Cardiomyocyte Progenitor Cells as a Functional Gene Delivery Vehicle for Long-Term Biological Pacing


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
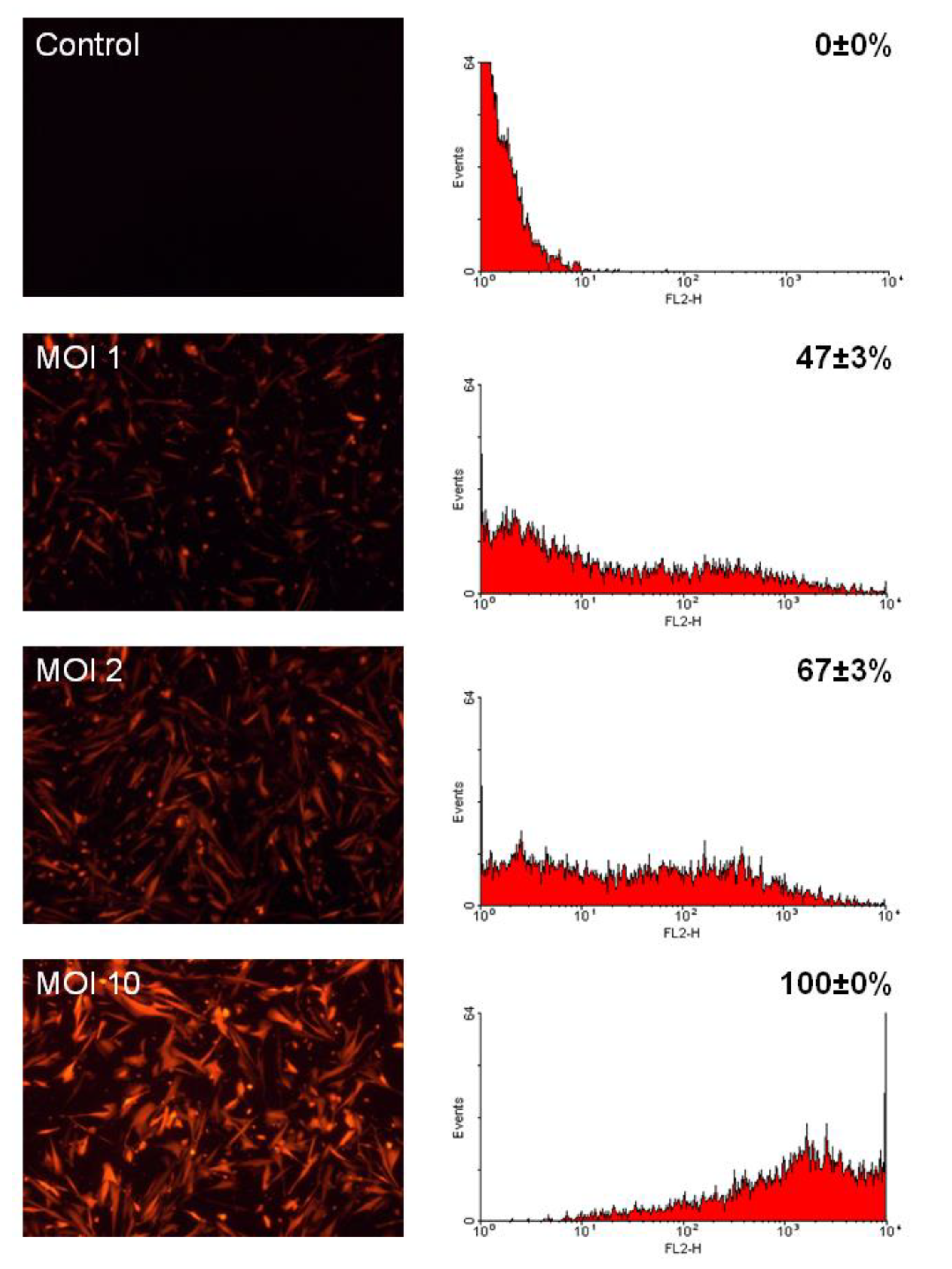
2.1. CMPCs are Efficiently Transduced by LV Vectors
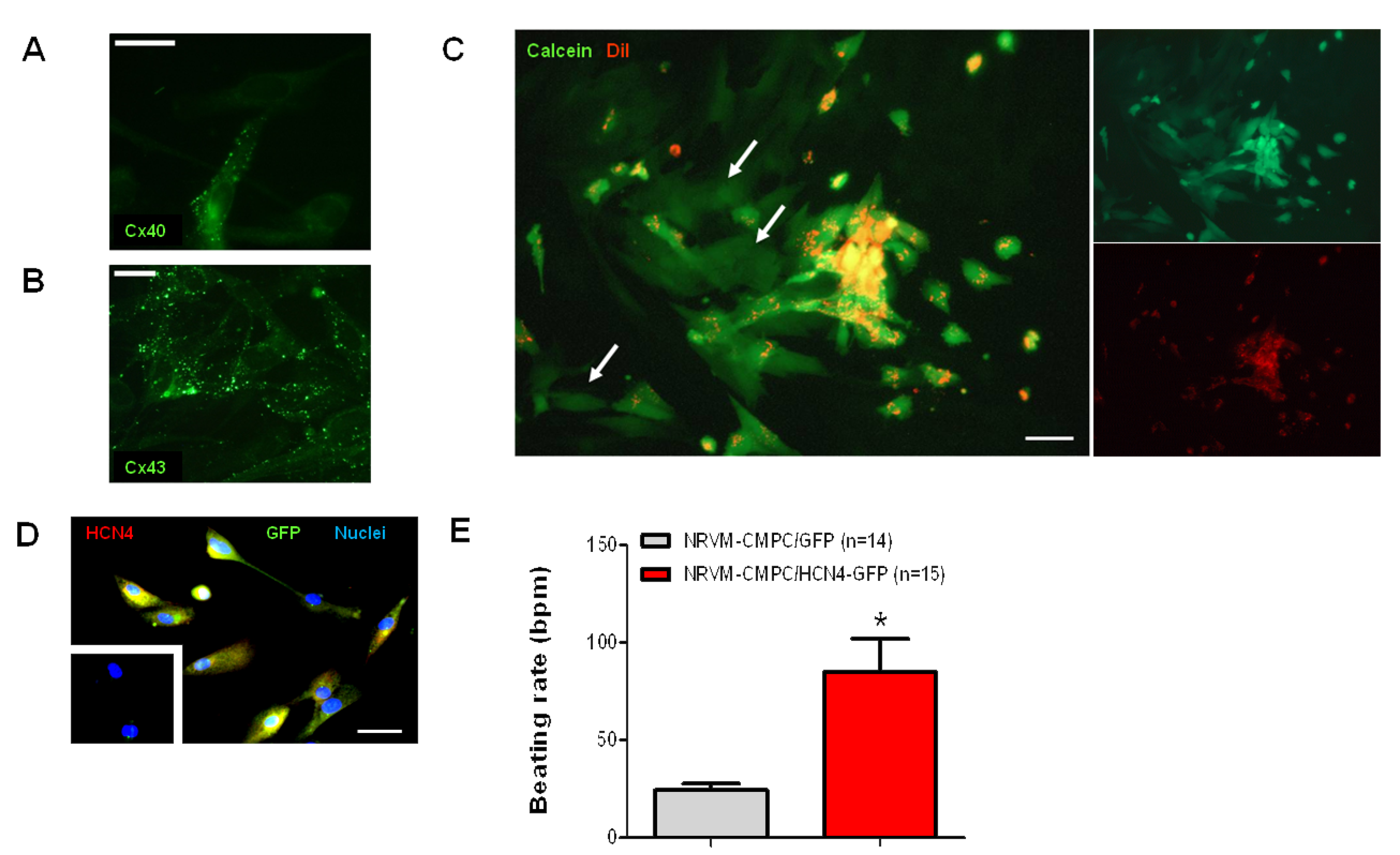
2.2. Undifferentiated CMPCs Couple to Cardiomyocytes
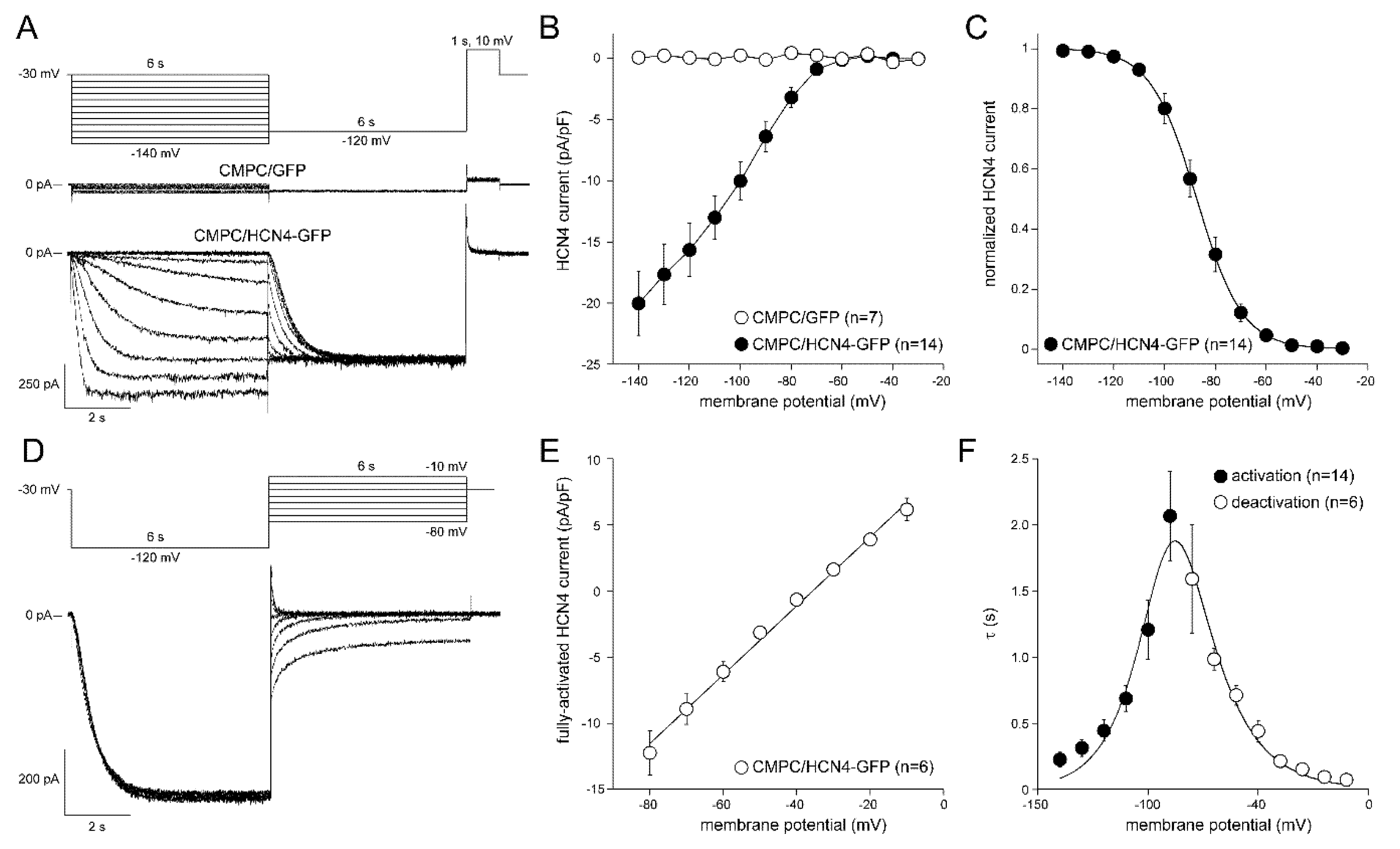
2.3. If Properties in LV-Transduced CMPCs
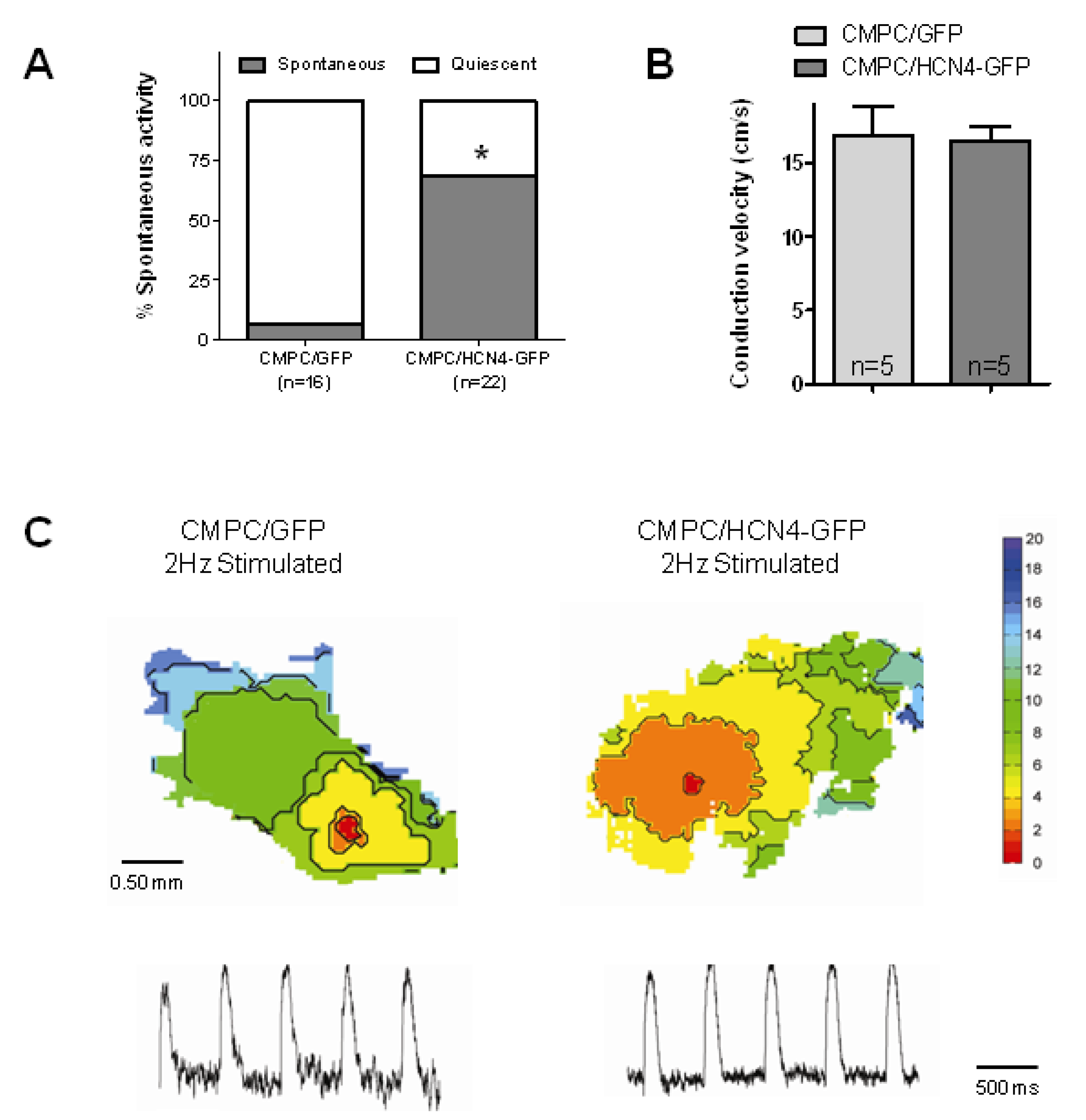
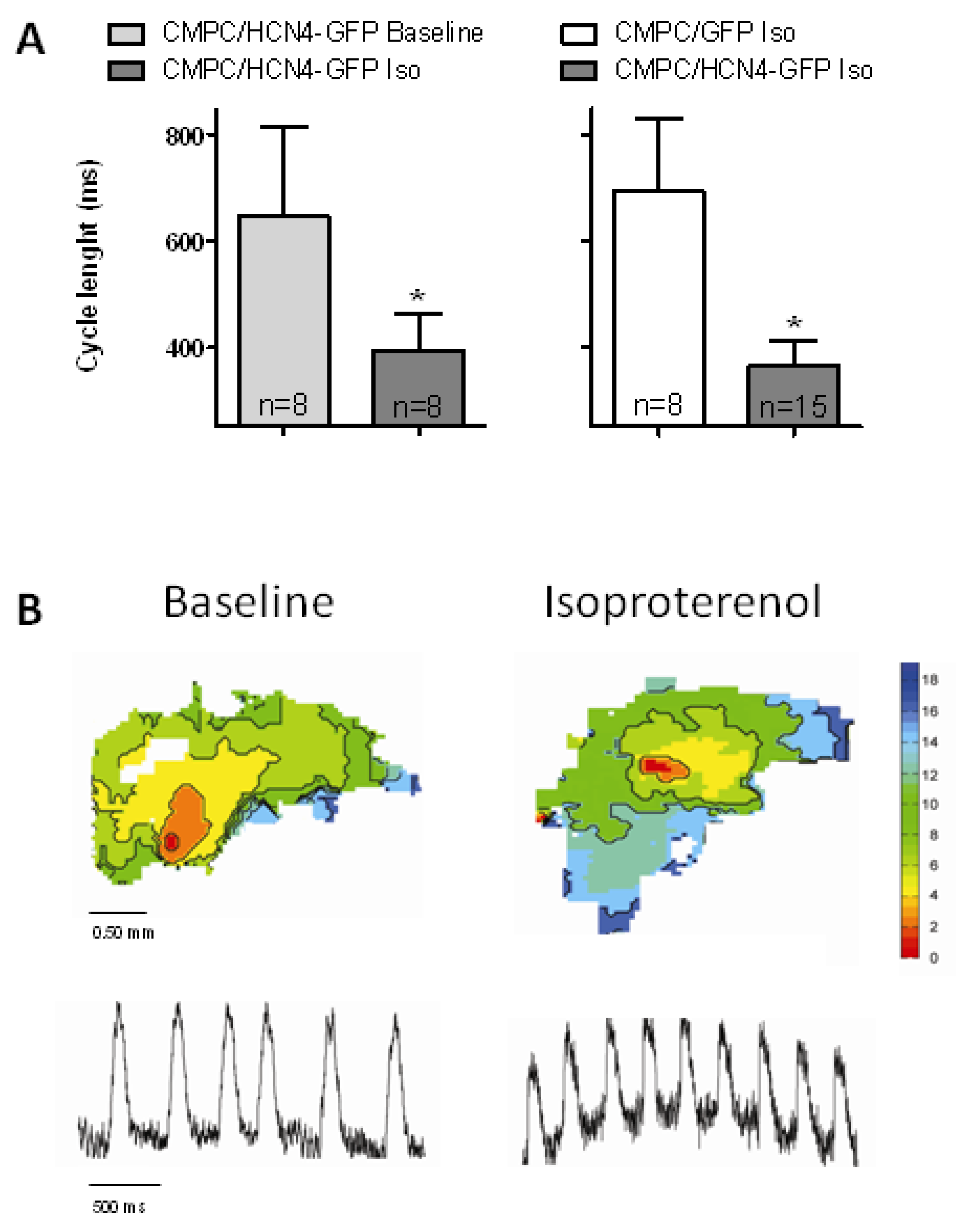
2.4. Biological Pacemaker Function in Organ Explant Cultures
3. Discussion
3.1. CMPC Transduction
3.2. Functional Interaction Between CMPCs and Cardiomyocytes
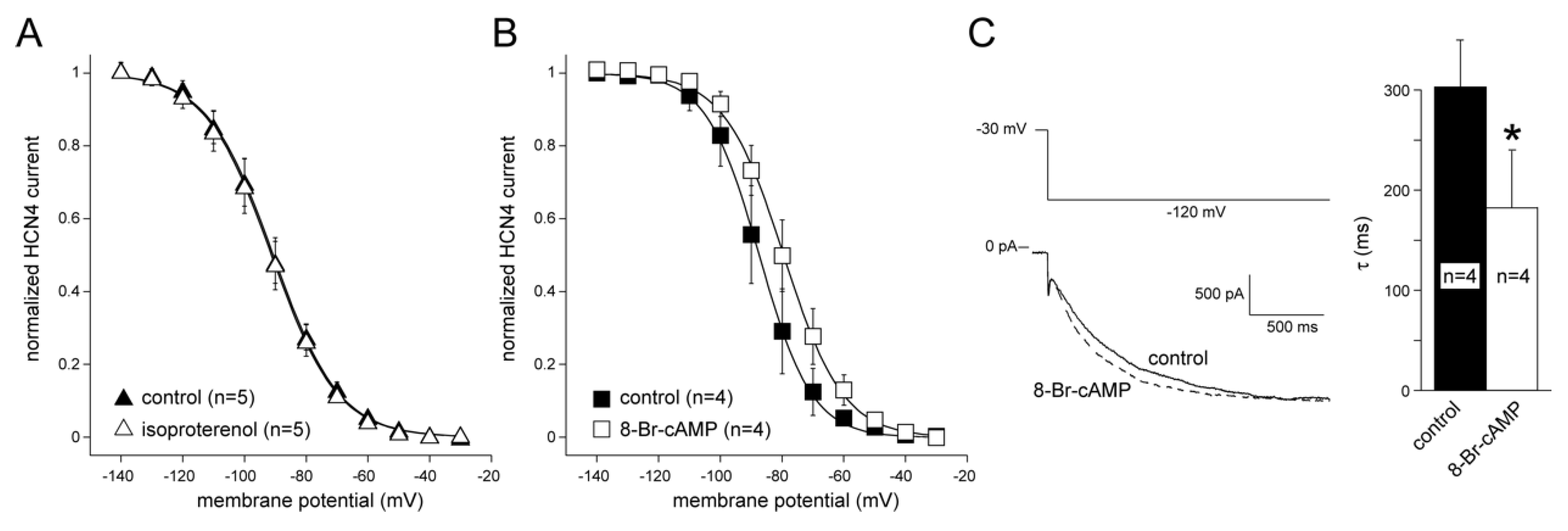
3.3. HCN4 Expression and Modulation
3.4. Biological Pacemaker Function Induced by HCN4-Expressing CMPCs
3.5. Safety Issues Related to CMPC-Based Biological Pacemakers
4. Materials and Methods
4.1. Cardiac Progenitor Cells Isolation and Culture
4.2. LV Vectors, Transduction, and Efficiency
4.3. Cell Isolation and Co-Culture of Neonatal Rat Ventricular Cardiac Myocytes
4.4. Immunofluoresent Labeling
4.5. CMPC-to-Cardiomyocyte Dye Transfer
4.6. Functional Interaction Between CMPCs and Cardiomyocytes
4.7. Single Cell Measurements
4.8. Organ Explant Cultures
4.9. Optical Mapping of Impulse Generation and Action Potential Propagation
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rosen, M.R.; Robinson, R.B.; Brink, P.R.; Cohen, I.S. The road to biological pacing. Nat. Rev. Cardiol. 2011, 8, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Boink, G.J.J.; Christoffels, V.M.; Robinson, R.B.; Tan, H.L. The past, present, and future of pacemaker therapies. Trends Cardiovasc. Med. 2015, 25, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zaiss, A.K.; Colarusso, P.; Patel, K.; Haljan, G.; Wickham, T.J.; Muruve, D.A. The role of capsid-endothelial interactions in the innate immune response to adenovirus vectors. Hum. Gene Ther. 2003, 14, 627–643. [Google Scholar] [CrossRef]
