The Potential of Indonesian Heterobranchs Found around Bunaken Island for the Production of Bioactive Compounds

 ,
,
Abstract
:1. Introduction
2. Scope of the Review and Methodology Applied
3. Natural Products with Biological Activity from Heterobranchia Families Found near Bunaken
3.1. Biodiversity of Sea Slugs Found in the Bunaken National Park Survey
3.2. Anaspidae (Sea Hares)
Aplysiidae
3.3. Sacoglossa
3.3.1. Caliphyllidae
3.3.2. Oxynoidae
3.3.3. Plakobranchidae
3.4. Cephalaspidea
3.4.1. Aglajidae
3.4.2. Gastropteridae
3.4.3. Haminoeidae
3.5. Pleurobranchomorpha
3.5.1. Pleurobranchidae
3.6. Nudibranchia
3.6.1. Aegiridae
3.6.2. Chromodorididae
Ceratosoma
Chromodoris
Doriprismatica and Glossodoris
Goniobranchus
Hypselodoris
3.6.3. Cladobranchia (Families Arminidae, Dotidae, Eubranchidae, Facelinidae, Flabellinidae Proctonotidae)
3.6.4. Dendrodorididae
3.6.5. Discodorididae
3.6.6. Hexabranchidae
3.6.7. Phyllidiidae
3.6.8. Polyceridae
4. Discussion and Conclusions
Acknowledgments
Conflicts of Interest
References
- Bogdanov, A.; Kehraus, S.; Bleidissel, S.; Preisfeld, G.; Schillo, D.; Piel, J.; Brachmann, A.O.; Wägele, H.; König, G.M. Defense in the Aeolidoidean Genus Phyllodesmium (Gastropoda). J. Chem. Ecol. 2014, 40, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Vardaro, R.R.; Petrocellis, L.D.; Villani, G.; Minei, R.; Cimino, G. Cyercenes, novel pyrones from the ascoglossan molluscCyerce cristallina. Tissue Distribution, Biosynthesis and Possible Involvement in Defense and Regenerative Processes. Experientia 1991, 47, 1221–1227. [Google Scholar] [CrossRef]
- Gavagnin, M.; Marin, A.; Mollo, E.; Crispino, A.; Villani, G.; Cimino, G. Secondary Metabolites from Mediterranean Elysioidea: Origin and Biological Role. Comp. Biochem. Physiol. Part B Comp. Biochem. 1994, 108, 107–115. [Google Scholar] [CrossRef]
- Cutignano, A.; Tramice, A.; De Caro, S.; Villani, G.; Cimino, G.; Fontana, A. Biogenesis of 3-Alkylpyridine Alkaloids in the Marine Mollusc Haminoea orbignyana. Angew. Chem.-Int. Ed. 2003, 42, 2633–2636. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Fujii, Y.; Kizu, H.; Boyd, M.R.; Boettner, F.E.; Doubek, D.L.; Schmidt, J.M.; Chapuis, J.C.; et al. Isolation of Dolastatins 10–15 from the Marine Mollusc Dolabella auricularia. Tetrahedron 1993, 49, 9151–9170. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Tuinman, A.A.; Boettner, F.E.; Kizu, H.; Schmidt, J.M.; Baczynskyj, L.; Tomer, K.B.; Bontems, R.J. The Isolation and Structure of a Remarkable Marine Animal Antineoplastic Constituent: Dolastatin 10. J. Am. Chem. Soc. 1987, 109, 6883–6885. [Google Scholar] [CrossRef]
- Luesch, H.; Moore, R.E.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H. Isolation of Dolastatin 10 from the Marine Cyanobacterium Symploca species VP642 and Total Stereochemistry and Biological Evaluation of Its Analogue Symplostatin 1. J. Nat. Prod. 2001, 64, 907–910. [Google Scholar] [PubMed]
- Cimino, G.; Gavagnin, M. (Eds.) Molluscs: From Chemo-ecological Study to Biotechnological Application; Progress in Molecular and Subcellular Biology; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, ISBN 978-3-540-30879-9. [Google Scholar]
- Benkendorff, K. Molluscan Biological and Chemical Diversity: Secondary Metabolites and Medicinal Resources Produced by Marine Molluscs. Biol. Rev. Camb. Philos. Soc. 2010, 85, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Debelius, H.; Kuiter, R.H. Nudibranchs of the World; IKAN-Unterwasserarchiv: Frankfurt, Germany, 2007; ISBN 978-3-939767-06-0. [Google Scholar]
- Gosliner, T.; Behrens, D.W.; Valdés, Á. Indo-Pacific Nudibranchs and Sea Slugs: A Field Guide to the World’s Most Diverse Fauna; Sea Challengers Natural History Books: San Francisco, CA, USA, 2008; ISBN 978-0-9700574-3-3. [Google Scholar]
- Gosliner, T.; Valdés, Á.; Behrens, D.W. Nudibranch and Sea Slug Identification: Indo-Pacific; New World Publications, Inc.: Jacksonville, FL, USA, 2015; ISBN 978-1-878348-59-3. [Google Scholar]
- Yonow, N. Results of the Rumphius Biohistorical Expedition to Ambon (1990) Part 15. The Suborder Doridina (Mollusca, Gastropods, Opisthobranchia, Nudibranchia). Zool. Meded. 2011, 85, 905–956. [Google Scholar]
- Martynov, A.V.; Korshunova, T.A. Opisthobranch Molluscs of Vietnam (Gastropoda: Opisthobranchia). In Benthic fauna of the Bay of Nhatrang, Southern Vietnam; Britayev, T.A., Pavlov, D.S., Eds.; KMK Scientific Press Ltd.: Moscow, Russia, 2012; Volume 2, pp. 142–257. ISBN 978-5-87317-860-5. [Google Scholar]
- Yonow, N. Results of the Rumphius Biohistorical Expedition to Ambon (1990). Part 11. Doridacea of the Families Chromodorididae and Hexabranchidae (Mollusca, Gastropoda, Opisthobranchia, Nudibranchia), Including Additional Moluccan Material. Zool. Meded. 2001, 75, 1–50. [Google Scholar]
- Yonow, N. Results of the Rumphius Biohistorical Expedition to Ambon (1990). Part 16. The Nudibranchia—Dendronotina, Arminina, Aeolidina, and Doridina (Mollusca: Gastropoda: Heterobranchia). Arch. Molluskenkd. Int. J. Malacol. 2017, 146, 135–172. [Google Scholar] [CrossRef]
- Takamasa, Tonozuka. Opisthobranchs of Bali and Indonesia; Hankyu Communications Co. Ltd.: Tokyo, Japan, 2003; ISBN 4-484-03409-3. Available online: https://combinedhunterunderwatergroup.weebly.com/uploads/1/1/2/3/11235330/indonesia-ssc_june_2017_-_album.pdf (accessed on 5 December 2017).
- Burghardt, I.; Carvalho, R.; Eheberg, D.; Gerung, G.; Kaligis, F.; Mamangkey, G.; Schrödl, M.; Schwabe, E.; Vonnemann, V.; Wägele, H. Molluscan Diversity at Bunaken National Park, Sulawesi. J. Zool. Soc. Wallacea 2006, 2, 29–43. [Google Scholar]
- Jensen, K.R. Evolution of the Sacoglossa (Mollusca, Opisthobranchia) and the ecological associations with their food plants. Evol. Ecol. 1997, 11, 301–335. [Google Scholar] [CrossRef]
- Pereira, R.B.; Andrade, P.B.; Valentão, P. Chemical Diversity and Biological Properties of Secondary Metabolites from Sea Hares of Aplysia Genus. Mar. Drugs 2016, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, G.G.; Luesch, H.; Moore, R.E.; Paul, V.J. The Anticancer Dolastatins as Cyanobacterial Metabolites. In Biodiversity; Wrigley, S.K., Hayes, M.A., Thomas, R., Chrystal, E.J.T., Nicholson, N., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2000; pp. 126–139. [Google Scholar]
- Kato, Y.; Scheuer, P.J. Aplysiatoxin and Debromoaplysiatoxin, Constituents of the Marine Mollusk Stylocheilus longicauda (Quoy and Gaimard, 1824). J. Am. Chem. Soc. 1974, 96, 2245–2246. [Google Scholar] [CrossRef] [PubMed]
- Watson, M. Midgut Gland Toxins of Hawaiian Sea Hares. I. Isolation and Preliminary Toxicological Observations. Toxicon 1973, 11, 259–267. [Google Scholar] [CrossRef]
- Watson, M.; Rayner, M.D. Midgut Gland Toxins of Hawaiian Sea Hares. II. A Preliminary Pharmacological Study. Toxicon 1973, 11, 269–276. [Google Scholar] [CrossRef]
- Moore, R.E. Toxins, Anticancer Agents, and Tumor Promoters from Marine Prokaryotes. Pure Appl. Chem. 1982, 54, 1919–1934. [Google Scholar] [CrossRef]
- Ashida, Y.; Yanagita, R.C.; Takahashi, C.; Kawanami, Y.; Irie, K. Binding Mode Prediction of Aplysiatoxin, a Potent Agonist of Protein Kinase C, Through Molecular Simulation and Structure–Activity Study on Simplified Analogs of the Receptor-Recognition Domain. Bioorg. Med. Chem. 2016, 24, 4218–4227. [Google Scholar] [CrossRef] [PubMed]
- Gallimore, W.A.; Galario, D.L.; Lacy, C.; Zhu, Y.; Scheuer, P.J. Two Complex Proline Esters from the Sea Hare Stylocheilus longicauda. J. Nat. Prod. 2000, 63, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Orjala, J.; Nagle, D.G.; Hsu, V.; Gerwick, W.H. Antillatoxin: An Exceptionally Ichthyotoxic Cyclic Lipopeptide from the Tropical Cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc. 1995, 117, 8281–8282. [Google Scholar] [CrossRef]
- White, J.D.; Hanselmann, R.; Wardrop, D.J. Synthesis of Epiantillatoxin, a Stereoisomer of the Potent Ichthyotoxin from Lyngbya majuscula. J. Am. Chem. Soc. 1999, 121, 1106–1107. [Google Scholar] [CrossRef]
- Nagle, D.G.; Paul, V.J.; Ann Roberts, M. Ypaoamide, a New Broadly Acting Feeding Deterrent from the Marine Cyanobacterium Lyngbya majuscula. Tetrahedron Lett. 1996, 37, 6263–6266. [Google Scholar] [CrossRef]
- Klein, D.; Braekman, J.-C.; Daloze, D.; Hoffmann, L.; Demoulin, V. Laingolide, a Novel 15-Membered Macrolide from Lyngbya bouillonii (cyanophyceae). Tetrahedron Lett. 1996, 37, 7519–7520. [Google Scholar] [CrossRef]
