Antiproliferative and Molecular Mechanism of Eugenol-Induced Apoptosis in Cancer Cells

Abstract
:1. Introduction
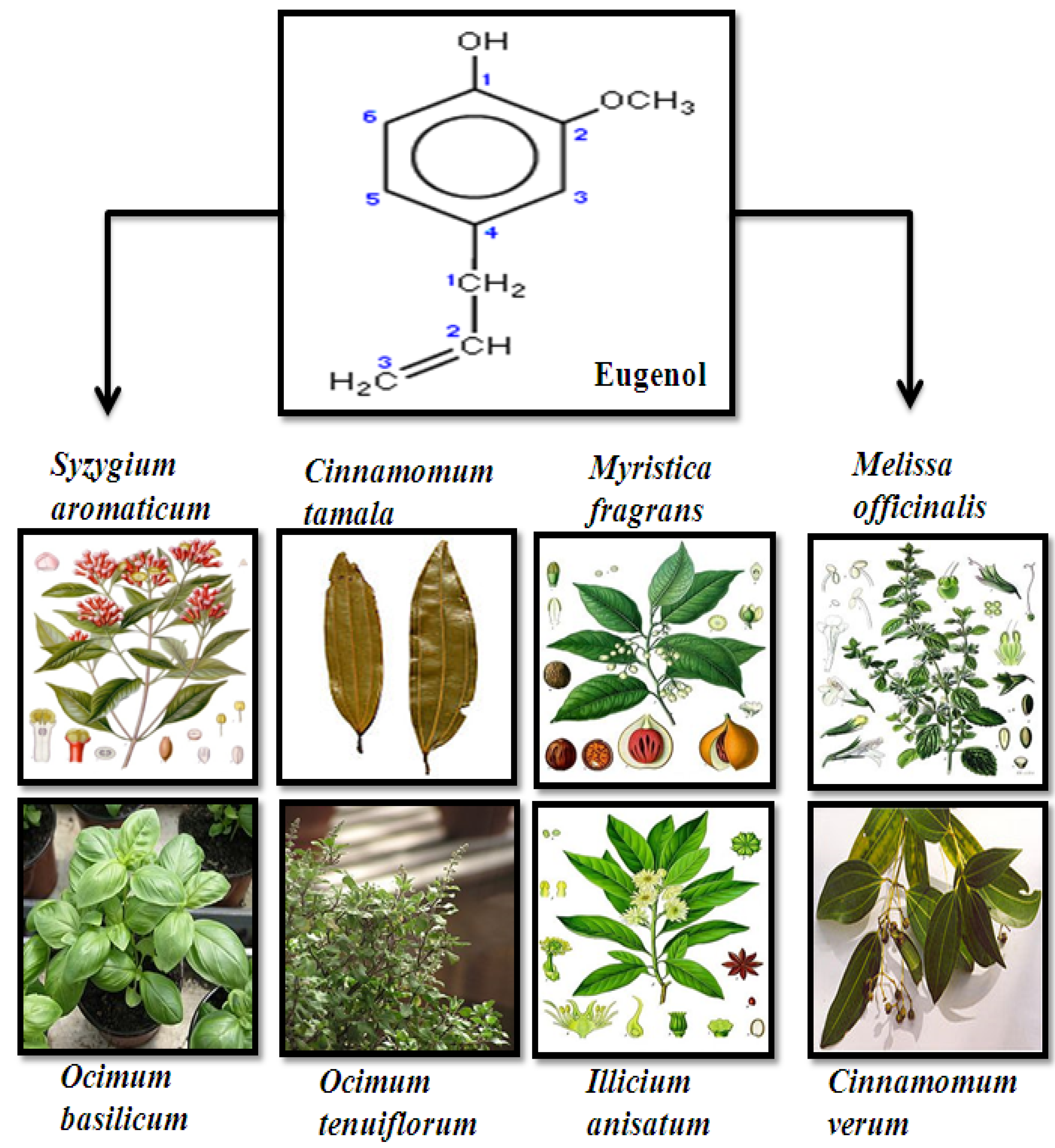
2. Source, Chemistry and Structure of Eugenol
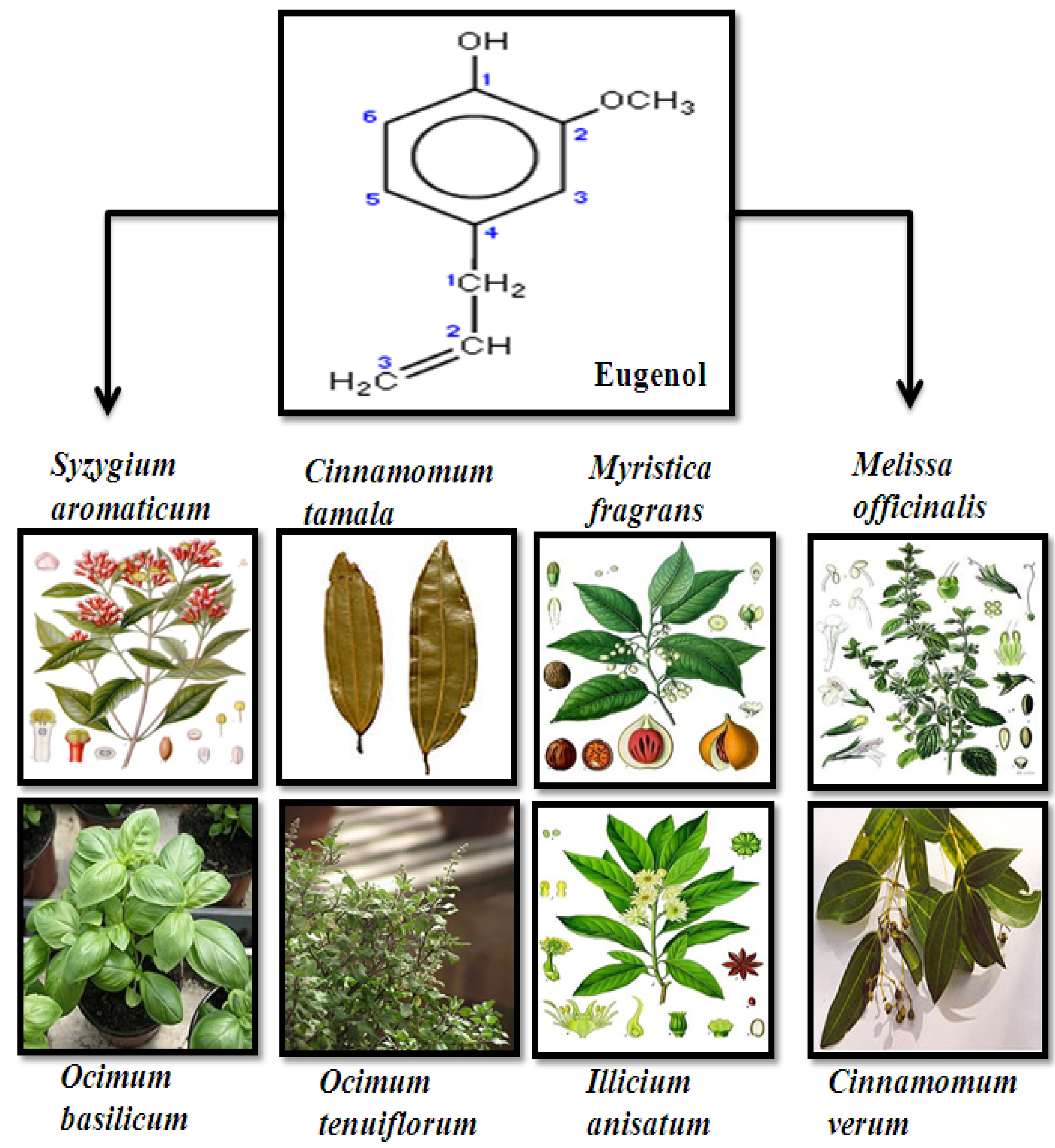
3. Anti-Proliferative and Molecular Mechanism of Eugenol-Induced Apoptosis
3.1. Anti-Proliferative Mechanism of Eugenol against Melanoma Cells
3.2. Antiproliferative Mechanism of Eugenol against Skin Tumors
3.3. Antiproliferative Mechanism of Eugenol against Osteosarcoma Cells
3.4. Antiproliferative Mechanism of Eugenol against Leukemia Cells
3.5. Antiproliferative Mechanism of Eugenol against Gastric Cancer
3.6. Antiproliferative Mechanism of Eugenol against Mast Cells
3.7. Antiproliferative Activity of Eugenol against Other Cancer Cells and Animal Model
4. Conclusions

{kind=link}
| Tested Compound | Cancer Type | Observations/Results | Reference |
|---|---|---|---|
| Eugenol/Isoeugenol | Melanoma | ► no significant activity at 100 µM | [8] |
| Dehydrodieugenol | Melanoma | ► 40–60% growth inhibition | [8] |
| O, O'-Dimethyl-dehydrodieugenol (S7) | Melanoma | ► 70–80% growth inhibition | [8] |
| 6,6'-Dibromo-dehydrodieugenol | Melanoma | ► nearly 100% growth inhibition | [8] |
| ► The IC50 of S7-S against WM, GR, PNP, GILIN, LAN-5 cell lines were found to be 27, 23, 29, 19 and 16 µM respectively | |||
| ► 50 µM of S7-S exposure for 24 h resulted in DNA fragmentation as detected by TUNEL assay | |||
| Eugenol | Melanoma | ► cytotoxic effect was observed in G361 cells in the range of 0.5 to 2 mM | [21] |
| ► caspase-3 and caspase-6 activation | |||
| ► caspase’s substrate like DFF45, PARP, Lamin A were cleaved | |||
| Eugenol | Melanoma | ► concentration of 0.5 µM inhibited 50% cell growth in Sbcl2 and WM3211 after 24 h | [22] |