- Muruve, D.A. The innate immune response to adenovirus vectors. Hum. Gene Ther. 2004, 15, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, E.; Goldhaber, J.I.; Marbán, E. Next-generation pacemakers: From small devices to biological pacemakers. Nat. Rev. Cardiol. 2018, 15, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Jessup, M.; Greenberg, B.; Mancini, D.; Cappola, T.; Pauly, D.F.; Jaski, B.; Yaroshinsky, A.; Zsebo, K.M.; Dittrich, H.; Hajjar, R.J. Calcium Upregulation by Percutaneous Administration of Gene Therapy in Cardiac Disease (CUPID) Investigators Calcium Upregulation by Percutaneous Administration of Gene Therapy in Cardiac Disease (CUPID): A phase 2 trial of intracoronary gene therapy of sarcoplasmic reticulum Ca2+-ATPase in patients with advanced heart failure. Circulation 2011, 124, 304–313. [Google Scholar] [PubMed]
- Boink, G.J.J.; Duan, L.; Nearing, B.D.; Shlapakova, I.N.; Sosunov, E.A.; Anyukhovsky, E.P.; Bobkov, E.; Kryukova, Y.; Ozgen, N.; Danilo, P.; et al. HCN2/SkM1 gene transfer into canine left bundle branch induces stable, autonomically responsive biological pacing at physiological heart rates. J. Am. Coll. Cardiol. 2013, 61, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.C.; Kashiwakura, Y.; Marbán, E. Creation of a biological pacemaker by cell fusion. Circ. Res. 2007, 100, 1112–1115. [Google Scholar] [CrossRef]
- Pedrazzoli, F.; Chrysantzas, I.; Dezzani, L.; Rosti, V.; Vincitorio, M.; Sitar, G. Cell fusion in tumor progression: the isolation of cell fusion products by physical methods. Cancer Cell Int. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.J.; Fan, X.; Guo, X.X.; Medin, J.A. Promoter-specific lentivectors for long-term, cardiac-directed therapy of Fabry disease. J. Cardiol. 2011, 57, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwano, K.; Arai, M.; Koitabashi, N.; Watanabe, A.; Ikeda, Y.; Miyoshi, H.; Kurabayashi, M. Lentiviral vector-mediated SERCA2 gene transfer protects against heart failure and left ventricular remodeling after myocardial Infarction in rats. Mol. Ther. 2008, 16, 1026–1032. [Google Scholar] [CrossRef] [PubMed]
