Hispidulin Inhibits Mast Cell-Mediated Allergic Inflammation through Down-Regulation of Histamine Release and Inflammatory Cytokines


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
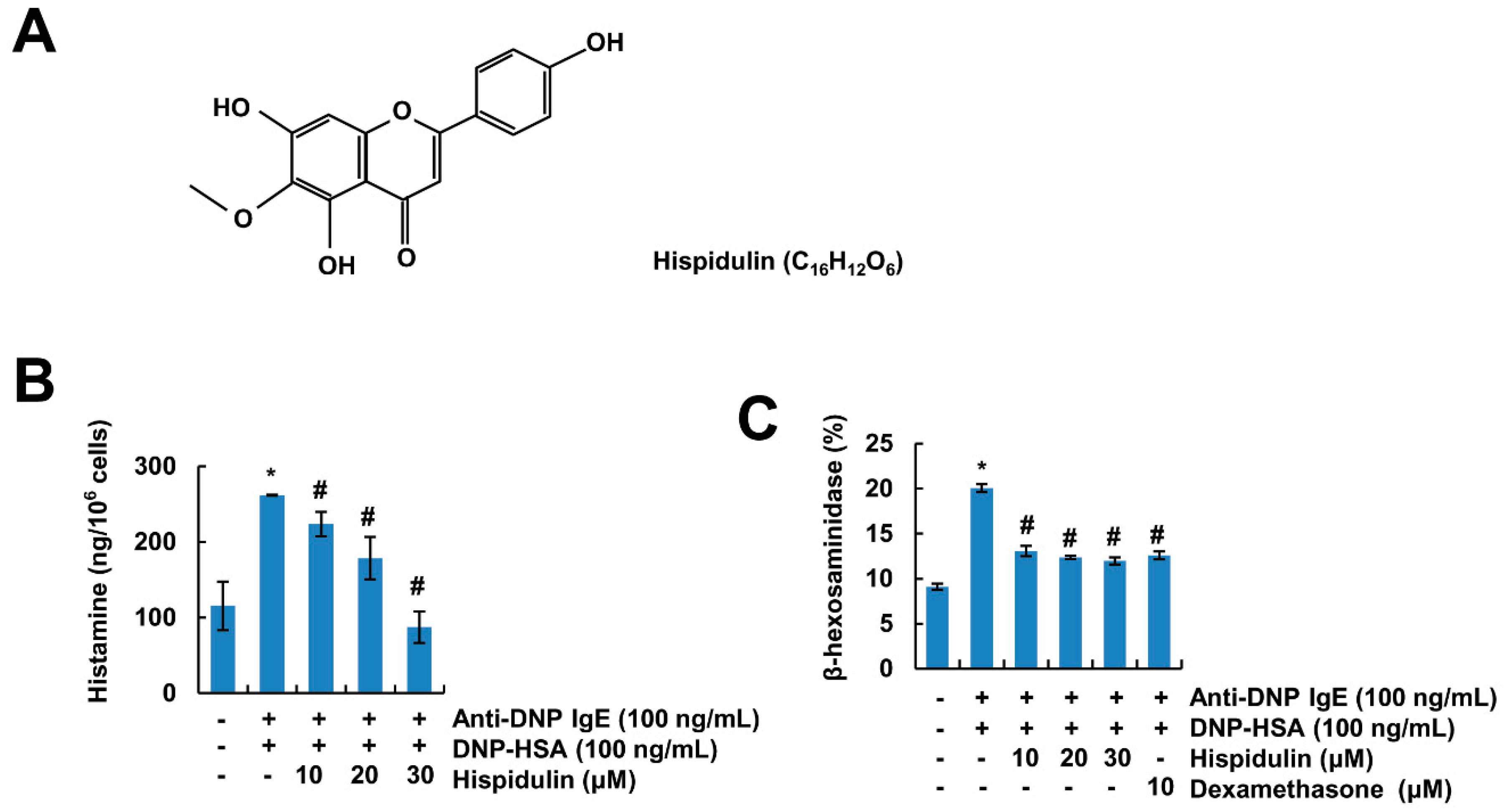
2.1. Hispidulin Inhibits Mast Cell Degranulation
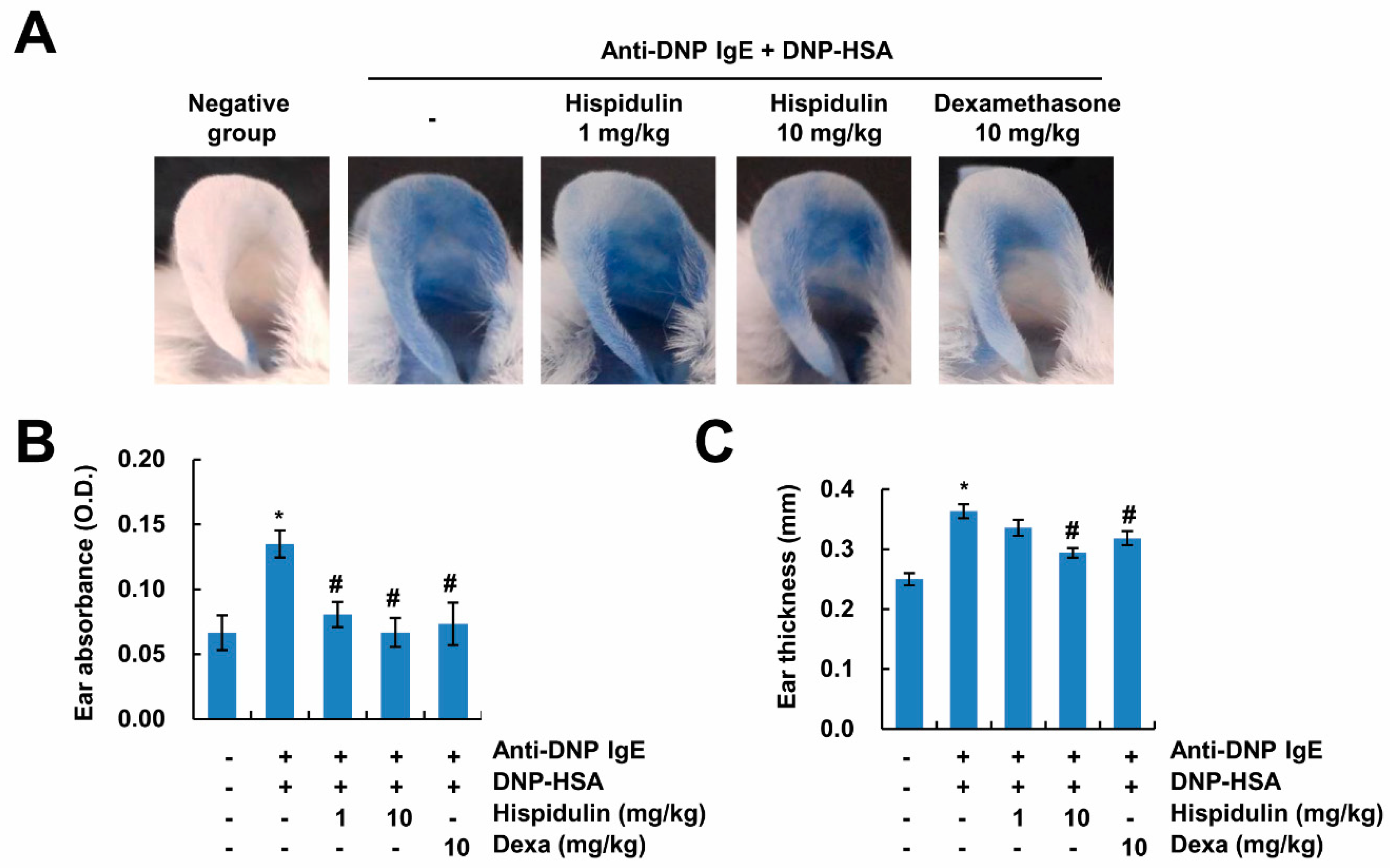
2.2. Hispidulin Reduces IgE-Mediated Local Cutaneous Anaphylaxis Reaction
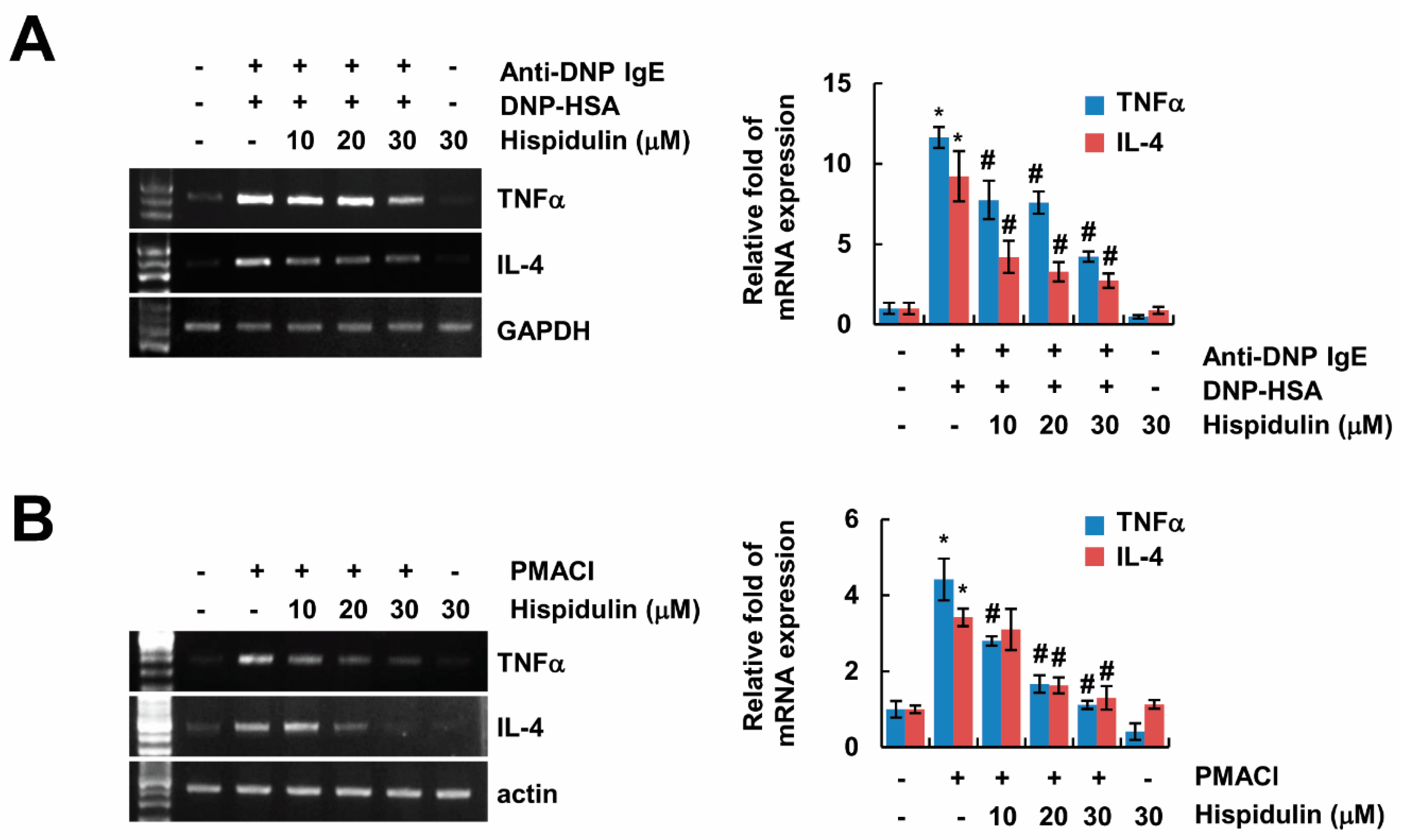
2.3. Hispidulin Inhibits Expression of Inflammatory Cytokines
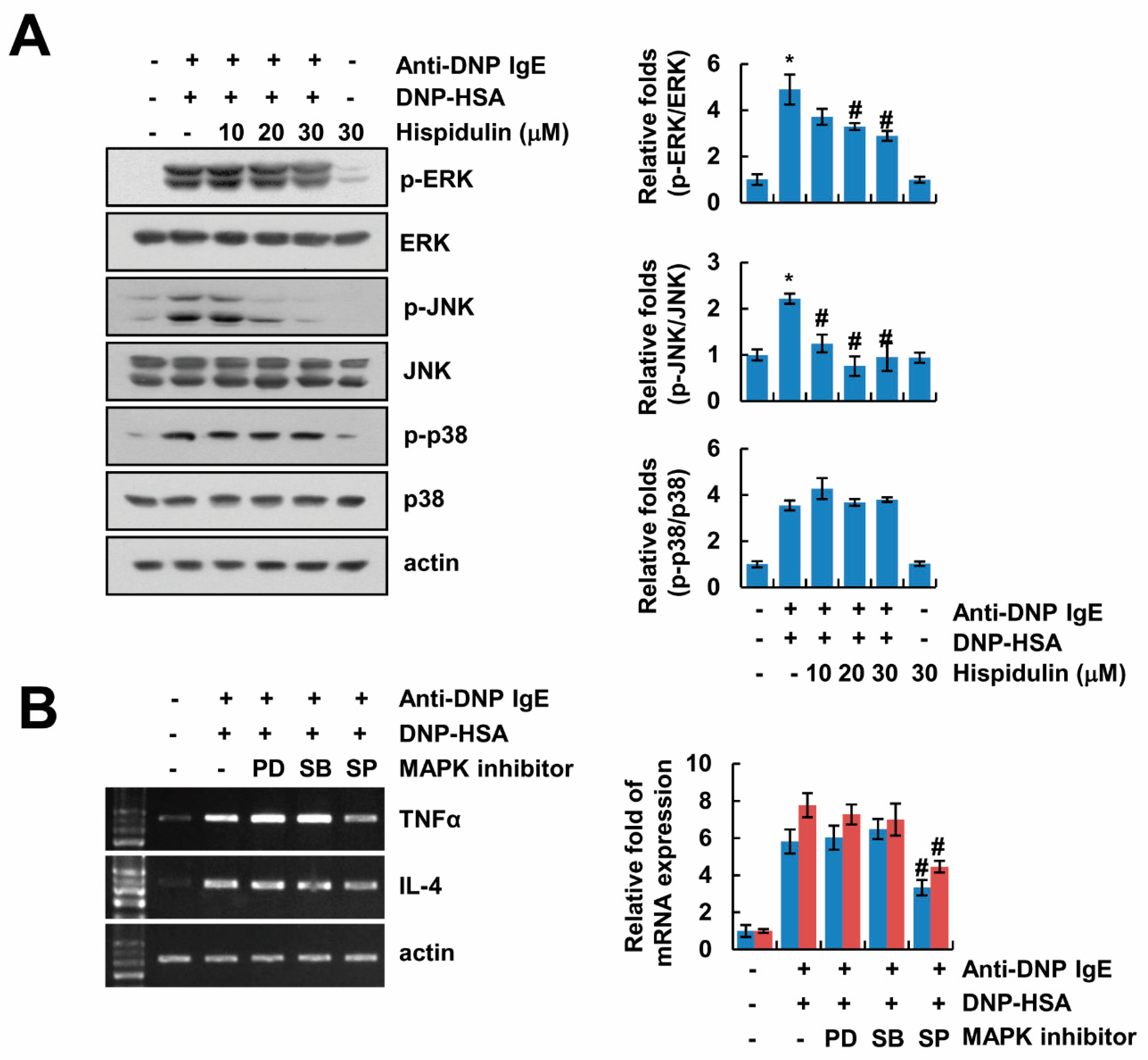
2.4. Hispidulin Inhibits Expression of Inflammatory Cytokines via Inhibition of JNK Phosphorylation
3. Discussion
4. Materials and Methods
4.1. Reagents and Cell Culture
4.2. Histamine Release
4.3. β-Hexosaminidase Release
4.4. Animals
4.5. Passive Cutaneous Anaphylaxis (PCA)
4.6. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TNF | Tumor necrosis factor |
| IL | Interleukin |
| HAS | Human serum albumin |
| PCA | Passive cutaneous anaphylaxis |
| MAPK | Mitogen-activated protein kinase |
References
- Abraham, S.N.; St John, A.L. Mast cell-orchestrated immunity to pathogens. Nat. Rev. Immunol. 