
Antitumor Effect of the Mannich Base(1,3-bis-((3-Hydroxynaphthalen-2-yl)phenylmethyl)urea) on Hepatocellular Carcinoma

Abstract
:1. Introduction
2. Results
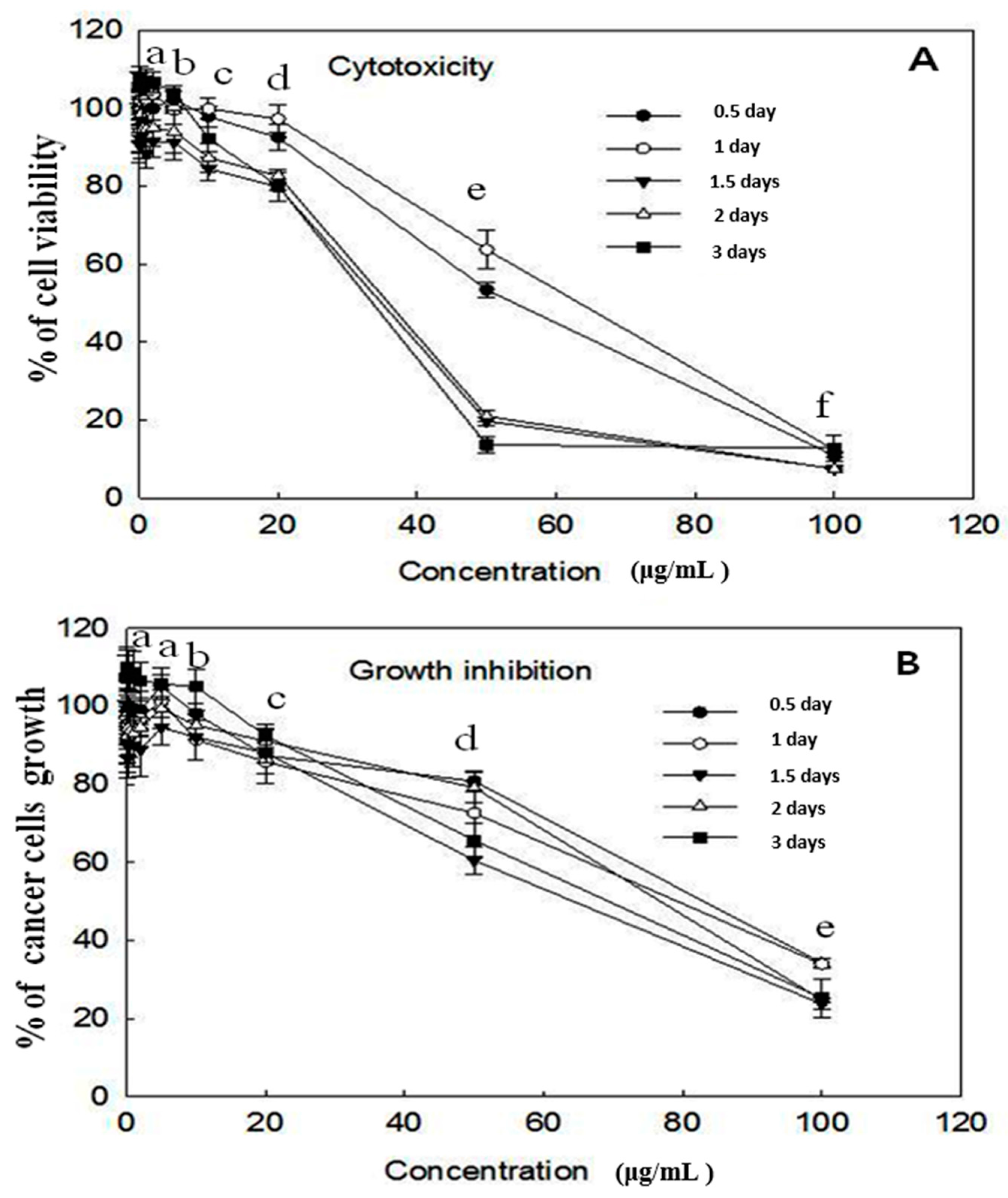
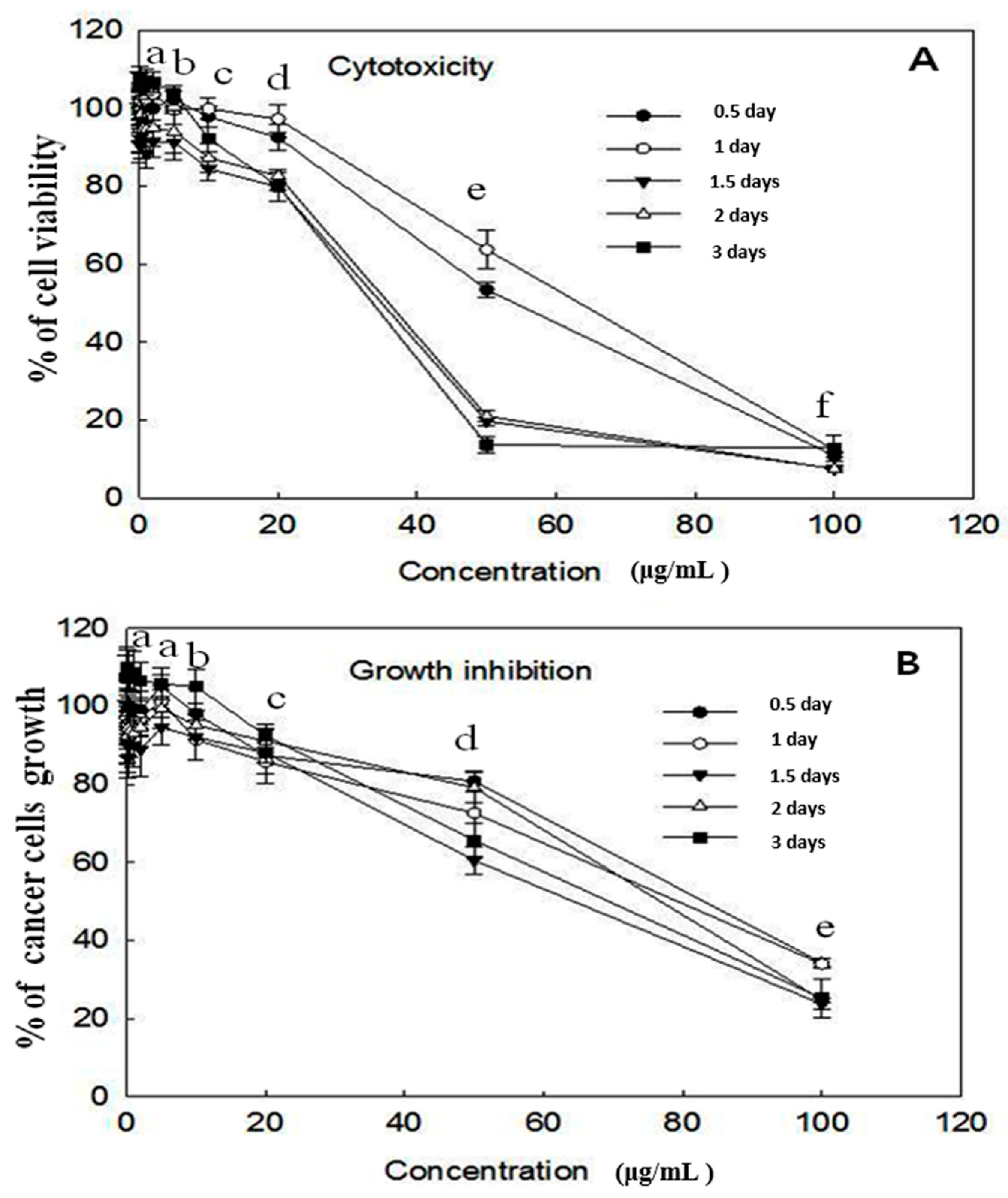
2.1. Cytotoxicity and Growth Inhibition Assay
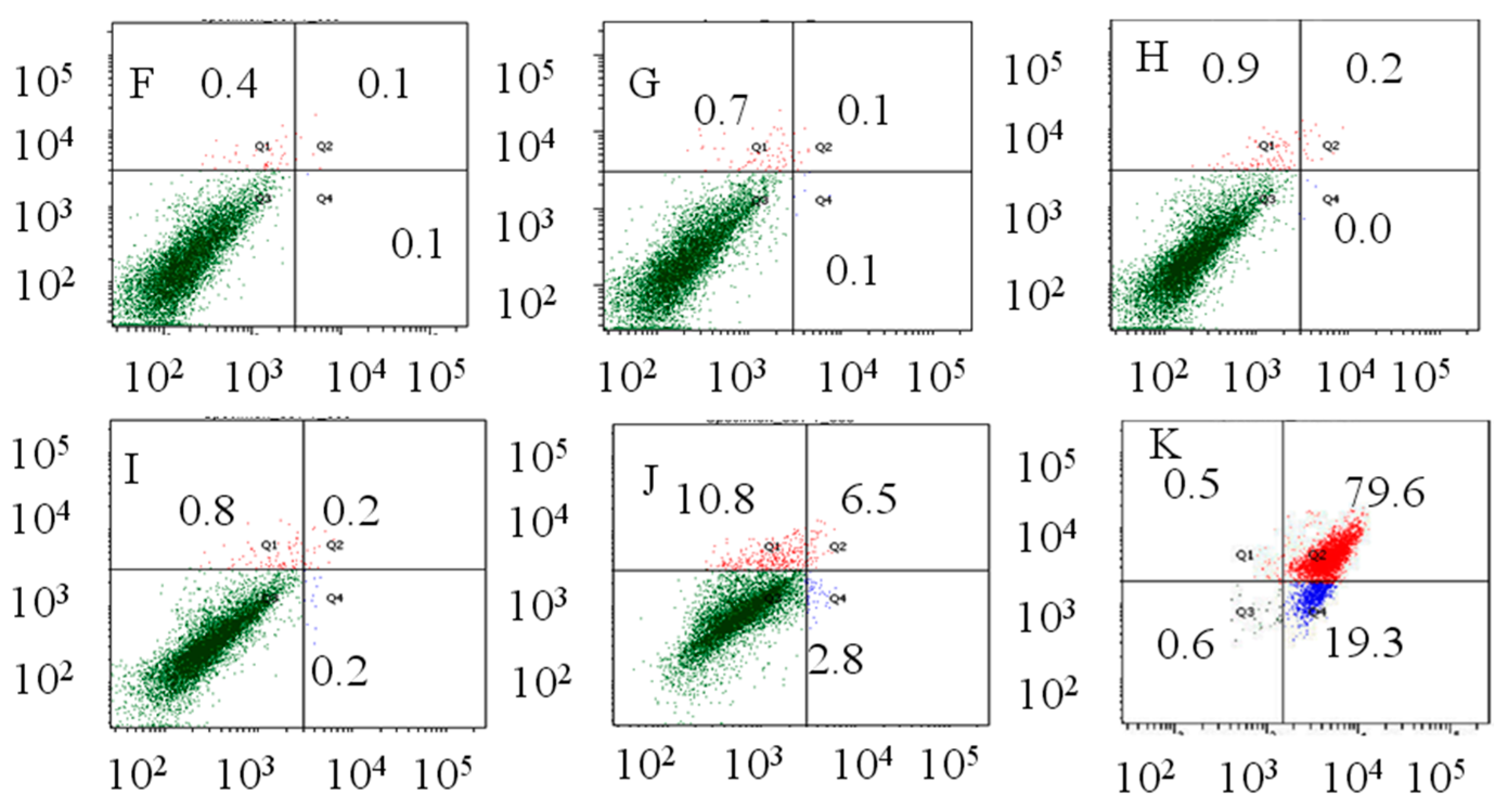
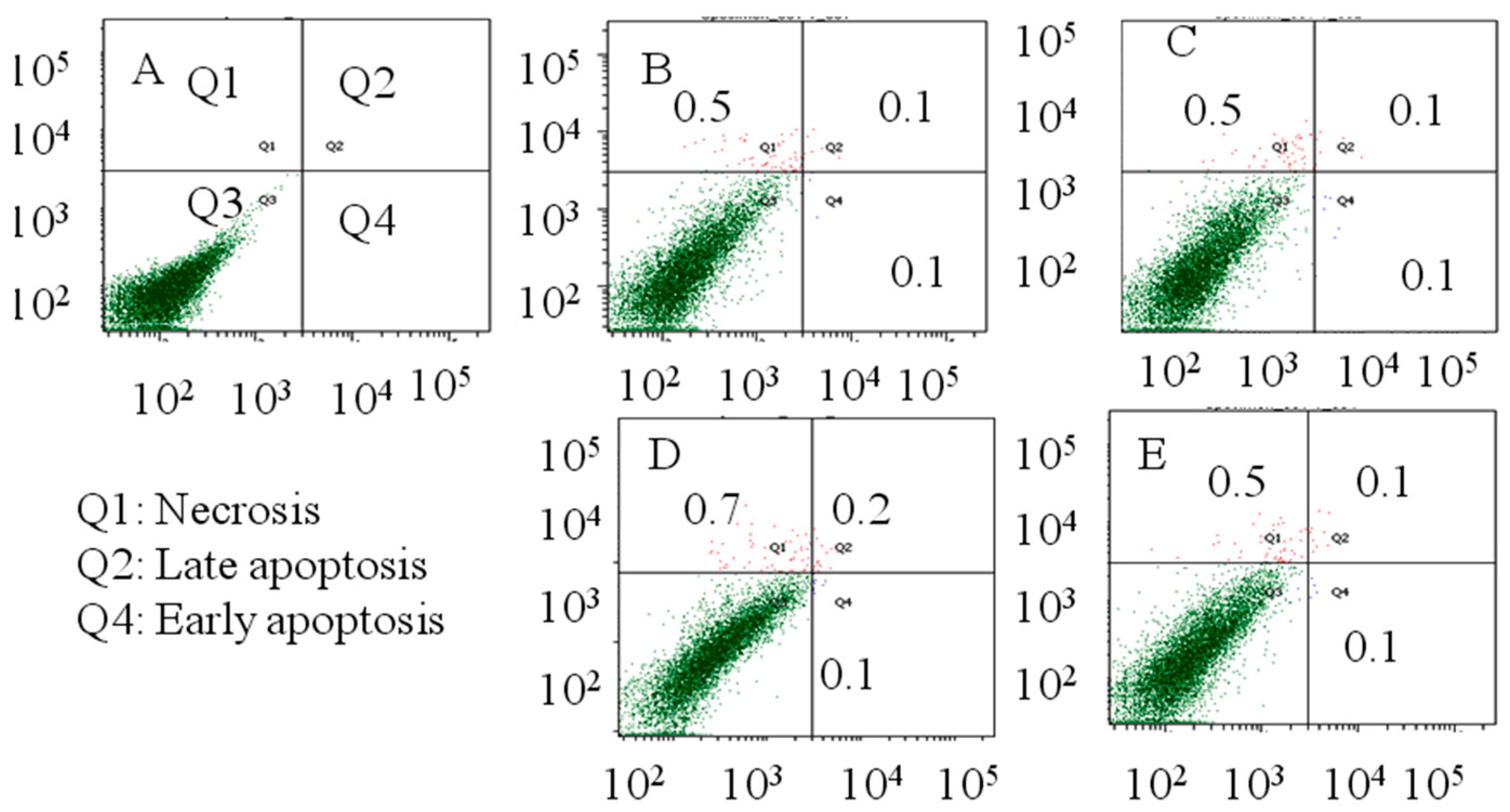
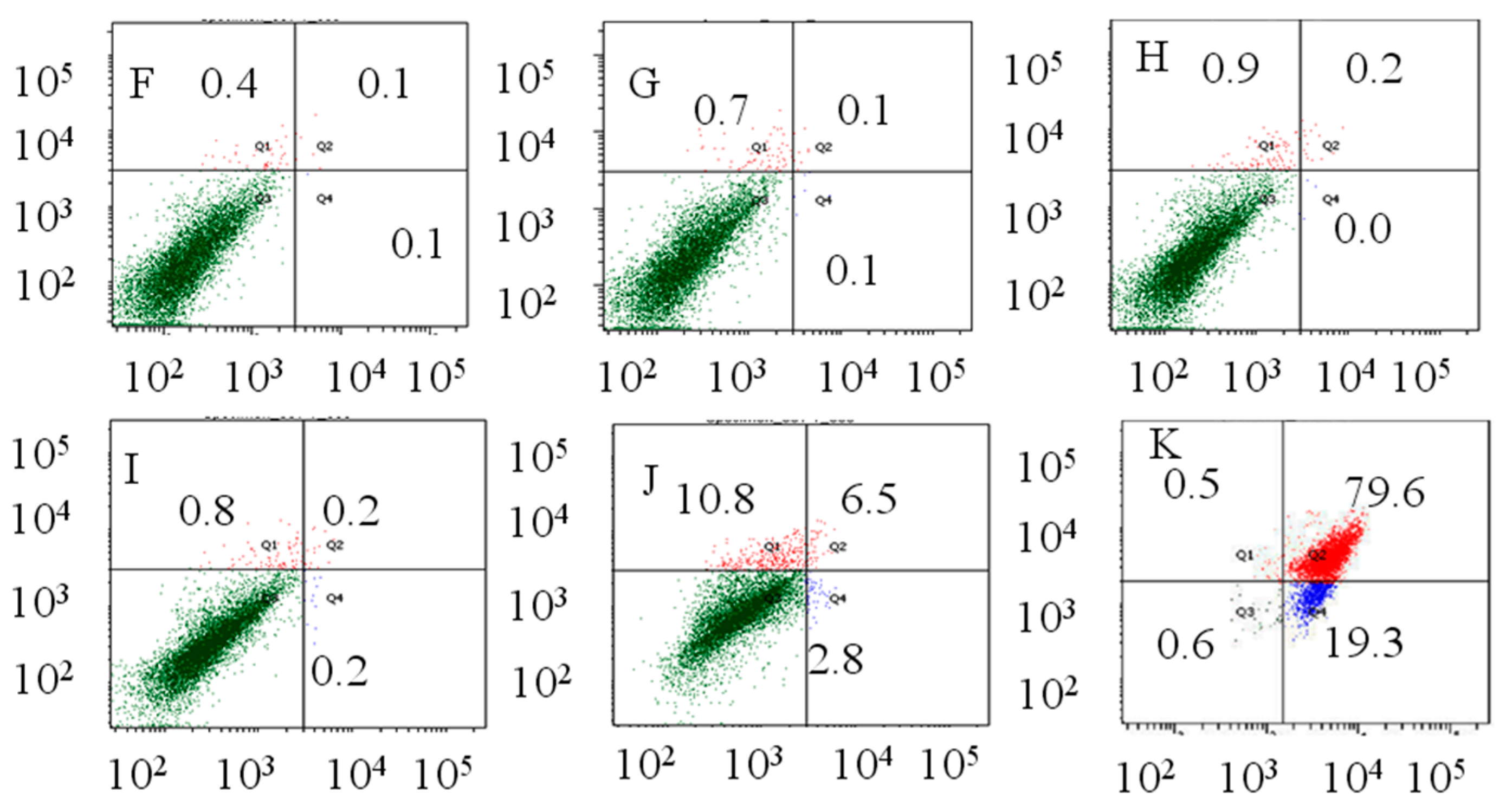
2.2. Flow Cytometry Analysis
2.3. Growth Conditions of the Rats
2.4. Inhibitory Effect of 1,3-BPMU on Tumor Growth in DEN-Induced Rats
2.5. Estimation of DNA and RNA
2.6. Estimation of Tumor Markers (AFP and CEA) Concentration
2.7. Activities of Serum Marker Enzymes, Lipid Peroxidation and Antioxidant Status
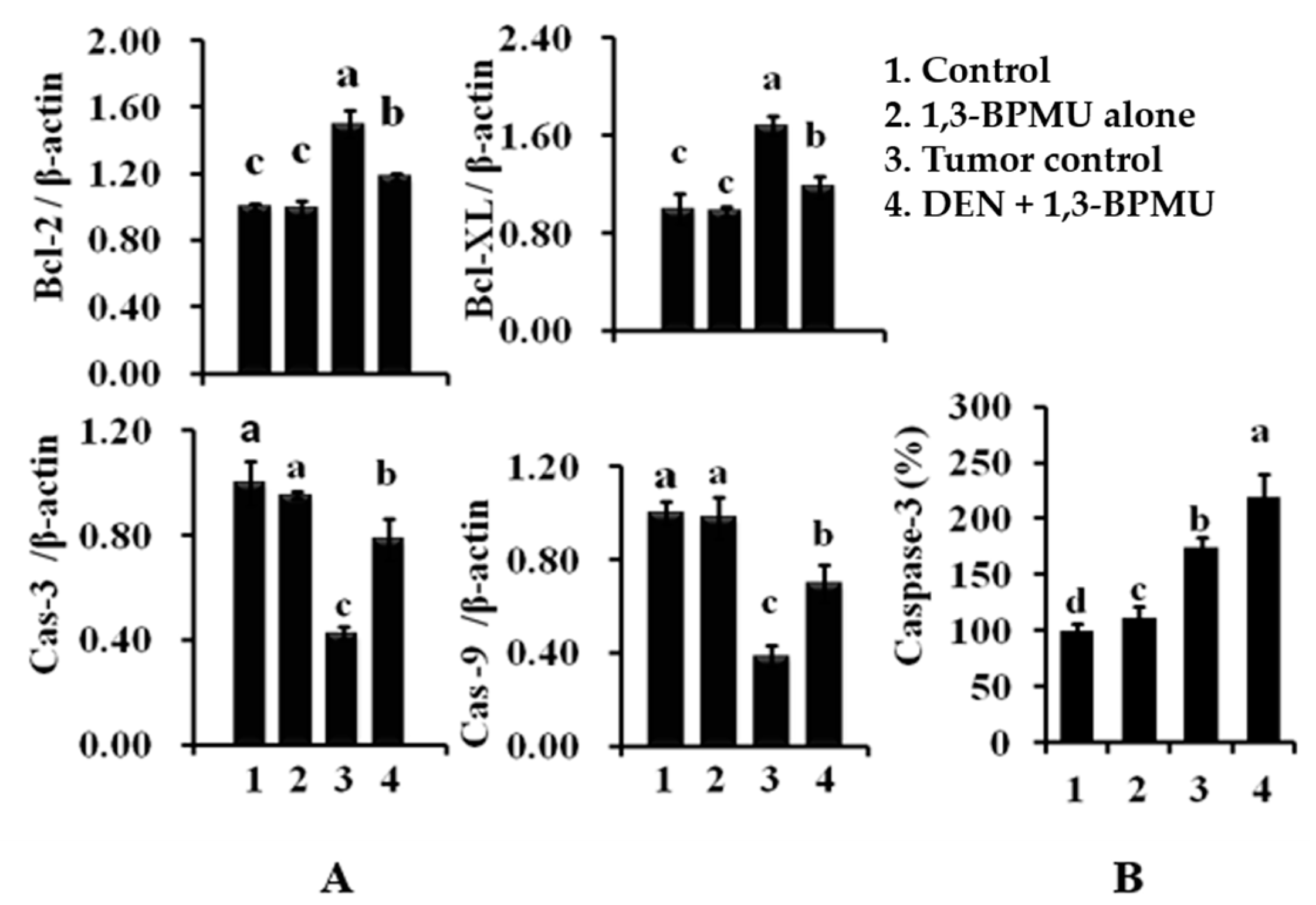
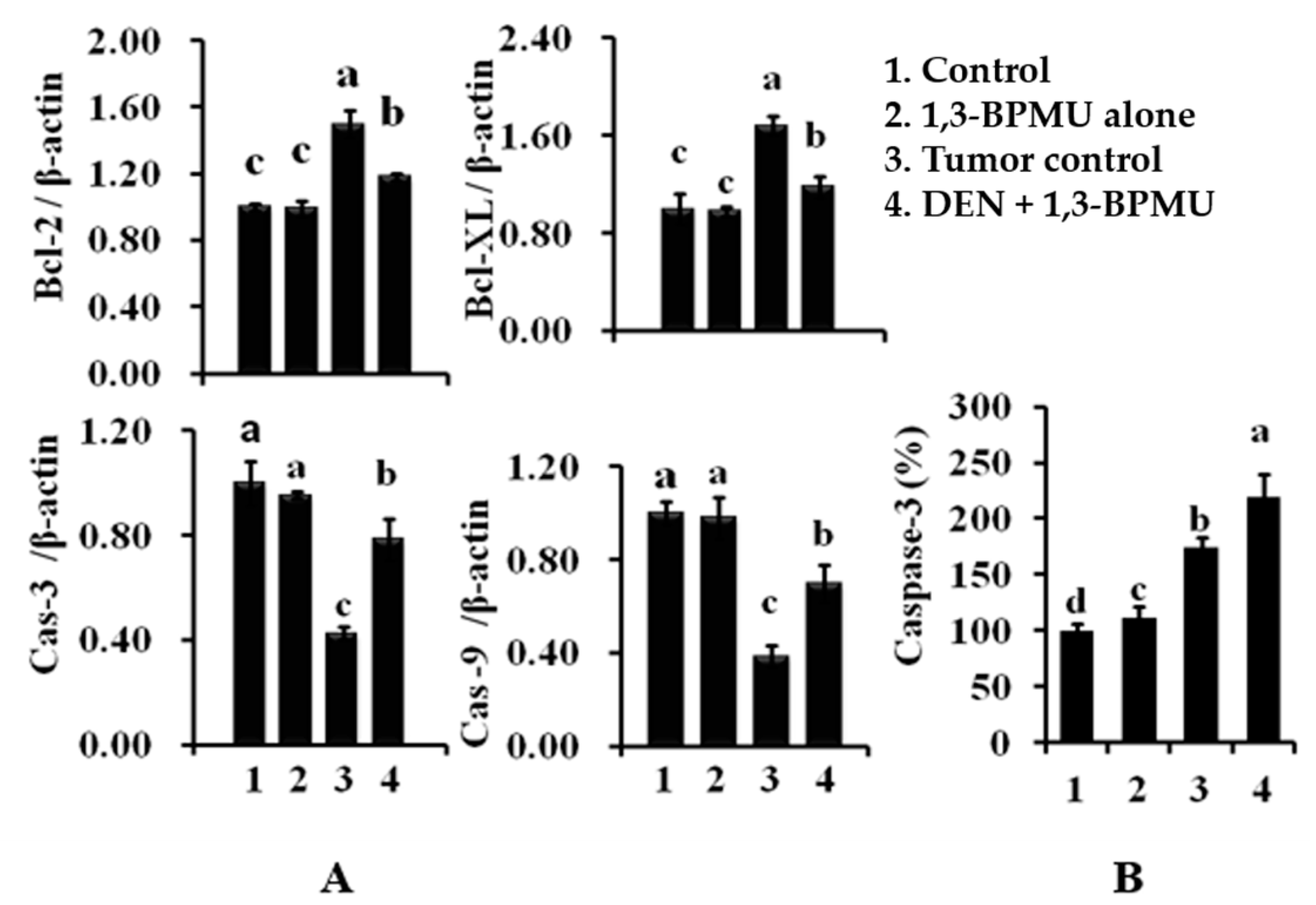
2.8. Apoptotic and Anti-Apoptotic Gene Expression Analysis by qPCR
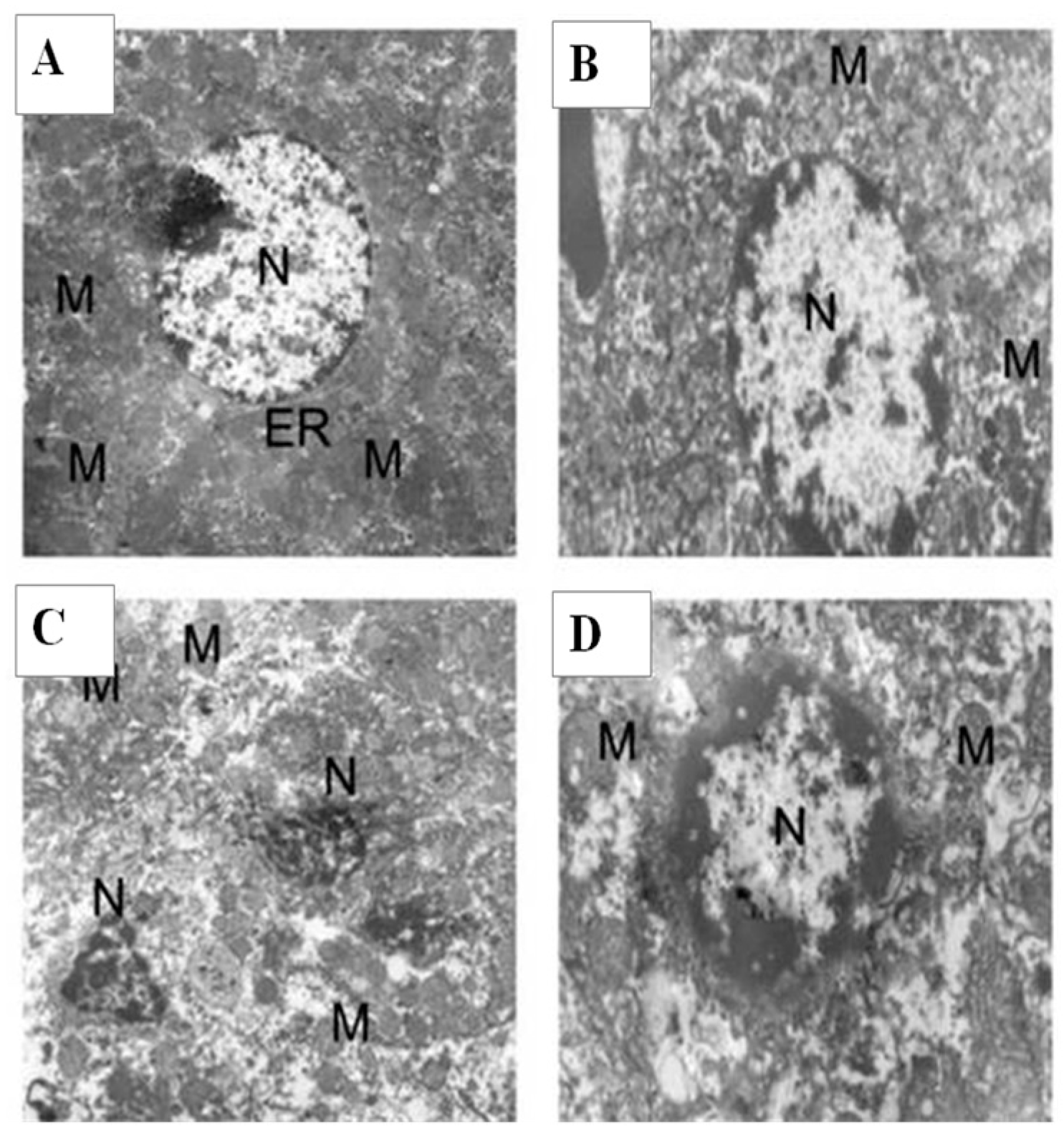
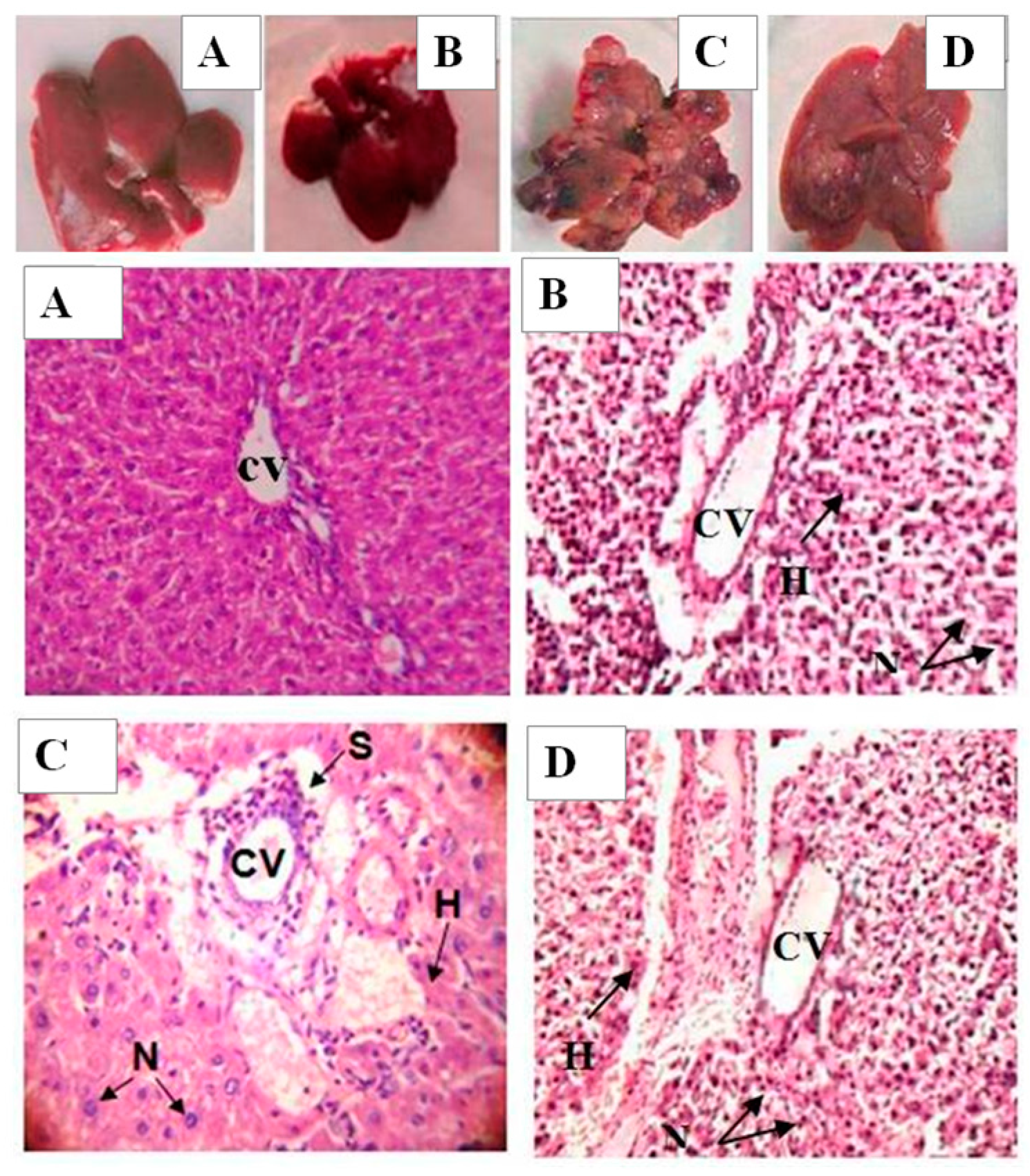
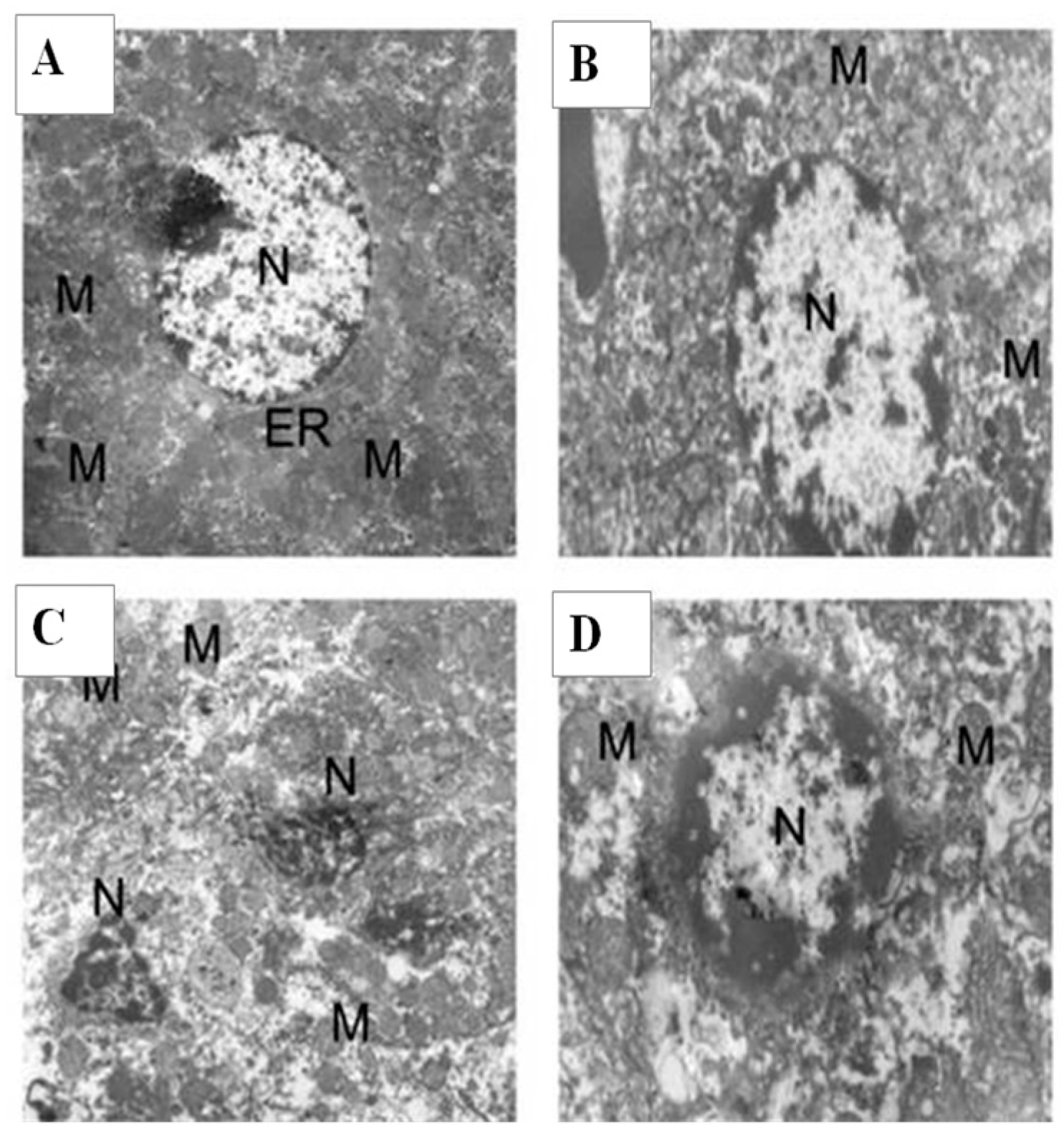
2.9. Histology and Ultrastructural Study
3. Discussion
4. Materials and Methods
4.1. Chemicals
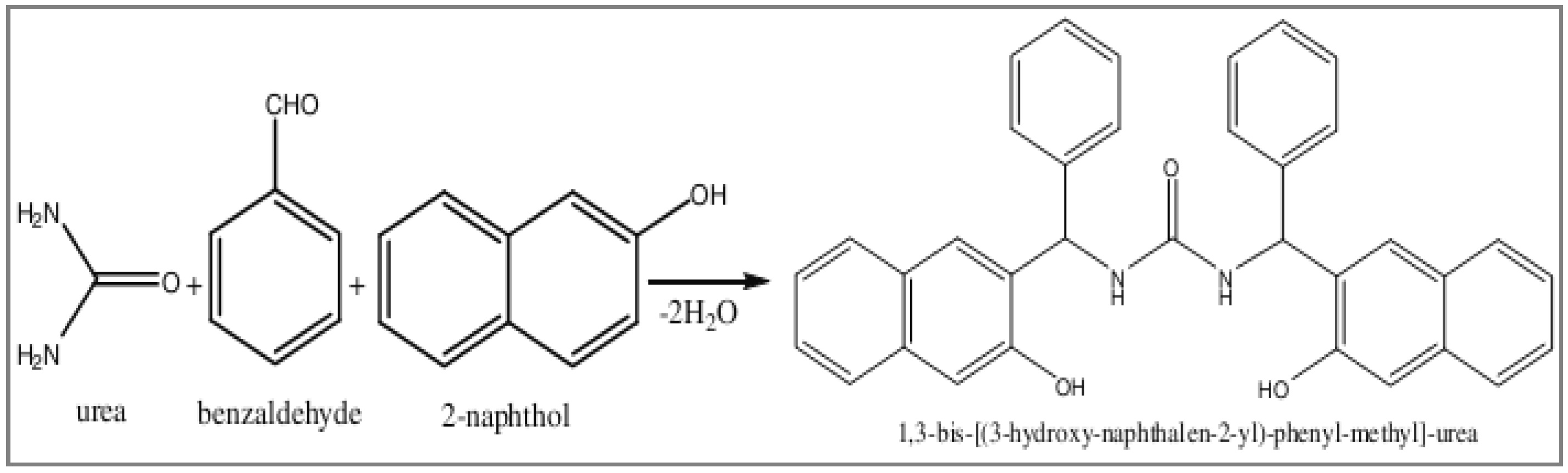
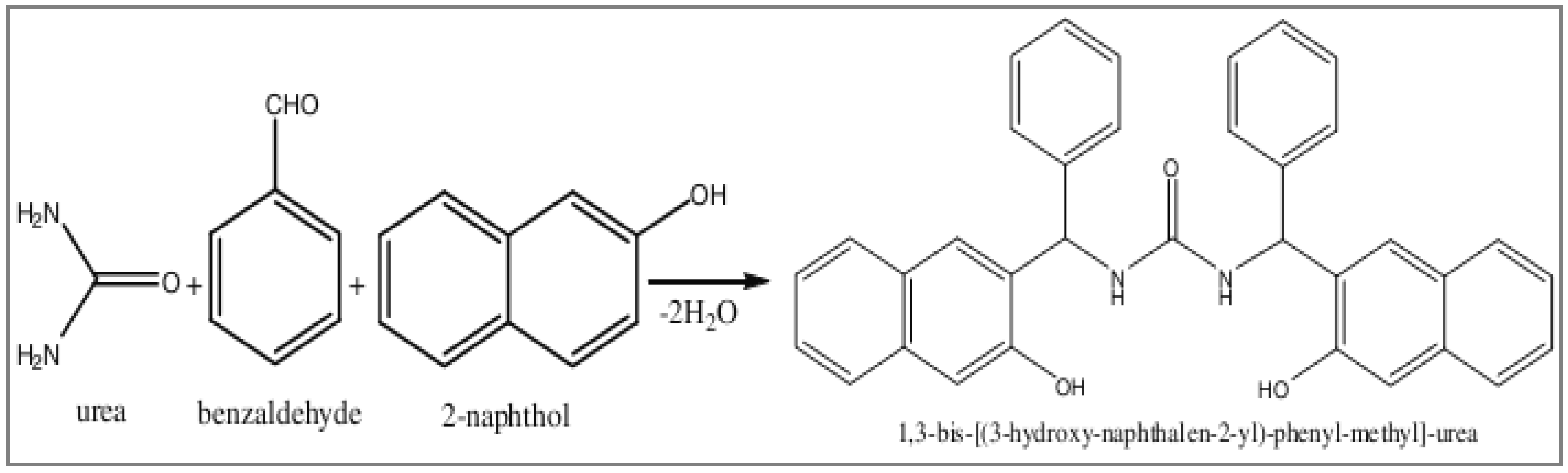
4.2. Synthesis of Mannich Base
4.3. Cytotoxicity and Growth Inhibition Assay
4.4. Flow Cytometric Analysis of Apoptosis
4.5. Animals
4.6. Experimental Design
- Group A: Normal rats fed with standard diet with ad libitum access to water.
- Group B: Rats received 1,3-BPMU alone (50 mg/kg/bw/day/orally based on the IC50) at 24-h intervals.
- Group C: Tumor control; hepatocarcinoma was induced by a single dose intraperitoneal (ip) injection of DEN (200 mg/kg/bw), and further, rats were administered with phenobarbital (250 mg/kg/bw/day/orally for two weeks) as a cancer-promoting agent [39].
- Group D: 1,3-BPMU-treated; hepatocarcinoma was induced by a single dose intraperitoneal (ip) injection of DEN (200 mg/kg/bw); then, rats were administered with phenobarbital (250 mg/kg/bw/day/orally) for two weeks as a cancer-promoting agent; then, animals were treated with 1,3-BPMU (50 mg/kg/bw/day/orally) up to the end of the experiment periods.