| ► in B16 melanoma Xenograft, it caused significant tumor decrease (almost 40%) | |||
| ► TUNEL assay of WM1205Lu cells confirmed apoptosis | |||
| ► E2F family of transcription factors have a role in the apoptosis | |||
| Eugenol | Osteosarcoma | ► inhibited the HOS cell proliferation both in dose and time-dependent manner | [25] |
| ► increased levels of p53, caspase 3 and cleaved PARP | |||
| ► cleavage of lamin A and cytosolic reduction of DFF-45 | |||
| Eugenol | Leukemia | ► concentration of 23.7 µM inhibited 50% cell growth in HL-60 | [26] |
| ► increased ROS generation and GSH depletion | |||
| ► increased bax translocation, bcl2 reduction, cytochrome c release and caspase-9 and -3 activation | |||
| Eugenol | Gastric cancer | ► decreased the expression of NF-κB (p50 and p65), pIκBα and IKKβ and increased the expression of IκBα | [28,29] |
| ► decreased the expression of cyclin D1, cyclin B, and PCNA and increased the expression of p21, p53 and Gadd45 | |||
| ► decreased Bcl2 and Bcl-xL expression, and increased the expression of Bax, Bid, Bad, Apaf-1, cytochrome C, and caspase-9, and -3 and PARP | |||
| ► decreased the activities of MMPS (expression of MMP-2, MMP-9), VEGF and VEGFR1 and increased the expression of TIMP-2 and RECK | |||
| Eugenol | Skin tumor | ► number of mice that developed tumors was less in the eugenol treated group (42%) | [23] |
| ► resulted in the down-regulation of c-Myc, H-ras and Bcl2 expression along with up-regulation of p53, Bax and active caspase-3 expression in the skin lesions | |||
| Eugenol | Skin tumor | ► PCNA and TUNEL analysis confirmed apoptosis | [24] |
| ► recovery of cellular GSH and various enzymes like activities of GR, CAT, GPX, GST, and XO | |||
| ► increased p53 and p21 WAF1 levels | |||
| ► inhibition of ODC activity, iNOS, COX-2 expression | |||