- Cardellina, J.H.; Marner, F.J.; Moore, R.E. Seaweed Dermatitis: Structure of Lyngbyatoxin A. Science 1979, 204, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.J.; Gerwick, W.H. Lyngbyatoxin Biosynthesis: Sequence of Biosynthetic Gene Cluster and Identification of a Novel Aromatic Prenyltransferase. J. Am. Chem. Soc. 2004, 126, 11432–11433. [Google Scholar] [CrossRef] [PubMed]
- Awakawa, T.; Zhang, L.; Wakimoto, T.; Hoshino, S.; Mori, T.; Ito, T.; Ishikawa, J.; Tanner, M.E.; Abe, I. A Methyltransferase Initiates Terpene Cyclization in Teleocidin B Biosynthesis. J. Am. Chem. Soc. 2014, 136, 9910–9913. [Google Scholar] [CrossRef] [PubMed]
- Gallimore, W.A.; Scheuer, P.J. Malyngamides O and P from the Sea Hare Stylocheilus longicauda. J. Nat. Prod. 2000, 63, 1422–1424. [Google Scholar] [CrossRef] [PubMed]
- Cardellina, J.H.; Marner, F.J.; Moore, R.E. Malyngamide A, a Novel Chlorinated Metabolite of the Marine Cyanophyte Lyngbya majuscula. J. Am. Chem. Soc. 1979, 101, 240–242. [Google Scholar] [CrossRef]
- Capper, A.; Tibbetts, I.R.; O’Neil, J.M.; Shaw, G.R. The Fate of Lyngbya majuscula Toxins in Three Potential Consumers. J. Chem. Ecol. 2005, 31, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- Vardaro, R.R.; Di Marzo, V.; Crispino, A.; Cimino, G. Cyercenes, Novel Polypropionate Pyrones from the Autotomizing Mediterranean Mollusc Cyerce cristallina. Tetrahedron 1991, 47, 5569–5576. [Google Scholar] [CrossRef]
- Hay, M.E.; Pawlik, J.R.; Duffy, J.E.; Fenical, W. Seaweed-Herbivore-Predator Interactions: Host-Plant Specialization Reduces Predation on Small Herbivores. Oecologia 1989, 81, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.J.; Fenical, W. Chemical Defense in Tropical Green Algae, Order Caulerpales. Mar. Ecol. Prog. Ser. 1986, 34, 157–169. [Google Scholar] [CrossRef]
- Gavagnin, M.; Marin, A.; Castelluccio, F.; Villani, G.; Cimino, G. Defensive Relationships between Caulerpa prolifera and Its Shelled Sacoglossan Predators. J. Exp. Mar. Biol. Ecol. 1994, 175, 197–210. [Google Scholar] [CrossRef]
- Mozzachiodi, R.; Scuri, R.; Roberto, M.; Brunelli, M. Caulerpenyne, a Toxin from the Seaweed Caulerpa taxifolia, Depresses Afterhyperpolarization in Invertebrate Neurons. Neuroscience 2001, 107, 519–526. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; Lopez Gresa, M.P.; Gavagnin, M.; Manzo, E.; Mollo, E.; D’Souza, L.; Cimino, G. New Caulerpenyne-Derived Metabolites of an Elysia Sacoglossan from the South Indian Coast. Molecules 2006, 11, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Cimino, G.; Fontana, A.; Gavagnin, M. Marine Opisthobranch Molluscs: Chemistry and Ecology in Sacoglossans and Dorids. Curr. Org. Chem. 1999, 3, 327–372. [Google Scholar]
- Cimino, G.; Ghiselin, M.T. Chemical Defense and the Evolution of Opisthobranch Gastropods; California Academy of Sciences: San Francisco, CA, USA, 2009; ISBN 978-0-940228-79-5. [Google Scholar]
- Gao, J.; Hamann, M.T. Chemistry and Biology of Kahalalides. Chem. Rev. 2011, 111, 3208–3235. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Muniain, C.; Castelluccio, F.; Iannicelli, O.; Gavagnin, M. First Chemical Study of the Sacoglossan Elysia patagonica: Isolation of a γ-pyrone propionate hydroperoxide. Biochem. Syst. Ecol. 2013, 49, 172–175. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; Devi, P.; Carbone, M.; Mathieu, V.; Kiss, R.; Casapullo, A.; Gavagnin, M. Kahalalide F Analogues from the Mucous Secretion of Indian Sacoglossan Mollusc Elysia ornata. Tetrahedron 2016, 72, 625–631. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Montanaro, D.; Ortea, J.; Cimino, G. Chemical Studies of Caribbean Sacoglossans: Dietary Relationships with Green Algae and Ecological Implications. J. Chem. Ecol. 2000, 26, 1563–1578. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Cimino, G. Is Phototridachiahydropyrone a True Natural Product? Rev. Bras. Farmacogn. 2015, 25, 588–591. [Google Scholar] [CrossRef]
- Cutignano, A.; Cimino, G.; Villani, G.; Fontana, A. Shaping the Polypropionate Biosynthesis in the Solar-Powered Mollusc Elysia viridis. ChemBioChem 2009, 10, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.J.; Van Alstyne, K.L. Use of Ingested Algal Diterpenoids by Elysia halimedae Macnae (Opisthobranchia : Ascoglossa) As Antipredator Defenses. J. Exp. Mar. Biol. Ecol. 1988, 119, 15–29. [Google Scholar] [CrossRef]
- Hamann, M.T.; Scheuer, P.J. Kahalalide F: A Bioactive Depsipeptide from the Sacoglossan Mollusk Elysia rufescens and the Green Alga Bryopsis sp. J. Am. Chem. Soc. 1993, 115, 5825–5826. [Google Scholar] [CrossRef]
- Ashour, M.; Edrada, R.; Ebel, R.; Wray, V.; Wätjen, W.; Padmakumar, K.; Müller, W.E.G.; Lin, W.H.; Proksch, P. Kahalalide Derivatives from the Indian Sacoglossan Mollusk Elysia grandifolia. J. Nat. Prod. 2006, 69, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.T.; Otto, C.S.; Scheuer, P.J.; Dunbar, D.C. Kahalalides: Bioactive Peptides from a Marine Mollusk Elysia rufescens and Its Algal Diet Bryopsis sp. (1). J. Org. Chem. 1996, 61, 6594–6600. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Waters, A.L.; Valeriote, F.A.; Hamann, M.T. An Efficient and Cost-Effective Approach to Kahalalide F N-Terminal Modifications Using a Nuisance Algal Bloom of Bryopsis pennata. Biochim. Biophys. Acta BBA—Gen. Subj. 2015, 1850, 1849–1854. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Fricke, W.F.; Hamann, M.T.; Esquenazi, E.; Dorrestein, P.C.; Hill, R.T. Characterization of the Bacterial Community of the Chemically Defended Hawaiian Sacoglossan Elysia rufescens. Appl. Environ. Microbiol. 2013, 79, 7073–7081. [Google Scholar] [CrossRef] [PubMed]
- Gavagnin, M.; Spinella, A.; Crispino, A.; Epifanio, R.; Marin, A.; Cimino, G. Chemical-Components of the Mediterranean Ascoglossan Thuridilla hopei. Gazzetta Chim. Ital. 1993, 123, 205–208. [Google Scholar]
- Somerville, M.J.; Katavic, P.L.; Lambert, L.K.; Pierens, G.K.; Blanchfield, J.T.; Cimino, G.; Mollo, E.; Gavagnin, M.; Banwell, M.G.; Garson, M.J. Isolation of Thuridillins D–F, Diterpene Metabolites from the Australian Sacoglossan Mollusk Thuridilla splendens; Relative Configuration of the Epoxylactone Ring. J. Nat. Prod. 2012, 75, 1618–1624. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.; Ciminiello, P.; Fenical, W. Diterpenoid Feeding Deterrents from the Pacific Green-Alga Pseudochlorodesmis furcellata. Phytochemistry 1988, 27, 1011–1014. [Google Scholar] [CrossRef]
- Reese, M.T.; Gulavita, N.K.; Nakao, Y.; Hamann, M.T.; Yoshida, W.Y.; Coval, S.J.; Scheuer, P.J. Kulolide: A Cytotoxic Depsipeptide from a Cephalaspidean Mollusk, Philinopsis speciosa1. J. Am. Chem. Soc. 1996, 118, 11081–11084. [Google Scholar] [CrossRef]
- Kimura, J.; Takada, Y.; Inayoshi, T.; Nakao, Y.; Goetz, G.; Yoshida, W.Y.; Scheuer, P.J. Kulokekahilide-1, a Cytotoxic Depsipeptide from the Cephalaspidean Mollusk Philinopsis speciosa. J. Org. Chem. 2002, 67, 1760–1767. [Google Scholar] [CrossRef] [PubMed]
- Nakao, Y.; Yoshida, W.Y.; Szabo, C.M.; Baker, B.J.; Scheuer, P.J. More Peptides and Other Diverse Constituents of the Marine Mollusk Philinopsis speciosa. J. Org. Chem. 1998, 63, 3272–3280. [Google Scholar] [CrossRef]
- Nakao, Y.; Yoshida, W.Y.; Takada, Y.; Kimura, J.; Yang, L.; Mooberry, S.L.; Scheuer, P.J. Kulokekahilide-2, a Cytotoxic Depsipeptide from a Cephalaspidean Mollusk Philinopsis speciosa. J. Nat. Prod. 2004, 67, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Rivera, E.; Paul, V.J. Chemical Deterrence of a Cyanobacterial Metabolite Against Generalized and Specialized Grazers. J. Chem. Ecol. 2007, 33, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, P.D.; Byrum, T.; Liu, W.-T.; Dorrestein, P.C.; Gerwick, W.H. Viequeamide A, a Cytotoxic Member of the Kulolide Superfamily of Cyclic Depsipeptides from a Marine Button Cyanobacterium. J. Nat. Prod. 2012, 75, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- Sleeper, H.L.; Fenical, W. Navenones A-C: Trail-Breaking Alarm Pheromones from the Marine Opisthobranch Navanax inermis. J. Am. Chem. Soc. 1977, 99, 2367–2368. [Google Scholar] [CrossRef]
- Marin, A.; Alvarez, L.A.; Cimino, G.; Spinella, A. Chemical Defence in Cephalaspidean Gastropods: Origin, Anatomical Location and Ecological Roles. J. Molluscan Stud. 1999, 65, 121–131. [Google Scholar] [CrossRef]