4.7. Analysis of Biochemical Parameters
4.8. Expression of Genetic Analysis by qPCR (Caspse-3, Caspase-9, Bcl-2 and Bcl-XL)
4.9. Caspase-3 Quantification
4.10. Histology and Ultra-Structural Investigation
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 1,3-BPMU: | 1,3-bis-[(3-hydroxy-naphthalen-2-yl)-phenyl-methyl]-urea |
| DEN: | Diethyl nitrosamine |
| HCC: | Hepatocarcinoma |
| AFP: | α-fetoprotein |
| CEA: | Carcinoembryonic antigen |
| AST: | Aspartate transaminase |
| ALT: | Alanine transaminase |
| ALP: | Alkaline phosphatase |
| LDH: | Lactate dehydrogenase |
| LPO: | Lipid peroxide |
| SOD: | Superoxide dismutase |
| CAT: | Catalase |
| GPx: | Glutathione peroxidase |
| GST: | Glutathione-S-transferase |
| GR: | Glutathione reductase |
| γ-GT: | γ-glutamyl transferase |
References
- Pascual, S.; Herrera, I.; Irurzun, L. New advances in hepatocellular carcinoma. World J. Hepatol. 2016, 28, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Bruix, J.; Sherman, M. Management of hepatocellular carcinoma. Hepatology 2005, 42, 1208–1236. [Google Scholar] [CrossRef] [PubMed]
- Waghray, A.; Murali, A.R.; Menon, K.N. Hepatocellular carcinoma: From diagnosis to treatment. World J. Hepatol. 2015, 18, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Blonski, W.; Kotlyar, D.S.; Forde, K.A. Non-viral causes of hepatocellular carcinoma. World J. Gastroenterol. 2010, 16, 3603–3615. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 1, 1397–1421. [Google Scholar] [CrossRef] [PubMed]
- Heindryckx, F.; Colle, I.; Van Vlierberghe, H. Experimental mouse models for hepato cellular carcinoma research. Int. J. Exp. Pathol. 2009, 90, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Ziech, D.; Franco, R.; Georgakilas, A.G.; Georgakila, S.; Malamou-Mitsi, V. The role of reactive oxygen species and oxidative stress in environmental carcinogenesis and biomarker development. Chem. Biol. Interact. 