2010, 10, 440–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.L.; Metcalfe, D.D. Mast cells in allergy and host defense. Allergy. Asthma. Proc. 2001, 22, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kempuraj, D.; Tagen, M.; Conti, P.; Kalogeromitros, D. Differential release of mast cell mediators and the pathogenesis of inflammation. Immunol. Rev. 2007, 217, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardamone, C.; Parente, R.; Feo, G.D.; Triggiani, M. Mast cells as effector cells of innate immunity and regulators of adaptive immunity. Immunol. Lett. 2016, 178, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J.; Gilfillan, A.M. Molecular regulation of mast cell activation. J. Allergy. Clin. Immunol. 2006, 117, 1214–1225. [Google Scholar] [CrossRef]
- Yin, Y.; Gong, F.Y.; Wu, X.X.; Sun, Y.; Li, Y.H.; Chen, T.; Xu, Q. Anti-inflammatory and immunosuppressive effect of flavones isolated from Artemisia vestita. J. Ethnopharmacol. 2008, 120, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Merfort, I. Methylated Flavonoids from Arnica montana and Arnica chamissonis. Planta Med. 1984, 50, 107–108. [Google Scholar] [CrossRef]
- Lin, T.Y.; Lu, C.W.; Wang, S.J.; Huang, S.K. Protective effect of hispidulin on kainic acid-induced seizures and neurotoxicity in rats. Eur. J. Pharm. 2015, 755, 6–15. [Google Scholar] [CrossRef]
- Tan, R.X.; Lu, H.; Wolfender, J.L.; Yu, T.T.; Zheng, W.F.; Yang, L.; Gafner, S.; Hostettmann, K. Mono- and sesquiterpenes and antifungal constituents from Artemisia species. Planta Med. 1999, 65, 64–67. [Google Scholar] [CrossRef]