- Cimino, G.; Sodano, G.; Spinella, A. New Propionate-Derived Metabolites from Aglaja depicta and from Its Prey Bulla striata (Opisthobranch Mollusks). J. Org. Chem. 1987, 52, 5326–5331. [Google Scholar] [CrossRef]
- Spinella, A.; Alvarez, L.A.; Cimino, G. Predator—Prey Relationship between Navanax inermis and Bulla gouldiana : A Chemical Approach. Tetrahedron 1993, 49, 3203–3210. [Google Scholar] [CrossRef]
- Becerro, M.A.; Starmer, J.A.; Paul, V.J. Chemical Defenses of Cryptic and Aposematic Gastropterid Molluscs Feeding on Their Host Sponge Dysidea granulosa. J. Chem. Ecol. 2006, 32, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Ortlepp, S.; Pedpradap, S.; Dobretsov, S.; Proksch, P. Antifouling Activity of Sponge-Derived Polybrominated Diphenyl Ethers and Synthetic Analogues. Biofouling 2008, 24, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Mollo, E.; Gavagnin, M.; Carbone, M.; Castelluccio, F.; Pozone, F.; Roussis, V.; Templado, J.; Ghiselin, M.T.; Cimino, G. Factors Promoting Marine Invasions: A Chemoecological Approach. Proc. Natl. Acad. Sci. USA 2008, 105, 4582–4586. [Google Scholar] [CrossRef] [PubMed]
- Poiner, A.; Paul, V.J.; Scheuer, P.J. Kumepaloxane, a Rearranged Trisnor Sesquiterpene from the Bubble Shell Haminoea cymbalum. Tetrahedron 1989, 45, 617–622. [Google Scholar] [CrossRef]
- Spinella, A.; Alvarez, L.; Passeggio, A.; Cimino, G. New 3-Alkylpyridines from 3 Mediterranean Cephalaspidean Mollusks—Structure, Ecological Role and Taxonomic Relevance. Tetrahedron 1993, 49, 1307–1314. [Google Scholar] [CrossRef]
- Blihoghe, D.; Manzo, E.; Villela, A.; Cutignano, A.; Picariello, G.; Faimali, M.; Fontana, A. Evaluation of the Antifouling Properties of 3-Alyklpyridine Compounds. Biofouling 2011, 27, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Cutignano, A.; Blihoghe, D.; Fontana, A.; Villani, G.; d’Ippolito, G.; Cimino, G. Fusaripyrones, Novel Polypropionates from the Mediterranean Mollusc Haminoea fusari. Tetrahedron 2007, 63, 12935–12939. [Google Scholar] [CrossRef]
- Wesson, K.J.; Hamann, M.T. Keenamide A, a Bioactive Cyclic Peptide from the Marine Mollusk Pleurobranchus forskalii. J. Nat. Prod. 1996, 59, 629–631. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.C.; Wakimoto, T.; Takada, K.; Ohtsuki, T.; Uchiyama, N.; Goda, Y.; Abe, I. Cycloforskamide, a Cytotoxic Macrocyclic Peptide from the Sea Slug Pleurobranchus forskalii. J. Nat. Prod. 2013, 76, 1388–1391. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Leikoski, N.; Fewer, D.P.; Jokela, J. Cyanobactins—Ribosomal Cyclic Peptides Produced by Cyanobacteria. Appl. Microbiol. Biotechnol. 2010, 86, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Wakimoto, T.; Tan, K.C.; Abe, I. Ergot Alkaloid from the Sea Slug Pleurobranchus forskalii. Toxicon 2013, 72, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Mulac, D.; Humpf, H.-U. Cytotoxicity and Accumulation of Ergot Alkaloids in Human Primary Cells. Toxicology 2011, 282, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Palomar, A.J.; Hong, E.P.; Schmitz, F.J.; Valeriote, F.A. Cytotoxic Lissoclimide-Type Diterpenes from the Molluscs Pleurobranchus albiguttatus and Pleurobranchus forskalii. J. Nat. Prod. 2004, 67, 1415–1418. [Google Scholar] [CrossRef] [PubMed]
- Robert, F.; Gao, H.Q.; Donia, M.; Merrick, W.C.; Hamann, M.T.; Pelletier, J. Chlorolissoclimides: New Inhibitors of Eukaryotic Protein Synthesis. RNA 2006, 12, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Carmelya, S.; Ilanb, M.; Kashmana, Y. 2-Amino Imidazole Alkaloids from the Marine Sponge Leucetta chagosensis. Tetrahedron 1989, 45, 2193–2200. [Google Scholar] [CrossRef]
- Copp, B.R.; Fairchild, C.R.; Cornell, L.; Casazza, A.M.; Robinson, S.; Ireland, C.M. Naamidine A Is an Antagonist of the Epidermal Growth Factor Receptor and an in Vivo Active Antitumor Agent. J. Med. Chem. 1998, 41, 3909–3911. [Google Scholar] [CrossRef] [PubMed]
- LaBarbera, D.V.; Modzelewska, K.; Glazar, A.I.; Gray, P.D.; Kaur, M.; Liu, T.; Grossman, D.; Harper, M.K.; Kuwada, S.K.; Moghal, N.; et al. The Marine Alkaloid Naamidine A Promotes Caspase-Dependent Apoptosis in Tumor Cells. Anticancer Drugs 2009, 20, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Mai, T.; Tintillier, F.; Lucasson, A.; Moriou, C.; Bonno, E.; Petek, S.; Magré, K.; Al Mourabit, A.; Saulnier, D.; Debitus, C. Quorum Sensing Inhibitors from Leucetta chagosensis Dendy, 1863. Lett. Appl. Microbiol. 2015, 61, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Alvi, K.A.; Crews, P.; Loughhead, D.G. Structures and Total Synthesis of 2-Aminoimidazoles from a Notodoris Nudibranch. J. Nat. Prod. 1991, 54, 1509–1515. [Google Scholar] [CrossRef]
- Carroll, A.R.; Bowden, B.F.; Coll, J.C. New Imidazole Alkaloids from the Sponge Leucetta sp. and the Associated Predatory Nudibranch Notodoris gardineri. Aust. J. Chem. 1993, 468, 1229–1234. [Google Scholar] [CrossRef]
- Alvi, K.A.; Peters, B.M.; Lisa, H.M.; Phillip, C. 2-Aminoimidazoles and Their Zinc Complexes from Indo-Pacific Leucetta sponges and Notodoris nudibranchs. Tetrahedron 1993, 49, 329–336. [Google Scholar] [CrossRef]
- Ciminiello, P.; Fattorusso, E.; Magno, S.; Mangoni, A. Clathridine and Its Zinc Complex, Novel Metabolites from the Marine Sponge Clathrina clathrus. Tetrahedron 1989, 45, 3873–3878. [Google Scholar] [CrossRef]
- Roué, M.; Domart-Coulon, I.; Ereskovsky, A.; Djediat, C.; Perez, T.; Bourguet-Kondracki, M.-L. Cellular Localization of Clathridimine, an Antimicrobial 2-Aminoimidazole Alkaloid Produced by the Mediterranean Calcareous Sponge Clathrina clathrus. J. Nat. Prod. 2010, 73, 1277–1282. [Google Scholar] [CrossRef] [PubMed]
- Grkovic, T.; Appleton, D.R.; Copp, B.R. Chemistry and Chemical Ecology of some of the Common Opisthobranch Molluscs Found on the Shores of NE New Zealand. Chem. N. Z. 2005, 69, 12. [Google Scholar]
- König, G.M.; Wright, A.D. Sesquiterpene Content of the Antibacterial Dichloromethane Extract of the Marine Red Alga Laurencia obtusa. Planta Med. 1997, 63, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Kawamoto, T.; Miwa, H.; Suzuki, M. Potent Antibacterial Activity of Halogenated Compounds against Antibiotic-Resistant Bacteria. Planta Med. 2004, 70, 1087–1090. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Gavagnin, M.; Haber, M.; Guo, Y.-W.; Fontana, A.; Manzo, E.; Genta-Jouve, G.; Tsoukatou, M.; Rudman, W.B.; Cimino, G.; et al. Packaging and Delivery of Chemical Weapons: A Defensive Trojan Horse Stratagem in Chromodorid Nudibranchs. PLoS ONE 2013, 8, e62075. [Google Scholar] [CrossRef] [PubMed]
- Kakou, Y.; Crews, P.; Bakus, G.J. Dendrolasin and Latrunculin A from the Fijian Sponge Spongia mycofijiensis and an Associated Nudibranch Chromodoris lochi. J. Nat. Prod. 1987, 50, 482–484. [Google Scholar] [CrossRef]
- Kashman, Y.; Groweiss, A.; Shmueli, U. Latrunculin, a New 2-Thiazolidinone Macrolide from the Marine Sponge Latrunculia magnifica. Tetrahedron Lett. 1980, 21, 3629–3632. [Google Scholar] [CrossRef]
- Pika, J.; Faulkner, D.J. Unusual Chlorinated Homo-Diterpenes from the South African Nudibranch Chromodoris hamiltoni. Tetrahedron 1995, 51, 8189–8198. [Google Scholar] [CrossRef]
- Cheney, K.L.; White, A.; Mudianta, I.W.; Winters, A.E.; Quezada, M.; Capon, R.J.; Mollo, E.; Garson, M.J. Choose Your Weaponry: Selective Storage of a Single Toxic Compound, Latrunculin A, by Closely Related Nudibranch Molluscs. PLoS ONE 2016, 11, e0145134. [Google Scholar] [CrossRef] [PubMed]
- El Sayed, K.A.; Youssef, D.T.A.; Marchetti, D. Bioactive Natural and Semisynthetic Latrunculins. J. Nat. Prod. 2006, 69, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Crews, P.; Kakou, Y.; Quinoa, E. Mycothiazole, a Polyketide Heterocycle from a Marine Sponge. J. Am. Chem. Soc. 1988, 110, 4365–4368. [Google Scholar] [CrossRef]