- Boink, G.J.J.; Rosen, M.R. Regenerative therapies in electrophysiology and pacing: Introducing the next steps. J. Interv. Card. Electrophysiol. 2011, 31, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Schlimgen, R.; Howard, J.; Wooley, D.; Thompson, M.; Baden, L.R.; Yang, O.O.; Christiani, D.C.; Mostoslavsky, G.; Diamond, D.V.; Duane, E.G.; et al. Risks Associated With Lentiviral Vector Exposures and Prevention Strategies. J. Occup. Environ. Med. 2016, 58, 1159–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmori, T.; Mizukami, H.; Katakai, Y.; Kawai, S.; Nakamura, H.; Inoue, M.; Shu, T.; Sugimoto, H.; Sakata, Y. Safety of intra-articular transplantation of lentivirally transduced mesenchymal stromal cells for haemophilic arthropathy in a non-human primate. Int. J. Hematol. 2018, 108, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Beegle, J.R.; Magner, N.L.; Kalomoiris, S.; Harding, A.; Zhou, P.; Nacey, C.; White, J.L.; Pepper, K.; Gruenloh, W.; Annett, G.; et al. Preclinical evaluation of mesenchymal stem cells overexpressing VEGF to treat critical limb ischemia. Mol. Ther. Methods Clin. Dev. 2016, 3. [Google Scholar] [CrossRef]
- McGarrity, G.J.; Hoyah, G.; Winemiller, A.; Andre, K.; Stein, D.; Blick, G.; Greenberg, R.N.; Kinder, C.; Zolopa, A.; Binder-Scholl, G.; et al. Patient monitoring and follow-up in lentiviral clinical trials. J. Gene Med. 2013, 15, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Marcucci, K.T.; Jadlowsky, J.K.; Hwang, W.T.; Suhoski-Davis, M.; Gonzalez, V.E.; Kulikovskaya, I.; Gupta, M.; Lacey, S.F.; Plesa, G.; Chew, A.; et al. Retroviral and Lentiviral Safety Analysis of Gene-Modified T Cell Products and Infused HIV and Oncology Patients. Mol. Ther. 2018, 26, 269–279. [Google Scholar] [CrossRef]
- Brendel, C.; Rothe, M.; Santilli, G.; Charrier, S.; Stein, S.; Kunkel, H.; Abriss, D.; Müller-Kuller, U.; Gaspar, B.; Modlich, U.; et al. Non-Clinical Efficacy and Safety Studies on G1XCGD, a Lentiviral Vector for Ex Vivo Gene Therapy of X-Linked Chronic Granulomatous Disease. Hum. Gene Ther. Clin. Dev. 2018, 29, 69–79. [Google Scholar] [CrossRef]
- Lidonnici, M.R.; Paleari, Y.; Tiboni, F.; Mandelli, G.; Rossi, C.; Vezzoli, M.; Aprile, A.; Lederer, C.W.; Ambrosi, A.; Chanut, F.; et al. Multiple Integrated Non-clinical Studies Predict the Safety of Lentivirus-Mediated Gene Therapy for β-Thalassemia. Mol. Ther. Methods Clin. Dev. 2018, 11, 9–28. [Google Scholar] [CrossRef]