| ► decreased levels of pro-inflammatory cytokines (IL-6, TNF-α, PGE-2) and NF-κB | |||
| Eugenol | Mast cells | ► half maximal inhibition (IC50) of Eugenol in RBL-2H3 cells was found to be 700 µM | [30] |
| ► apoptosis induction was confirmed by DNA ladder, activation of caspase-3 and the cleavage product of PARP 85 kda | |||
| ► phospho-ser 15-p53 plays a pivotal role in eugenol-induced apoptosis of RBL-2H3 cells | |||
| Eugenol | Prostate cancer | ► combinational effect of eugenol along with 2-methoxyestradiol (2-ME) against prostate cancer cells | [32] |
| ► cell cycle analysis displayed significant increase of G2M phase by 4.6-fold, when eugenol was combined with 2-ME | |||
| ► Bax and Bcl2 had a role in the synergistic combination |
Acknowledgements
References
- Sporn, M.B.; Newton, D.L. Chemoprevention of cancer with retinoids. Fed. Proc. 1979, 38, 2528–2534. [Google Scholar]
- Kelloff, G.J. Perspectives on cancer chemoprevention research and drug development. Adv. Cancer Res. 1999, 78, 320–334. [Google Scholar]
- Kelloff, G.J.; Boone, C.W. Cancer chemopreventive agents: Drug development status and future prospects. J. Cell. Biochem. 1994, 20, 1–303. [Google Scholar] [CrossRef]
- Kelloff, G.J.; Hawk, E.T.; Sigman, C.C. Cancer Chemoprevention: Promising Cancer Chemopreventive Agents; Humana Press: Totowa, NJ, USA, 2004. [Google Scholar]
- Ferguson, L.R. Antimutagens as cancer chemopreventive agents in the diet. Mutat. Res. 1994, 307, 395–410. [Google Scholar] [CrossRef]
- Ferguson, L.R.; Philpott, M.; Karunasinghe, N. Dietary cancer and prevention using antimutagens. Toxicology 2004, 198, 147–159. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Pisano, M.; Pagnan, G.; Loi, M.; Mura, M.E.; Tilocca, M.G.; Palmieri, G.; Fabbri, D.; Dettori, M.A.; Delogu, G.; Ponzoni, M.; et al. Antiproliferative and pro-apoptotic activity of eugenol-related biphenyls on malignant melanoma cells. Mol. Cancer 2007, 6, 8–20. [Google Scholar] [CrossRef]