- Sonnenschein, R.N.; Johnson, T.A.; Tenney, K.; Valeriote, F.A.; Crews, P. A Reassignment of (−)-Mycothiazole and the Isolation of a Related Diol. J. Nat. Prod. 2006, 69, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.B.; Mahdi, F.; Liu, Y.; Coothankandaswamy, V.; Jekabsons, M.B.; Johnson, T.A.; Sashidhara, K.V.; Crews, P.; Nagle, D.G.; Zhou, Y.-D. The Marine Sponge Metabolite Mycothiazole: A Novel Prototype Mitochondrial Complex I Inhibitor. Bioorg. Med. Chem. 2010, 18, 5988–5994. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.J.; Singh, A.J.; Cameron, A.; Tan, A.S.; Leahy, D.C.; O’Sullivan, D.; Joshi, P.; La Flamme, A.C.; Northcote, P.T.; Berridge, M.V.; et al. Mitochondrial Genome-Knockout Cells Demonstrate a Dual Mechanism of Action for the Electron Transport Complex I Inhibitor Mycothiazole. Mar. Drugs 2012, 10, 900–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corley, D.G.; Herb, R.; Moore, R.E.; Scheuer, P.J.; Paul, V.J. Laulimalides. New Potent Cytotoxic Macrolides from a Marine Sponge and a Nudibranch Predator. J. Org. Chem. 1988, 53, 3644–3646. [Google Scholar] [CrossRef]
- Mooberry, S.L.; Randall-Hlubek, D.A.; Leal, R.M.; Hegde, S.G.; Hubbard, R.D.; Zhang, L.; Wender, P.A. Microtubule-Stabilizing Agents Based on Designed Laulimalide Analogues. Proc. Natl. Acad. Sci. USA 2004, 101, 8803–8808. [Google Scholar] [CrossRef] [PubMed]
- Churchill, C.D.M.; Klobukowski, M.; Tuszynski, J.A. Analysis of the Binding Mode of Laulimalide to Microtubules: Establishing a Laulimalide-Tubulin Pharmacophore. J. Biomol. Struct. Dyn. 2016, 34, 1455–1469. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Sakamoto, K.; Amano, H.; Arakawa, Y.; Nagarekawa, Y.; Komori, T.; Higuchi, R.; Sasaki, T. New Cytotoxic Sesterterpenoids from the Nudibranch Chromodoris inornata. Tetrahedron 1999, 55, 9133–9142. [Google Scholar] [CrossRef]
- Gavagnin, M.; Vardaro, R.; Avila, C.; Cimino, G.; Ortea, J. Ichthyotoxic Diterpenoids from the Cantabrian Nudibranch Chromodoris luteorosea. J. Nat. Prod. 1992, 55, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Molinski, T.F.; Faulkner, D.J. Aromatic Norditerpenes from the Nudibranch Chromodoris macfarlandi. J. Org. Chem. 1986, 51, 2601–2603. [Google Scholar] [CrossRef]
- Molinski, T.F.; Faulkner, D.J.; He, C.H.; Van Duyne, G.D.; Clardy, J. Three New Rearranged Spongian Diterpenes from Chromodoris macfarlandi: Reappraisal of the Structures of Dendrillolides A and B. J. Org. Chem. 1986, 51, 4564–4567. [Google Scholar] [CrossRef]
- Schnermann, M.J.; Beaudry, C.M.; Egorova, A.V.; Polishchuk, R.S.; Suetterlin, C.; Overman, L.E. Golgi-Modifying Properties of Macfarlandin E and the Synthesis and Evaluation Of Its 2,7-Dioxabicyclo[3.2.1]Octan-3-One Core. Proc. Natl. Acad. Sci. USA 2010, 107, 6158–6163. [Google Scholar] [CrossRef] [PubMed]
- Suciati; Lambert, L.K.; Garson, M.J. Structures and Anatomical Distribution of Oxygenated Diterpenes in the Australian Nudibranch Chromodoris reticulata. Aust. J. Chem. 2011, 64, 757–765. [Google Scholar] [CrossRef]
- Schulte, G.; Scheuer, P.J.; McConnell, O.J. Two Furanosesquiterpene Marine Metabolites with Antifeedant Properties. Helv. Chim. Acta 1980, 63, 2159–2167. [Google Scholar] [CrossRef]
- De Silva, E.D.; Scheuer, P.J. Furanoditerpenoids from the Dorid Nudibranch Casella atromarginata. Heterocycles 1982, 17, 167–170. [Google Scholar] [CrossRef]
- Karuso, P. Chemical Ecology of the Nudibranchs. In Bioorganic Marine Chemistry; Scheuer, P.P.J., Ed.; Bioorganic Marine Chemistry; Springer: Berlin/Heidelberg, Germany, 1987; pp. 31–60. ISBN 978-3-642-72728-3. [Google Scholar]
- Manzo, E.; Gavagnin, M.; Somerville, M.J.; Mao, S.-C.; Ciavatta, M.L.; Mollo, E.; Schupp, P.J.; Garson, M.J.; Guo, Y.-W.; Cimino, G. Chemistry of Glossodoris Nudibranchs: Specific Occurrence of 12-Keto Scalaranes. J. Chem. Ecol. 2007, 33, 2325–2336. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; Cavaliere, P.; Ungur, N.; D’Souza, L.; Parameswaram, P.S.; Cimino, G. New Scalaranes from the Nudibranch Glossodoris atromarginata and Its Sponge Prey. J. Nat. Prod. 1999, 62, 1367–1370. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.; Wright, A.D.; Reinscheid, U.; König, G.M. Three New Spongian Diterpenes from the Fijian Marine Sponge Spongia sp. Nat. Prod. Commun. 2009, 4, 315–322. [Google Scholar] [PubMed]
- Somerville, M.J.; Mollo, E.; Cimino, G.; Rungprom, W.; Garson, M.J. Spongian Diterpenes from Australian Nudibranchs: An Anatomically Guided Chemical Study of Glossodoris atromarginata. J. Nat. Prod. 2006, 69, 1086–1088. [Google Scholar] [CrossRef] [PubMed]
- Betancur-Galvis, L.; Zuluaga, C.; Arnó, M.; González, M.A.; Zaragozá, R.J. Cytotoxic Effect (on Tumor Cells) and in Vitro Antiviral Activity against Herpes Simplex Virus of Synthetic Spongiane Diterpenes. J. Nat. Prod. 2002, 65, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Gavagnin, M.; Mollo, E.; Docimo, T.; Guo, Y.-W.; Cimino, G. Scalarane Metabolites of the Nudibranch Glossodoris rufomarginata and Its Dietary Sponge from the South China Sea. J. Nat. Prod. 2004, 67, 2104–2107. [Google Scholar] [CrossRef] [PubMed]
- Kamel, H.N.; Kim, Y.B.; Rimoldi, J.M.; Fronczek, F.R.; Ferreira, D.; Slattery, M. Scalarane Sesterterpenoids: Semisynthesis and Biological Activity. J. Nat. Prod. 2009, 72, 1492–1496. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-Y.; Sung, P.-J.; Chang, Y.-L.; Pan, S.-L.; Teng, C.-M. Heteronemin, a Spongean Sesterterpene, Induces Cell Apoptosis and Autophagy in Human Renal Carcinoma Cells. BioMed Res. Int. 2015, 2015, e738241. [Google Scholar] [CrossRef] [PubMed]
- Kohmoto, S.; Mcconnell, O.J.; Wright, A.; Cross, S. Isospongiadiol, a Cytotoxic and Antiviral Diterpene from a Caribbean Deep Water Marine Sponge, Spongia sp. Chem. Lett. 1987, 16, 1687–1690. [Google Scholar] [CrossRef]
- Agrawal, M. Isolation and Structural Elucidation of Cytotoxic Agents from Marine Invertebrates and Plants Sourced from the Great Barrier Reef, Australia. Ph.D. Thesis, James Cook University, Townsville, Australia, 2007. [Google Scholar]
- Yong, K.W.; Garson, M.J.; Bernhardt, P.V. Absolute Structures and Conformations of the Spongian Diterpenes Spongia-13(16), 14-dien-3-one, Epispongiadiol and Spongiadiol. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 2009, 65, o167–o170. [Google Scholar] [CrossRef] [PubMed]
- Longley, R.E.; McConnell, O.J.; Essich, E.; Harmody, D. Evaluation of Marine Sponge Metabolites for Cytotoxicity and Signal Transduction Activity. J. Nat. Prod. 1993, 56, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Wonganuchitmeta, S.; Yuenyongsawad, S.; Keawpradub, N.; Plubrukarn, A. Antitubercular Sesterterpenes from the Thai Sponge Brachiaster sp. J. Nat. Prod. 2004, 67, 1767–1770. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; Mollo, E.; Ortea, J.; Gavagnin, M.; Cimino, G. Scalarane and Homoscalarane Compounds from the Nudibranchs Glossodoris sedna and Glossodoris dalli: Chemical and Biological Properties. J. Nat. Prod. 2000, 63, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Sakamoto, K.; Arao, K.; Komori, T.; Higuchi, R.; Sasaki, T. Dorisenones, cytotoxic spongian diterpenoids, from the nudibranch Chromodoris obsoleta. Tetrahedron 1996, 52, 8187–8198. [Google Scholar] [CrossRef]
- Hirayama, Y.; Katavic, P.L.; White, A.M.; Pierens, G.K.; Lambert, L.K.; Winters, A.E.; Kigoshi, H.; Kita, M.; Garson, M.J. New Cytotoxic Norditerpenes from the Australian Nudibranchs Goniobranchus splendidus and Goniobranchus daphne. Aust. J. Chem. 2016, 69, 136–144. [Google Scholar] [CrossRef]
- Rueda, A.; Losada, A.; Fernandez, R.; Cabanas, C.; Garcia-Fernandez, L.F.; Reyes, F.; Cuevas, C. Gracilins G-I, Cytotoxic Bisnorditerpenes from Spongionella pulchella, and the Anti-Adhesive Properties of Gracilin B. Lett. Drug Des. Discov. 2006, 3, 753–760. [Google Scholar] [CrossRef]
- Andoni Sanchez, J.; Alfonso, A.; Leiros, M.; Alonso, E.; Rateb, M.E.; Jaspars, M.; Houssen, W.E.; Ebel, R.; Tabudravu, J.; Botana, L.M. Identification of Spongionella Compounds as Cyclosporine A Mimics. Pharmacol. Res. 2016, 107, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Leiros, M.; Alonso, E.; Rateb, M.E.; Houssen, W.E.; Ebel, R.; Jaspars, M.; Alfonso, A.; Botana, L.M. Gracilins: Spongionella-Derived Promising Compounds for Alzheimer Disease. Neuropharmacology 2015, 93, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Karuso, P.; Scheuer, P.J. Natural Products from Three Nudibranchs: Nembrotha kubaryana, Hypselodoris infucata and Chromodoris petechialis. Molecules 2002, 7, 1–6. [Google Scholar] [CrossRef]
- Putz, A.; König, G.M.; Wägele, H. Defensive Strategies of Cladobranchia (Gastropoda, Opisthobranchia). Nat. Prod. Rep. 2010, 27, 1386–1402. [Google Scholar] [CrossRef] [PubMed]