2010, 188, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Yeo, W.; Mok, T.S.; Zee, B.; Leung, T.W.; Lai, P.B.; Lau, W.Y.; Koh, J.; Mo, F.K.; Yu, S.C.; Chan, A.T.; et al. A Randomized Phase III Study of Doxorubicin versus Cisplatin/Interferon α-2b/Doxorubicin/Fluorouracil (PIAF) Combination Chemotherapy for Unresectable Hepatocellular Carcinoma. J. Natl. Cancer Inst. 2005, 97, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs overthe 30 Years from 1981 to 2010. J. Nat. Prod. 2005, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, K.W.; Napper, M.L.; Hoveyda, A.H. Combinatorial catalyst discovery. Curr. Opin. Chem. Biol. 1999, 3, 313–319. [Google Scholar] [CrossRef]
- Boucher, E.; Foner, A.; Reig, M.; Bruix, J. New drug for the treatment of hepatocellular carcinoma. Liver Int. 2009, 29, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Luo, H.; Wu, J.; Lan, L.B.; Fan, D.H.; Zhu, K.D.; Chen, X.Y.; Wen, M.; Liu, H.M. Galangin induces apoptosis of hepatocellular carcinoma cell via mitochondrial pathway. World J. Gastroenterol. 2010, 16, 3377–3384. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Pan, S.L.; Huang, Y.C.; Guh, J.H.; Chiang, P.C.; Huang, D.Y.; Kuo, S.C.; Lee, K.H.; Teng, C.M. CHM-1, a novel synthetic quinolone with potent and selective antimitotic antitumor activity against human hepatocellular carcinoma in vitro and in vivo. Mol. Cancer Ther. 2008, 7, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Tamilvendan, D.; Rajeswari, S.; Ilavenil, S.; Chakkaravarthy, K.; Venkatesa, P.G. Synthesis, spectral, crystallographic, antimicrobial, and anti-oxidant studies of few mannich bases. Med. Chem. Res. 2012, 21, 4129–4138. [Google Scholar] [CrossRef]
- Vadanasundari, V.; Ilavenil, S.; Ravikumar, S. Potential effect of bis-((hydroxyl-naphthalen-2-yl)-phenylmethyl)-urea (1,3BPMU) on diethyl nitrosamine (DEN) induced hepatocarcinoma in wistar albino rats. Int. J. Pharm. Biol. Sci. 2014, 5, 364–372. [Google Scholar]