- Naldini, L. Ex vivo gene transfer and correction for cell-based therapies. Nat. Rev. Genet. 2011, 12, 301–315. [Google Scholar] [CrossRef]
- Valiunas, V.; Doronin, S.; Valiuniene, L.; Potapova, I.; Zuckerman, J.; Walcott, B.; Robinson, R.B.; Rosen, M.R.; Brink, P.R.; Cohen, I.S. Human mesenchymal stem cells make cardiac connexins and form functional gap junctions. J. Physiol. 2004, 555, 617–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potapova, I.; Plotnikov, A.; Lu, Z.; Danilo, P.; Valiunas, V.; Qu, J.; Doronin, S.; Zuckerman, J.; Shlapakova, I.N.; Gao, J.; et al. Human mesenchymal stem cells as a gene delivery system to create cardiac pacemakers. Circ. Res. 2004, 94, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, A.N.; Shlapakova, I.; Szabolcs, M.J.; Danilo, P.; Lorell, B.H.; Potapova, I.A.; Lu, Z.; Rosen, A.B.; Mathias, R.T.; Brink, P.R.; et al. Xenografted adult human mesenchymal stem cells provide a platform for sustained biological pacemaker function in canine heart. Circulation 2007, 116, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Smits, A.M.; van Laake, L.W.; den Ouden, K.; Schreurs, C.; Szuhai, K.; van Echteld, C.J.; Mummery, C.L.; Doevendans, P.A.; Goumans, M.-J. Human cardiomyocyte progenitor cell transplantation preserves long-term function of the infarcted mouse myocardium. Cardiovasc. Res. 2009, 83, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Qu, J.; Kryukova, Y.; Potapova, I.A.; Doronin, S.V.; Larsen, M.; Krishnamurthy, G.; Cohen, I.S.; Robinson, R.B. MiRP1 modulates HCN2 channel expression and gating in cardiac myocytes. J. Biol. Chem. 2004, 279, 43497–43502. [Google Scholar] [CrossRef] [PubMed]
- Den Haan, A.D.; Veldkamp, M.W.; Bakker, D.; Boink, G.J.J.; Janssen, R.B.; de Bakker, J.M.T.; Tan, H.L. Organ explant culture of neonatal rat ventricles: A new model to study gene and cell therapy. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Valiunas, V.; Kanaporis, G.; Valiuniene, L.; Gordon, C.; Wang, H.Z.; Li, L.; Robinson, R.B.; Rosen, M.R.; Cohen, I.S.; Brink, P.R. Coupling an HCN2-expressing cell to a myocyte creates a two-cell pacing unit. J. Physiol. 2009, 587, 5211–5226. [Google Scholar] [CrossRef] [Green Version]