- Frick, K. Response in Nematocyst Uptake by the Nudibranch Flabellina verrucosa to the Presence of Various Predators in the Southern Gulf of Maine. Biol. Bull. 2003, 205, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Affeld, S.; Wägele, H.; Avila, C.; Kehraus, S.; König, G.M. Distribution of Homarine in some Opisthobranchia (Gastropoda: Mollusca). Bonn. Zool. Beitr. 2006, 55, 181–190. [Google Scholar]
- Sodano, G.; Spinella, A. Janolusimide, a Lipophilic Tripeptide Toxin from the Nudibranch Mollusk Janolus cristatus. Tetrahedron Lett. 1986, 27, 2505–2508. [Google Scholar] [CrossRef]
- Wang, J.; Prinsep, M.R.; Gordon, D.P.; Page, M.J.; Copp, B.R. Isolation and Stereospecific Synthesis of Janolusimide B from a New Zealand Collection of the Bryozoan Bugula flabellata. J. Nat. Prod. 2015, 78, 530–533. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K.; Davis, A.R.; Bremner, J.B. Chemical Defense in the Egg Masses of Benthic Invertebrates: An Assessment of Antibacterial Activity in 39 Mollusks and 4 Polychaetes. J. Invertebr. Pathol. 2001, 78, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Cimino, G.; Ghiselin, M.T. Chemical Defense and Evolutionary Trends in Biosynthetic Capacity among Dorid Nudibranchs (Mollusca: Gastropoda: Opisthobranchia). Chemoecology 1999, 9, 187–207. [Google Scholar] [CrossRef]
- Faulkner, D.J. Marine Natural Products. Nat. Prod. Rep. 2001, 18, 1R–49R. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Castelluccio, F.; Ghiselin, M.T.; Calado, G.; Cimino, G. Can Molluscs Biosynthesize Typical Sponge Metabolites? The Case of the Nudibranch Doriopsilla areolata. Tetrahedron 2001, 57, 8913–8916. [Google Scholar] [CrossRef]
- Sakio, Y.; Hirano, Y.J.; Hayashi, M.; Komiyama, K.; Ishibashi, M. Dendocarbins A–N, New Drimane Sesquiterpenes from the Nudibranch Dendrodoris carbunculosa. J. Nat. Prod. 2001, 64, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Nakanishi, K. Insect Antifeedants and Repellents from African Plants. In Host Plant Resistance to Pests; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1977; Volume 62, pp. 165–178. ISBN 978-0-8412-0389-1. [Google Scholar]
- Cimino, G.; De Rosa, S.; De Stefano, S.; Sodano, G.; Villani, G. Dorid Nudibranch Elaborates Its Own Chemical Defense. Science 1983, 219, 1237–1238. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Taniguchi, M. Polygodial, an Antifungal Potentiator. J. Nat. Prod. 1988, 51, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Cimino, G.; Sodano, G.; Spinella, A. Occurrence of Olepupuane in Two Mediterranean Nudibranchs: A Protected Form of Polygodial. J. Nat. Prod. 1988, 51, 1010–1011. [Google Scholar] [CrossRef] [PubMed]
- Fahey, S.J.; Carroll, A.R. Natural Products Isolated from Species of Halgerda Bergh, 1880 (Mollusca: Nudibranchia) and Their Ecological and Evolutionary Implications. J. Chem. Ecol. 2007, 33, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Marin, A.; Belluga, M.D.L.; Scognamiglio, G.; Cimino, G. Morphological and Chemical Camouflage of the Mediterranean Nudibranch Discodoris indecora on the Sponges Ircinia variabilis and Ircinia fasciculata. J. Molluscan Stud. 1997, 63, 431–439. [Google Scholar] [CrossRef]
- Fontana, A.; Cavaliere, P.; Wahidulla, S.; Naik, C.G.; Cimino, G. A New Antitumor Isoquinoline Alkaloid from the Marine Nudibranch Jorunna funebris. Tetrahedron 2000, 56, 7305–7308. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Marine-Sourced Anti-Cancer and Cancer Pain Control Agents in Clinical and Late Preclinical Development. Mar. Drugs 2014, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Drugs and Drug Candidates from Marine Sources: An Assessment of the Current “State of Play”. Planta Med. 2016, 82, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Menchaca, R.; Martínez, V.; Rodríguez, A.; Rodríguez, N.; Flores, M.; Gallego, P.; Manzanares, I.; Cuevas, C. Synthesis of Natural Ecteinascidins (ET-729, ET-745, ET-759B, ET-736, ET-637, ET-594) from Cyanosafracin B. J. Org. Chem. 2003, 68, 8859–8866. [Google Scholar] [CrossRef] [PubMed]
- Roesener, J.A.; Scheuer, P.J. Ulapualide A and B, Extraordinary Antitumor Macrolides from Nudibranch Eggmasses. J. Am. Chem. Soc. 1986, 108, 846–847. [Google Scholar] [CrossRef]
- Matsunaga, S.; Fusetani, N.; Hashimoto, K.; Koseki, K.; Noma, M. Bioactive Marine Metabolites. Part 13. Kabiramide C, a Novel Antifungal Macrolide from Nudibranch Eggmasses. J. Am. Chem. Soc. 1986, 108, 847–849. [Google Scholar] [CrossRef]
- Parrish, S.M.; Yoshida, W.; Yang, B.; Williams, P.G. Ulapualides C–E Isolated from a Hawaiian Hexabranchus sanguineus Egg Mass. J. Nat. Prod. 2017, 80, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, J.R.; Kernan, M.R.; Molinski, T.F.; Harper, M.K.; Faulkner, D.J. Defensive Chemicals of the Spanisch Dancer Nudibranch Hexabranchus sanguineus and Its Egg Ribbons: Macrolides Derived from a Sponge Diet. J. Exp. Mar. Biol. Ecol. 1988, 119, 99–109. [Google Scholar] [CrossRef]
- Matsunaga, S.; Fusetani, N.; Hashimoto, K.; Koseki, K.; Noma, M.; Noguchi, H.; Sankawa, U. Bioactive Marine Metabolites. 25. Further Kabiramides and Halichondramides, Cytotoxic Macrolides Embracing Trisoxazole, from the Hexabranchus egg masses. J. Org. Chem. 1989, 54, 1360–1363. [Google Scholar] [CrossRef]
- Kernan, M.R.; Molinski, T.F.; Faulkner, D.J. Macrocyclic Antifungal Metabolites from the Spanish Dancer Nudibranch Hexabranchus sanguineus and Sponges of the Genus Halichondria. J. Org. Chem. 1988, 53, 5014–5020. [Google Scholar] [CrossRef]
- Klenchin, V.A.; Allingham, J.S.; King, R.; Tanaka, J.; Marriott, G.; Rayment, I. Trisoxazole Macrolide Toxins Mimic the Binding of Actin-Capping Proteins to Actin. Nat. Struct. Biol. 2003, 10, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Melville, J.L.; Moal, I.H.; Baker-Glenn, C.; Shaw, P.E.; Pattenden, G.; Hirst, J.D. The Structural Determinants of Macrolide-Actin Binding: In Silico Insights. Biophys. J. 2007, 92, 3862–3867. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Fujii, Y.; Hirayama, Y.; Kobayashi, S.; Hayakawa, I.; Kigoshi, H. Design, Synthesis, and Biological Evaluations of Aplyronine A–Mycalolide B Hybrid Compound. Org. Lett. 2012, 14, 1290–1293. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.H.; Petchprayoon, C.; Hoepker, A.C.; Moriarty, N.W.; Fink, S.J.; Cecere, G.; Paterson, I.; Adams, P.D.; Marriott, G. Structural and Biochemical Studies of Actin in Complex with Synthetic Macrolide Tail Analogues. ChemMedChem 2014, 9, 2286–2293. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, S. Trisoxazole macrolides from Hexabranchus Nudibranchs and Other Marine Invertebrates. Prog. Mol. Subcell. Biol. 2006, 43, 241–260. [Google Scholar] [PubMed]
- Ueoka, R.; Uria, A.R.; Reiter, S.; Mori, T.; Karbaum, P.; Peters, E.E.; Helfrich, E.J.N.; Morinaka, B.I.; Gugger, M.; Takeyama, H.; et al. Metabolic and Evolutionary Origin of Actin-Binding Polyketides from Diverse Organisms. Nat. Chem. Biol. 2015, 11, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Yeung, K.-S.; Paterson, I. Actin-Binding Marine Macrolides: Total Synthesis and Biological Importance. Angew. Chem. Int. Ed. 2002, 41, 4632–4653. [Google Scholar] [CrossRef] [PubMed]
- Tetlow, D.J.; Winder, S.J.; Aïssa, C. The Synthesis and Biological Evaluation of a Kabiramide C Fragment Modified with a WH2 Consensus Actin-Binding Motif as a Potential Disruptor of the Actin Cytoskeleton. Chem. Commun. 2016, 52, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Stoffels, B.E.M.W.; van der Meij, S.E.T.; Hoeksema, B.W.; van Alphen, J.; van Alen, T.; Meyers-Muñoz, M.A.; Voogd, N.J.D.; Tuti, Y.; van der Velde, G. Phylogenetic relationships within the Phyllidiidae (Opisthobranchia, Nudibranchia). ZooKeys 2016, 605, 1–35. [Google Scholar] [CrossRef]
- Ritson-Williams, R.; Paul, V.J. Marine Benthic Invertebrates Use Multimodal Cues for Defense against Reef Fish. Mar. Ecol. Prog. Ser. 2007, 340, 29–39. [Google Scholar] [CrossRef]
- Manzo, E.; Ciavatta, M.L.; Gavagnin, M.; Mollo, E.; Guo, Y.-W.; Cimino, G. Isocyanide Terpene Metabolites of Phyllidiella pustulosa, a Nudibranch from the South China Sea. J. Nat. Prod. 2004, 67, 1701–1704. [Google Scholar] [CrossRef] [PubMed]
- Dumdei, E.J.; Flowers, A.E.; Garson, M.J.; Moore, C.J. The Biosynthesis of Sesquiterpene Isocyanides and Isothiocyanates in the Marine Sponge Acanthella cavernosa (Dendy); Evidence for Dietary Transfer to the Dorid Nudibranch Phyllidiella pustulosa. Comp. Biochem. Physiol. A Physiol. 1997, 118, 1385–1392. [Google Scholar] [CrossRef]