- Le Couteur, D.G.; Warren, A.; Cogger, V.C.; Smedsrod, B.; Sorensen, K.K.; De Cabo, R.; Fraser, R.; McCuskey, R.S. Old Age and the Hepatic Sinusoid. Anat Rec. 2008, 291, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Rayburn, E.R.; Velu, S.E.; Nadkarni, D.H.; Murugesan, S.; Zhang, R. In vitro and in vivo Anticancer Activity of Novel Synthetic Makaluvamine Analogues. Clin. Cancer Res. 2009, 15, 3511–3518. [Google Scholar] [CrossRef] [PubMed]
- Tao, D.; Wu, J.; Feng, Y.; Qin, J.; Hu, J.; Gong, J. New method for the analysis of cell cycle-specific apoptosis. Cytom. Part 2004, 57, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Vina, J.; Gomez-Cabrera, M.C.; Lloret, A.; Marquez, R.; Minana, J.B.; Pallardo, F.V.; Sastre, J. Free radicals in exhaustive physical exercise: Mechanism of production, and protection by antioxidants. IUBMB Life 2000, 50, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Tayob, N.; Lok, A.S.; Do, K.A.; Feng, Z. Improved detection of hepatocellular carcinoma using a longitudinal α-fetoprotein screening algorithm. Clin. Gastroenterol. Hepatol. 2016, 14, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Rich, N.; Singal, A.G. Hepatocellular carcinoma tumor markers: Current role and expectations. Best Pract. Res. Clin. Gastroenterol. 2014, 28, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Morrison, C.; Marsh, W., Jr.; Frankel, W.L. A comparison of CD10 to pCEA, MOC-31, and hepatocyte for the distinction of malignant tumors in the liver. Mod. Pathol. 2002, 12, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, O.; Cacho, B.; Morales-Espinosa, D.; Ruelas-Villavicencio, A.; Flores-Estrada, D.; Hernandez-Pedro, N. The progressive elevation of α-fetoprotein for the diagnosis of hepatocellular carcinoma in patients with liver cirrhosis. BMC Cancer 2007, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Yang, H.; Zhang, R.; Zhao, J.J.; Hao, D.J. Tumor-associated antigens and their anti-cancer applications. Eur. J. Cancer Care (Engl.) 2016. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Jeon, C.H.; Ko, G.; Kim, J.; Sohn, D.H. Protective effect of curcumin in rat liver injury induced by carbon tetrachloride. J. Pharm. Pharmacol. 