Use of Physcion to Improve Atopic Dermatitis-Like Skin Lesions through Blocking of Thymic Stromal Lymphopoietin


Abstract
:1. Introduction
2. Results
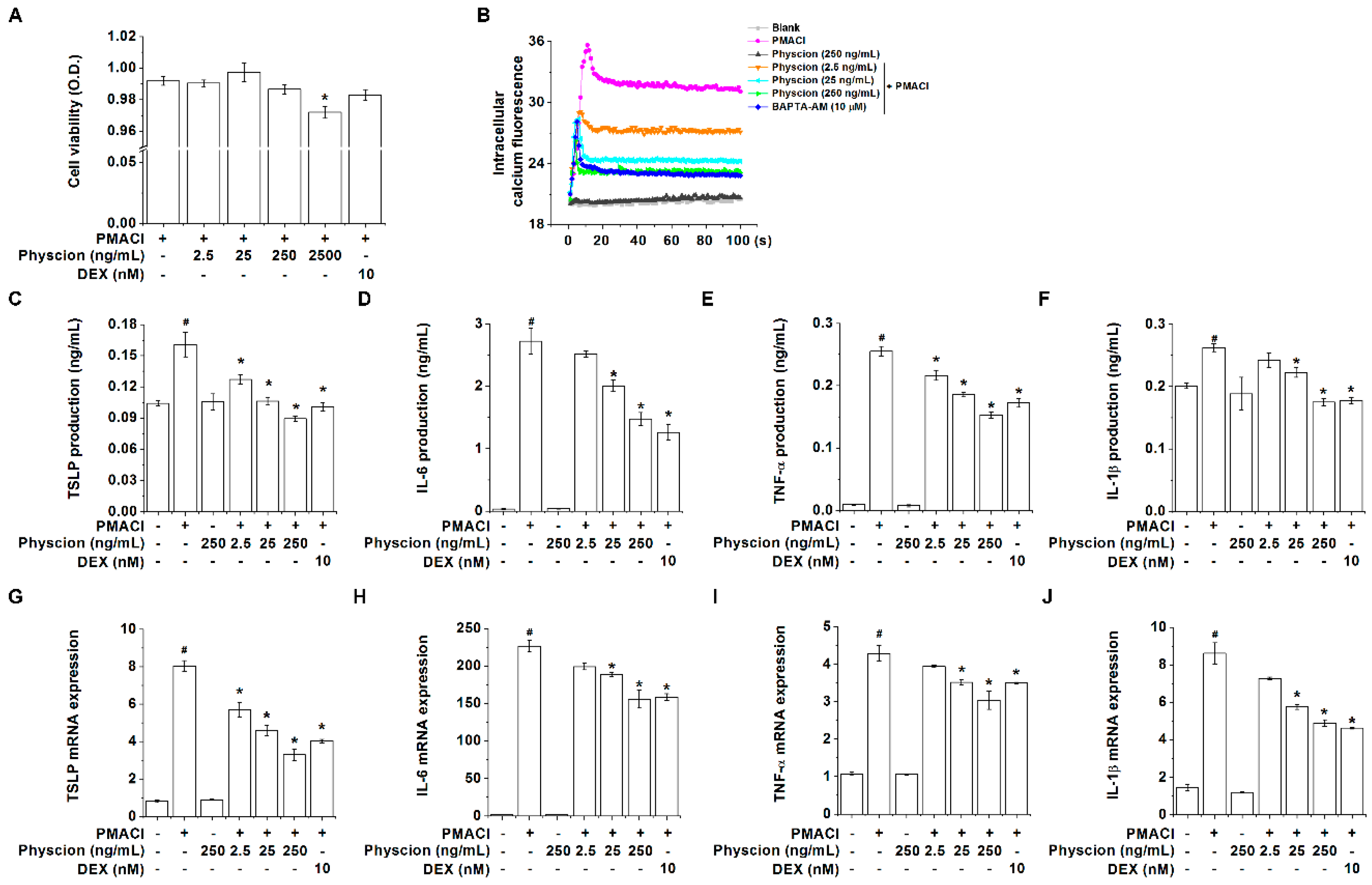
2.1. Physcion Attenuates TSLP Level in PMA Plus Calcium Ionophore (PMACI)-Stimulated HMC-1 Cells
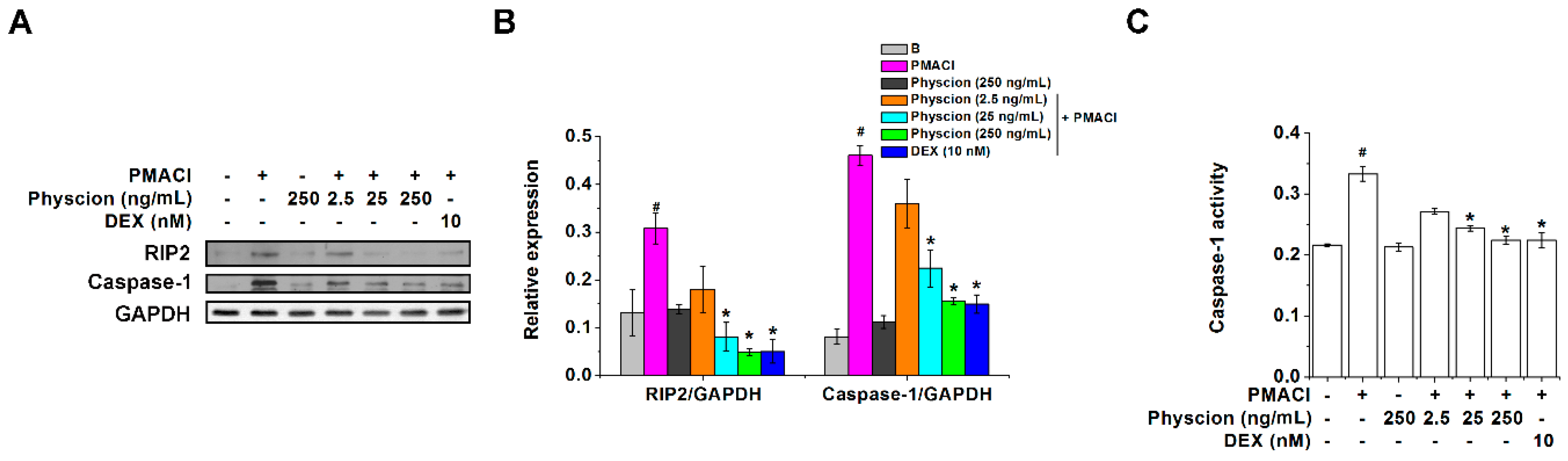
2.2. Physcion Downregulates RIP2 and Caspase-1 Expressions in PMACI-Stimulated HMC-1 Cells
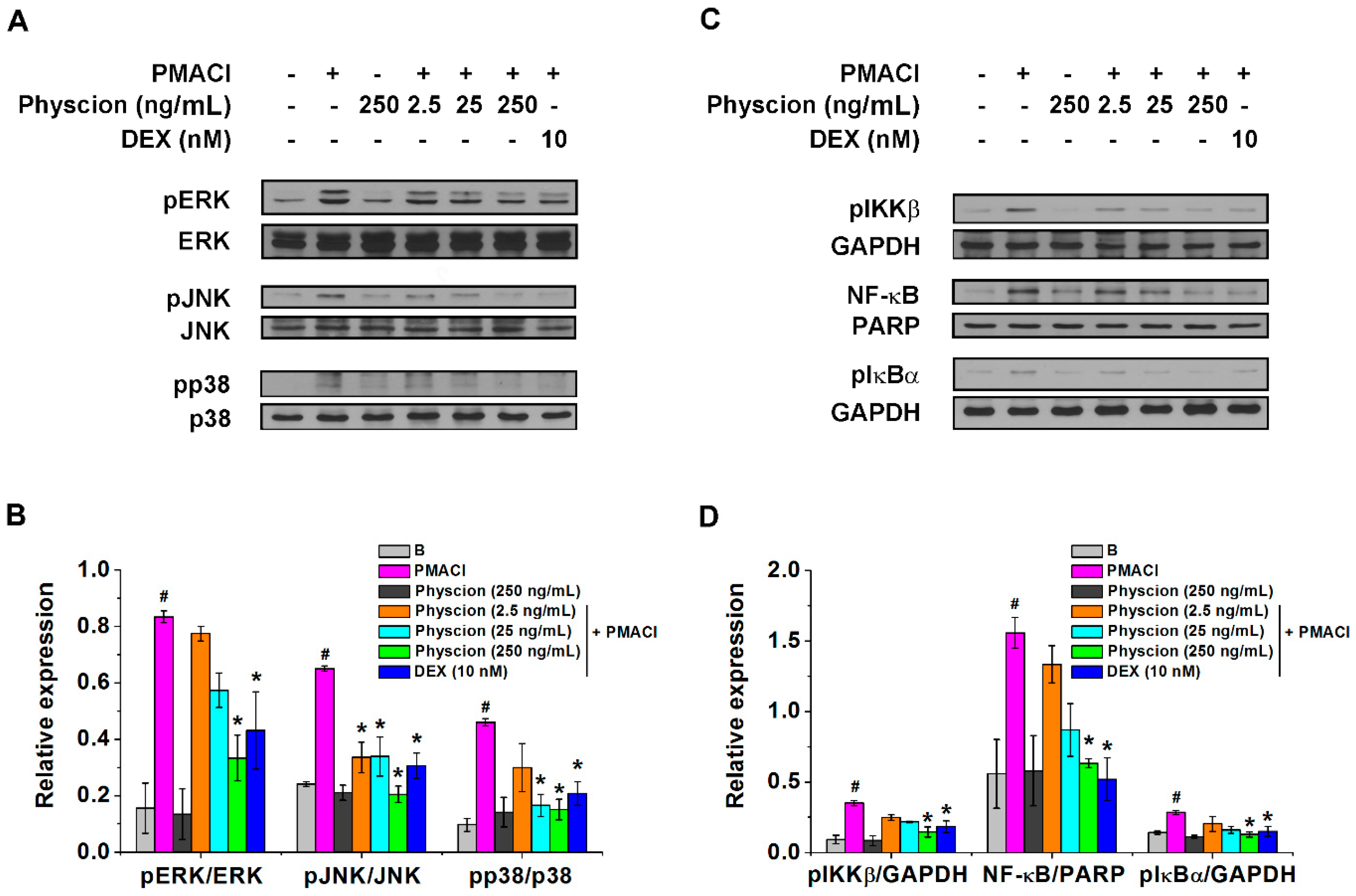
2.3. Physcion Attenuates Phospho (p)ERK/pJNK/pp38 And pIKKβ/NF-κB/pIkBα Expressions in PMACI-Stimulated HMC-1 Cells
2.4. Physcion Attenuates TSLP Production in Anti-CD3 And Anti-CD28 Antibodies-Stimulated Splenocytes
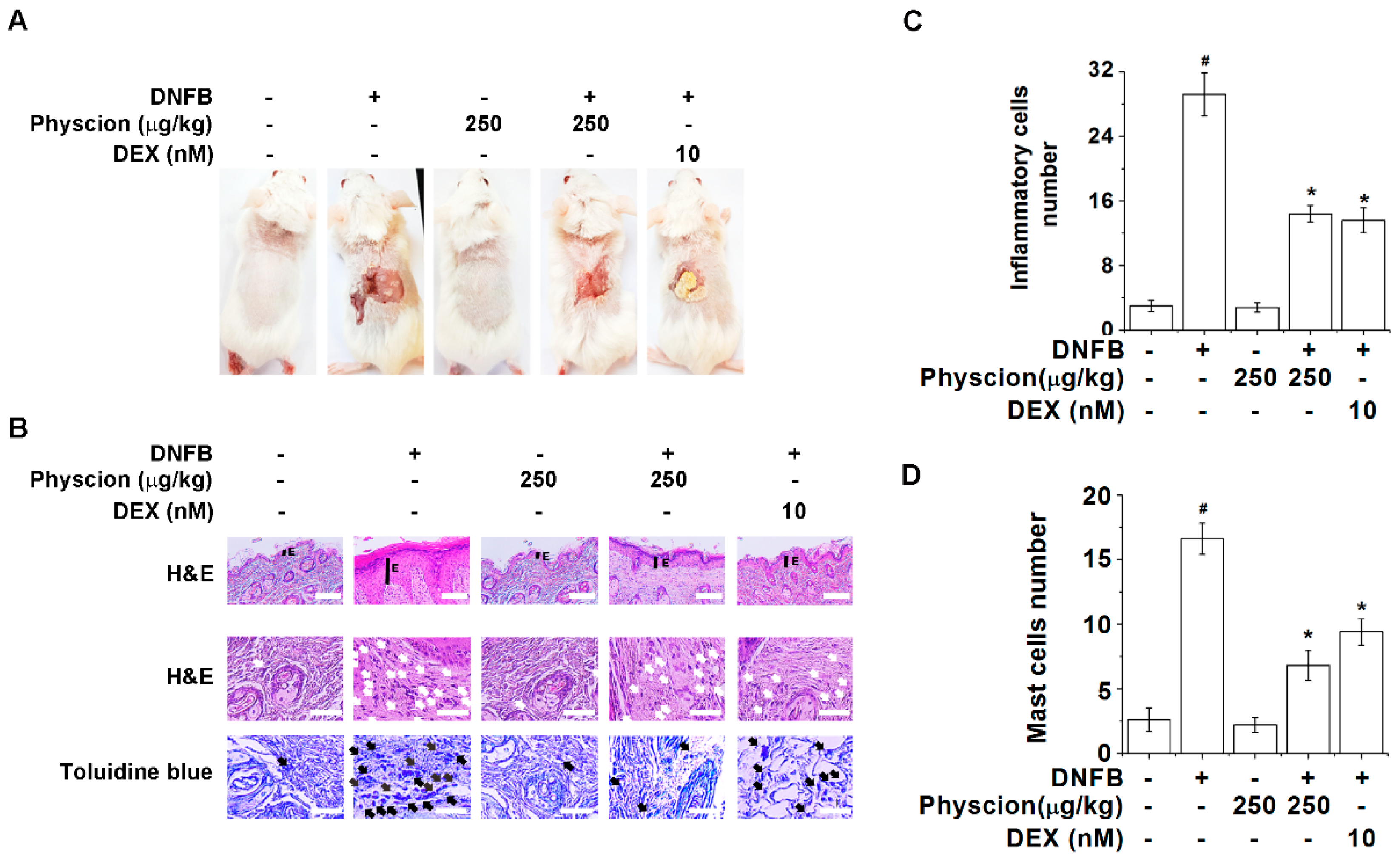
2.5. Physcion Relieves Pathological Changes of AD-Like Lesional Skin
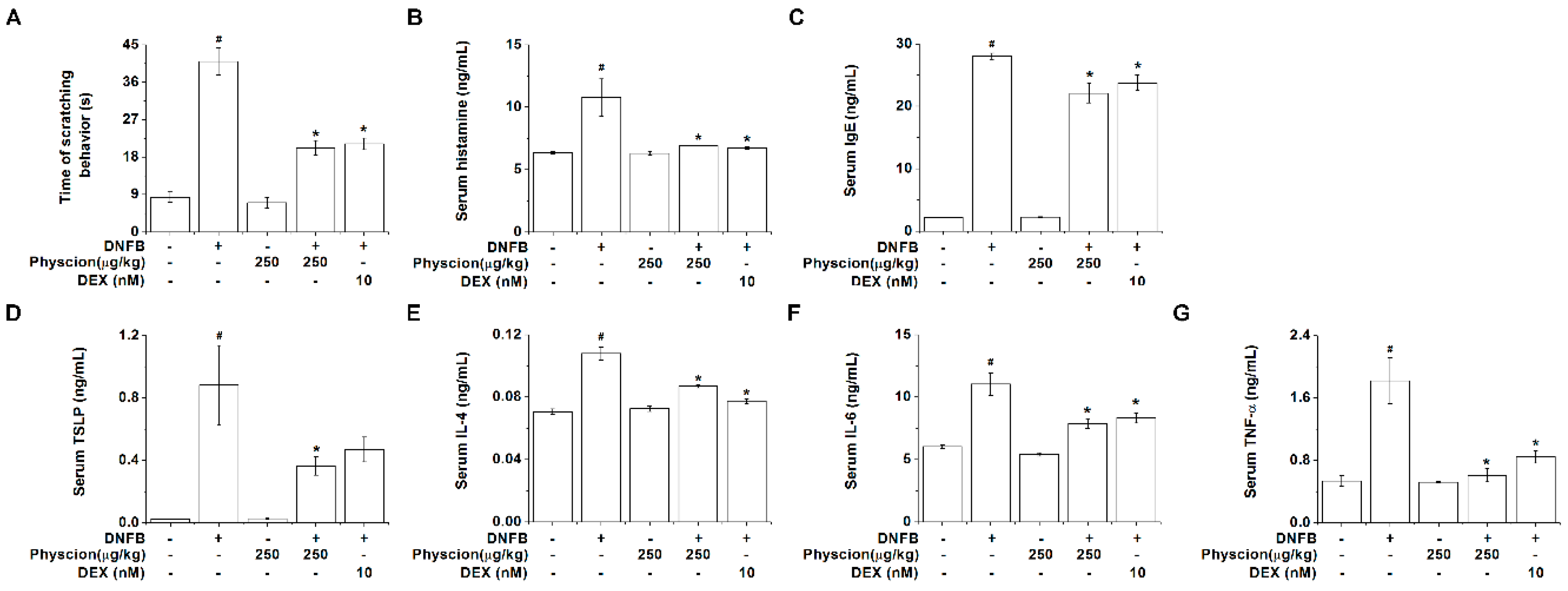
2.6. Physcion Inhibits Serum Histamine, IgE, And TSLP Levels in AD-Like Murine Model
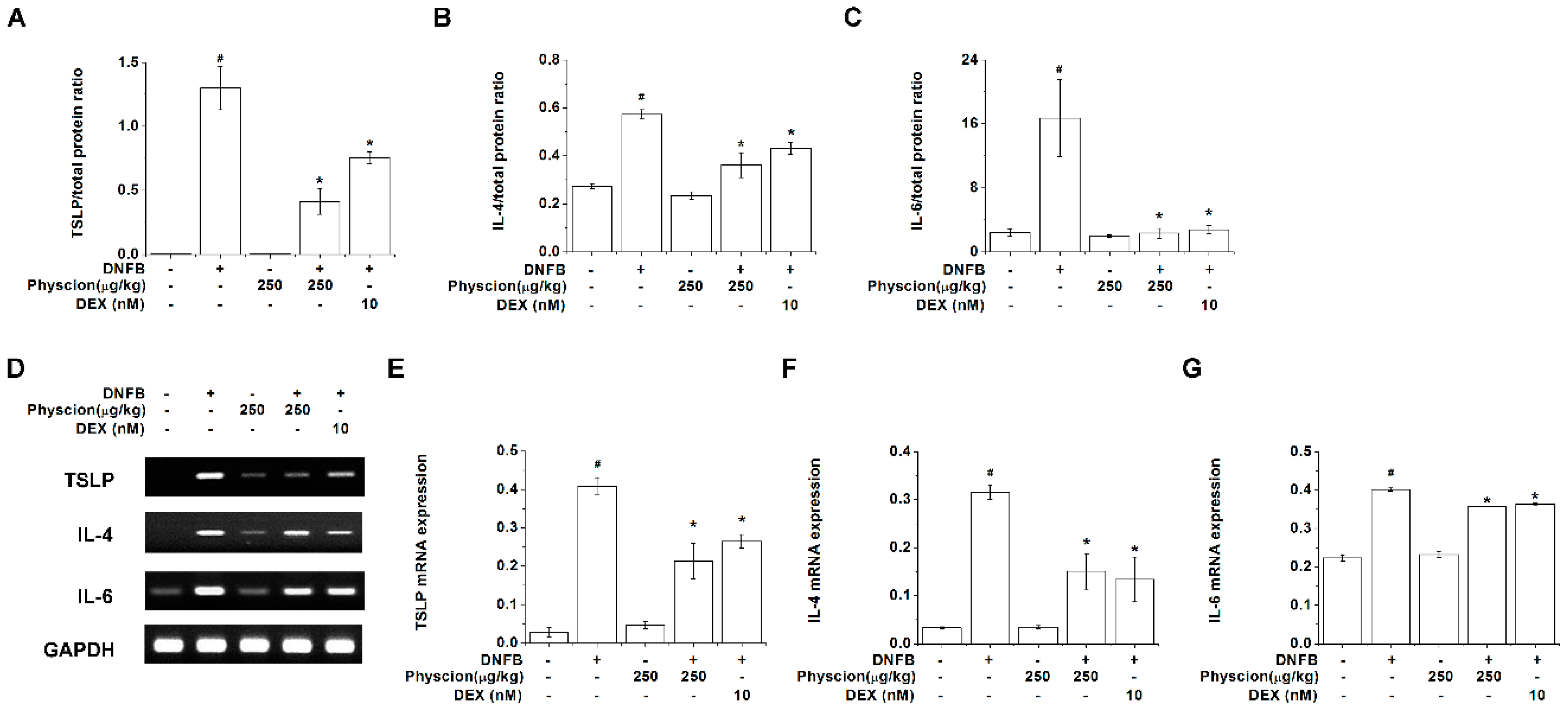
2.7. Physcion Reduces TSLP, IL-4, And IL-6 Expression Levels in Lesional Skin of DNFB-Induced AD-Like Murine Model
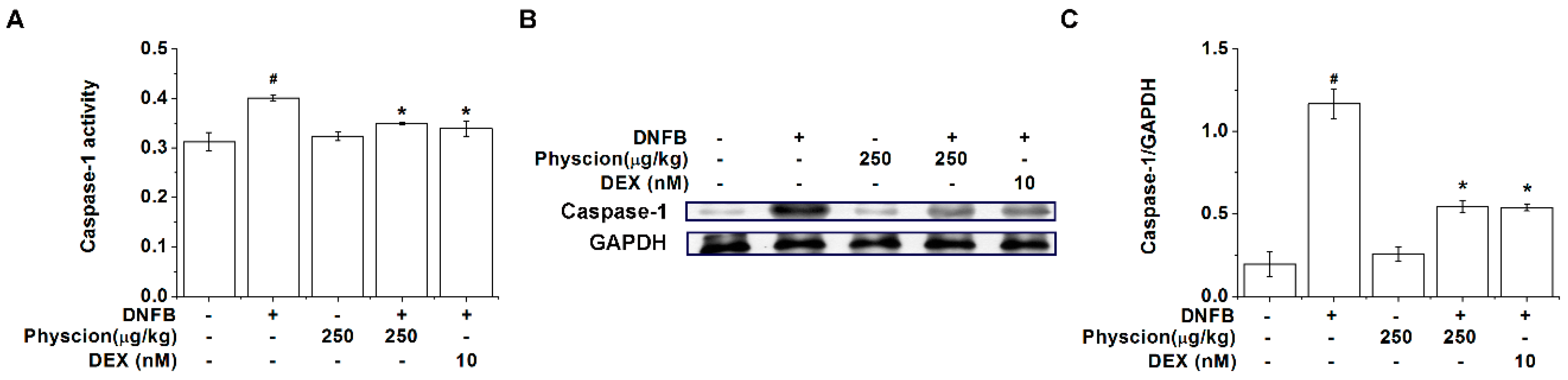
2.8. Physcion Suppresses Caspase-1 Activation in AD-Like Lesional Skin
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Physcion Preparation
4.3. Cell Culture
4.4. MTT Assay
4.5. Intracellular Calcium Levels
4.6. Cytokines Assay
4.7. Quantitative Real-Time Polymerase Chain Reaction (PCR) And Quantitative Reverse-Transcription PCR
4.8. Western Blotting
4.9. Caspase-1 Activity Assay
4.10. Nuclear Extracts And Cytoplasmic Extracts
4.11. Animals
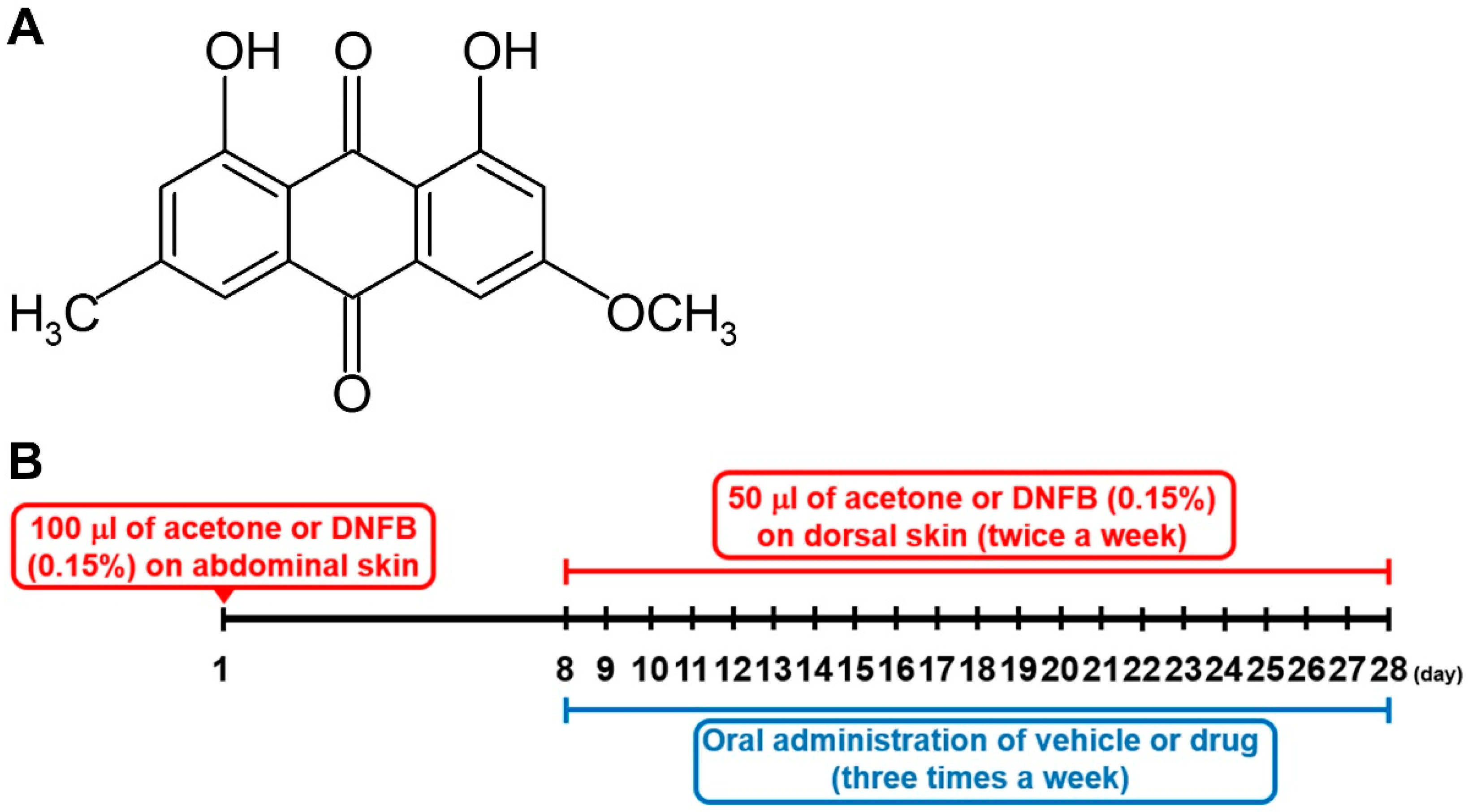
4.12. DNFB-Induced AD-Like Lesional Skin
4.13. Histological Analysis
4.14. Histamine Assay
4.15. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Camargo Lopes de Oliveira, L.; Pierotti, F.F.; Mallozi, M.; Cocco, R.R.; Rosário, N.; Rugue Genov, I.; Åberg, K.M.; Borres, M.P.; Solé, D. rBlo t 5 is a potential contributor to the severity of atopic dermatitis in a Brazilian population. Pediatr. Allergy Immunol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ingordo, V.; Cazzaniga, S.; Naldi, L.; Perrucci, S.; Barbierato, M.; Crociata, F.; Cusano, F. Atopic dermatitis in young adult italian males: Persistent and adult-onset varieties did not differ clinically and as for allergological variables. G. Ital. Dermatol. Venereol. 2019. [Google Scholar] [CrossRef]