- Ogata, M.; Hoshi, M.; Urano, S.; Endo, T. Antioxidant activity of eugenol and related monomeric and dimeric compounds. Chem. Pharm. Bull. 2000, 48, 1467–1469. [Google Scholar] [CrossRef]
- Benencia, F.; Courreges, M.C. In vitro and in vivo activity of eugenol on human herpesvirus. Phytother. Res. 2000, 14, 495–500. [Google Scholar] [CrossRef]
- Chogo, J.B.; Crank, G. Chemical composition and biological activity of the tanzanian plant Ocimum suave. J. Nat. Prod. 1981, 42, 308–311. [Google Scholar] [CrossRef]
- Asha, M.K.; Prashanth, D.; Murali, B.; Padmaja, R.; Amit, A. Anthelmintic activity of essential oil of Ocimum sanctum and eugenol. Fitoterapia 2001, 72, 669–670. [Google Scholar] [CrossRef]
- Miyazawa, M.; Hisama, M. Suppression of chemical mutagen-induced SOS response by alkylphenols from clove (Syzygium aromaticum) in the Salmonella typhimurium TA1535/pSK1002. J. Agric. Food Chem. 2001, 49, 4019–4025. [Google Scholar] [CrossRef]
- Han, E.H.; Hwang, Y.P.; Jeong, T.C.; Lee, S.S.; Shin, J.G.; Jeong, H.G. Eugenol inhibit 7,12-dimethylbenz[a]anthracene-induced genotoxicity in MCF-7 cells: Bifunctional effects on CYP1 and NAD (P) H:quinone oxidoreductase. FEBS Lett. 2007, 581, 749–756. [Google Scholar] [CrossRef]
- Abraham, S.K. Anti-genotoxicity of trans-anethole and eugenol in mice. Food Chem. Toxicol. 2001, 39, 493–498. [Google Scholar] [CrossRef]
- World Health Organization (WHO), Twenty-sixth report of the Joint FAO/WHO Expert Committee on Food Additives, WHO Technical Report Series, No. 683. In Evaluation of Certain Food Additives and Contaminants; Geneva, Switzerland, 1982.