2000, 52, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.H.; Keeffe, E.B.; Frederiks, W.M.; Van Noorden, C.J.; Aronson, D.C.; Marx, F.; Bosch, K.S.; Jonges, G.N.; Vogels, I.M.; James, J. Quantitative changes in acid phosphatase, alkaline phosphatase and 5’nucleodidase activity in rat liver after experimentally induced cholestasis. Liver 1990, 10, 158–166. [Google Scholar]
- Kamdem, L.; Siest, G.; Magdalou, J. Differential toxicity of aflotoxin B1 in male and female rats’ relationship with hepatic drug metabolizing enzymes. Biochem. Pharmacol. 1982, 31, 3057–3062. [Google Scholar] [CrossRef]
- Esterbauer, H.; Chessman, K.H. Determination of aldehydic lipid peroxidation products: Malonoldehyde and 4-hydroxy-nonenal. Methods Enzymol. 1990, 186, 407–421. [Google Scholar] [PubMed]
- Revathi, R.; Manju, V. Effects of umbelliferone on lipid peroxidation and antioxidant status in diethylnitrosamine-induced hepatocellular carcinoma. J. Acute Med. 2013, 3, 73–82. [Google Scholar]
- Jiang, B.; Xiao, W.; Shi, Y.; Liu, M.; Xiao, X. Heat shock pretreatment inhibited the release of Smac/DIABLO from mitochondria and apoptosis induced by hydrogen peroxide in cardiomyocytes and C2C12 myogenic cells. Cell Stress Chaperones. 2005, 10, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Klaunig, J.F.; Kamendulis, L.M. The role of oxidative stress in carcinogenesis. Ann. Rev. Pharmacol. Toxicol. 2004, 44, 239–267. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Kim, H.P.; Hoetzel, A.; Park, J.W.; Nakahira, K. Mechanisms of cell death in oxidative stress. Antioxid. Redox Signal. 2007, 9, 49–89. [Google Scholar] [CrossRef] [PubMed]
- Ozen, T.; Korkmaz, H. Modulatory effect of Urtica dioica L (Urticaceae) leaf extraction biotransformation of enzyme systems, antioxidant enzymes, and LDH and lipid peroxidation in mice. Phytomedicine 2003, 10, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.J.; Shi, Y. Molecular mechanisms of caspase regulation during apoptosis. Nat. Rev. Mol. Cell Biol. 2004, 5, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Earnshaw, W.C.; Martins, L.M.; Kaufmann, S.H. Mammalian caspases: Structure, activation, substrates, and functions during apoptosis. Ann. Rev. Biochem. 1999, 68, 383–424. [Google Scholar] [CrossRef] [PubMed]