- Kircik, L.H. Management of Atopic Dermatitis. J. Drugs Dermatol. 2019, 18, s111. [Google Scholar]
- Reed, B.; Blaiss, M.S. The burden of atopic dermatitis. Allergy Asthma Proc. 2018, 39, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Barr, T.P.; Garzia, C.; Guha, S.; Fletcher, E.K.; Nguyen, N.; Wieschhaus, A.J.; Ferrer, L.; Covic, L.; Kuliopulos, A. PAR2 Pepducin-Based Suppression of Inflammation and Itch in Atopic Dermatitis Models. J. Investig. Dermatol. 2019, 139, 412–421. [Google Scholar] [CrossRef]
- Han, N.R.; Moon, P.D.; Kim, H.M.; Jeong, H.J. Cordycepin ameliorates skin inflammation in a DNFB-challenged murine model of atopic dermatitis. Immunopharmacol. Immunotoxicol. 2018, 40, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Sasso, O.; Summa, M.; Armirotti, A.; Pontis, S.; De Mei, C.; Piomelli, D. The N-Acylethanolamine Acid Amidase Inhibitor ARN077 Suppresses Inflammation and Pruritus in a Mouse Model of Allergic Dermatitis. J. Investig. Dermatol. 2018, 138, 562–569. [Google Scholar] [CrossRef]
- Zhu, Y.; Pan, W.H.; Wang, X.R.; Liu, Y.; Chen, M.; Xu, X.G.; Liao, W.Q.; Hu, J.H. Tryptase and protease-activated receptor-2 stimulate scratching behavior in a murine model of ovalbumin-induced atopic-like dermatitis. Int. Immunopharmacol. 2015, 28, 507–512. [Google Scholar] [CrossRef]
- Schneider, C.; Döcke, W.D.; Zollner, T.M.; Röse, L. Chronic mouse model of TMA-induced contact hypersensitivity. J. Investig. Dermatol. 2009, 129, 899–907. [Google Scholar]
- Han, N.R.; Oh, H.A.; Nam, S.Y.; Moon, P.D.; Kim, D.W.; Kim, H.M.; Jeong, H.J. TSLP induces mast cell development and aggravates allergic reactions through the activation of MDM2 and STAT6. J. Investig. Dermatol. 2014, 134, 2521–2530. [Google Scholar] [CrossRef]
