The Health Promoting Bioactivities of Lactuca sativa can be Enhanced by Genetic Modulation of Plant Secondary Metabolites

 and
and 
Abstract
:1. Introduction
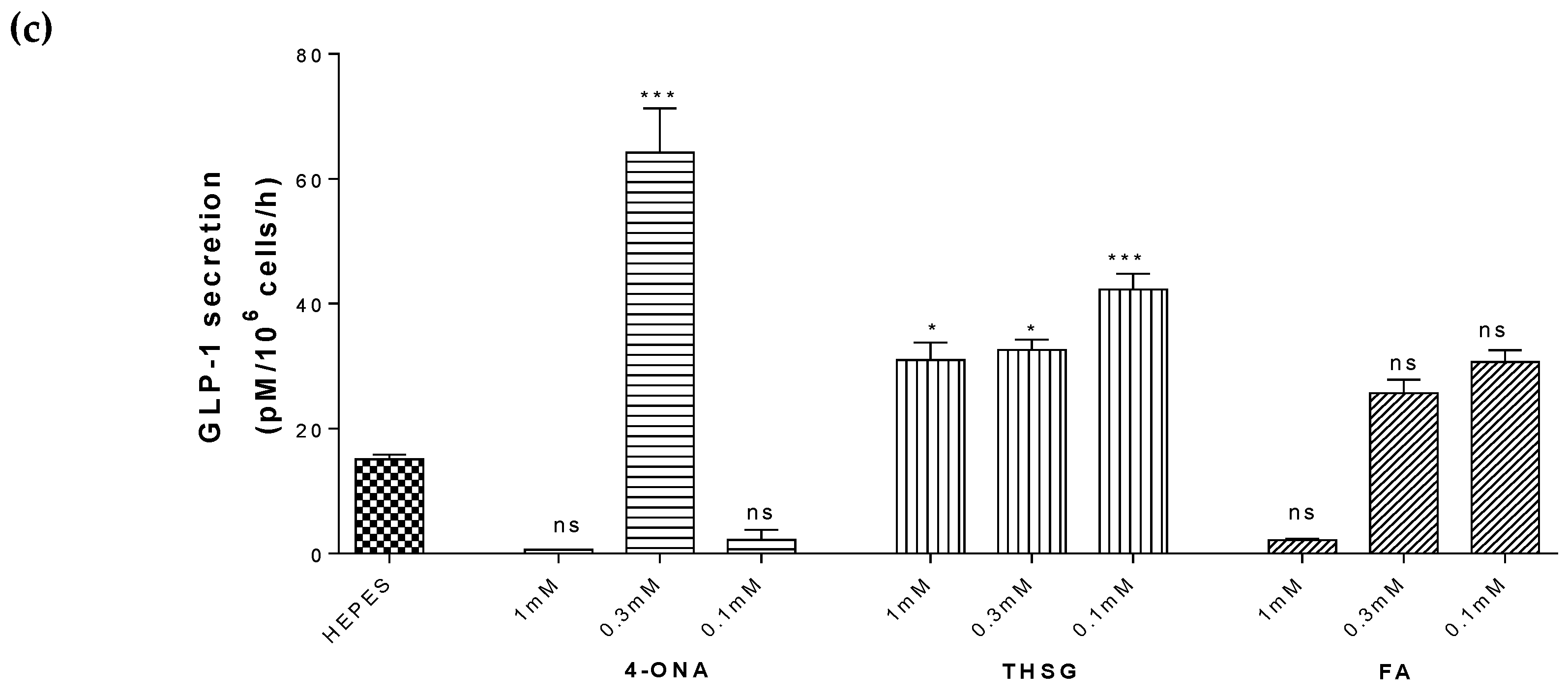
2. Results
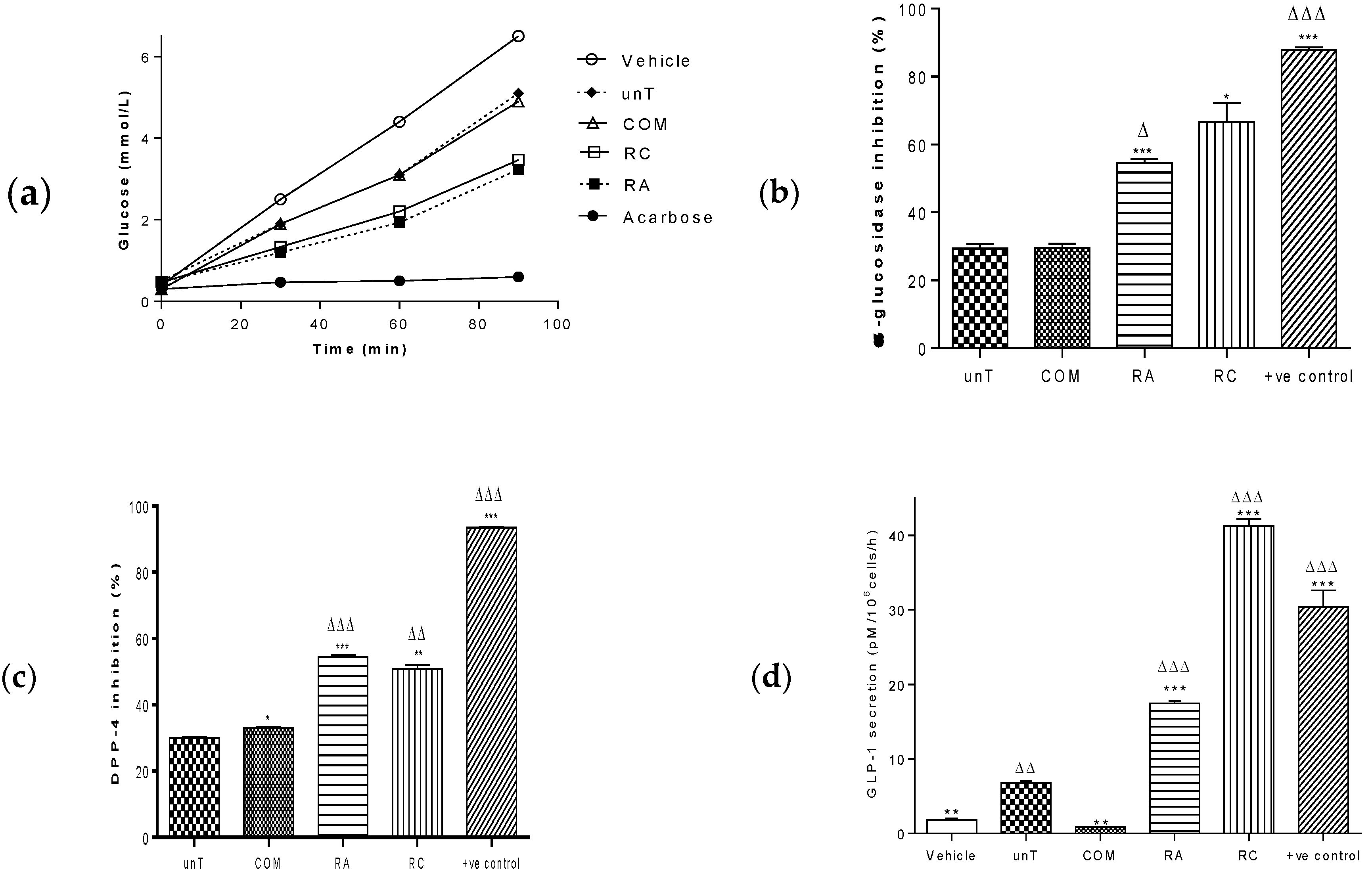
2.1. Agrobacterium Rhizogenes rol Genes Enhance the Antidiabetic Bioactivity of Lactuca sativa
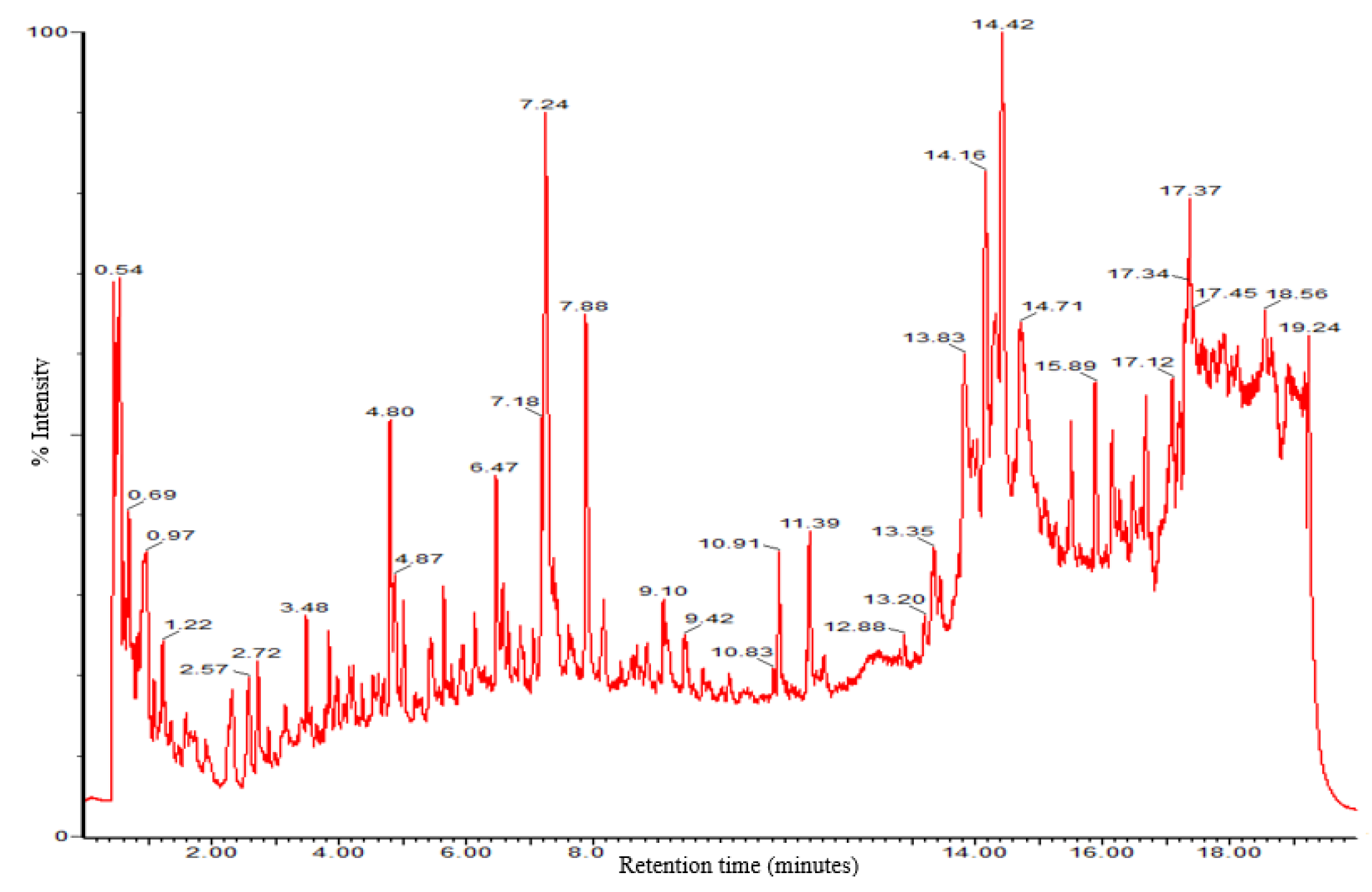
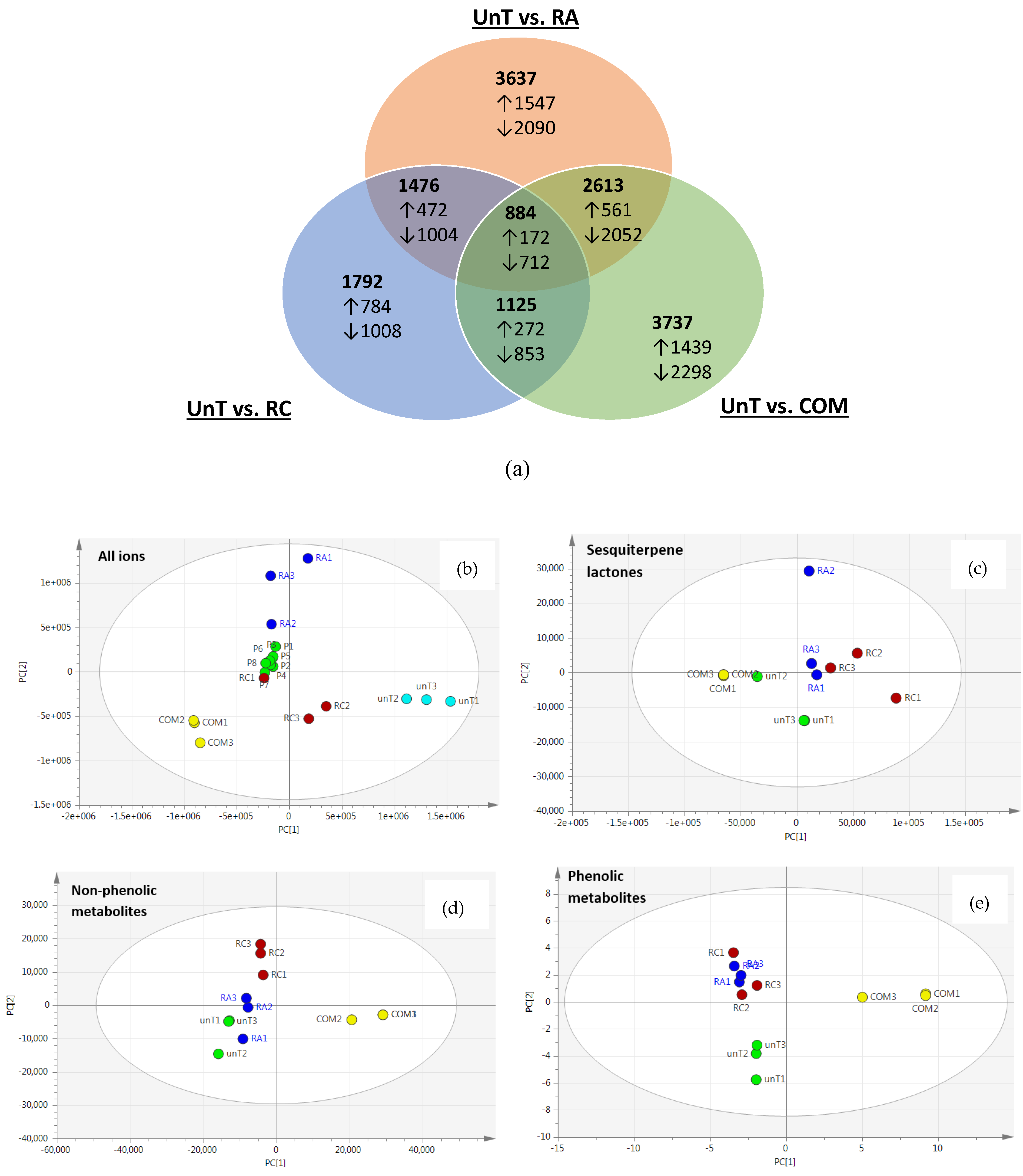
2.2. Agrobacterium rhizogenes rol Genes Induce Widespread Changes in the L. sativa Metabolome
2.3. Sesquiterpene Lactones and Related Derivatives/Conjugates
2.4. Non-Phenolic Metabolites
2.5. Phenolic Metabolites
2.6. Unidentified Metabolites
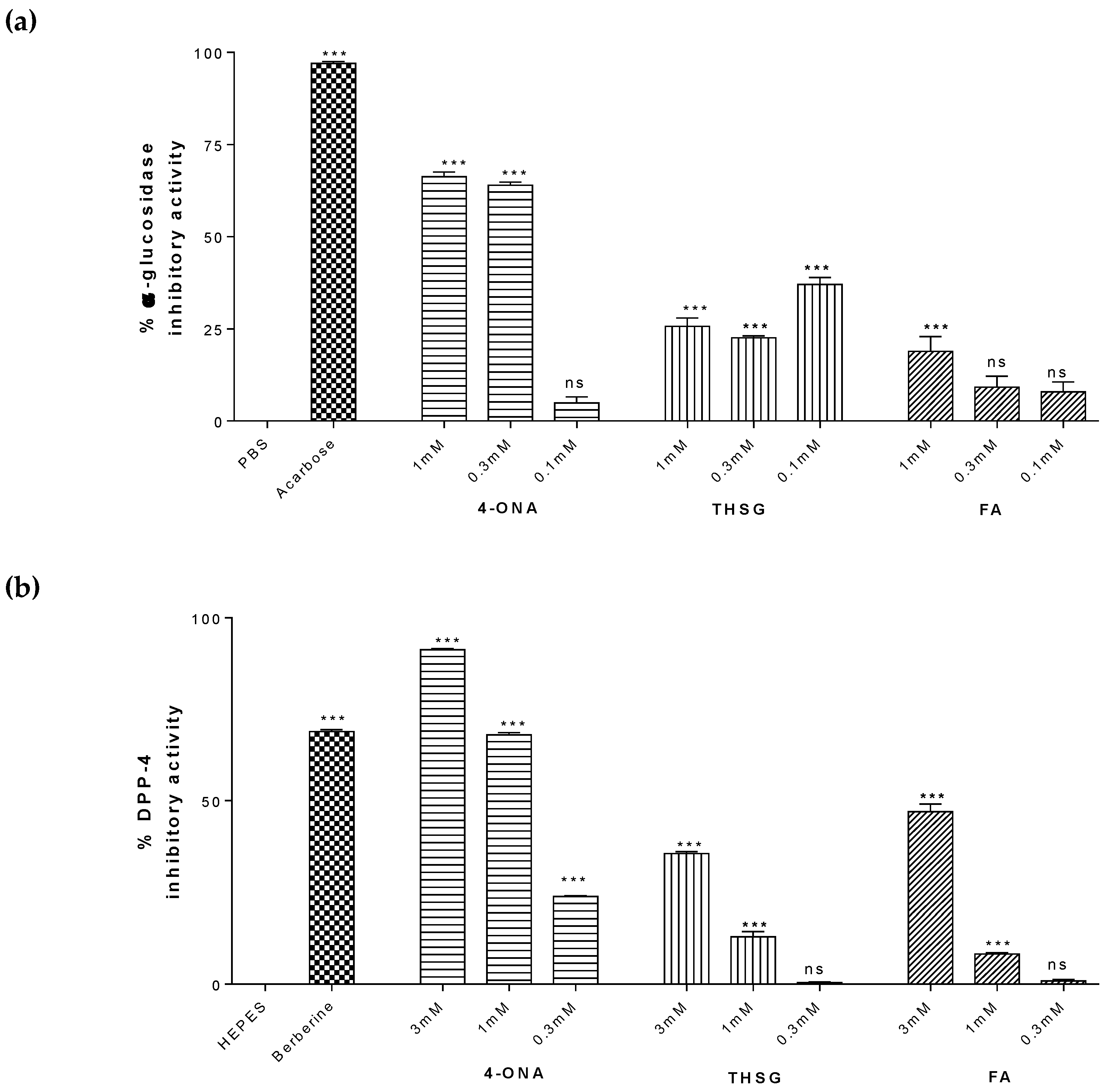
2.7. Antidiabetic Effects of Selected Compounds
3. Discussion
4. Materials and Methods
4.1. Plants
4.2. Extraction of Plant Material
4.3. UPLC-QTof-MS Analysis of Lactuca Sativa
4.4. Data Analysis
4.5. Plant Metabolite Identification
4.6. Measurement of Alpha-Glucosidase Inhibition
4.7. Measurement of DPP-4 Inhibitory Activity
4.8. GLP-1 Secretory Responses in pGIP/Neo STC-1 Cells
4.9. Data Analysis of Bioactivity Studies
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Spena, A.; Schmülling, T.; Koncz, C.; Schell, J. Independent and synergistic activity of rol A, B and C loci in stimulating abnormal growth in plants. Embo J. 1987, 6, 3891–3899. [Google Scholar] [CrossRef]
- Cardarelli, M.; Mariotti, D.; Pomponi, M.; Spano, L.; Capone, I.; Costantino, P. Agrobacterium rhizogenes T-DNA genes capable of inducing hairy root phenotype. Mol. Gen. Genet. 1987, 209, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Bulgakov, V.P. Functions of rol genes in plant secondary metabolism. Biotechnol. Adv. 2008, 26, 318–324. [Google Scholar] [CrossRef]
- Bulgakov, V.; Tchernoded, G.; Mischenko, N.; Shkryl, Y.N.; Glazunov, V.; Fedoreyev, S.; Zhuravlev, Y.N. Effects of Ca2+ channel blockers and protein kinase/phosphatase inhibitors on growth and anthraquinone production in Rubia cordifolia callus cultures transformed by the rolB and rolC genes. Planta 2003, 217, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Bulgakov, V.P.; Khodakovskaya, M.V.; Labetskaya, N.V.; Chernoded, G.K.; Zhuravlev, Y.N. The impact of plant rolC oncogene on ginsenoside production by ginseng hairy root cultures. Phytochemistry 1998, 49, 1929–1934. [Google Scholar] [CrossRef]