- Vadanasundari, V.; Karthik, D.; Ilavenil, S.; Ravikumar, S. Identification of Differential Protein Expression in Hepatocellular Carcinoma Induced Wistar Albino Rats by 2D Electrophoresis and MALDI-TOF–MS Analysis. Indian J. Clin. Biochem. 2015. [Google Scholar] [CrossRef]
- Tavakoli, M.; Fatemeh, Y.; Tehrani, K. Apoptosis Induced by 13-S-hydroxy octadecadienoic acid in the breast cancer cell lines, MCF-7 and MDA-MB-231. Iran. J. Basic Med. Sci. 2013, 16, 661–669. [Google Scholar]
- Su, N.W.; Lin, Y.L.; Lee, M.H.; Ho, C.Y. Ankaflavin from Monascus-fermented red rice exhibits selective cytotoxic effect and induces cell death on Hep G2 cells. J. Agric. Food Chem. 2005, 53, 1949–1954. [Google Scholar] [CrossRef] [PubMed]
- Peraino, C.; Fry, R. J.M.; Staffeldt, E. Reduction and enhancement by phenobarbital of hepatocarcinogenesis inducedin the rat by 2-acetylaminofluorene. Cancer Res. 1971, 31, 1506–1512. [Google Scholar]
- Bishayee, A.; Dhir, N. Resveratrol-mediated chemoprevention of diethy lnitro samine-initiated hepatocarcinogenesis: Inhibition of cell proliferation and induction of apoptosis. Chem. Biol. Interact. 2009, 179, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Stuhr, L.E.; Iversen, V.V.; Straume, O.; Maehle, B.O.; Reed, R.K. Hyperbaric oxygen alone or combined with 5-FU attenuates growth of DMBA-induced rat mammary tumors. Cancer Lett. 2004, 210, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Sivaramakrishnan, V.; Shilpa, P.N.; Praveenkumar, V.R.; Niranjali devaraj, S. Attenuation of N-nitrodiethylamine-induced hepatocellular carcinogenesis by a noval flavonol—Morin. Chem. Biol. Interact. 2008, 171, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Karthik, D.; Ravikumar, S. Proteome and phytochemical analysis of Cynodon dactylon leaves extract and its biological activity in diabetic rats. Biomed. Prev. Nutr. 2011, 1, 49–56. [Google Scholar] [CrossRef]