- Han, N.R.; Moon, P.D.; Yoou, M.S.; Chang, T.S.; Kim, H.M.; Jeong, H.J. Effect of massage therapy by VOSKIN 125+ painkiller® on inflammatory skin lesions. Dermatol. Ther. 2018, 31, e12628. [Google Scholar] [CrossRef]
- Schneider, K.S.; Groß, C.J.; Dreier, R.F.; Saller, B.S.; Mishra, R.; Gorka, O.; Heilig, R.; Meunier, E.; Dick, M.S.; Ćiković, T.; et al. The Inflammasome Drives GSDMD-Independent Secondary Pyroptosis and IL-1 Release in the Absence of Caspase-1 Protease Activity. Cell Rep. 2017, 21, 3846–3859. [Google Scholar] [CrossRef]
- Han, N.R.; Moon, P.D.; Kim, N.R.; Kim, H.Y.; Jeong, H.J.; Kim, H.M. Schisandra chinensis and Its Main Constituent Schizandrin Attenuate Allergic Reactions by Down-Regulating Caspase-1 in Ovalbumin-Sensitized Mice. Am. J. Chin. Med. 2017, 45, 159–172. [Google Scholar] [CrossRef]
- Błażejewski, A.J.; Thiemann, S.; Schenk, A.; Pils, M.C.; Gálvez, E.J.C.; Roy, U.; Heise, U.; de Zoete, M.R.; Flavell, R.A.; Strowig, T. Microbiota Normalization Reveals that Canonical Caspase-1 Activation Exacerbates Chemically Induced Intestinal Inflammation. Cell Rep. 2017, 19, 2319–2330. [Google Scholar] [CrossRef]
- Moon, P.D.; Kim, H.M. Thymic stromal lymphopoietin is expressed and produced by caspase-1/NF-κB pathway in mast cells. Cytokine 2011, 54, 239–243. [Google Scholar] [CrossRef]
- Mueller, S.O.; Schmitt, M.; Dekant, W.; Stopper, H.; Schlatter, J.; Schreier, P.; Lutz, W.K. Occurrence of emodin, chrysophanol and physcion in vegetables, herbs and liquors. Genotoxicity and anti-genotoxicity of the anthraquinones and of the whole plants. Food Chem. Toxicol. 1999, 37, 481–491. [Google Scholar] [CrossRef]
- Qin, X.; Peng, Y.; Zheng, J. In Vitro and in Vivo Studies of the Electrophilicity of Physcion and its Oxidative Metabolites. Chem. Res. Toxicol. 2018, 31, 340–349. [Google Scholar] [CrossRef]
- Pan, X.P.; Wang, C.; Li, Y.; Huang, L.H. Physcion induces apoptosis through triggering endoplasmic reticulum stress in hepatocellular carcinoma. Biomed. Pharmacother. 2018, 99, 894–903. [Google Scholar] [CrossRef]
- Pan, X.; Wang, H.; Tong, D.; Wang, C.; Sun, L.; Zhao, C.; Li, Y.; Zhu, L.; Wu, D. Physcion induces apoptosis in hepatocellular carcinoma by modulating miR-370. Am. J. Cancer Res. 2016, 6, 2919–2931. [Google Scholar]