- Jaisamut, S.; Prabpai, S.; Tancharoen, C.; Yuenyongsawad, S.; Hannongbua, S.; Kongsaeree, P.; Plubrukarn, A. Bridged Tricyclic Sesquiterpenes from the Tubercle Nudibranch Phyllidia coelestis Bergh. J. Nat. Prod. 2013, 76, 2158–2161. [Google Scholar] [CrossRef] [PubMed]
- Zhukova, N.V. Lipid Classes and Fatty Acid Composition of the Tropical Nudibranch Mollusks Chromodoris sp. and Phyllidia coelestis. Lipids 2007, 42, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N.; Wolstenholme, H.J.; Shinoda, K.; Asai, N.; Matsunaga, S.; Onuki, H.; Hirota, H. Two Sesquiterpene Isocyanides and a Sesquiterpene Thiocyanate from the Marine Sponge Acanthella cf. cavernosa and the Nudibranch Phyllidia ocellata. Tetrahedron Lett. 1992, 33, 6823–6826. [Google Scholar] [CrossRef]
- White, A.M.; Pierens, G.K.; Skinner-Adams, T.; Andrews, K.T.; Bernhardt, P.V.; Krenske, E.H.; Mollo, E.; Garson, M.J. Antimalarial Isocyano and Isothiocyanato Sesquiterpenes with Tri- and Bicyclic Skeletons from the Nudibranch Phyllidia ocellata. J. Nat. Prod. 2015, 78, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Fahrner, A.; Schrodl, M. Taxonomic Revision of the Common Indo-West Pacific Nudibranch Phyllidia varicosa Lamarck, 1801. Veliger 2000, 43, 164–171. [Google Scholar]
- Burreson, B.J.; Scheuer, P.J.; Finer, J.; Clardy, J. 9-Isocyanopupukeanane, a Marine Invertebrate Allomone with a New Sesquiterpene Skeleton. J. Am. Chem. Soc. 1975, 97, 4763–4764. [Google Scholar] [CrossRef]
- Hagadone, M.R.; Burreson, B.J.; Scheuer, P.J.; Finer, J.S.; Clardy, J. Defense Allomones of the Nudibranch Phyllidia varicosa Lamarck 1801. Helv. Chim. Acta 1979, 62, 2484–2494. [Google Scholar] [CrossRef]
- Fusetani, N.; Wolstenholme, H.J.; Matsunaga, S. Co-Occurrence of 9-Isocyanopupukeanane and Its C-9 Epimer in the Nudibranch Phyllidia bourguini. Tetrahedron Lett. 1990, 31, 5623–5624. [Google Scholar] [CrossRef]
- Gulavita, N.K.; de Silva, E.D.; Hagadone, M.R.; Karuso, P.; Scheuer, P.J.; van Duyne, G.D.; Clardy, J. Nitrogenous Bisabolene Sesquiterpenes from Marine Invertebrates. J. Org. Chem. 1986, 51, 5136–5139. [Google Scholar] [CrossRef]
- Yasman; Edrada, R.A.; Wray, V.; Proksch, P. New 9-Thiocyanatopupukeanane Sesquiterpenes from the Nudibranch Phyllidia varicosa and Its Sponge-Prey Axinyssa aculeata. J. Nat. Prod. 2003, 66, 1512–1514. [Google Scholar] [CrossRef]
- Wright, A.D. GC-MS and NMR analysis of Phyllidiella pustulosa and One of Its Dietary Sources, the Sponge Phakellia carduus. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2003, 134, 307–313. [Google Scholar] [CrossRef]
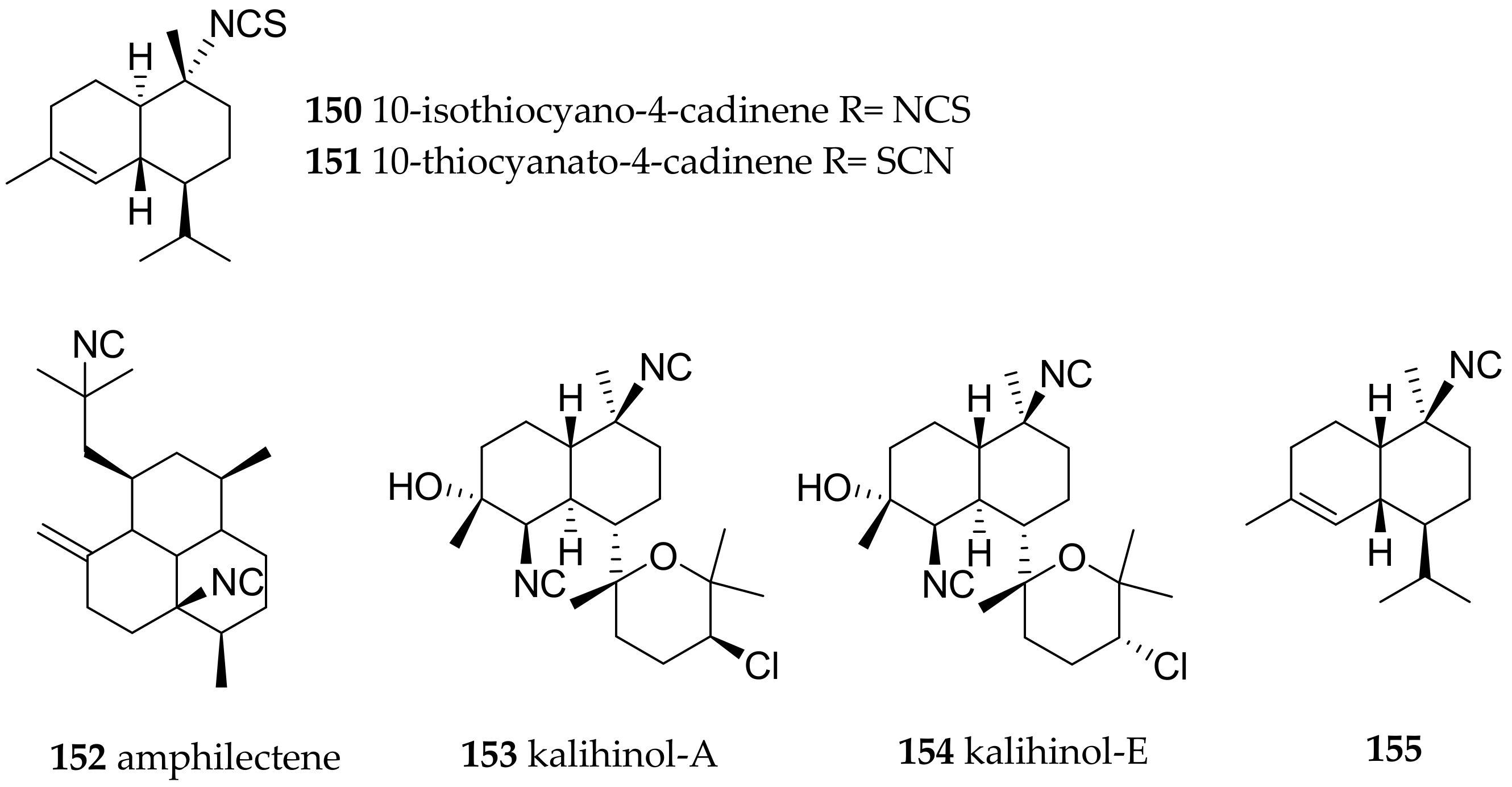
- Nishikawa, K.; Umezawa, T.; Garson, M.J.; Matsuda, F. Confirmation of the Configuration of 10-Isothiocyanato-4-cadinene Diastereomers through Synthesis. J. Nat. Prod. 2012, 75, 2232–2235. [Google Scholar] [CrossRef] [PubMed]
- Lyakhova, E.G.; Kolesnikova, S.A.; Kalinovskii, A.I.; Stonik, V.A. Secondary Metabolites of the Vietnamese Nudibranch Mollusk Phyllidiella pustulosa. Chem. Nat. Compd. 2010, 46, 534–538. [Google Scholar] [CrossRef]
- Jomori, T.; Shibutani, T.; Ahmadi, P.; Suzuka, T.; Tanaka, J. A New Isocyanosesquiterpene from the Nudibranch Phyllidiella pustulosa. Nat. Prod. Commun. 2015, 10, 1913–1914. [Google Scholar]
- Carte, B.; Faulkner, D. Role of Secondary Metabolites in Feeding Associations between a Predatory Nudibranch, 2 Grazing Nudibranchs, and a Bryozoan. J. Chem. Ecol. 1986, 12, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.; Lindquist, N.; Fenical, W. Chemical Defenses of the Tropical Ascidian Atapozoa sp. and Its Nudibranch Predators Nembrotha spp. Mar. Ecol. Prog. Ser. 1990, 59, 109–118. [Google Scholar] [CrossRef]
- Trindade-Silva, A.E.; Lim-Fong, G.E.; Sharp, K.H.; Haygood, M.G. Bryostatins: Biological Context and Biotechnological Prospects. Curr. Opin. Biotechnol. 2010, 21, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.K. The Biology of the Bryostatins in the Marine Bryozoan Bugula neritina. Ph. D. Thesis, University of California, San Diego, CA, USA, 1999. [Google Scholar]
- Lim, G.E. Bugula (Bryozoa) and Their Bacterial Symbionts: A Study in Symbiosis, Molecular Phylogenetics and Secondary Metabolism. Ph. D. Thesis, University of California, San Diego, CA, USA, 2004. [Google Scholar]
- Sudek, S.; Lopanik, N.B.; Waggoner, L.E.; Hildebrand, M.; Anderson, C.; Liu, H.; Patel, A.; Sherman, D.H.; Haygood, M.G. Identification of the Putative Bryostatin Polyketide Synthase Gene Cluster from “Candidatus endobugula sertula”, the Uncultivated Microbial Symbiont of the Marine Bryozoan Bugula neritina. J. Nat. Prod. 2007, 70, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Ciavatta, M.L.; Lefranc, F.; Carbone, M.; Mollo, E.; Gavagnin, M.; Betancourt, T.; Dasari, R.; Kornienko, A.; Kiss, R. Marine Mollusk-Derived Agents with Antiproliferative Activity as Promising Anticancer Agents to Overcome Chemotherapy Resistance. Med. Res. Rev. 2017, 37, 702–801. [Google Scholar] [CrossRef] [PubMed]
- Böhringer, N.; Fisch, K.M.; Schillo, D.; Bara, R.; Hertzer, C.; Grein, F.; Eisenbarth, J.-H.; Kaligis, F.; Schneider, T.; Wägele, H.; et al. Antimicrobial Potential of Bacteria Associated with Marine Sea Slugs from North Sulawesi, Indonesia. Front. Microbiol. 2017, 8, 1092. [Google Scholar] [CrossRef] [PubMed]
- Fisch, K.M.; Schäberle, T.F. Toolbox for Antibiotics Discovery from Microorganisms. Arch. Pharm. 2016, 349, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Krug, D.; Bozkurt, N.; Duddela, S.; Jansen, R.; Garcia, R.; Gerth, K.; Steinmetz, H.; Müller, R. Bacterial versus Chemical Diversity: The Taxonomy Paradigm in Microbial Natural Product Discovery. Nat. Commun. 2017. accepted. [Google Scholar]
- Hartmann, A.C.; Petras, D.; Quinn, R.A.; Protsyuk, I.; Archer, F.I.; Ransome, E.; Williams, G.J.; Bailey, B.A.; Vermeij, M.J.A.; Alexandrov, T.; et al. Meta-Mass Shift Chemical Profiling of Metabolomes from Coral Reefs. Proc. Natl. Acad. Sci. USA 2017, 114, 11685–11690. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.G.; Winters, A.E.; Cheney, K.L. Tropical Range Extension for the Temperate, Endemic South-Eastern Australian Nudibranch Goniobranchus splendidus (Angas, 1864). Diversity 2016, 8, 16. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clade | Family | Species | Locality | Depth and Abundance a | Bioactive Compounds | |
|---|---|---|---|---|---|---|
| Anaspidae (sea hares) | Aplysiidae Lamarck, 1809 | Stylocheilus striatus (Quoy and Gaimard, 1832) | Bunaken | 10 m; rare | see text and Table 2 | |
| Sacoglossa | Caliphyllidae Tiberi, 1881 | Cyerce sp. 4 (cf. Cyerce bourbonica Yonow, 2012) | Bunaken | 3–10 m; uncommon | nothing published | |
| Cyerce sp. 2 | Bunaken | 3–7 m; uncommon | nothing published | |||
| Oxynoidae Stoliczka, 1868 (1847) | Lobiger sp. 1 | Bunaken | 7 m; rare | nothing published | ||
| Lobiger viridis (Pease, 1863) | Bunaken | 8 m; rare | nothing published | |||
| Plakobranchidae Gray, 1840 | Elysia asbecki Wägele, Stemmer, (Burghardt and Händeler, 2010) | Bunaken, Manado Tua, Siladen, Tiwoho | 4–15 m; uncommon | nothing published | ||
| 3 undescribed Elysia species | Bunaken, Siladen | 5–9 m; rare | nothing published | |||
| Thuridilla albopustulosa (Gosliner, 1995) | Bunaken | 6 m; rare | nothing published | |||
| Thuridilla flavomaculata (Gosliner, 1995) | Bunaken | 2–7 m; rare | nothing published | |||
| Thuridilla gracilis (Risbec, 1928) | Bunaken, Siladen | 3–8 m; uncommon | nothing published | |||
| Thuridilla lineolata (Bergh, 1905) | Bunaken, Siladen, Tiwoho | 1–9 m; abundant | nothing published | |||
| Cephalaspidea | Aglajidae Pilsbry, 1895 (1847) | unidentified specimen | Tiwoho | 5 m; rare | nothing published | |
| Chelidonura amoena (Bergh, 1905) | Bunaken | 1 m; rare | nothing published | |||
| Chelidonura hirundinina (Quoy and Gaimard, 1833) | Bunaken, Manado Tua | 5 m; rare | nothing published | |||
| Odontoglaja guamensis (Rudman, 1978) | Bunaken, Manado Tua, Tiwoho | 3–19 m; uncommon | nothing published | |||
| Diaphanidae Odhner, 1914 (1857) | Colpodaspis thompsoni (G. H. Brown, 1979) | Bunaken, Manado Tua, Siladen, Tiwoho | 4–11 m; uncommon | nothing published | ||