- Bonhomme, V.; Laurain-Mattar, D.; Fliniaux, M. Effects of the rol C gene on hairy root: Induction development and tropane alkaloid production by Atropa belladonna. J. Nat. Prod. 2000, 63, 1249–1252. [Google Scholar] [CrossRef]
- Palazón, J.; Cusidó, R.; Roig, C.; Pinol, M. Expression of the rolC gene and nicotine production in transgenic roots and their regenerated plants. Plant Cell Rep. 1998, 17, 384–390. [Google Scholar]
- Sevón, N.; Oksman-Caldentey, K.-M. Agrobacterium rhizogenes-mediated transformation: Root cultures as a source of alkaloids. Planta Med. 2002, 68, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Graefe, E.; Veit, M. Urinary metabolites of flavonoids and hydroxycinnamic acids in humans after application of a crude extract from Equisetum arvense. Phytomedicine 1999, 6, 239–246. [Google Scholar] [CrossRef]
- Olthof, M.R.; Hollman, P.C.; Katan, M.B. Chlorogenic acid and caffeic acid are absorbed in humans. J. Nutr. 2001, 131, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Abu-Reidah, I.; Contreras, M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Reversed-phase ultra-high-performance liquid chromatography coupled to electrospray ionization-quadrupole-time-of-flight mass spectrometry as a powerful tool for metabolic profiling of vegetables: Lactuca sativa as an example of its application. J. Chromatogr. A 2013, 1313, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.L.; Clifford, M.N.; Morgan, L.M. Coffee acutely modifies gastrointestinal hormone secretion and glucose tolerance in humans: Glycemic effects of chlorogenic acid and caffeine. Am. J. Clin. Nutr. 2003, 78, 728–733. [Google Scholar] [CrossRef]
- Dao, T.-M.A.; Waget, A.; Klopp, P.; Serino, M.; Vachoux, C.; Pechere, L.; Drucker, D.J.; Champion, S.; Barthélemy, S.; Barra, Y.; et al. Resveratrol increases glucose induced GLP-1 secretion in mice: A mechanism which contributes to the glycemic control. PLoS ONE 2011, 6, e20700. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, C.R. Alkylperoxyl radical scavenging activity of red leaf lettuce (Lactuca sativa L.) phenolics. J. Agric. Food Chem. 2003, 51, 4589–4595. [Google Scholar] [CrossRef]
- Llorach, R.; Tomás-Barberán, F.A.; Ferreres, F. Lettuce and chicory byproducts as a source of antioxidant phenolic extracts. J. Agric. Food Chem. 2004, 52, 5109–5116. [Google Scholar] [CrossRef]