- Atif, M.; Ali, I.; Hussain, A.; Hyder, S.V.; Niaz, B.; Khan, F.A.; Maalik, A.; Farooq, U. Pharmacological Assessment of Hispidulin--a Natural Bioactive Flavone. Acta Pol. Pharm. 2015, 72, 829–842. [Google Scholar] [PubMed]
- Park, J.C.; Yoo, H.; Kim, C.E.; Shim, S.Y.; Lee, M. Hispidulin-7-O-Neohesperidoside from Cirsium japonicum var. ussuriense Attenuates the Production of Inflammatory Mediators in LPS-Induced Raw 264.7 Cells and HT-29 Cells. Pharm. Mag. 2017, 13, 707–711. [Google Scholar]
- Bae, Y.; Lee, S.; Kim, S.H. Chrysin suppresses mast cell-mediated allergic inflammation: Involvement of calcium, caspase-1 and nuclear factor-κB. Toxicol. Appl. Pharm. 2011, 254, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Gao, M.Q.; Peng, J.J.; Han, M.; Liu, K.L.; Han, Y.T. Hispidulin mediates apoptosis in human renal cell carcinoma by inducing ceramide accumulation. Acta Pharm. Sin. 2017, 38, 1618–1631. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Gao, H.; Xie, J.; Yuan, Y.P.; Yuan, Q.; Gao, M.Q.; Liu, K.L.; Chen, X.H.; Han, Y.T.; Han, Z.W. Hispidulin induces ER stress-mediated apoptosis in human hepatocellular carcinoma cells in vitro and in vivo by activating AMPK signaling pathway. Acta Pharm. Sin. 