| Gastropteridae Swainson, 1840 | Sagaminopteron psychedelicum (Carlson and Hoff, 1974) | Bunaken, Manado Tua | 4–15 m; uncommon | see text and Table 2 | ||
| Siphopteron brunneomarginatum (Carlson and Hoff, 1974) | Bunaken, Manado Tua, Siladen | 4–10 m; uncommon | nothing published | |||
| Siphopteron cf. ladrones (Carlson and Hoff, 1974) | Bunaken | 5 m; rare | nothing published | |||
| Siphopteron nigromarginatum (Gosliner, 1989) | Manado Tua | 5 m; rare | nothing published | |||
| Siphopteron spec. | Bunaken | 4–5 m; rare | nothing published | |||
| Siphopteron tigrinum (Gosliner, 1989) | Bunaken | 5–6 m; rare | nothing published | |||
| Haminoeidae Pilsbry, 1895 | 2 undescribed Haminoea species | Bunaken, Manado Tua, Siladen | 3–13 m; rare | nothing published | ||
| Pleurobranchomorpha | Pleurobranchidae Gray, 1827 | Pleurobranchus forskalii (Rüppell and Leuckart, 1828) | Bunaken, Siladen | 4–8 m; common | see text and text and Table 2 | |
| Nudibranchia | Anthobranchia | Aegiridae P. Fischer, 1883 | Notodoris serenae (Gosliner and Behrens, 1997) | Manado Tua | 13 m; rare | nothing published |
| Chromodorididae Bergh, 1891 | Ceratosoma sp. 2 | Bunaken | 5–8 m; rare | nothing published | ||
| Chromodoris annae (Bergh, 1877) | Bunaken, Manado Tua, Siladen, Tiwoho | 4–23 m; abundant | nothing published | |||
| Chromodoris cf. boucheti (Rudman, 1982) | Siladen | 8 m; rare | nothing published | |||
| Chromodoris dianae (Gosliner and Behrens, 1998) | Bunaken, Manado Tua, Siladen, Tiwoho | 4–21 m; abundant | nothing published | |||
| Chromodoris lochi (Rudman, 1982) | Bunaken, Manado Tua, Siladen | 5–17 m; common | see text and Table 2 | |||
| Chromodoris strigata (Rudman, 1982) | Siladen | 11 m; rare | nothing published | |||
| Chromodoris willani (Rudman, 1982) | Bunaken, Manado Tua, Siladen | 7–21 m; common | nothing published | |||
| Doriprismatica (=Glossodoris) stellata (Rudman, 1986) | Bunaken | 4–21 m; uncommon | nothing published | |||
| Glossodoris (=Casella) cincta (Bergh, 1888) | Tiwoho | 6 m; rare | nothing published | |||
| Goniobranchus geometricus (Risbec, 1928) | Bunaken, Tiwoho | 4–8 m; rare | nothing published | |||
| Goniobranchus reticulatus (Quoy and Gaimard, 1832) | Manado Tua | 15 m; rare | nothing published | |||
| Hypselodoris maculosa (Pease, 1871) | Bunaken, Tiwoho | 4–6 m; rare | nothing published | |||
| Dendrodorididae O’Donoghue, 1924 (1864) | Dendrodoris albobrunnea (Allan, 1933) | Bunaken | 4 m; rare | nothing published | ||
| Dendrodoris nigra (Stimpson, 1855) | Bunaken | 4 m; rare | see text and Table 2 | |||
| Discodorididae Bergh, 1891 | Taringa halgerda (Gosliner and Behrens, 1998) | Bunaken, Tiwoho | 6 m; rare | nothing published | ||
| Halgerda carlsoni (Rudman, 1978) | Manado Tua | 5 m; rare | nothing published | |||
| Halgerda tessellata (Bergh, 1880) | Siladen | 5 m; rare | nothing published | |||
| Rostanga sp. 4 | Manado Tua | 13m; rare | nothing published | |||
| Hexabranchidae Bergh, 1891 | Hexabranchus sanguineus (Rüppell and Leuckart, 1830) | Bunaken | 2 m; rare | see text and Table 2 | ||
| Goniodorididae H. Adams and A. Adams, 1854 | Trapania euryeia (Gosliner and Fahay, 2008) | Bunaken | 6 m; rare | nothing published | ||
| Gymnodorididae Odhner, 1941 | Gymnodoris sp. | Bunaken, Manado Tua | 5–7 m; rare | nothing published | ||
| Phyllidiidae Rafinesque, 1814 | Phyllidia coelestis (Bergh, 1905) | Bunaken, Manado Tua, Tiwoho | 2–15 m | see text and Table 2 | ||
| Phyllidia elegans (Bergh, 1869) | Bunaken, Siladen | 2–19 m; uncommon | nothing published | |||
| Phyllidia ocellata (Cuvier, 1804) | Tiwoho | 5 m; rare | see text and Table 2 | |||
| Phyllidia varicosa (Lamarck, 1801) | Bunaken | 4–21 m; uncommon | see text and Table 2 | |||
| Phyllidiella annulata (Gray, 1853) | Bunaken, Manado Tua | 11–13 m; rare | nothing published | |||
| Phyllidiella pustulosa (Cuvier, 1804) | Bunaken, Manado Tua, Siladen, Tiwoho | 5–19 | see text and Table 2 | |||
| Phyllidiella striata (Bergh, 1889) | Bunaken | 15 m; rare | nothing published | |||
| Phyllidiopsis pipeki (Brunckhorst, 1993) | Bunaken | 14–15 m; rare | nothing published | |||
| Phyllidiopsis sphingis (Brunckhorst, 1993) | Manado Tua | 19 m; rare | nothing published | |||
| Polyceridae Alder and Hancock, 1845 | Nembrotha cristata (Bergh, 1877) | Bunaken, Siladen | 4–15 m; rare | see text and Table 2 | ||
| Nembrotha kubaryana (Bergh, 1877) | Tiwoho | 6 m; rare | see text and Table 2 | |||
| Kaloplocamus dokte (Vallès and Gosliner, 2006) | Bunaken | 7 m; rare | nothing published | |||
| Polycera japonica (Baba, 1949) | Bunaken | 7–8 m; rare | nothing published | |||
| Polycera risbeci (Odhner, 1941) | Bunaken | 7–8 m; rare | nothing published | |||
| Nudibranchia | Subclade Cladobranchia | Arminidae Iredale and O’Donoghue, 1923 (1841) | Dermatobranchus fasciatus (Gosliner and Fahey, 2011) | Bunaken | 7 m; rare | nothing published |
| Dermatobranchus striatus (van Hasselt, 1824) | Manado Tua | 7 m; rare | nothing published | |||
| Eubranchidae Odhner, 1934 | Eubranchus sp. 4 | Bunaken | 18 m; rare | nothing published | ||
| Facelinidae Bergh, 1889 | Caloria indica (Bergh, 1896) | Bunaken, Manado Tua, Siladen, Tiwoho | 3–6 m; uncommon | nothing published | ||
| Favorinus japonicus (Baba, 1949) | Bunaken, Siladen | 5–10 m; uncommon | nothing published | |||
| Favorinus mirabilis (Baba, 1955) | Bunaken | 23 m; rare | nothing published | |||
| Favorinus tsuruganus (Baba and Abe, 1964) | Bunaken | 6–23 m; uncommon | nothing published | |||
| Noumeaella sp. No. 1–2 | Bunaken | 4–12 m; uncommon | nothing published | |||
| Phyllodesmium briareum (Bergh, 1896) | Bunaken, Tiwoho | 2–7 m; abundant | see text and Table 2 | |||
| Phyllodesmium poindimiei (Risbec, 1928) | Bunaken | 17 m; rare | nothing published | |||
| Facelina rhodopos (Yonow, 2000) | Manado Tua | 15 m; rare | nothing published | |||
| Pteraeolidia semperi (Bergh, 1870) | Bunaken, Manado Tua, Siladen, Tiwoho | 4–15 m; common | nothing published | |||
| Flabellinidae Bergh, 1889 | Flabellina bicolor (Kelaart, 1858) | Siladen, Tiwoho | 3–6 m; rare | nothing published | ||
| Flabellina exoptata (Gosliner and Willan, 1991) | Bunaken, Siladen | 5–8 m; uncommon | see text and Table 2 | |||
| Flabellina rubrolineata (O’Donoghue, 1929) | Bunaken | 6 m; rare | nothing published | |||
| Proctonotidae Gray, 1853 | Janolus sp. (sp. 11 Gosliner et al., 2015: 308) | Bunaken | 7 m; rare | nothing published |
| Clade | Family | Species | Location | Chemistry | (Presumable) Origin of Compound | References |
|---|---|---|---|---|---|---|
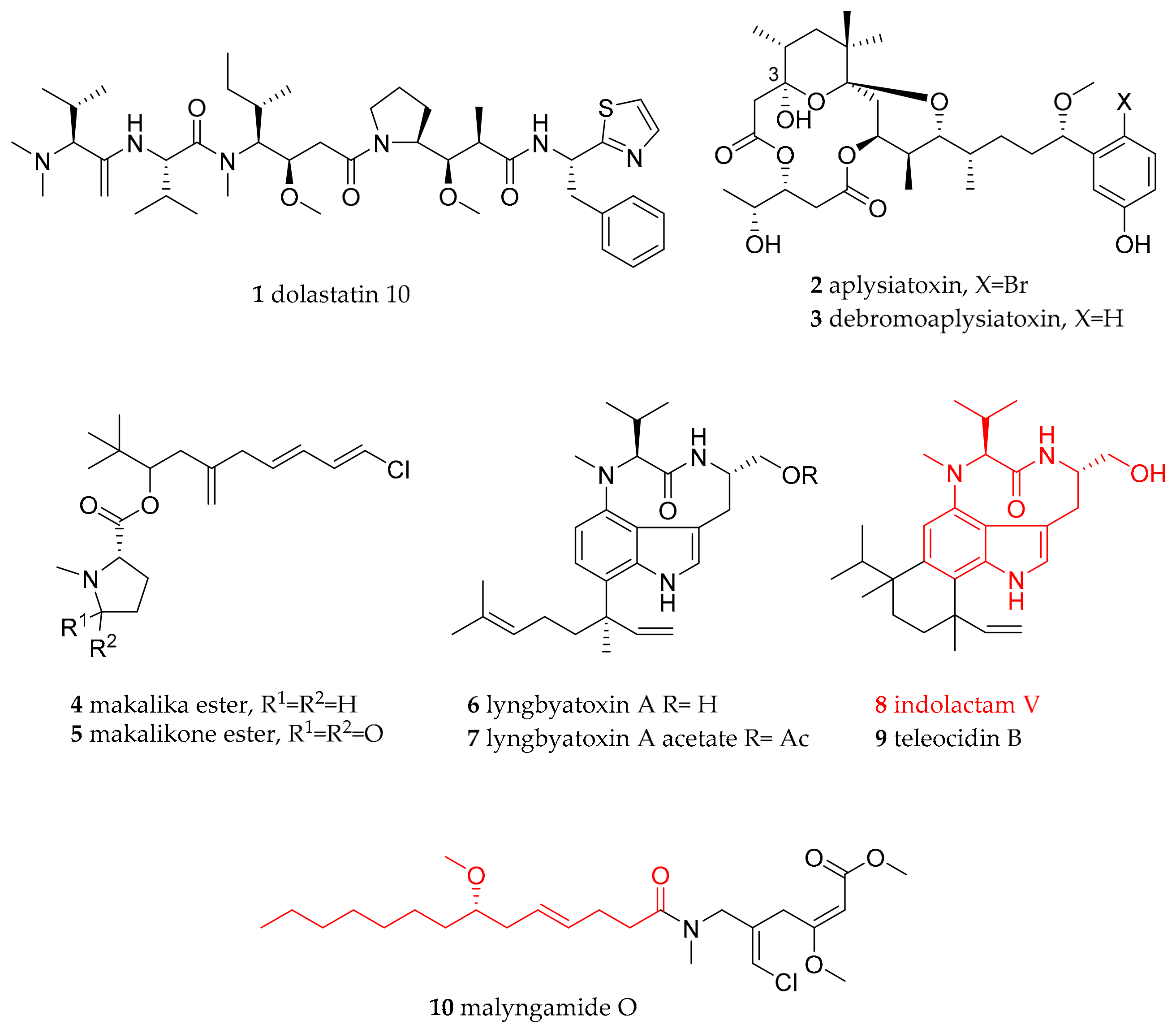
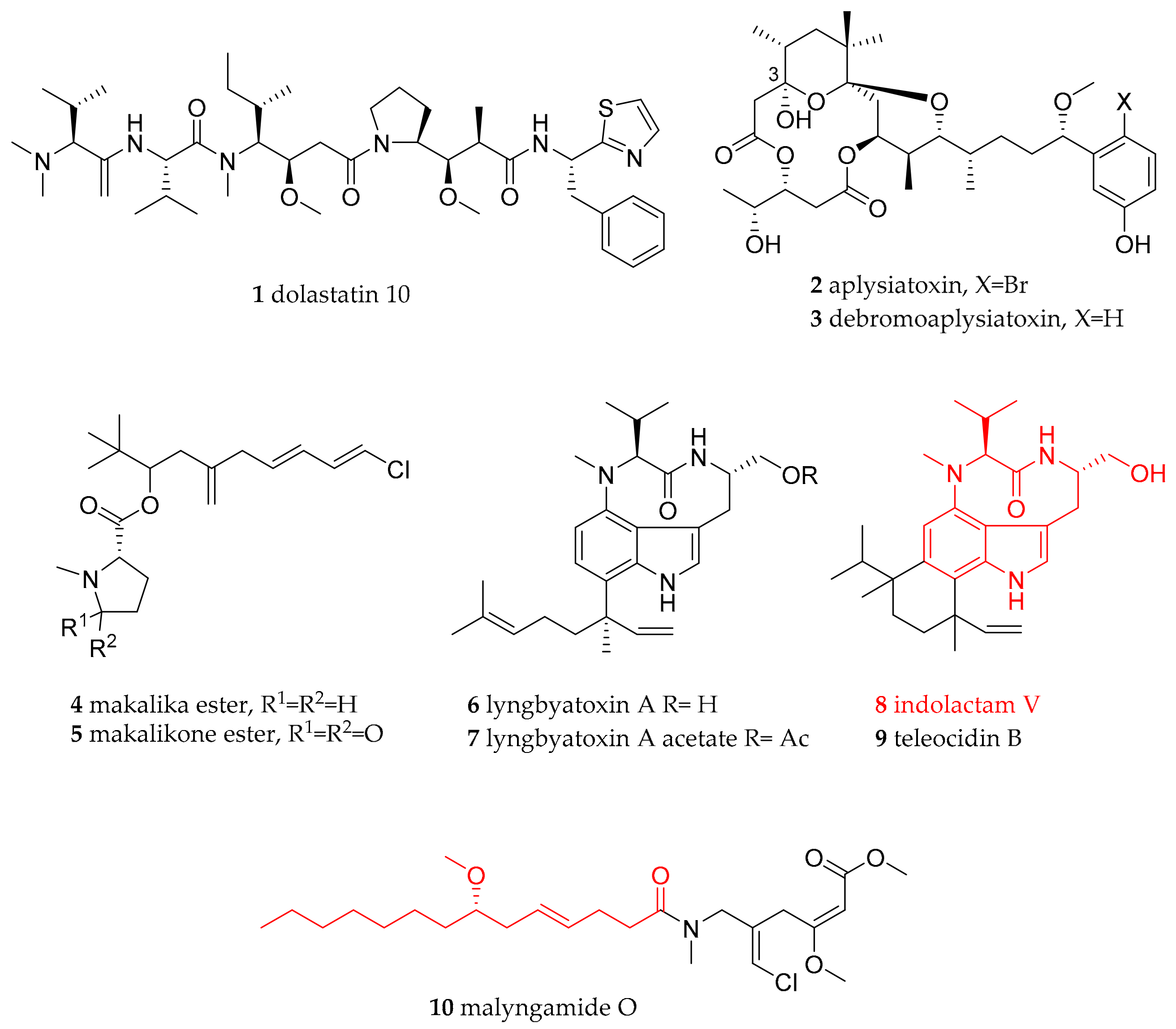
| Anaspidae (sea hares) | Aplysiidae Lamarck, 1809 | Dolabella auricularia (Lightfoot, 1786) | Western Indian Ocean (Mauritius) | dolastatin 10 (1) | dietary (cyanobacteria) | [21] |
| Stylocheilus striatus (Quoy and Gaimard, 1832) | Australia (feeding study) | debromoaplysiatoxin (3), lyngbyatoxin A (6), | dietary (cyanobacteria Lyngbya majuscula) | [22,23,24,25,26,32,37] | ||
| Stylocheilus longicauda (Quoy and Gaimard, 1825) | not given, presumably Hawaii | aplysiatoxin (2), debromoaplysiatoxin (3) | dietary (cyanobacteria) | [22,23,24,25,26] | ||
| Stylocheilus longicauda (Quoy and Gaimard, 1825) | Black Point, Oahu, Hawaii | makalika ester (4), makalikone ester (5), lyngbyatoxin A acetate (7), malyngamide O (10) | dietary (cyanobacteria) | [27,28,29,30,31,35] | ||
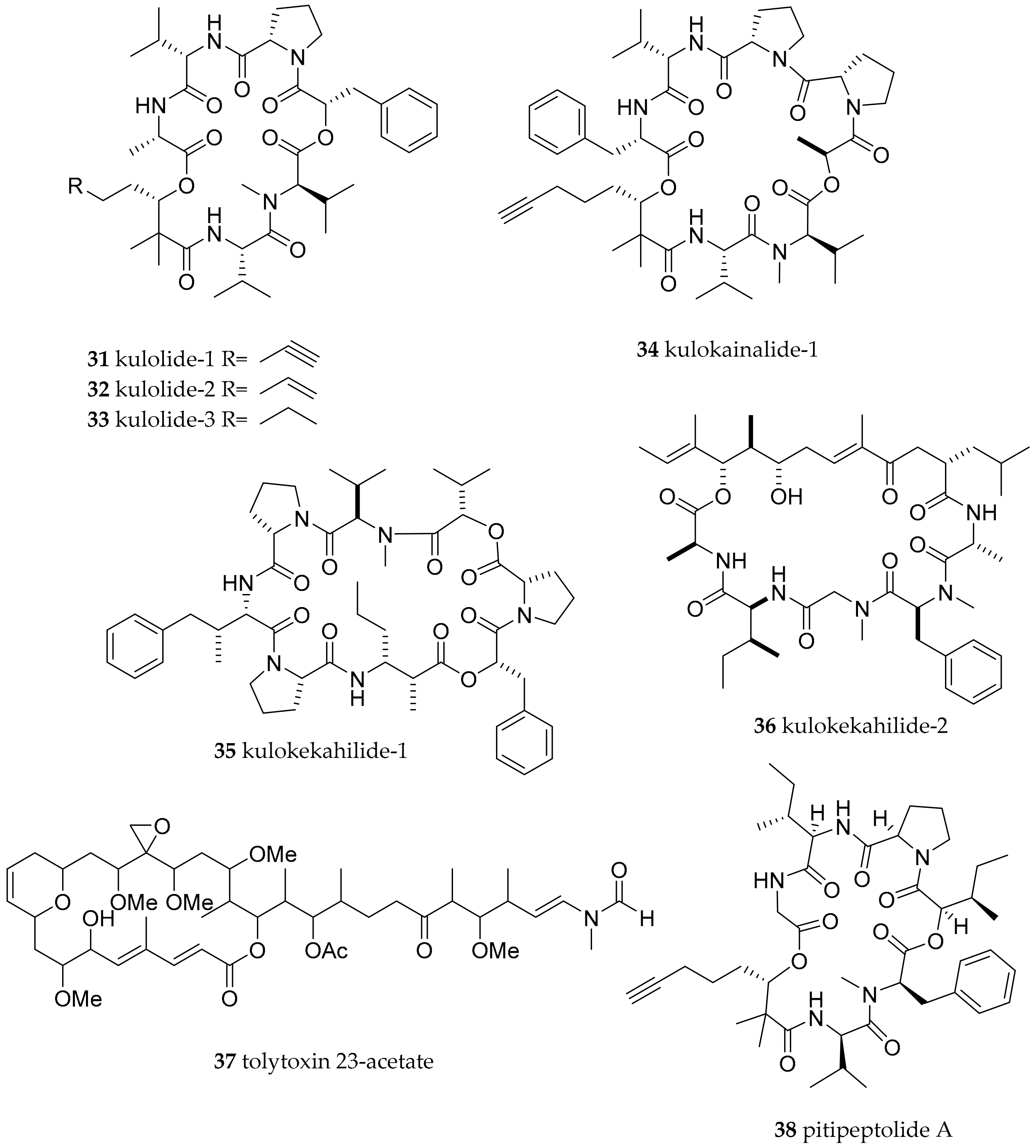
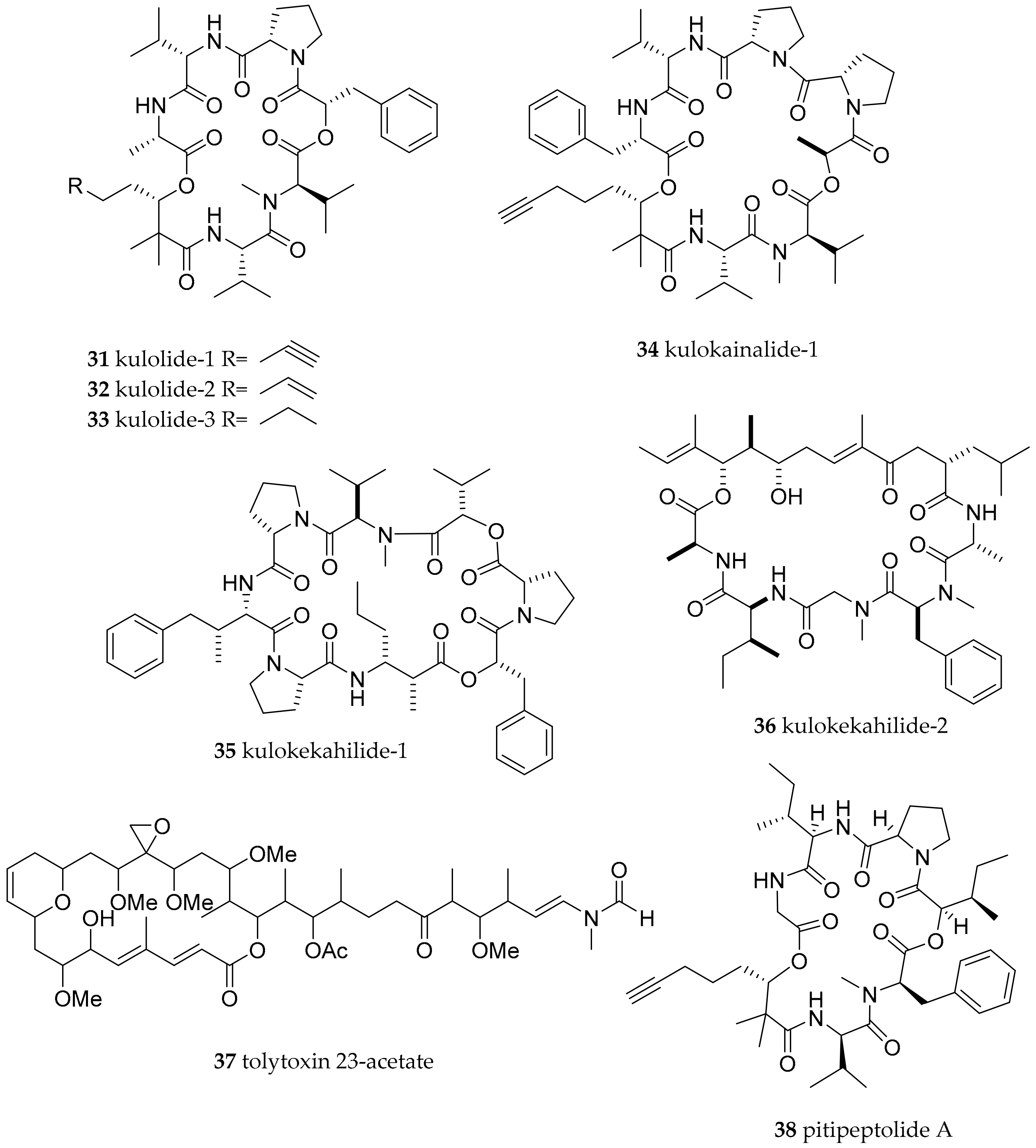
| Stylocheilus longicauda (Quoy and Gaimard, 1825) | not given, presumably Hawaii | kulolide-1 (31) | dietary (cyanobacteria) | [63] | ||
| Bursatella leachii (Blainville, 1817) | Australia (feeding study) | lyngbyatoxin A (6) | dietary (cyanobacteria) | [37] | ||
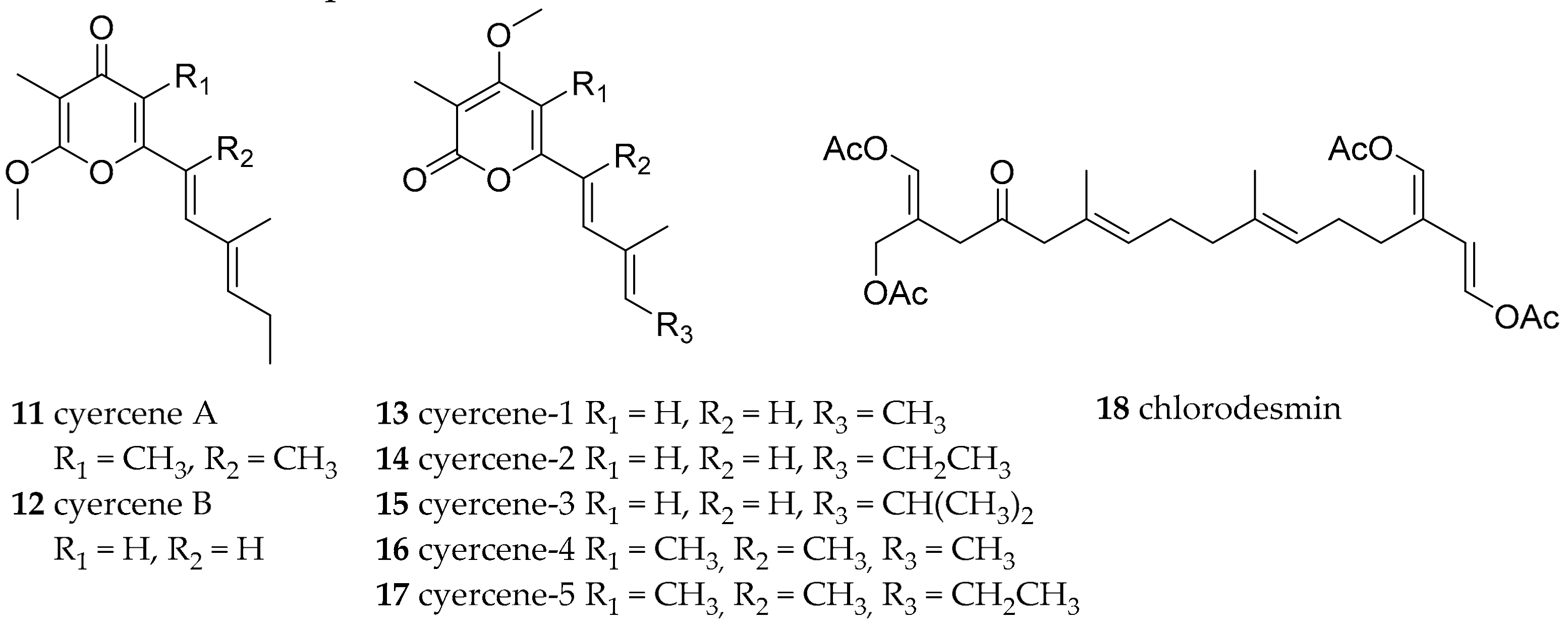
| Sacoglossa | Caliphyllidae Tiberi, 1881 | Cyerce cristallina (Trinchese, 1881) | Mediterranean Sea | cyercene A 11) and B (12) and 1–5 (13–17) | de novo | [2,38] |
| Cyerce nigricans (Pease, 1866) | Lizard Island (Australia) | chlorodesmin (18) | diatery (green alga Chlorodesmis fastigiata) | [39,40] | ||
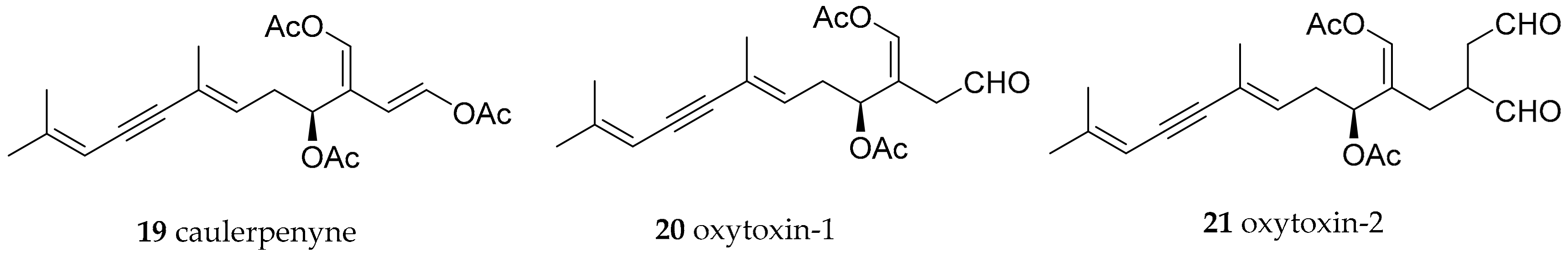
| Oxynoidae Stoliczka, 1868 (1847) | Lobiger serradifalci (Calcara, 1840) | Capo Miseno (Bay of Naples, Italy) | oxytoxin-1 (20) | modified from dietary caulerpynene (19) from green alga Caulerpa prolifera | [41,42,43] | |