- Nicolle, C.; Cardinault, N.; Gueux, E.; Jaffrelo, L.; Rock, E.; Mazur, A.; Amouroux, P.; Rémésy, C. Health effect of vegetable-based diet: Lettuce consumption improves cholesterol metabolism and antioxidant status in the rat. Clin. Nutr. 2004, 23, 605–614. [Google Scholar] [CrossRef]
- Sayyah, M.; Hadidi, N.; Kamalinejad, M. Analgesic and anti-inflammatory activity of Lactuca sativa seed extract in rats. J. Ethnopharmacol. 2004, 92, 325–329. [Google Scholar] [CrossRef]
- Ismail, H.; Mirza, B. Evaluation of analgesic, anti-inflammatory, anti-depressant and anti-coagulant properties of Lactuca sativa (CV. Grand Rapids) plant tissues and cell suspension in rats. BMC Complement. Altern. Med. 2015, 15, 199. [Google Scholar] [CrossRef]
- Derosa, G.; Maffioli, P. α-Glucosidase inhibitors and their use in clinical practice. Arch. Med. Sci. 2012, 8, 899. [Google Scholar] [CrossRef]
- Green, B.D.; Flatt, P.R.; Bailey, C.J. Inhibition of dipeptidyl peptidase IV activity as a therapy of type 2 diabetes. Expert Opin. Emerg. Drugs 2006, 11, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-glucosidase and α-amylase by flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef]
- Jafri, L.; Saleem, S.; Calderwood, D.; Gillespie, A.; Mirza, B.; Green, B.D. Naturally-occurring TGR5 agonists modulating glucagon-like peptide-1 biosynthesis and secretion. Peptides 2016, 78, 51–58. [Google Scholar] [CrossRef]
- Saleem, S.; Jafri, L.; ul Haq, I.; Chang, L.C.; Calderwood, D.; Green, B.D.; Mirza, B. Plants Fagonia cretica L. and Hedera nepalensis K. Koch contain natural compounds with potent dipeptidyl peptidase-4 (DPP-4) inhibitory activity. J. Ethnopharmacol. 2014, 156, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, A.L.; Calderwood, D.; Hobson, L.; Green, B.D. Whey proteins have beneficial effects on intestinal enteroendocrine cells stimulating cell growth and increasing the production and secretion of incretin hormones. Food Chem. 2015, 189, 120–128. [Google Scholar] [CrossRef]
- Gillespie, A.L.; Green, B.D. The bioactive effects of casein proteins on enteroendocrine cell health, proliferation and incretin hormone secretion. Food Chem. 2016, 211, 148–159. [Google Scholar] [CrossRef]
- Shkryl, Y.N.; Veremeichik, G.N.; Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Fedoreyev, S.A.; Zhuravlev, Y.N. Individual and combined effects of the rolA, B, and C genes on anthraquinone production in Rubia cordifolia transformed calli. Biotechnol. Bioeng. 2008, 100, 118–125. [Google Scholar] [CrossRef]