- Chen, X.; Gao, H.; Han, Y.; Ye, J.; Xie, J.; Wang, C. Physcion induces mitochondria-driven apoptosis in colorectal cancer cells via downregulating EMMPRIN. Eur. J. Pharmacol. 2015, 764, 124–133. [Google Scholar] [CrossRef]
- Han, N.R.; Kim, H.M.; Jeong, H.J. Thymic stromal lymphopoietin is regulated by the intracellular calcium. Cytokine 2012, 59, 215–217. [Google Scholar] [CrossRef]
- Kim, M.H.; Seo, J.H.; Kim, H.M.; Jeong, H.J. Aluminum-doped zinc oxide nanoparticles attenuate the TSLP levels via suppressing caspase-1 in activated mast cells. J. Biomater. Appl. 2016, 30, 1407–1416. [Google Scholar] [CrossRef]
- Debnath, T.; Lee, Y.M.; Lim, J.H.; Lim, B.O. Anti-allergic and anti-atopic dermatitis effects of Gardenia Fructus extract. Food Agric. Immunol. 2018, 29, 665–674. [Google Scholar] [CrossRef]
- Gonzalez, J.; Orlofsky, A.; Prystowsky, M.B. A1 is a growth-permissive antiapoptotic factor mediating postactivation survival in T cells. Blood 2003, 101, 2679–2685. [Google Scholar] [CrossRef]
- Han, N.R.; Moon, P.D.; Kim, H.M.; Jeong, H.J. Effect of Pyeongwee-San (KMP6) on 2,4-dinitrofluorobenzene-induced atopic dermatitis-like skin lesions in NC/Nga mice. Life Sci. 2012, 90, 147–153. [Google Scholar] [CrossRef]
- Weber, M.B.; Petry, V.; Weis, L.; Mazzotti, N.G.; Cestari, T.F. Evaluating the relation between pruritus, serum IgE levels and severity of clinical manifestations in atopic dermatitis patients. An. Bras. Dermatol. 2005, 80, 245–248. [Google Scholar] [CrossRef]
- Wilson, S.R.; Thé, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef]
- Wong, L.S.; Wu, T.; Lee, C.H. Inflammatory and Noninflammatory Itch: Implications in Pathophysiology-Directed Treatments. Int. J. Mol. Sci. 2017, 18, 1485. [Google Scholar] [CrossRef]
- Moon, P.D.; Han, N.R.; Lee, J.S.; Kim, H.M.; Jeong, H.J. Effects of Linalyl Acetate on Thymic Stromal Lymphopoietin Production in Mast Cells. Molecules 2018, 23, 1711. [Google Scholar] [CrossRef]
- Ziegler, S.F. The role of thymic stromal lymphopoietin (TSLP) in allergic disorders. Curr. Opin. Immunol. 2010, 22, 795–799. [Google Scholar] [CrossRef]