- Lee, K.G.; Shibamoto, T. Antioxidant property of aroma extract isolated from clove buds Syzygium aromaticum (L.) Merr. et Perry. Food Chem. 2001, 74, 443–448. [Google Scholar]
- Usta, J.; Copti, R. Effect of cinnamon, clove and some of their constituents on the Na+-K+-ATPase activity and alanine absorption in the rat jejunum. Food Chem. Toxicol. 2000, 38, 755–762. [Google Scholar] [CrossRef]
- Gursale, A.A.; Sane, R.T.; Menon, S.; Patel, P.H. Quantitative determination of eugenol from Cinnamomum tamala nees and eberm. Leaf powder and polyherbal formulation using reverse phase liquid chromatography. Chromatographia 2005, 61, 443–446. [Google Scholar] [CrossRef]
- Stamford, I.F.; Tavares, I.A.; Jacobs, S.; Capasso, F.; Mascolo, N.; Autore, G.; Romano, V. The biological activity of eugenol, a major constituent of nutmeg (Myristica fragrans): Studies on prostaglandins, the intestine and other tissues. Phytother. Res. 1988, 2, 124–130. [Google Scholar] [CrossRef]
- Kim, G.C.; Choi, D.S.; Lim, J.S.; Jeong, H.C.; Kim, I.R.; Lee, M.H.; Park, B.S. Caspases-dependent apoptosis in human melanoma cell by eugenol. Korean J. Anat. 2006, 39, 245–253. [Google Scholar]
- Ghosh, R.; Nadiminty, N.; Fitzpatrick, J.E.; Alworth, W.L.; Slaga, T.J.; Kumar, A.P. Eugenol causes melanoma growth suppression through inhibition of E2F1 transcriptional activity. J. Biol. Chem. 2005, 280, 5812–5819. [Google Scholar]
- Pal, D.; Banerjee, S.; Mukherjee, S.; Roy, A.; Panda, C.K.; Das, S. Eugenol restricts DMBA croton oil induced skin carcinogenesis in mice: Downregulation of c-Myc and H-ras, and activation of p53 dependent apoptotic pathway. J. Dermatol. Sci. 2010, 59, 31–39. [Google Scholar] [CrossRef]
- Kaur, G.; Athar, M.; Alam, M.S. Eugenol precludes cutaneous chemical carcinogenesis in mouse by preventing oxidative stress and inflammation and by inducing apoptosis. Mol. Carcinog. 2010, 49, 290–301. [Google Scholar]
- Shin, S.H.; Park, J.H.; Kim, G.C. The mechanism of apoptosis induced by eugenol in human osteosarcoma cells. J. Korean Oral Maxillofac. Surg. 2007, 33, 20–27. [Google Scholar]
- Yoo, C.B.; Han, K.T.; Cho, K.S.; Ha, J.; Park, H.J.; Nam, J.H.; Kil, U.H.; Lee, K.T. Eugenol isolated from the essential oil of Eugenia caryophyllata induces a reactive oxygen species-mediated apoptosis in HL-60 human promyelocytic leukemia cells. Cancer Lett. 2005, 225, 41–52. [Google Scholar] [CrossRef]
- Okada, N.; Hirata, A.; Murakami, Y.; Shoji, M.; Sakagami, H.; Fujisawa, S. Induction of cytotoxicity and apoptosis and inhibition of cyclooxygenase-2 gene expression by eugenol-related compounds. Anticancer Res. 2005, 25, 3263–3269. [Google Scholar]
- Manikandan, P.; Vinothini, G.; Priyadarsini, R.V.; Prathiba, D.; Nagini, S. Eugenol inhibits cell proliferation via NF-κB suppression in a rat model of gastric carcinogenesis induced by MNNG. Invest. New Drugs 2011, 29, 110–117. [Google Scholar] [CrossRef]
- Manikandan, P.; Murugan, R.S.; Priyadarsini, R.V.; Vinothini, G.; Nagini, S. Eugenol induces apoptosis and inhibits invasion and angiogenesis in a rat model of gastric carcinogenesis induced by MNNG. Life Sci. 2010, 86, 936–941. [Google Scholar] [CrossRef]