- Kiselev, K.; Dubrovina, A.; Veselova, M.; Bulgakov, V.; Fedoreyev, S.; Zhuravlev, Y.N. The rol B gene-induced overproduction of resveratrol in Vitis amurensis transformed cells. J. Biotechnol. 2007, 128, 681–692. [Google Scholar] [CrossRef]
- Panwar, H.; Calderwood, D.; Grant, I.R.; Grover, S.; Green, B.D. Lactobacillus strains isolated from infant faeces possess potent inhibitory activity against intestinal alpha-and beta-glucosidases suggesting anti-diabetic potential. Eur. J. Nutr. 2014, 53, 1465–1474. [Google Scholar] [CrossRef]
- Chang, M.-J.; Xiao, J.-H.; Wang, Y.; Yan, Y.-L.; Yang, J.; Wang, J.-L. 2,3,5,4′-Tetrahydroxystilbene-2-O-beta-D-glucoside improves gastrointestinal motility disorders in STZ-induced diabetic mice. PLoS ONE 2012, 7, e50291. [Google Scholar] [CrossRef] [PubMed]
- Balasubashini, M.; Rukkumani, R.; Viswanathan, P.; Menon, V.P. Ferulic acid alleviates lipid peroxidation in diabetic rats. Phytother. Res. 2004, 18, 310–314. [Google Scholar] [CrossRef]
- Ismail, H.; Dilshad, E.; Waheed, M.T.; Mirza, B. Transformation of lettuce with rol ABC genes: Extracts show enhanced antioxidant, analgesic, anti-inflammatory, antidepressant, and anticoagulant activities in rats. Appl. Biochem. Biotechnol. 2017, 181, 1179–1198. [Google Scholar] [CrossRef]
- Ismail, H.; Dilshad, E.; Waheed, M.T.; Sajid, M.; Kayani, W.K.; Mirza, B. Transformation of Lactuca sativa L. with rol C gene results in increased antioxidant potential and enhanced analgesic, anti-inflammatory and antidepressant activities in vivo. 3 Biotech 2016, 6, 215. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Graham, S.F.; Chevallier, O.P.; Roberts, D.; Holscher, C.; Elliott, C.T.; Green, B.D. Investigation of the human brain metabolome to identify potential markers for early diagnosis and therapeutic targets of Alzheimer’s disease. Anal. Chem. 2013, 85, 1803–1811. [Google Scholar] [CrossRef]
- Fujiwara, K.; Tsuru, D. New chromogenic and fluorogenic substrates for pyrrolidonyl peptidase. J. Biochem. 1978, 83, 1145–1149. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | Rol ABC (RA) | Rol C (RC) | Commercial (COM) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ↑/↓ | % Change ± SEM | p Value | ↑/↓ | % Change ± SEM | p Value | ↑/↓ | % Change ± SEM | P Value | ||
| 1 | Lactucopicrin | ↑ | 17 ± 10.3 | 0.272 | ↑ | 85 ± 31.6 | 0.048 * | ↑ | 90 ± 8.9 | 0.009 ** |