- Jang, H.; Matsuda, A.; Jung, K.; Karasawa, K.; Matsuda, K.; Oida, K.; Ishizaka, S.; Ahn, G.; Amagai, Y.; Moon, C.; et al. Skin pH is the Master Switch of Kallikrein 5-Mediated Skin Barrier Destruction in a Murine Atopic Dermatitis Model. J. Investig. Dermatol. 2016, 136, 127–135. [Google Scholar] [CrossRef]
- Moon, P.D.; Han, N.R.; Kim, H.M.; Jeong, H.J. High-Fat Diet Exacerbates Dermatitis through Up-Regulation of TSLP. J. Investig. Dermatol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, L.; Cios, A.; Lewicki, S.; Szymanski, P.; Stankiewicz, W. Fas/FasL pathway and cytokines in keratinocytes in atopic dermatitis—Manipulation by the electromagnetic field. PLoS ONE 2018, 13, e0205103. [Google Scholar] [CrossRef] [PubMed]
- Szegedi, K.; Lutter, R.; Res, P.C.; Bos, J.D.; Luiten, R.M.; Kezic, S.; Middelkamp-Hup, M.A. Cytokine profiles in interstitial fluid from chronic atopic dermatitis skin. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 2136–2144. [Google Scholar] [CrossRef] [PubMed]
- Batista, D.I.; Perez, L.; Orfali, R.L.; Zaniboni, M.C.; Samorano, L.P.; Pereira, N.V.; Sotto, M.N.; Ishizaki, A.S.; Oliveira, L.M.; Sato, M.N.; et al. Profile of skin barrier proteins (filaggrin, claudins 1 and 4) and Th1/Th2/Th17 cytokines in adults with atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Humke, E.W.; Shriver, S.K.; Starovasnik, M.A.; Fairbrother, W.J.; Dixit, V.M. ICEBERG: A novel inhibitor of interleukin-1beta generation. Cell 2000, 103, 99–111. [Google Scholar] [CrossRef]
- Moon, P.D.; Han, N.R.; Ryu, K.J.; Kang, S.W.; Go, J.H.; Jang, J.B.; Choi, Y.; Kim, H.M.; Jeong, H.J. A novel compound 2-(4-{2-[(phenylthio)acetyl]carbonohydrazonoyl}phenoxy)acetamide downregulates TSLP through blocking of caspase-1/NF-κB pathways. Int. Immunopharmacol. 2016, 38, 420–425. [Google Scholar] [CrossRef]
- Han, N.R.; Moon, P.D.; Kim, H.M.; Jeong, H.J. Tryptanthrin ameliorates atopic dermatitis through down-regulation of TSLP. Arch. Biochem. Biophys. 2014, 542, 14–20. [Google Scholar] [CrossRef]