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, W.; He, X.; Fei, Z. Hispidulin enhances the anti-tumor effects of temozolomide in glioblastoma by activating AMPK. Cell Biochem. Biophys. 2015, 71, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Liu, Y.; Li, K.; Wu, T.; Peng, J.; Jing, F. Hispidulin induces mitochondrial apoptosis in acute myeloid leukemia cells by targeting extracellular matrix metalloproteinase inducer. Am. J. Transl. Res. 2016, 8, 1115–1132. [Google Scholar]
- Xie, J.; Gao, H.; Peng, J.; Han, Y.; Chen, X.; Jiang, Q.; Wang, C. Hispidulin prevents hypoxia-induced epithelial-mesenchymal transition in human colon carcinoma cells. Am. J. Cancer Res. 2015, 5, 1047–1061. [Google Scholar]
- Gao, M.Q.; Gao, H.; Han, M.; Liu, K.L.; Peng, J.J.; Han, Y.T. Hispidulin suppresses tumor growth and metastasis in renal cell carcinoma by modulating ceramide-sphingosine 1-phosphate rheostat. Am. J. Cancer Res. 2017, 7, 1501–1514. [Google Scholar]
- Wu, X.; Xu, J. New Role of Hispidulin in Lipid Metabolism: PPARalpha Activator. Lipids 2016, 51, 1249–1257. [Google Scholar] [CrossRef]
- Lee, S.G.; Kim, J.S.; Min, K.; Kwon, T.K.; Nam, J.O. Hispidulin inhibits adipogenesis in 3T3-L1 adipocytes through PPARgamma pathway. Chem. Biol. Interact. 2018, 293, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-Y.; Lu, C.-W.; Wang, C.-C.; Lu, J.