| 2 | Lactucopicrin-15-oxalate | ↓ | 79 ± 14.1 | 0.020 * | ↑ | 29 ± 18.3 | 0.183 | ↓ | 6 ± 0.5 | 0.403 |
| 3 | 11β,13-dihydrolactucopicrin | ↑ | 132 ± 6.3 | 0.002 ** | ↑ | 257 ± 4.4 | 0.006 ** | ↑ | 127 ± 4.9 | 0.003 ** |
| 4 | Lactucin | - | Unchanged | N/A | ↑ | unchanged | N/A | ↑ | 667 ± 145.3 | 0.005 ** |
| 5 | Lactucin-sulfate | ↑ | 25 ± 5.6 | 0.177 | ↑ | 95 ± 26.7 | 0.027 * | ↑ | 90 ± 11.1 | 0.009 ** |
| 6 | 15-deoxylactucin-8-sulfate | ↓ | 93 ± 3.1 | 0.008 ** | ↓ | 95 ± 1.5 | 0.007 ** | ↓ | 56 ± 1.0 | 0.035 * |
| 7 | 15-deoxylactucin-8-sulfate-Gln | ↑ | 535 ± 98.9 | 0.005 ** | ↑ | 511 ± 68.8 | 0.001 ** | ↓ | 89 ± 5.3 | 0.009 * |
| 8 | 15-deoxylactucin-8-sulfate-Pro | ↓ | 96 ± 1.4 | 0.007 ** | ↓ | 97 ± 0.4 | 0.007 ** | ↑ | 50 ± 1.8 | 0.048 ** |
| 9 | 15-deoxylactucin-8-sulfate-Val | ↑ | 1103 ± 99.1 | 0.061 | ↑ | 1028 ±164.9 | 0.002 ** | ↓ | 20 ± 3.1 | 0.214 |
| 10 | 8-deacetylmatricarin-8-sulfate | ↑ | 28 ± 15.8 | 0.182 | ↑ | 128 ± 43.8 | 0.030 * | ↑ | 207 ± 13.1 | 0.001 ** |
| 11 | Cichorioside B-sulfate | ↑ | 83 ± 10.3 | 0.014 * | ↑ | 149 ± 26.9 | 0.006 * | ↑ | 52 ± 8.6 | 0.047 * |
| 12 | 1-β-(4-hydroxyphenylacetyl)-15-O-β-d-glucopyranosyl-5α, 6βH-eudesma-3-en-12, 6α-olide | ↑ | 327 ± 48.2 | 0.029 * | ↓ | 29 ± 23.6 | 0.220 | ↓ | 100 ± 9.8 | 0.007 ** |
| No. | Compound | Rol ABC (RA) | Rol C (RC) | Commercial (COM) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ↑/↓ | % Change ± SEM | p Value | ↑/↓ | % Change ± SEM | p Value | ↑/↓ | % Change ± SEM | p Value | ||
| 1 | Leucine/Isoleucine | ↑ | 168 ± 19.5 | 0.003 ** | ↑ | 457 ± 67.2 | 0.002 ** | ↑ | 794 ± 7.9 | 0.001 ** |
| 2 | Glycine-Leucine | ↑ | 57 ± 36.6 | 0.126 | ↑ | 69 ± 11.9 | 0.024 * | ↑ | 289 ± 5.4 | 0.001 ** |
| 3 | Glycine-Leu/Ileu-hexose | ↑ | 2192 ± 659.5 | 0.015 * | ↑ | 2039 ± 960 | 0.050 * | ↑ | 2292 ± 38.7 | 0.001 ** |
| 4 | Tryptophan | ↓ | 22 ± 18.4 | 0.241 | ↑ | 107 ± 65.1 | 0.096 | ↑ | 494 ± 9.8 | 0.001 ** |
| 5 | Tryptophan-hexose | ↑ | 523 ± 193.4 | 0.031 * | ↑ | 516 ± 277.1 | 0.071 | ↑ | 1514 ± 664.6 | 0.043 * |
| 6 | Tyrosine | ↑ | 160 ± 32.9 | 0.007 ** | ↑ | 401 ± 76.0 | 0.004 ** | ↑ | 282 ± 2.9 | 0.001 ** |
| 7 | Tyrosine-hexose | ↑ | 688 ± 164.9 | 0.007 ** | ↑ | 1295 ± 41.9 | 0.001 ** | ↑ | 2897 ± 78.9 | 0.001 ** |
| 8 | Phenylalanine | ↑ | 168 ± 35.5 | 0.008 ** | ↑ | 540 ± 148.3 | 0.011 * | ↑ | 2968 ± 13.5 | 0.001 ** |
| 9 | Phenylalanine-hexose | ↑ | 95 ± 16.4 | 0.012 * | ↑ | 206 ± 1.8 | 0.001 ** | ↑ | 2567 ± 20.3 | 0.001 ** |