- Moon, P.D.; Choi, I.H.; Kim, H.M. Epigallocatechin-3-O-gallate inhibits the production of thymic stromal lymphopoietin by the blockade of caspase-1/NF-κB pathway in mast cells. Amino Acids 2012, 42, 2513–2519. [Google Scholar] [CrossRef] [PubMed]
- Kakeda, M.; Yamanaka, K.; Kitagawa, H.; Tsuda, K.; Akeda, T.; Kurokawa, I.; Gabazza, E.C.; Mizutani, H. Heat-killed bacillus Calmette-Guérin and Mycobacterium kansasii antigen 85B combined vaccination ameliorates dermatitis in a mouse model of atopic dermatitis by inducing regulatory T cells. Br. J. Dermatol. 2012, 166, 953–963. [Google Scholar] [CrossRef]
- Hiramoto, K.; Yamate, Y.; Yokoyama, S. Ultraviolet B eye irradiation aggravates atopic dermatitis via adrenocorticotropic hormone and NLRP3 inflammasome in NC/Nga mice. Photodermatol. Photoimmunol. Photomed. 2018, 34, 200–210. [Google Scholar] [CrossRef]
- Kobayashi, K.; Inohara, N.; Hernandez, L.D.; Galán, J.E.; Núñez, G.; Janeway, C.A.; Medzhitov, R.; Flavell, R.A. RICK/Rip2/CARDIAK mediates signalling for receptors of the innate and adaptive immune systems. Nature 2002, 416, 194–199. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, L.; Wang, J.J.; Luo, P.F.; Wang, X.T.; Xia, Z.F. The caspase-1 inhibitor AC-YVAD-CMK attenuates acute gastric injury in mice: Involvement of silencing NLRP3 inflammasome activities. Sci. Rep. 2016, 6, 24166. [Google Scholar] [CrossRef]
- Hedl, M.; Abraham, C. Distinct roles for Nod2 protein and autocrine interleukin-1beta in muramyl dipeptide-induced mitogen-activated protein kinase activation and cytokine secretion in human macrophages. J. Biol. Chem. 2011, 286, 26440–26449. [Google Scholar] [CrossRef]
- Lee, H.C.; Ziegler, S.F. Inducible expression of the proallergic cytokine thymic stromal lymphopoietin in airway epithelial cells is controlled by NFkappaB. Proc. Natl. Acad. Sci. USA 2007, 104, 914–919. [Google Scholar] [CrossRef]
- Rizzo, J.M.; Oyelakin, A.; Min, S.; Smalley, K.; Bard, J.; Luo, W.; Nyquist, J.; Guttman-Yassky, E.; Yoshida, T.; De Benedetto, A.; et al. ΔNp63 regulates IL-33 and IL-31 signaling in atopic dermatitis. Cell Death Differ. 2016, 23, 1073–1085. [Google Scholar] [CrossRef]