- Van Vliet, P.; Roccio, M.; Smits, A.M.; Van Oorschot, A.A.M.; Metz, C.H.G.; Van Veen, T.A.B.; Sluijter, J.P.G.; Doevendans, P.A.; Goumans, M.J. Progenitor cells isolated from the human heart: A potential cell source for regenerative therapy. Neth. Heart J. 2008, 16, 163–169. [Google Scholar] [CrossRef]
- Goumans, M.-J.; de Boer, T.P.; Smits, A.M.; van Laake, L.W.; van Vliet, P.; Metz, C.H.G.G.; Korfage, T.H.; Kats, K.P.; Hochstenbach, R.; Pasterkamp, G.; et al. TGF-beta1 induces efficient differentiation of human cardiomyocyte progenitor cells into functional cardiomyocytes in vitro. Stem Cell Res. 2008, 1, 138–149. [Google Scholar] [CrossRef]
- De Boer, T.P.; van Veen, T.A.B.; Jonsson, M.K.B.; Kok, B.G.J.M.; Metz, C.H.G.; Sluijter, J.P.G.; Doevendans, P.A.; de Bakker, J.M.T.; Goumans, M.-J.; van der Heyden, M.A.G. Human cardiomyocyte progenitor cell-derived cardiomyocytes display a maturated electrical phenotype. J. Mol. Cell. Cardiol. 2010, 48, 254–260. [Google Scholar] [CrossRef]
- Shadrin, I.Y.; Yoon, W.; Li, L.; Shepherd, N.; Bursac, N. Rapid fusion between mesenchymal stem cells and cardiomyocytes yields electrically active, non-contractile hybrid cells. Sci. Rep. 2015, 5, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Boink, G.J.J.; Verkerk, A.O.; van Amersfoorth, S.C.M.; Tasseron, S.J.; van der Rijt, R.; Bakker, D.; Linnenbank, A.C.; van der Meulen, J.; de Bakker, J.M.T.; Seppen, J.; et al. Egineering physiologically controlled pacemaker cells with lentiviral HCN4 gene transfer. J. Gene Med. 2008, 10, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Barbuti, A.; Protas, L.; Santoro, B.; Cohen, I.S.; Robinson, R.B. HCN2 overexpression in newborn and adult ventricular myocytes: distinct effects on gating and excitability. Circ. Res. 2001, 89, E8–E14. [Google Scholar] [CrossRef] [PubMed]
- Bucchi, A.; Plotnikov, A.N.; Shlapakova, I.; Danilo, P.; Kryukova, Y.; Qu, J.; Lu, Z.; Liu, H.; Pan, Z.; Potapova, I.; et al. Wild-type and mutant HCN channels in a tandem biological-electronic cardiac pacemaker. Circulation 2006, 114, 992–999. [Google Scholar] [CrossRef]
