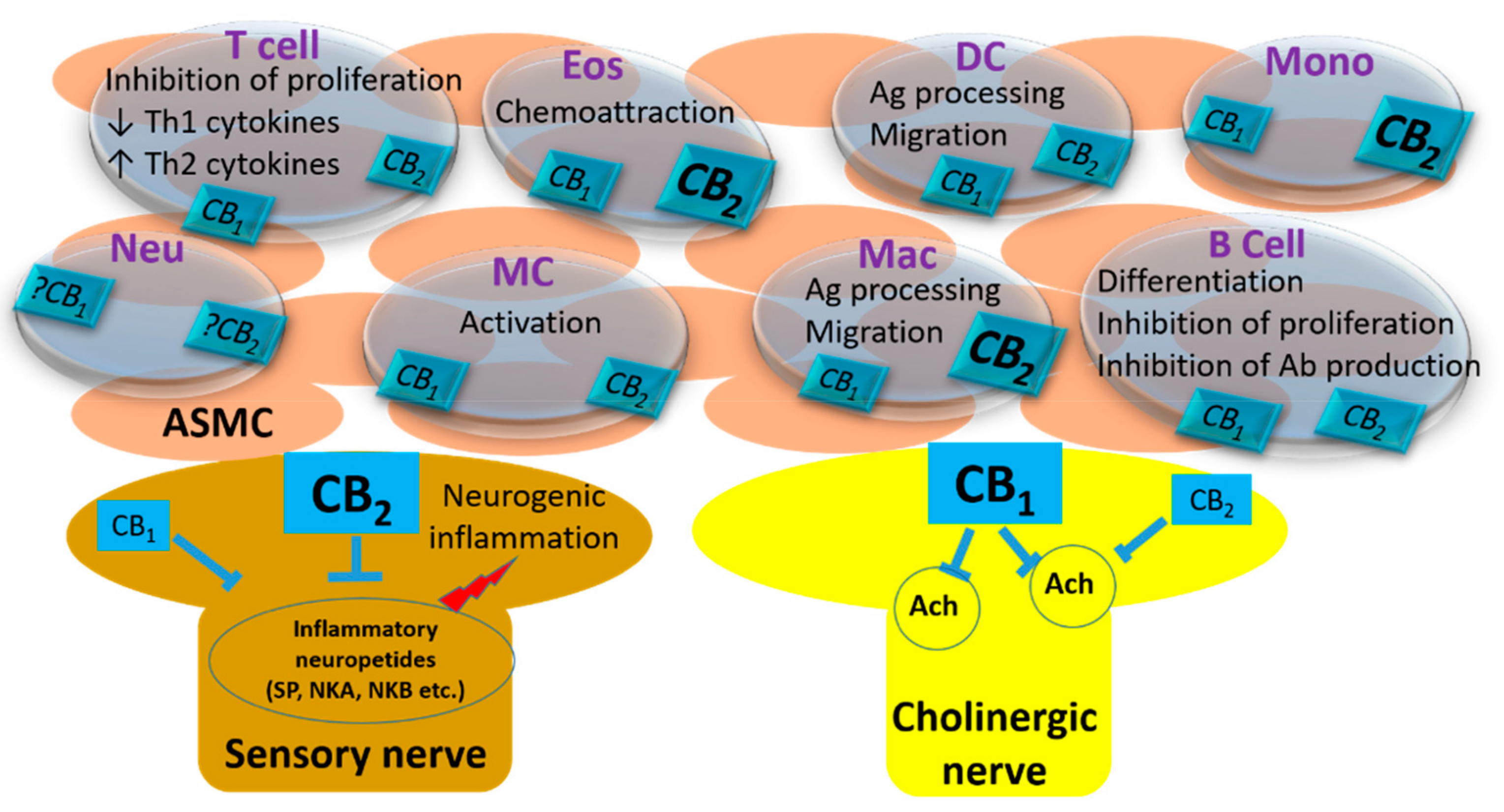
Figure 1.
The localization CB receptors and their reported effects in the airways. (ASMC: Airway smooth muscle cell; Eos: Eosinophil; DC: Dendritic cell; Mono: Monocyte; Neu: Neutrophil; MC: Mast cell; Mac: Macrophage; Ag: Antigen; Ab: Antibody). The receptors in bold characters represent a denser expression in that cell type.
Figure 1.
The localization CB receptors and their reported effects in the airways. (ASMC: Airway smooth muscle cell; Eos: Eosinophil; DC: Dendritic cell; Mono: Monocyte; Neu: Neutrophil; MC: Mast cell; Mac: Macrophage; Ag: Antigen; Ab: Antibody). The receptors in bold characters represent a denser expression in that cell type.

{kind=link}
Table 1.
Functional cannabinoid (CB) receptors in the airways and immune cells.
| Cell Type | Receptor | Species and Used Cells/Tissues in Related Studies |
|---|---|---|
| Alveolar Type-II epithelial cells | CB1 | Human [130] |
| Nerves | CB1 | Rat, Human [89,92] |
| Macrophages | CB1/CB2 | Human CD68(+), CD36(+) macrophages (CD: cluster of differentiation); THP-1 human monocytic cell line-derived macrophages; Murine RAW264.7 macrophage cell line [73] Human lung macrophages; lung cancer-associated macrophages [136] Human monocyte-derived macrophages [137] |
| Mast cells | CB1/CB2 | RBL2H3 mast cell line [58] Human CTS-mast cells [59] Human mucosal type mast cells [60] |
| Eosinophils | CB1/CB2 | EoL-1 cells [69] Human eosinophils from peripheral blood [69,70,71,138] |
| Dendritic cells | CB1/CB2 | Mouse bone marrow-derived dendritic cells [49,80] Human dendritic cells from peripheral blood [79] |
| Neutrophils | CB1/CB2 | Human promyelocytic cell line HL60 [33,81] Human neutrophils from peripheral blood [33,69,70,71,81] |
| B cells | CB1/CB2 | Human B cells from peripheral blood [33,54,74] Human tonsillar B cells [54] Human B lymphoblastoid cell line DAUDI [33] |
| T lymphocytes | CB1/CB2 | Human T cells from peripheral blood [74,139] |
| Basophils | CB1/CB2 | Human basophils from peripheral blood [138] |
| NK cells | CB1/CB2 | Human NK cells from peripheral blood [33] |
| Monocytes | CB1/CB2 | Human monocytes from peripheral blood [33,72,73,74] Human monocytic cell line U937 [33] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bozkurt, T.E. Endocannabinoid System in the Airways. Molecules 2019, 24, 4626. https://doi.org/10.3390/molecules24244626
AMA Style
Bozkurt TE. Endocannabinoid System in the Airways. Molecules. 2019; 24(24):4626. https://doi.org/10.3390/molecules24244626
Chicago/Turabian StyleBozkurt, Turgut Emrah. 2019. "Endocannabinoid System in the Airways" Molecules 24, no. 24: 4626. https://doi.org/10.3390/molecules24244626