- Choi, E.J.; Iwasa, M.; Han, K.I.; Kim, W.J.; Tang, Y.; Hwang, Y.J.; Chae, J.R.; Han, W.C.; Shin, Y.S.; Kim, E.K. Heat-Killed Enterococcus faecalis EF-2001 Ameliorates Atopic Dermatitis in a Murine Model. Nutrients 2016, 8, 146. [Google Scholar] [CrossRef]
- Imaizumi, A.; Kawakami, T.; Murakami, F.; Soma, Y.; Mizoguchi, M. Effective treatment of pruritus in atopic dermatitis using H1 antihistamines (second-generation antihistamines): Changes in blood histamine and tryptase levels. J. Dermatol. Sci. 2003, 33, 23–29. [Google Scholar] [CrossRef]
- Tian, H.; Liu, Z.; Pu, Y.; Bao, Y. Immunomodulatory effects exerted by Poria Cocos polysaccharides via TLR4/TRAF6/NF-κB signaling in vitro and in vivo. Biomed. Pharmacother. 2019, 112, 108709. [Google Scholar] [CrossRef]
- Liu, G.; Park, Y.J.; Tsuruta, Y.; Lorne, E.; Abraham, E. p53 Attenuates lipopolysaccharide-induced NF-kappaB activation and acute lung injury. J. Immunol. 2009, 182, 5063–5071. [Google Scholar] [CrossRef]
- Lee, S.A.; Moon, S.M.; Han, S.H.; Hwang, E.J.; Hong, J.H.; Park, B.R.; Choi, M.S.; Ahn, H.; Kim, J.S.; Kim, H.J.; et al. In Vivo and In Vitro Anti-Inflammatory Effects of Aqueous Extract of Anthriscus sylvestris Leaves. J. Med. Food. 2018, 21, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.S.; Gu, P.S.; Kim, N.; Jang, Y.P.; Oh, M.S. Effects of Rhei Undulati Rhizoma on lipopolysaccharide-induced neuroinflammation in vitro and in vivo. Environ. Toxicol. 2018, 33, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.Y.; Liu, Y.J.; Don, M.J.; Liu, H.Y.; Chen, Z.W.; Mettling, C.; Corbeau, P.; Chiang, C.K.; Jang, Y.S.; Li, T.H.; et al. Combined phytochemistry and chemotaxis assays for identification and mechanistic analysis of anti-inflammatory phytochemicals in Fallopia japonica. PLoS ONE 2011, 6, e27480. [Google Scholar] [CrossRef]
- Chen, X.; Murakami, T.; Oppenheim, J.J.; Howard, O.M.Z. Differential response of murine CD4+CD25+ and CD4+CD25− T cells to dexamethasone-induced cell death. Eur. J. Immunol. 2004, 34, 859–869. [Google Scholar] [CrossRef]