- Barman, P.; Choisy, S.C.M.; Hancox, J.C.; James, A.F. β-Adrenoceptor/PKA-stimulation, Na(+)-Ca(2+) exchange and PKA-activated Cl(−) currents in rabbit cardiomyocytes: A conundrum. Cell Calcium 2011, 49, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Harmati, G.; Bányász, T.; Bárándi, L.; Szentandrássy, N.; Horváth, B.; Szabõ, G.; Szentmiklõsi, J.A.; Szénási, G.; Nánási, P.P.; Magyar, J. Effects of β-adrenoceptor stimulation on delayed rectifier K+ currents in canine ventricular cardiomyocytes. Br. J. Pharmacol. 2011, 162, 890–896. [Google Scholar] [CrossRef]
- Ogrodnik, J.; Niggli, E. Increased Ca2+ leak and spatiotemporal coherence of Ca2+ release in cardiomyocytes during β-adrenergic stimulation. J. Physiol. 2010, 588, 225–242. [Google Scholar] [CrossRef]
- Kanaporis, G.; Mese, G.; Valiuniene, L.; White, T.W.; Brink, P.R.; Valiunas, V. Gap Junction Channels Exhibit Connexin-specific Permeability to Cyclic Nucleotides. J. Gen. Physiol. 2008, 131, 293–305. [Google Scholar] [CrossRef] [Green Version]
- Jun, C.; Zhihui, Z.; Lu, W.; Yaoming, N.; Lei, W.; Yao, Q.; Zhiyuan, S. Canine bone marrow mesenchymal stromal cells with lentiviral mHCN4 gene transfer create cardiac pacemakers. Cytotherapy 2012, 14, 529–539. [Google Scholar] [CrossRef]
- Lu, W.; Yaoming, N.; Boli, R.; Jun, C.; Changhai, Z.; Yang, Z.; Zhiyuan, S. mHCN4 genetically modified canine mesenchymal stem cells provide biological pacemaking function in complete dogs with atrioventricular block. Pacing Clin. Electrophysiol. 2013, 36, 1138–1149. [Google Scholar] [CrossRef]
- Boink, G.J.J.; Lu, J.; Driessen, H.E.; Duan, L.; Sosunov, E.A.; Anyukhovsky, E.P.; Shlapakova, I.N.; Lau, D.H.; Rosen, T.S.; Danilo, P.; et al. Effect of skeletal muscle Na(+) channel delivered via a cell platform on cardiac conduction and arrhythmia induction. Circ. Arrhythm. Electrophysiol. 2012, 5, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wang, H.Z.; Jia, Z.; Zuckerman, J.; Lu, Z.; Guo, Y.; Boink, G.J.J.; Brink, P.R.; Robinson, R.B.; Entcheva, E.; et al. Improving cardiac conduction with a skeletal muscle sodium channel by gene and cell therapy. J. Cardiovasc. Pharmacol. 