- Graham, L.; Orenstein, J.M. Processing tissue and cells for transmission electron microscopy in diagnostic pathology and research. Nat. Protocol. 2007, 2, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Cogger, V.C.; Mross, P.E.; Hosie, M.J.; Ansselin, A.D.; McLean, A.J.; Le Couteur, D.G. The effect of acute oxidative stress on the ultra-structure of the perfused rat liver. Pharmacol. Toxicol. 2001, 89, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (g) | Control | 1,3-BPMU Alone | Tumor Control | DEN + 1,3-BPMU * |
|---|---|---|---|---|
| Initial | 156 ± 10.21 a | 162 ±12.8 a | 157 ± 12.9 b | 159 ± 12.5 c |
| Final | 202 ± 11.20 a | 205 ± 9.20 a | 184 ± 11.8 b | 208 ± 10.8 c |
| Liver | 7.45 ± 0.59 a | 8.20 ± 0.65 a | 13.29 ±1.2 b | 8.32 ± 0.67 c |
| Groups | Average Number of Nodules in Liver | Volume of Nodules (mm3) | |
|---|---|---|---|
| (>2 mm) | (>4 mm) | ||
| Normal control | 0 a | 0 a | 0 a |
| 1,3-BPMU alone | 0 a | 0 a | 0 a |
| Tumor control | 58.65 ± 1.02 b | 6.3 ± 0.05 b | 192.55 ± 2.85 b |
| DEN + 1,3-BPMU * | 34.21 ± 0.98 c | 2.6 ± 0.03 c | 72.25 ± 0.24 c |
| Nucleic Acids (mg/g Wet Tissue) | Normal Control | 1,3-BPMU Alone | Tumor Control | DEN + 1,3-BPMU * |
|---|---|---|---|---|
| DNA | 1.8 ± 0.04 a | 5.72 ± 0.14 a | 8.10 ± 0.08 b | 2.28 ± 0.13 c |
| RNA | 1.3 ± 0.09 a | 2.70 ± 0.07 a | 5.3 ± 0.20 b | 1.58 ± 0.08 c |
| Groups | AFP (ng/mL) | CEA (ng/mL) |
|---|---|---|
| Normal control | 0.05 ± 0.003 a | 0.04 ± 0.003 a |
| 1,3-BPMU alone | 0.05 ± 0.004 a | 0.03 ± 0.005 a |
| Tumor control | 1.58 ± 0.07 b | 2.47 ± 0.050 b |
| DEN + 1,3-BPMU * | 0.43 ± 0.05 c | 0.18 ± 0.090 c |
| Groups | AST (U/L) | ALT (U/L) | ALP (U/L) | LDH (U/L) | γGT (U/L) | 5’NT (U/L) |
|---|---|---|---|---|---|---|
| Normal control | 44.0 ± 2.42 a | 27.08 ± 1.08 a | 33.17 ± 1.21 a | 5.9 ± 0.36 a | 6.09 ± 0.32 a | 3.08 ± 0.38 a |