- Moon, P.D.; Choi, I.S.; Go, J.H.; Lee, B.J.; Kang, S.W.; Yoon, S.; Han, S.J.; Nam, S.Y.; Oh, H.A.; Han, N.R.; et al. Inhibitory effects of BiRyuChe-bang on mast cell-mediated allergic reactions and inflammatory cytokines production. Am. J. Chin. Med. 2013, 41, 1267–1282. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blank | - | Physcion (250 ng/mL) | Physcion (2.5 ng/mL) | Physcion (25 ng/mL) | Physcion (250 ng/mL) | DEX (10 nM) | |
|---|---|---|---|---|---|---|---|
| - | CD3/CD28 | - | CD3/CD28 | CD3/CD28 | CD3/CD28 | CD3/CD28 | |
| TSLP (ng/mL) | 0.16 ± 0.02 | 0.80 ± 0.14 # | 0.18 ± 0.01 | 0.65 ± 0.11 | 0.51 ± 0.08 | 0.25 ± 0.01 * | 0.56 ± 0.11 |
| IL-4 (ng/mL) | 0.00 ± 0.00 | 0.60 ± 0.15 # | 0.00 ± 0.00 | 0.42 ± 0.02 | 0.31 ± 0.04 | 0.24 ± 0.01 * | 0.03 ± 0.00 * |
| IL-6 (ng/mL) | 0.02 ± 0.00 | 1.85 ± 0.05 # | 0.02 ± 0.00 | 1.55 ± 0.11 * | 1.14 ± 0.02 * | 1.03 ± 0.02 * | 0.22 ± 0.01 * |
| TNF-α (ng/mL) | 0.14 ± 0.02 | 0.56 ± 0.06 # | 0.11 ± 0.00 | 0.46 ± 0.05 | 0.30 ± 0.01 * | 0.22 ± 0.01 * | 0.29 ± 0.02 * |
| IFN-γ (ng/mL) | 0.00 ± 0.00 | 35.95 ± 6.55 # | 0.00 ± 0.00 | 16.78 ± 1.00 * | 10.99 ± 0.76 * | 8.51 ± 0.69 * | 4.85 ± 0.10 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, P.-D.; Han, N.-R.; Lee, J.S.; Hong, S.; Yoo, M.-S.; Kim, H.-J.; Kim, J.-H.; Kang, S.; Jee, H.-W.; Kim, H.-M.; et al. Use of Physcion to Improve Atopic Dermatitis-Like Skin Lesions through Blocking of Thymic Stromal Lymphopoietin. Molecules 2019, 24, 1484. https://doi.org/10.3390/molecules24081484
Moon P-D, Han N-R, Lee JS, Hong S, Yoo M-S, Kim H-J, Kim J-H, Kang S, Jee H-W, Kim H-M, et al. Use of Physcion to Improve Atopic Dermatitis-Like Skin Lesions through Blocking of Thymic Stromal Lymphopoietin. Molecules. 2019; 24(8):1484. https://doi.org/10.3390/molecules24081484
Chicago/Turabian StyleMoon, Phil-Dong, Na-Ra Han, Jin Soo Lee, Sungwei Hong, Min-Sun Yoo, Hyeong-Jin Kim, Ji-Hyeon Kim, Soonsik Kang, Hyun-Woo Jee, Hyung-Min Kim, and et al. 2019. "Use of Physcion to Improve Atopic Dermatitis-Like Skin Lesions through Blocking of Thymic Stromal Lymphopoietin" Molecules 24, no. 8: 1484. https://doi.org/10.3390/molecules24081484