- Park, B.S.; Song, Y.S.; Yee, S.B.; Lee, B.G.; Seo, S.Y.; Park, Y.C.; Kim, J.M.; Kim, H.M.; Yoo, Y.H. Phospho-ser 15-p53 translocates into mitochondria and interacts with Bcl-2 and Bcl-xL in eugenol-induced apoptosis. Apoptosis 2005, 10, 193–200. [Google Scholar] [CrossRef]
- Rothschild, A.M. Mechanisms of histamine release by compound 48/80. Br. J. Pharmacol. 1970, 38, 253–262. [Google Scholar] [CrossRef]
- Ghosh, R.; Ganapathy, M.; Alworth, W.L.; Chan, D.C.; Kumar, A.P. Combination of 2-methoxyestradiol (2-ME2) and eugenol for apoptosis induction synergistically in androgen independent prostate cancer cells. J. Steroid Biochem. Mol. Biol. 2009, 113, 25–35. [Google Scholar] [CrossRef]
- Atsumi, T.; Fujisawa, S.; Satoh, K.; Sakagami, H.; Iwakura, I.; Ueha, T.; Sugita, Y.; Yokoe, I. Cytotoxicity and radical intensity of eugenol, isoeugenol or related dimers. Anticancer Res. 2000, 20, 2519–2524. [Google Scholar]
- Carrasco, A.; Espinoza, C.; Cardile, V.; Gallardo, C.; Cardona, W.; Lombardo, L.; Catalán, M.; Cuellar, F.; Russo, A. Eugenol and its synthetic analogues inhibit cell growth of human cancer cells. J. Braz. Chem. Soc. 2008, 19, 543–548. [Google Scholar] [CrossRef]
- Jaganathan, S.K.; Mondhe, D.; Wani, Z.A.; Pal, H.C.; Mandal, M. Effect of honey and eugenol on ehrlich ascites and solid carcinoma. J. Biomed. Biotechnol. 2010, 2010, 989163–1. [Google Scholar]
- Jaganathan, S.K.; Mazumdar, A.; Mondhe, D.; Mandal, M. Apoptotic effect of eugenol in human colon cancer cell lines. Cell Biol. Int. 2011, 35, 607–615. [Google Scholar] [CrossRef]
- Sukumaran, K.; Unnikrishnan, M.C.; Kuttan, R. Inhibition of tumour promotion in mice by eugenol. Indian J. Physiol. Pharmacol. 1994, 38, 306–308. [Google Scholar]
- Hussain, A.; Priyani, A.; Sadrieh, L.; Brahmbhatt, K.; Ahmed, M.; Sharma, C. Concurrent sulforaphane and eugenol induces differential effects on human cervical cancer cells. Integr. Cancer Ther. 2011. [Google Scholar] [CrossRef]
- Hussain, A.; Brahmbhatt, K.; Priyani, A.; Ahmed, M.; Rizvi, T.A.; Sharma, C. Eugenol enhances the chemotherapeutic potential of gemcitabine and induces anticarcinogenic and anti-inflammatory activity in human cervical cancer cells. Cancer Biother. Radiopharm. 2011, 26, 519–527. [Google Scholar] [CrossRef]
- Vidhya, N.; Devaraj, S.N. Induction of apoptosis by eugenol in human breast cancer cells. Indian J. Exp. Biol. 2011, 49, 871–878. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jaganathan, S.K.; Supriyanto, E. Antiproliferative and Molecular Mechanism of Eugenol-Induced Apoptosis in Cancer Cells. Molecules 2012, 17, 6290-6304. https://doi.org/10.3390/molecules17066290
Jaganathan SK, Supriyanto E. Antiproliferative and Molecular Mechanism of Eugenol-Induced Apoptosis in Cancer Cells. Molecules. 2012; 17(6):6290-6304. https://doi.org/10.3390/molecules17066290
Chicago/Turabian StyleJaganathan, Saravana Kumar, and Eko Supriyanto. 2012. "Antiproliferative and Molecular Mechanism of Eugenol-Induced Apoptosis in Cancer Cells" Molecules 17, no. 6: 6290-6304. https://doi.org/10.3390/molecules17066290