-F.; Wang, S.-J. Hispidulin inhibits the release of glutamate in rat cerebrocortical nerve terminals. Toxicol. Appl. Pharm. 2012, 263, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Akram, M.; Syed, A.S.; Kim, K.A.; Lee, J.S.; Chang, S.Y.; Kim, C.Y.; Bae, O.N. Heme oxygenase 1-mediated novel anti-inflammatory activities of Salvia plebeia and its active components. J. Ethnopharmacol. 2015, 174, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Oscherwitz, J.; Cease, K.B.; Chan, S.M.; Munoz-Planillo, R.; Hasegawa, M.; Villaruz, A.E.; Cheung, G.Y.; McGavin, M.J.; Travers, J.B.; et al. Staphylococcus delta-toxin induces allergic skin disease by activating mast cells. Nature 2013, 503, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Qu, C.; Srivastava, K.; Yang, N.; Busse, P.; Zhao, W.; Li, X.M. Food allergy herbal formula 2 protection against peanut anaphylactic reaction is via inhibition of mast cells and basophils. J. Allergy. Clin. Immunol. 2010, 126, 1208–1217. [Google Scholar] [CrossRef]
- Chung, M.J.; Kim, J.M.; Lee, S.; Kim, T.; Kim, D.; Baek, J.; Kim, T.; Lee, J.; Kim, K.; Yoon, J.A.; et al. Suppressive effects of Schizandra chinensis Baillon water extract on allergy-related cytokine generation and degranulation in IgE-antigen complex-stimulated RBL-2H3 cells. Nutr. Res. Pr. 2012, 6, 97–105. [Google Scholar] [CrossRef]