| 10 | Malic acid | ↓ | 100 ± 0.9 | 0.005 ** | ↓ | 100 ± 0.9 | 0.005 ** | ↓ | 51 ± 3.0 | 0.038 * |
| 11 | Citramalic acid | ↓ | 38 ± 5.8 | 0.080 | ↓ | 29 ± 4.7 | 0.126 | ↑ | 59 ± 20.3 | 0.059 |
| 12 | Pyroglutamic acid | ↑ | 53 ± 10.3 | 0.047 * | ↑ | 154 ± 22.8 | 0.004 ** | ↑ | 51 ± 2.2 | 0.042 * |
| 13 | Pyroglutamic acid-Leucine/Isoleucine | ↑ | 152 ± 60.2 | 0.038 * | ↑ | 294 ± 77.8 | 0.011 * | ↑ | 87 ± 0.8 | 0.008 ** |
| 14 | Uridine | ↑ | 108 ± 44.0 | 0.047 * | ↑ | 207 ± 20.5 | 0.001 ** | ↑ | 126 ± 3.2 | 0.002 ** |
| 15 | Adenosine | ↑ | 84 ± 58.1 | 0.124 | ↑ | 148 ± 27.3 | 0.007 ** | ↑ | 224 ± 1.9 | 0.001 ** |
| 16 | Guanosine | ↑ | 200 ± 67.7 | 0.024 * | ↑ | 255 ± 54.4 | 0.006 ** | ↑ | 113 ± 0.2 | 0.003 ** |
| 17 | Isopropylmalic acid | ↑ | 56 ± 7.8 | 0.038 * | ↑ | 43 ± 45.7 | 0.223 | ↑ | 537 ± 19.3 | 0.001 ** |
| 18 | 1,2,3,4-Tetrahydro-beta-carboline-3-carboxylic acid | ↓ | 50 ± 9.5 | 0.071 | ↑ | 125 ± 96.4 | 0.139 | ↑ | 1698 ± 13.2 | 0.001 ** |
| 19 | 1,2,3,4-Tetrahydro-b-carboline-1,3-dicarboxylic acid | ↓ | 38 ± 6.6 | 0.081 | ↑ | 147 ± 90.3 | 0.094 | ↑ | 1567 ± 27.7 | 0.001 ** |
| 20 | Azelaic acid | ↓ | 66 ± 4.8 | 0.022 * | ↑ | 52 ± 52.6 | 0.208 | ↓ | 26 ± 1.4 | 0.155 |
| 21 | Quinic acid | ↓ | 61 ± 3.5 | 0.026 * | ↓ | 46 ± 10.3 | 0.065 | ↓ | 72 ± 0.4 | 0.016 * |
| 22 | Citric Acid | ↑ | 169 ± 26.6 | 0.004 ** | ↑ | 508 ± 137.3 | 0.011 * | ↑ | 1137 ± 241.7 | 0.005 ** |
| 23 | Amino oxononanoic acid | ↑ | 1803 ± 388.3 | 0.005 ** | ↑ | 1141 ± 337.1 | 0.015 * | ↑ | 188 ± 41.5 | 0.030 * |
| 24 | Pantothenic acid (vitamin B5) | ↑ | 128 ± 10.0 | 0.003 ** | ↑ | 79 ± 167.3 | 0.332 | ↓ | 100 ± 0.9 | 0.005 ** |
| No. | Compound | Rol ABC (RA) | Rol C (RC) | Commercial (COM) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ↑/↓ | % Change ± SEM | p Value | ↑/↓ | % Change ± SEM | p Value | ↑/↓ | % Change ± SEM | p Value | ||
| 1 | Hydroxybenzoic acid | ↓ | 25 ± 14.9 | 0.170 | ↑ | 71 ± 35.2 | 0.074 | ↓ | 70 ± 4.0 | 0.009 ** |
| 2 | Dihydroxybenzoic acid | ↓ | 59 ± 5.5 | 0.017 * | ↑ | 48 ± 50.8 | 0.213 | ↑ | 223 ± 53.1 | 0.008 ** |
| 3 | 3-Methoxy-4-hydroxyphenyl-1-O-β-d-glucoside | ↑ | 115 ± 47.9 | 0.044 * | ↑ | 153 ± 26.1 | 0.004 ** | ↑ | 28 ± 17.7 | 0.163 |
| 4 | Syringic acid | ↑ | Unchanged | N/A | ↑ | unchanged | N/A | ↑ | unchanged | N/A |
| 5 | Syringic acid hexose | ↓ | 49 ± 1.1 | 0.027 * | ↑ | 35 ± 33.5 | 0.203 | ↓ | 76 ± 3.4 | 0.007 ** |
| 6 | Vanillic Acid | ↑ | Unchanged | N/A | ↑ | unchanged | N/A | ↑ | unchanged | N/A |