2012, 60, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Boink, G.J.J.; Nearing, B.D.; Shlapakova, I.N.; Duan, L.; Kryukova, Y.; Bobkov, Y.; Tan, H.L.; Cohen, I.S.; Danilo, P.; Robinson, R.B.; et al. Ca(2+)-stimulated adenylyl cyclase AC1 generates efficient biological pacing as single gene therapy and in combination with HCN2. Circulation 2012, 126, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, A.N.; Bucchi, A.; Shlapakova, I.; Danilo, P.; Brink, P.R.; Robinson, R.B.; Cohen, I.S.; Rosen, M.R. HCN212-channel biological pacemakers manifesting ventricular tachyarrhythmias are responsive to treatment with I(f) blockade. Heart Rhythm 2008, 5, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Montini, E.; Cesana, D.; Schmidt, M.; Sanvito, F.; Ponzoni, M.; Bartholomae, C.; Sergi, L.S.; Benedicenti, F.; Ambrosi, A.; Di Serio, C.; et al. Hematopoietic stem cell gene transfer in a tumor-prone mouse model uncovers low genotoxicity of lentiviral vector integration. Nat. Biotechnol. 2006, 24, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Montini, E.; Cesana, D.; Schmidt, M.; Sanvito, F.; Bartholomae, C.C.; Ranzani, M.; Benedicenti, F.; Sergi, L.S.; Ambrosi, A.; Ponzoni, M.; et al. The genotoxic potential of retroviral vectors is strongly modulated by vector design and integration site selection in a mouse model of HSC gene therapy. J. Clin. Investig. 2009, 119, 964–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zychlinski, D.; Schambach, A.; Modlich, U.; Maetzig, T.; Meyer, J.; Grassman, E.; Mishra, A.; Baum, C. Physiological promoters reduce the genotoxic risk of integrating gene vectors. Mol. Ther. 2008, 16, 718–725. [Google Scholar] [CrossRef]
- Smits, A.M.; van Vliet, P.; Metz, C.H.; Korfage, T.; Sluijter, J.P.; Doevendans, P.A.; Goumans, M.J. Human cardiomyocyte progenitor cells differentiate into functional mature cardiomyocytes: An in vitro model for studying human cardiac physiology and pathophysiology. Nat. Protoc. 2009, 4, 232–243. [Google Scholar] [CrossRef]
- Bell, R.M.; Mocanu, M.M.; Yellon, D.M. Retrograde heart perfusion: The Langendorff technique of isolated heart perfusion. J. Mol. Cell. Cardiol. 2011, 50, 940–950. [Google Scholar] [CrossRef]