- Han, S.Y.; Bae, J.Y.; Park, S.H.; Kim, Y.H.; Park, J.H.; Kang, Y.H. Resveratrol inhibits IgE-mediated basophilic mast cell degranulation and passive cutaneous anaphylaxis in mice. J. Nutr. 2013, 143, 632–639. [Google Scholar] [CrossRef]
- Kang, B.C.; Kim, M.J.; Lee, S.; Choi, Y.A.; Park, P.H.; Shin, T.Y.; Kwon, T.K.; Khang, D.; Kim, S.H. Nothofagin suppresses mast cell-mediated allergic inflammation. Chem. Biol. Interact. 2019, 298, 1–7. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, Y.Y.; Choi, Y.A.; Baek, M.C.; Lee, B.; Park, P.H.; Shin, T.Y.; Kwon, T.K.; Khang, D.; Kim, S.H. Elaeocarpusin Inhibits Mast Cell-Mediated Allergic Inflammation. Front. Pharm. 2018, 9, 591. [Google Scholar] [CrossRef]
- Do, H.J.; Oh, T.W.; Yang, J.H.; Park, K.I.; Ma, J.Y. Davallia mariesii Moore Improves FcepsilonRI-Mediated Allergic Responses in the Rat Basophilic Leukemia Mast Cell Line RBL-2H3 and Passive Cutaneous Anaphylaxis in Mice. Mediat. Inflamm. 2017, 2017, 8701650. [Google Scholar] [CrossRef]
- Choi, J.K.; Jang, Y.H.; Lee, S.; Lee, S.R.; Choi, Y.A.; Jin, M.; Choi, J.H.; Park, J.H.; Park, P.H.; Choi, H.; et al. Chrysin attenuates atopic dermatitis by suppressing inflammation of keratinocytes. Food Chem. Toxicol. 