| 1,3-BPMU alone | 43.0 ± 1.63 a | 26.05 ± 1.11 a | 34.12 ± 1.12 a | 5.6 ± 0.35 a | 6.08 ± 0.33 a | 3.0 ± 0.38 a |
| Tumor control | 192.2 ± 2.03 b | 82.36 ± 1.67 b | 75.46 ± 1.39 b | 12.3 ± 0.30 b | 12.38 ± 1.05 b | 6.19 ± 0.78 b |
| DEN + 1,3-BPMU * | 98.2 ± 3.79 c | 36.04 ± 1.33 c | 34.23 ± 1.99 c | 8.1 ± 0.24 c | 5.13 ± 0.46 c | 4.38 ± 0.51 c |
| Groups | LPO | SOD (U/mg) | CAT (U/mg) | GPx (U/mg) | GR (U/mg) | GST (U/mg) |
|---|---|---|---|---|---|---|
| Normal control | 2.97 ± 0.11 a | 3.99 ± 0.19 a | 4.82 ± 0.23 a | 6.21 ± 0.43 a | 5.28 ± 0.26 a | 0.89 ± 0.02 a |
| 1,3-BPMU alone | 2.42 ± 1.16 a | 4.35 ± 0.21 a | 4.61 ± 0.22 a | 6.72 ± 0.33 a | 5.63 ± 0.28 a | 0.96 ± 0.02 a |
| Tumor control | 3.96 ± 0.19 b | 2.56 ± 0.27 b | 3.88 ± 0.28 b | 2.63 ± 0.38 b | 3.82 ± 0.19 b | 0.59 ± 0.01 b |
| DEN + 1,3-BPMU * | 2.99 ± 0.14 c | 4.87 ± 0.24 c | 4.72 ± 0.24 c | 5.73 ± 0.33 c | 4.83 ± 0.24 c | 0.71 ± 0.02 c |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vedarethinam, V.; Dhanaraj, K.; Ilavenil, S.; Arasu, M.V.; Choi, K.C.; Al-Dhabi, N.A.; Srisesharam, S.; Lee, K.D.; Kim, D.H.; Dhanapal, T.; et al. Antitumor Effect of the Mannich Base(1,3-bis-((3-Hydroxynaphthalen-2-yl)phenylmethyl)urea) on Hepatocellular Carcinoma. Molecules 2016, 21, 632. https://doi.org/10.3390/molecules21050632
Vedarethinam V, Dhanaraj K, Ilavenil S, Arasu MV, Choi KC, Al-Dhabi NA, Srisesharam S, Lee KD, Kim DH, Dhanapal T, et al. Antitumor Effect of the Mannich Base(1,3-bis-((3-Hydroxynaphthalen-2-yl)phenylmethyl)urea) on Hepatocellular Carcinoma. Molecules. 2016; 21(5):632. https://doi.org/10.3390/molecules21050632
Chicago/Turabian StyleVedarethinam, Vadanasundari, Karthik Dhanaraj, Soundharrajan Ilavenil, Mariadhas Valan Arasu, Ki Choon Choi, Naif Abdullah Al-Dhabi, Srigopalram Srisesharam, Kyung Dong Lee, Da Hye Kim, Tamilvenvendan Dhanapal, and et al. 2016. "Antitumor Effect of the Mannich Base(1,3-bis-((3-Hydroxynaphthalen-2-yl)phenylmethyl)urea) on Hepatocellular Carcinoma" Molecules 21, no. 5: 632. https://doi.org/10.3390/molecules21050632