| 7 | Vanillic acid glucoside | ↓ | 51 ± 11.5 | 0.037 * | ↓ | 44 ± 8.2 | 0.046 * | ↓ | 10 ± 14.5 | 0.340 |
| 8 | Dihydroxybenzoic acid hexose | ↓ | 77 ± 1.2 | 0.007 ** | ↓ | 25 ± 19.8 | 0.205 | ↑ | 66 ± 23.1 | 0.044 * |
| 9 | Hydroxybenzoyl dihydroxybenzoyl-hexose | ↓ | 35 ± 18.5 | 0.123 | ↑ | 44 ± 35.6 | 0.166 | ↑ | 36 ± 18.8 | 0.121 |
| 10 | Caffeic acid | ↓ | 3 ± 9.9 | 0.439 | ↑ | 131 ± 35.1 | 0.015 * | ↑ | 326 ± 59.1 | 0.003 ** |
| 11 | Dihydrocaffeic acid hexose | ↑ | 65 ± 40.0 | 0.105 | ↑ | 56 ± 17.8 | 0.047 * | ↓ | 9 ± 12.3 | 0.344 |
| 12 | Caffeoyl-hexose | ↑ | 55 ± 15.5 | 0.040 * | ↑ | 62 ± 10.1 | 0.020 * | ↑ | 69 ± 23.2 | 0.039 * |
| 13 | Ferulic acid | ↓ | 34 ± 10.2 | 0.089 | ↑ | 20 ± 33.2 | 0.310 * | ↑ | 38 ± 19.3 | 0.114 |
| 14 | Ferulic acid glucoside | ↓ | 71 ± 3.9 | 0.009 ** | ↓ | 49 ± 10.1 | 0.038 * | ↑ | 8 ± 14.9 | 0.376 |
| 15 | Ferulic acid methyl ester | ↑ | 5616 ± 231 | 0.036 * | ↑ | 9777 ± 291 | 0.014 * | ↑ | 3033 ± 458 | 0.001 ** |
| 16 | Sinapoyl glucoside | ↑ | 21 ± 11.8 | 0.193 | ↑ | 41 ± 54.9 | 0.259 | ↑ | 230 ± 46.1 | 0.005 ** |
| 17 | p-Coumaroyl glucoside | ↓ | 46 ± 5.6 | 0.036 * | ↓ | 28 ± 6.2 | 0.107 | ↑ | 880 ± 140.3 | 0.002 ** |
| 18 | Caftaric acid | ↓ | 80 ± 8.4 | 0.008 ** | ↓ | 91 ± 2.3 | 0.004 ** | ↑ | 2290 ± 332 | 0.001 ** |
| 19 | Tartaric acid | ↑ | Unchanged | N/A | ↑ | unchanged | N/A | ↑ | unchanged | N/A |
| 20 | Coutaric acid | ↓ | 92 ± 2.4 | 0.004 ** | ↓ | 83 ± 6.4 | 0.006 ** | ↑ | 69 ± 24.0 | 0.042 * |
| 21 | Caffeoyltartaric-p-coumaroyl acid | ↓ | 87 ± 4.6 | 0.005 ** | ↓ | 73 ± 11.8 | 0.014 * | ↑ | 822 ± 134.9 | 0.002 ** |
| 22 | Caffeoylquinic acid | ↓ | 39 ± 5.9 | 0.055 | ↓ | 11 ± 13.2 | 0.322 | ↓ | 78 ± 3.0 | 0.006 ** |
| 23 | Dicaffeoylquinic acid | ↓ | 75 ± 6.2 | 0.009 ** | ↓ | 61 ± 10.4 | 0.021 * | ↓ | 94 ± 0.9 | 0.003 ** |
| 24 | p-Coumaroyl-caffeoylquinic acid | ↓ | 5 ± 17.2 | 0.425 | ↓ | 52 ± 6.7 | 0.027 * | ↓ | 86 ± 2.0 | 0.004 ** |
| 25 | p-Coumaroylquinic acid | ↓ | 75 ± 9.1 | 0.011 * | ↓ | 18 ± 4.5 | 0.196 | ↑ | 3978 ± 572 | 0.001 ** |
| 26 | 4-hydroxyphenylacetyl glucoside | ↑ | 119 ± 27.4 | 0.011 | ↑ | 40 ± 102.7 | 0.360 | ↓ | 65 ± 6.0 | 0.013 * |
| 27 | Di(4-hydroxyphenylacetyl)-hexose | ↑ | 1839 ± 438 | 0.007 ** | ↑ | 1454 ± 114 | 0.136 | ↓ | 86 ± 3.4 | 0.005 ** |
| 28 | Quercetin | ↓ | 26 ± 45.6 | 0.314 | ↓ | 73 ± 18.3 | 0.023 * | ↑ | 3952 ± 672 | 0.002 ** |