2017, 110, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Bae, Y.; Kim, S.H. Galangin attenuates mast cell-mediated allergic inflammation. Food Chem. Toxicol. 2013, 57, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Gi Ho, L.; Sun Woo, J.; Se Jong, K.; Thi Hoa, P.; Jae Ho, C.; Hye Gwang, J. Tetrabromobisphenol A Induces MMP-9 Expression via NADPH Oxidase and the activation of ROS, MAPK, and Akt Pathways in Human Breast Cancer MCF-7 Cells. Toxicol. Res. 2019, 35, 93–101. [Google Scholar]
Sample Availability: Not available. |




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.E.; Min, K.-j.; Kim, M.-J.; Kim, S.-H.; Kwon, T.K. Hispidulin Inhibits Mast Cell-Mediated Allergic Inflammation through Down-Regulation of Histamine Release and Inflammatory Cytokines. Molecules 2019, 24, 2131. https://doi.org/10.3390/molecules24112131
Kim DE, Min K-j, Kim M-J, Kim S-H, Kwon TK. Hispidulin Inhibits Mast Cell-Mediated Allergic Inflammation through Down-Regulation of Histamine Release and Inflammatory Cytokines. Molecules. 2019; 24(11):2131. https://doi.org/10.3390/molecules24112131
Chicago/Turabian StyleKim, Dong Eun, Kyoung-jin Min, Min-Jong Kim, Sang-Hyun Kim, and Taeg Kyu Kwon. 2019. "Hispidulin Inhibits Mast Cell-Mediated Allergic Inflammation through Down-Regulation of Histamine Release and Inflammatory Cytokines" Molecules 24, no. 11: 2131. https://doi.org/10.3390/molecules24112131