- Seppen, J.; van der Rijt, R.; Looije, N.; van Til, N.P.; Lamers, W.H.; Oude Elferink, R.P.J. Long-term correction of bilirubin UDPglucuronyltransferase deficiency in rats by in utero lentiviral gene transfer. Mol. Ther. 2003, 8, 593–599. [Google Scholar] [CrossRef]
- Rohr, S.; Schölly, D.M.; Kléber, A.G. Patterned growth of neonatal rat heart cells in culture. Morphological and electrophysiological characterization. Circ. Res. 1991, 68, 114–130. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, A.O.; Tan, H.L.; Ravesloot, J.H. Ca2+-activated Cl− current reduces transmural electrical heterogeneity within the rabbit left ventricle. Acta Physiol. Scand. 2004, 180, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Potse, M.; Linnenbank, A.C.; Grimbergen, C.A. Software design for analysis of multichannel intracardial and body surface electrocardiograms. Comput. Methods Programs Biomed. 2002, 69, 225–236. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds LV-GFP and LV-HCN4-GFP are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Végh, A.M.D.; Den Haan, A.D.; Cócera Ortega, L.; Verkerk, A.O.; Sluijter, J.P.G.; Bakker, D.; Van Amersfoorth, S.; Van Veen, T.A.B.; Klerk, M.; Seppen, J.; et al. Cardiomyocyte Progenitor Cells as a Functional Gene Delivery Vehicle for Long-Term Biological Pacing. Molecules 2019, 24, 181. https://doi.org/10.3390/molecules24010181
Végh AMD, Den Haan AD, Cócera Ortega L, Verkerk AO, Sluijter JPG, Bakker D, Van Amersfoorth S, Van Veen TAB, Klerk M, Seppen J, et al. Cardiomyocyte Progenitor Cells as a Functional Gene Delivery Vehicle for Long-Term Biological Pacing. Molecules. 2019; 24(1):181. https://doi.org/10.3390/molecules24010181
Chicago/Turabian StyleVégh, Anna M. D., A. Dénise Den Haan, Lucía Cócera Ortega, Arie O. Verkerk, Joost P. G. Sluijter, Diane Bakker, Shirley Van Amersfoorth, Toon A. B. Van Veen, Mischa Klerk, Jurgen Seppen, and et al. 2019. "Cardiomyocyte Progenitor Cells as a Functional Gene Delivery Vehicle for Long-Term Biological Pacing" Molecules 24, no. 1: 181. https://doi.org/10.3390/molecules24010181