| 29 | Quercetin hexose | ↓ | 69 ± 3.2 | 0.011 * | ↓ | 38 ± 27.6 | 0.155 | ↑ | 12,022 ± 173 | 0.001 ** |
| 30 | Quercetin 3-glucuronide | ↓ | 78 ± 12.0 | 0.012 * | ↓ | 82 ± 7.4 | 0.007 ** | ↑ | 44,309 ± 6178 | 0.001 ** |
| 31 | Quercetin malonylglucoside | ↓ | 91 ± 3.2 | 0.004 ** | ↓ | 92 ± 1.2 | 0.004 ** | ↑ | 361 ± 65.3 | 0.003 ** |
| 32 | Quercetin-3-O-(6″-O-crotonyl)-β-glucoside | ↑ | 559 ± 59.9 | 0.001 ** | ↑ | 1509 ±319 | 0.005 ** | ↑ | 470 ± 78.7 | 0.002 ** |
| 33 | Naringenin 7-neohesperidoside | ↓ | 51 ± 7.7 | 0.030 * | ↓ | 27 ± 10.9 | 0.133 | ↓ | 81 ± 2.5 | 0.006 ** |
| 34 | Apigenin 7-O-glucuronide | ↑ | 199 ± 76.5 | 0.032 * | ↑ | 81 ± 57.3 | 0.124 | ↑ | 3740 ± 531 | 0.001 ** |
| 35 | Syringaresinol-β-d-glucoside | ↓ | 43 ± 1.4 | 0.039 * | ↓ | 7 ± 10.9 | 0.382 | ↓ | 55 ± 6.5 | 0.023 * |
| 36 | Syringaresinol malonylhexose | ↓ | 95 ± 1.1 | 0.003 ** | ↓ | 97 ± 0.4 | 0.003 * | ↓ | 100 ± 0.0 | 0.003 ** |
| 37 | Alangilignoside C | ↑ | 224 ± 22.3 | 0.001 ** | ↑ | 372 ± 110.2 | 0.015 * | ↓ | 99 ± 0.2 | 0.003 ** |
| 38 | Coniferoside | ↑ | 111 ± 64.1 | 0.085 * | ↑ | 357 ± 154.5 | 0.042 * | ↑ | 272 ± 51.5 | 0.004 ** |
| 39 | 2,3,5,4β-Tetrahydroxystilbene2-O-β-D-glucoside | ↑ | 48,008 ± 365 | 0.001 ** | ↑ | 40,272 ± 1204 | 0.014 * | ↑ | 11,944 ± 207 | 0.002 ** |
| 40 | Eugenol malonylglucoside | ↑ | 171 ± 78.7 | 0.051 | ↑ | 387 ± 88.6 | 0.006 ** | ↓ | 92 ± 1.6 | 0.004 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, H.; Gillespie, A.L.; Calderwood, D.; Iqbal, H.; Gallagher, C.; Chevallier, O.P.; Elliott, C.T.; Pan, X.; Mirza, B.; Green, B.D. The Health Promoting Bioactivities of Lactuca sativa can be Enhanced by Genetic Modulation of Plant Secondary Metabolites. Metabolites 2019, 9, 97. https://doi.org/10.3390/metabo9050097
Ismail H, Gillespie AL, Calderwood D, Iqbal H, Gallagher C, Chevallier OP, Elliott CT, Pan X, Mirza B, Green BD. The Health Promoting Bioactivities of Lactuca sativa can be Enhanced by Genetic Modulation of Plant Secondary Metabolites. Metabolites. 2019; 9(5):97. https://doi.org/10.3390/metabo9050097
Chicago/Turabian StyleIsmail, Hammad, Anna L. Gillespie, Danielle Calderwood, Haroon Iqbal, Colene Gallagher, Olivier P. Chevallier, Christopher T. Elliott, Xiaobei Pan, Bushra Mirza, and Brian D. Green. 2019. "The Health Promoting Bioactivities of Lactuca sativa can be Enhanced by Genetic Modulation of Plant Secondary Metabolites" Metabolites 9, no. 5: 97. https://doi.org/10.3390/metabo9050097