| Oxynoe olivacea (Rafinesque, 1814) | Murcia (SE Spain) | oxytoxin-1 (20), oxytoxin-2 (21) | modified from dietary caulerpynene (19) from green alga Caulerpa prolifera | [41,42,43] | ||
| Oxynoe olivacea (Rafinesque, 1814) | Bay of Naples, Italy | oxytoxin-1 (20), oxytoxin-2 (21) | modified from dietary caulerpynene (19) from green alga Caulerpa prolifera | [41,42,43] | ||
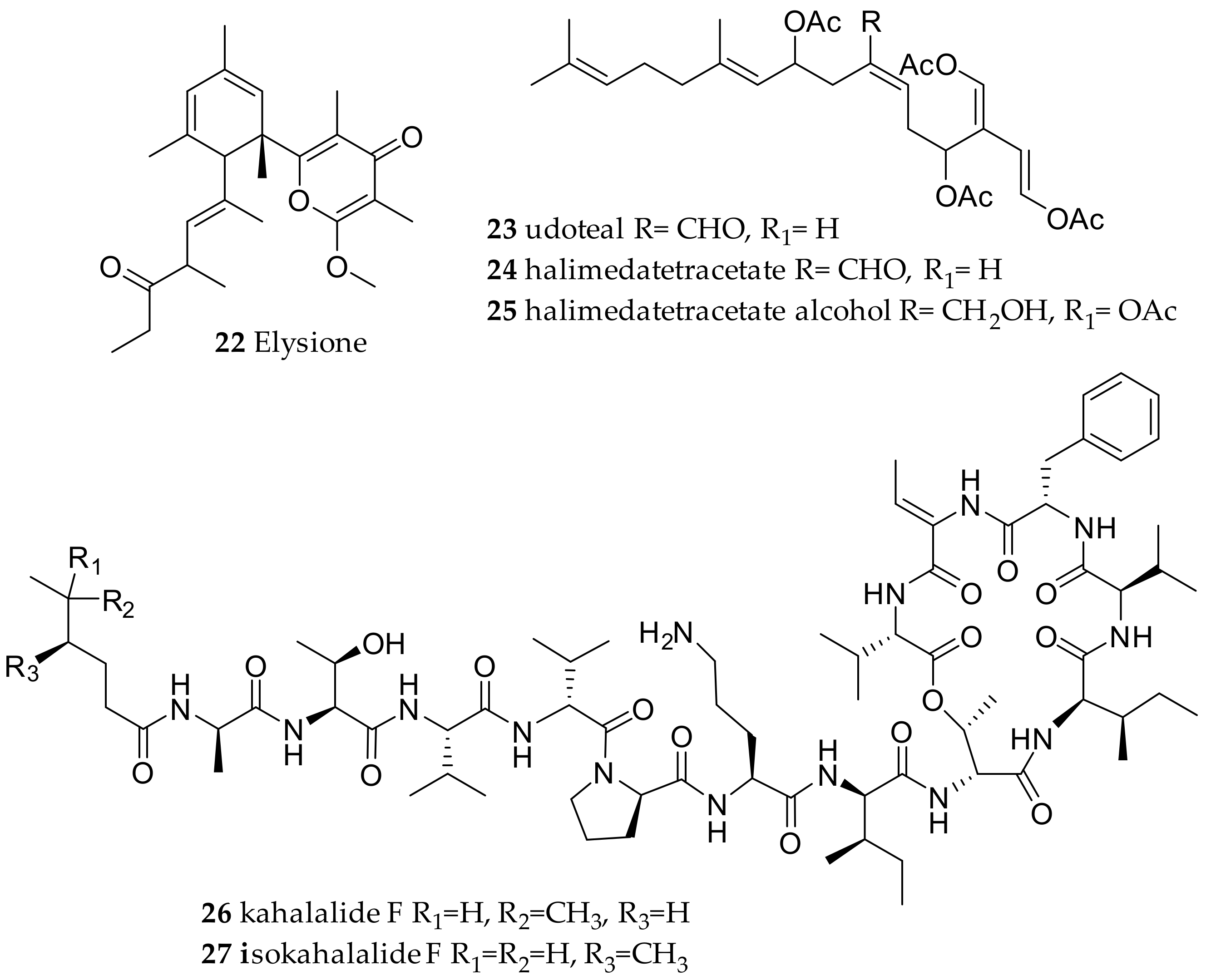
| Plakobranchidae Gray, 1840 | Elysia chlorotica (Gould, 1870) | Bay of Fundy, Canada | elysione (22) | de novo | [3,51] | |
| Elysia viridis (Montagu, 1804) | Fusaro Lake, Bay of Naples, Italy | elysione (22) | de novo | [3,51] | ||
| Elysia translucens (Pruvot-Fol, 1957) | Capo Miseno, Bay of Naples, Italy | udoteal (23) | dietary from the green alga Udotea petiolata | [3,40] | ||
| Elysia halimedae (Macnae 1954) (accepted as Elysia pusilla (Bergh, 1871)) | Agat Bay, Guam | halimedatetracetate alcohol (25) | dietary (chemical modification of halimedatetraacetate (24) from Halimeda mucroloba Decaisne (Carlson and Hoff, 1978)) | [52] | ||
| Elysia rufescens (Pease, 1871) | Hawaii | kahalalide F (26) and isokahalalide F (27) | dietary (algae Bryopsis pennata; might be of bacterial origin, with Mycoplasma spp. and Vibrio spp. as possible producers) | [46,53,55,57] | ||
| Elysia ornata (Swainson, 1840) | coasts of Okha (India) | kahalalide F (26) and other kalahalides | dietary (algae Bryopsis pennata; might be of bacterial origin, with Mycoplasma spp. and Vibrio spp. as possible producers) | [46,48] | ||
| Elysia grandifolia (Kelaart, 1858) | Gulf of Mannar and Palk Bay, Rameswaram, India at 1 to 2 m depth. | kahalalide F (26) and other kalahalides | dietary (slugs were feeding on algae Bryopsis plumosa (Hudson); might be of bacterial origin, with Mycoplasma spp. and Vibrio spp. as possible producers) | [46,54] | ||
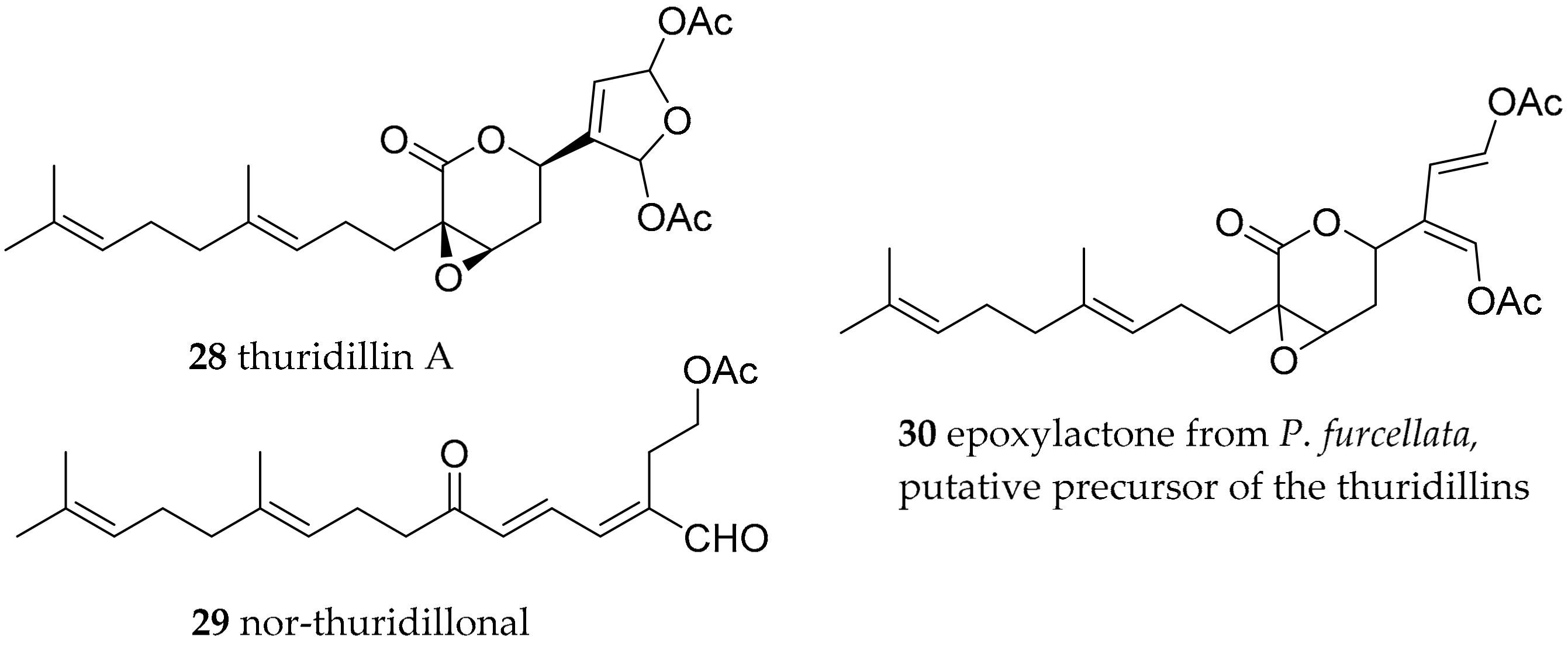
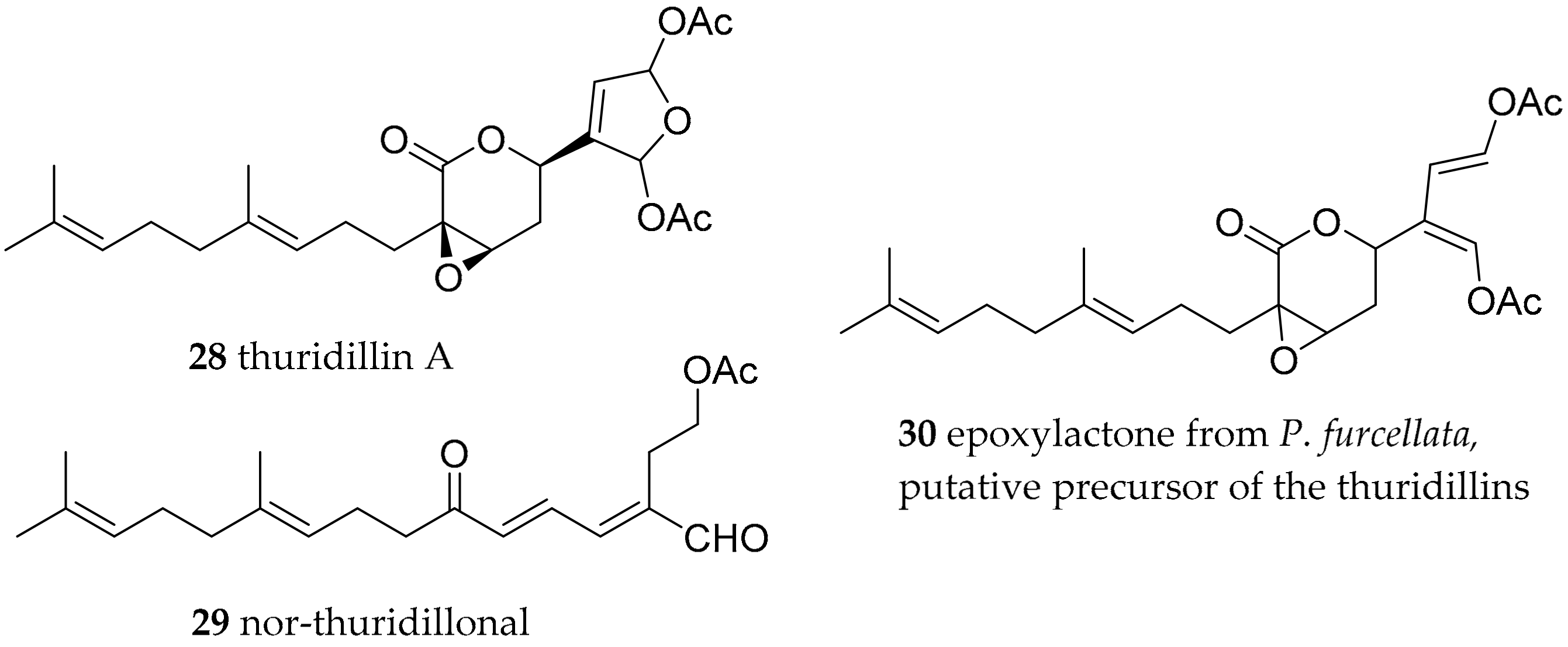
| Thuridilla hopei (Vérany, 1853) | Italy | thuridillins, e.g., thuridillin A (28) | de novo, with precursor derived from algae Pseudochlorodesmis furcellata (30) | [58,60] | ||
| Thuridilla splendens (Baba, 1949) | Australia | thuridillins | de novo, with precursor derived from algae P. furcellata 30) | [59,60] | ||
| Thuridilla hopei (Vérany, 1853) | Italy | thuridillin-related aldehydes, e.g., nor-thuridillinal (29) | de novo, with precursor derived from algae P. furcellata (30) | [58,60] | ||
| Cephalaspidea | Aglajidae Pilsbry, 1895 (1847) | Philinopsis speciosa (Pease, 1860) | Hawaii | Kulolide-1 (31), kulolide-2 (32), kulolide-3 (33), kulokainalide-1 (34), kulokahilide-1 (35), kulokahilide-1 (36) as well as tolytoxin 23-acetate (37) | dietary (cyanobacteria; transfer most likely mediated via herbivorous molluscs like Stylocheilus longicauda and Dolabella auricularia which in turn were readily accepted by P. speciosa in feeding experiment) | [61,62,63,64] |
| Navanax inermis (J. G. Cooper, 1862) | Pacific | navenones A-C (39-41), isopulo’upone (42), 5,6-dehydroaglajne-3 (43) | dietary (Bulla species, e.g., Bulla gouldiana) | [67,68,70] | ||
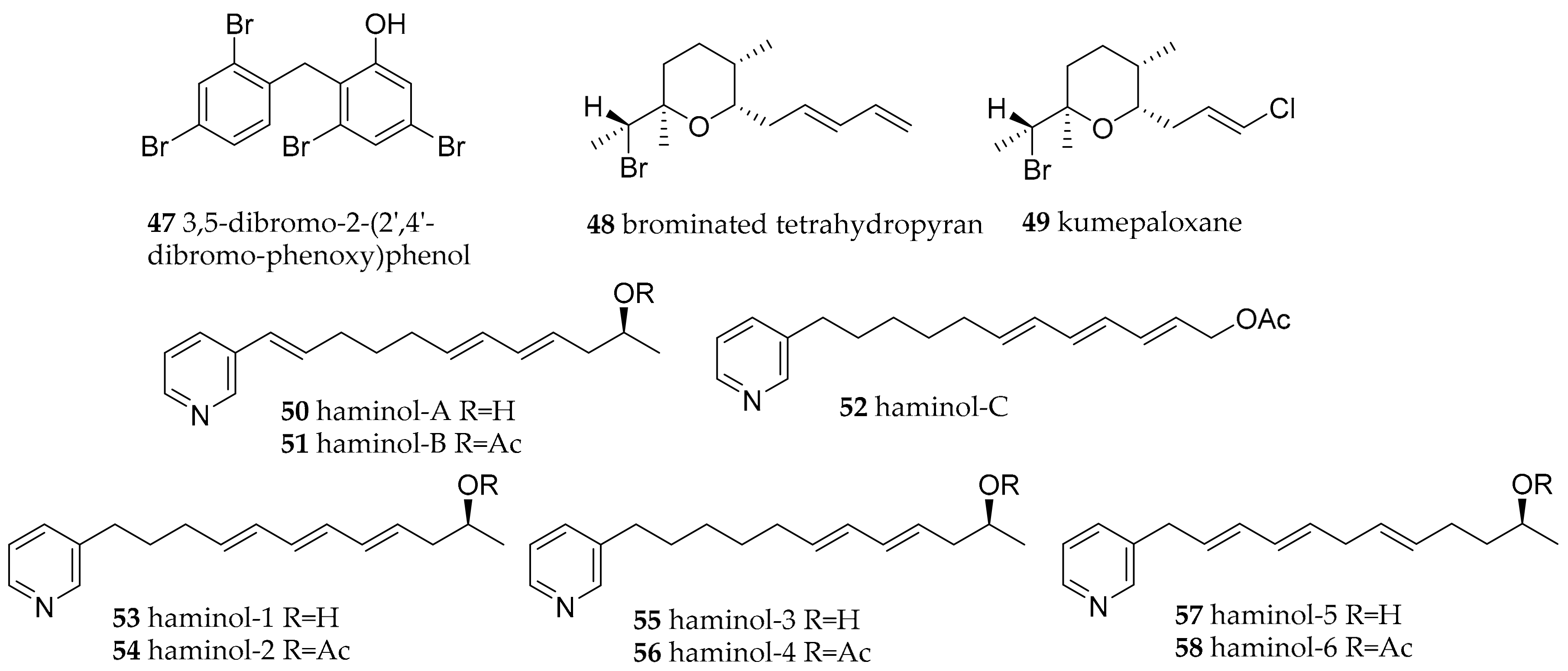
| Gastropteridae Swainson, 1840 | Sagaminopteron psychedelicum (Carlson and Hoff, 1974) | Guam | 3,5 dibromo-2-(2′,4′-dibromo-phenoxy)phenol (47) | dietary (sponge Dysidea granulosa) | [71,72] | |
| Sagaminopteron nigropunctatum (Carlson and Hoff, 1973) | Guam | 3,5 dibromo-2-(2′,4′-dibromo-phenoxy)phenol (47) | dietary (sponge Dysidea granulosa) | [71,73] | ||
| Haminoeidae Pilsbry, 1895 | Haminoea cyanomarginata (Heller and Thompson, 1983) | Gulf of Corinth (Greece) | brominated tetrahydropyran (48) | dietary (Western Australian sponge Haliclona sp. Grant, 1841) | [73] | |
| Haminoea cymbalum (Quoy and Gaimard, 1832) | Indian coasts | brominated tetrahydropyran (48) | dietary (sponge) | [73] | ||
| Haminoea cymbalum (Quoy and Gaimard, 1832) | Guam | kumepaloxane (49) | dietary (sponge) | [74] | ||
| Haminoea species | Naples (Italy) | haminol A–C (50–52), and haminol 1–6 (53–58 | de novo | [75,76] | ||
| Haminoea fusari (Alvarez, Garcia and Villani, 1993) | Naples (Italy) | polypropionates, haminol 1–6 (53–58) | de novo | [77] | ||
| Haminoea orbignyana (Férussac, 1822) | Naples (Italy) | haminol 1 and 2 (53–54) | de novo synthesis, shown by feeding study | [4] | ||
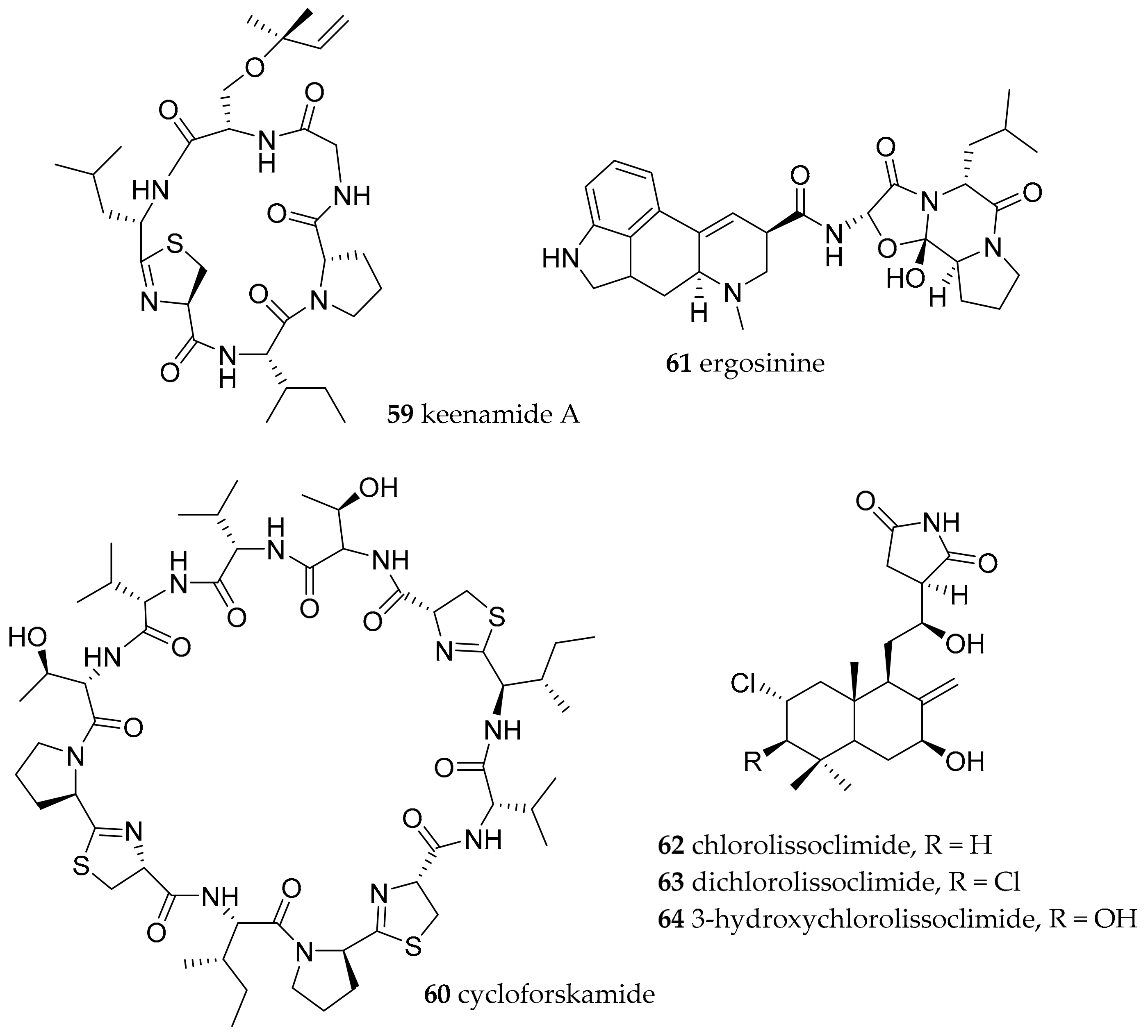
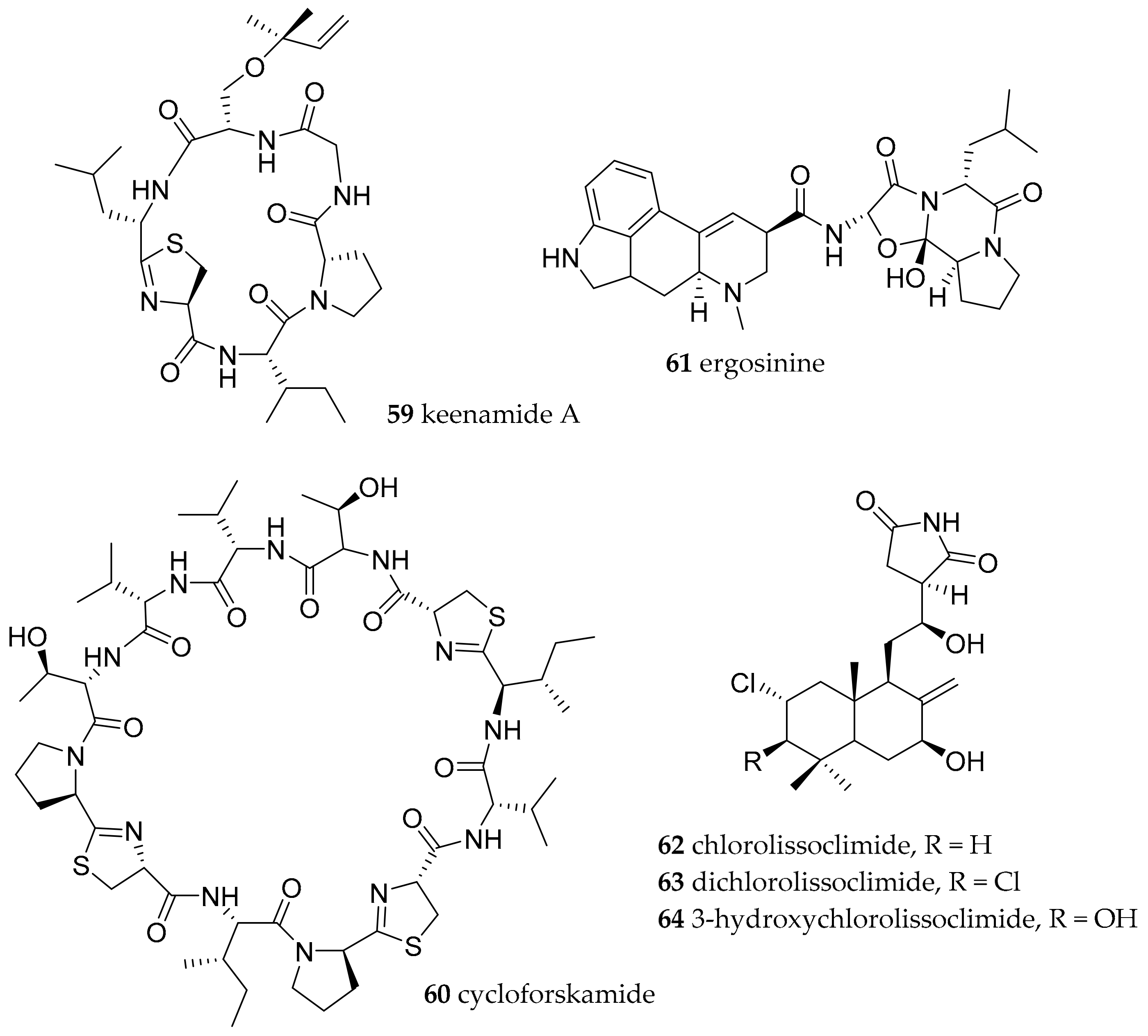
| Pleurobranchomorpha | Pleurobranchidae Gray, 1827 | Pleurobranchus forskalii (Rüppell and Leuckart, 1828) | Manado, Indonesia | keenamide A (59) | dietary (presumable cyanobacterial origin) | [78] |
| Pleurobranchus forskalii (Rüppell and Leuckart, 1828) | Ishigaki Island, Japan | cycloforskamide (60) | dietary (sponge with associated cyanobacteria) or symbiotic cyanobacteria | [79] | ||
| Pleurobranchus forskalii (Rüppell and Leuckart, 1828) | Ishigaki Island, Japan | ergosinine (61) | dietary (ascidian and/or endophytic fungi) | [81] | ||
| Pleurobranchus forskalii (Rüppell and Leuckart, 1828) | Philippines | chlorolissoclimide (62) and dichlorolissoclimide 63) | dietary (Lissoclinum species of ascidian) | [83,84] | ||
| Pleurobranchus. albiguttatus (Bergh, 1905) | Philippines | chlorolissoclimide (62), dichlorolissoclimide (63) and 3β-hydroxychlorolissoclimide (64) | dietary (Lissoclinum species of ascidian) | [83,84] | ||
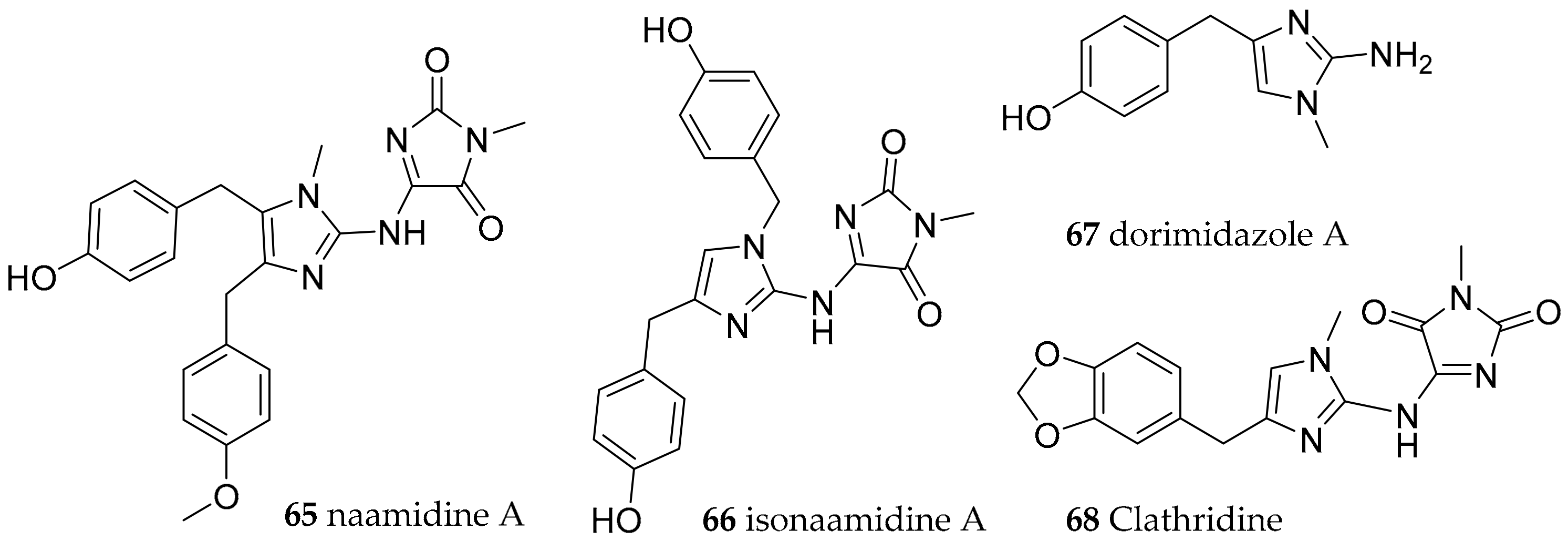
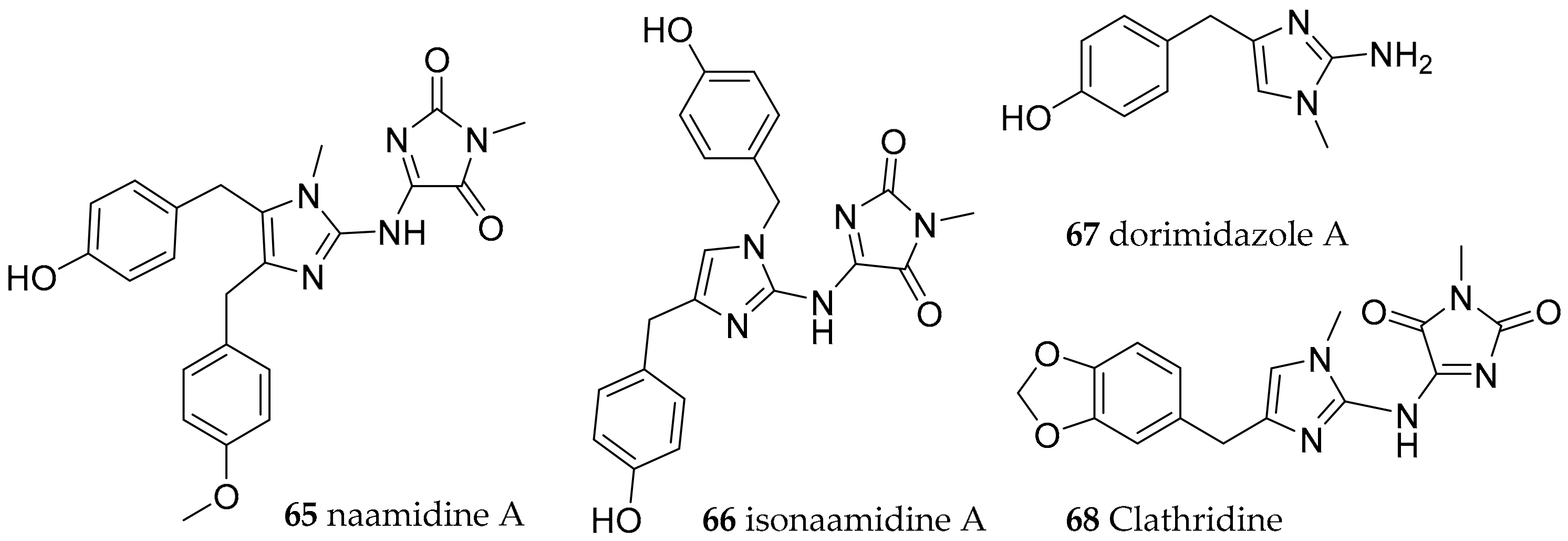
| Nudibranchia Anthobranchia | Aegiridae P. Fischer, 1883 | Notodoris citrina (Bergh, 1875) | Gulf of Eilat, The Red Sea | naamidine A (65), isonaamidine-A (66) | dietary (sponge Leucetta chagosensis Dendy, 1913) | [85,86,87,88] |
| Notodoris gardineri (Eliot, 1906) | Philippines | isonaamidine-A (66), dorimidazole-A (67) | [88,89] | |||
| Notodoris gardineri (Eliot, 1906) | Great Barrier Reef | clathridine (68) | dietary (sponge) | [90,92,93] | ||
| Notodoris gardineri (Eliot, 1906) | Papua New Guinea | clathridine (68) | dietary (sponge Clathrina clathrus Schmidt, 1864) | [91,92,93] | ||
| Chromodorididae Bergh, 1891 | Ceratosoma amoenum (Cheeseman, 1886) | Great Barrier Reef | allolaurinterol (69) | dietary (origin could be red algae, e.g., of the genus Laurencia; 69 also found in cyanobacteria, via herbivorous sea slugs) | [94,95,96] | |
| Ceratosoma trilobatum (J.E. Gray, 1827) | South China Sea Coast | (−)-furodysinin (70) | dietary (sponge) | [97] | ||
| Ceratosoma gracillimum (Semper in Bergh, 1876) | South China Sea Coast | (−)-furodysinin (70) | dietary (sponge) | [97] | ||
| Ceratosoma gracillimum (Semper in Bergh, 1876) | South Coast of Hainan Island | (−)-furodysinin (70), nakafuran-9 (71) | dietary (sponge) | [97] | ||
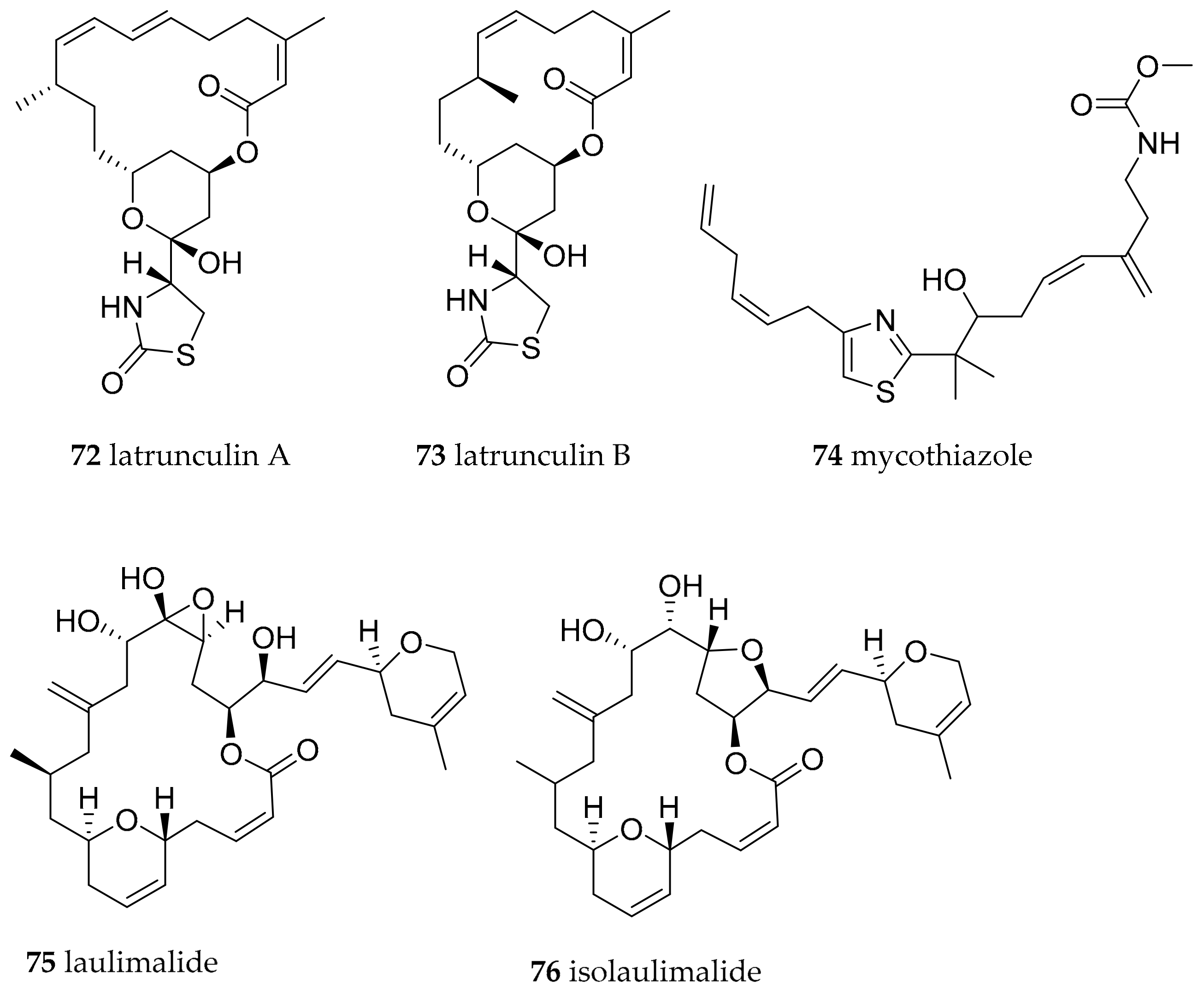
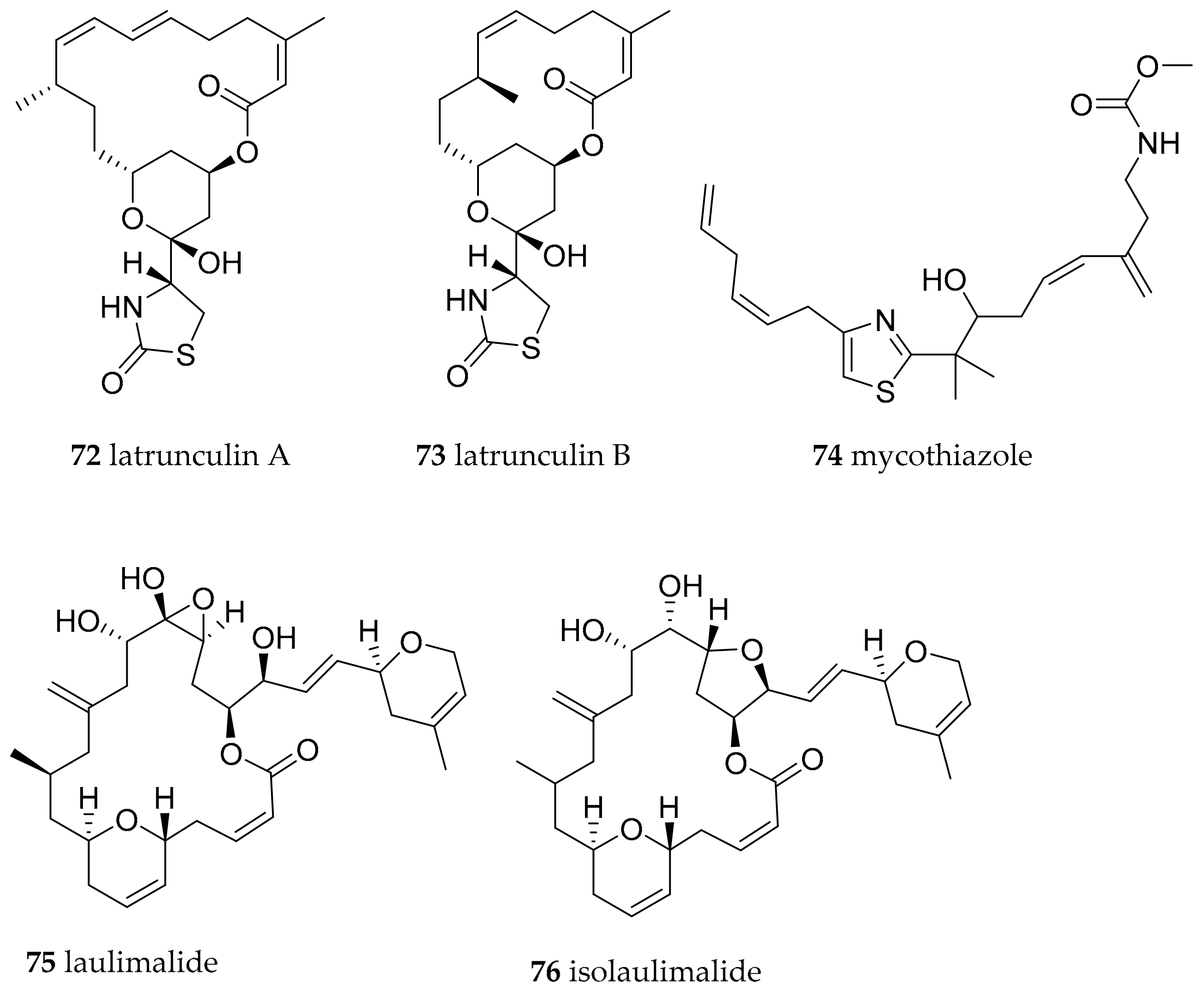
| Chromodoris lochi (Rudman, 1982) | Fiji | latrunculin A (72) | dietary, 72 in Spongia (=Cacospongia) mycofijiensis, but could be produced by as yet uncultivated microorganism | [98,102] | ||
| Chromodoris lochi (Rudman, 1982) | Vanuatu | mycothiazole (74) | dietary, sponges, but could be produced by as yet uncultivated microorganism | [103,104] | ||
| Chromodoris lochi (Rudman, 1982) | Indonesia | laulimalide (syn fijianolide B) (75), isolaulimalide (syn fijianolide A) (76) | dietary, sponges, but could be produced by as yet uncultivated microorganism | [107,108,109] | ||
| Chromodoris hamiltoni (Rudman, 1977) | South Africa | Lantrunculin A (72) and B (73) | dietary (sponge) | [100,102] | ||
| Chromodoris elisabethina (Bergh, 1877) | Queensland, Australia | Lantrunculin A (72) and B (73) | dietary (sponge) | [101,102] | ||
| Chromodoris magnifica (Quoy and Gaimard, 1832) | Queensland, Australia | Lantrunculin A (72) and B (73) | dietary (sponge) | [101,102] | ||
| Chromodoris kuiteri (Rudman, 1982) | Queensland, Australia | Lantrunculin A (72) and B (73) | dietary (sponge) | [101,102] | ||
| Chromodoris annae (Bergh, 1877) | Queensland, Australia | Lantrunculin A (72) and B (73) | dietary (sponge) | [101,102] | ||
| Chromodoris quadricolor (Rüppell and Leuckart, 1830) | Red Sea | Lantrunculin A (72) and B (73) | dietary (sponge) | [101,102] | ||
| Chromodoris inorata (Pease, 1871) (accepted as Chromodoris aspersa (Gould, 1852)) | Japan | inorolide A (77), B (78), C (79) and various scalaranes | [110] | |||
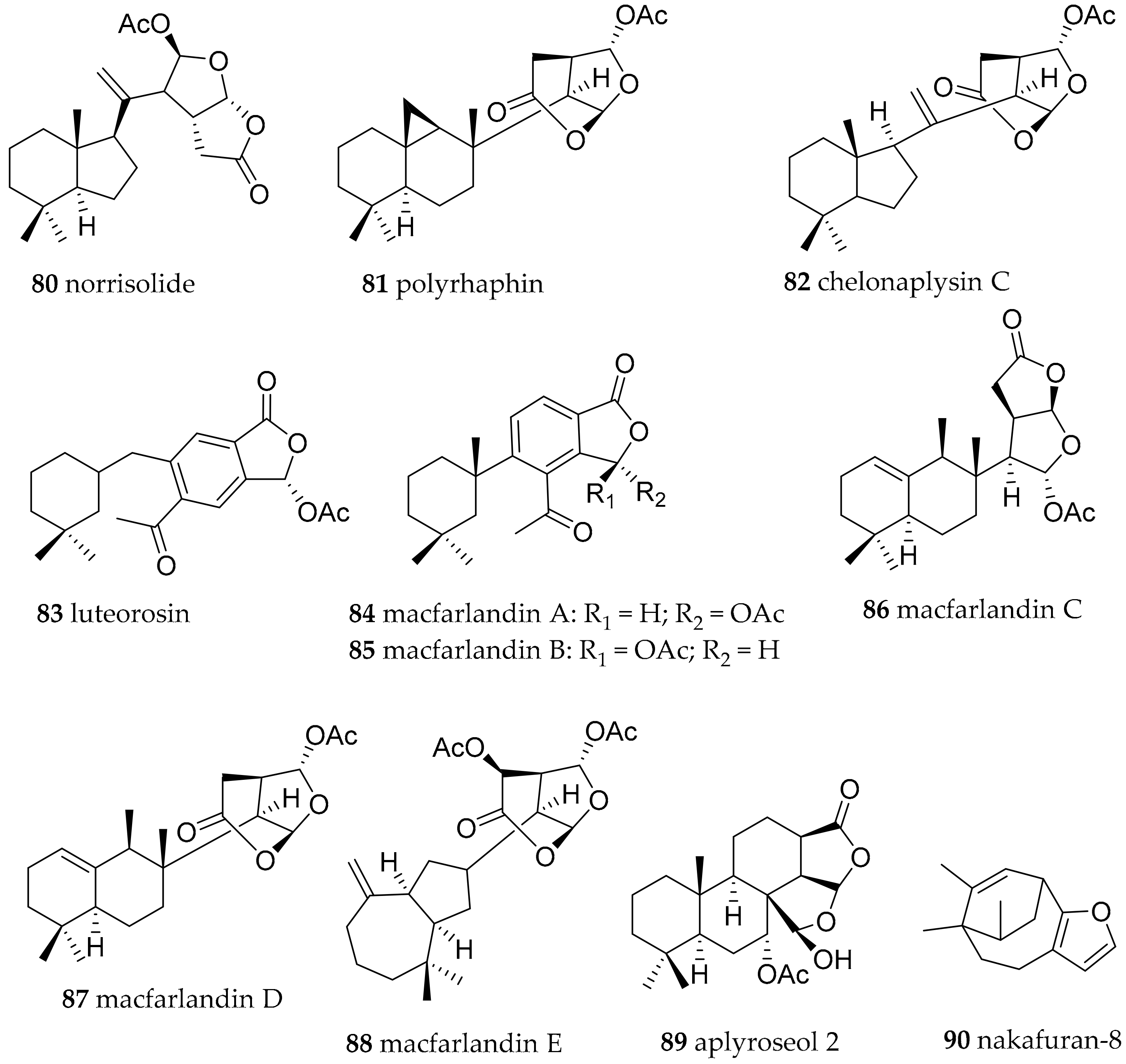
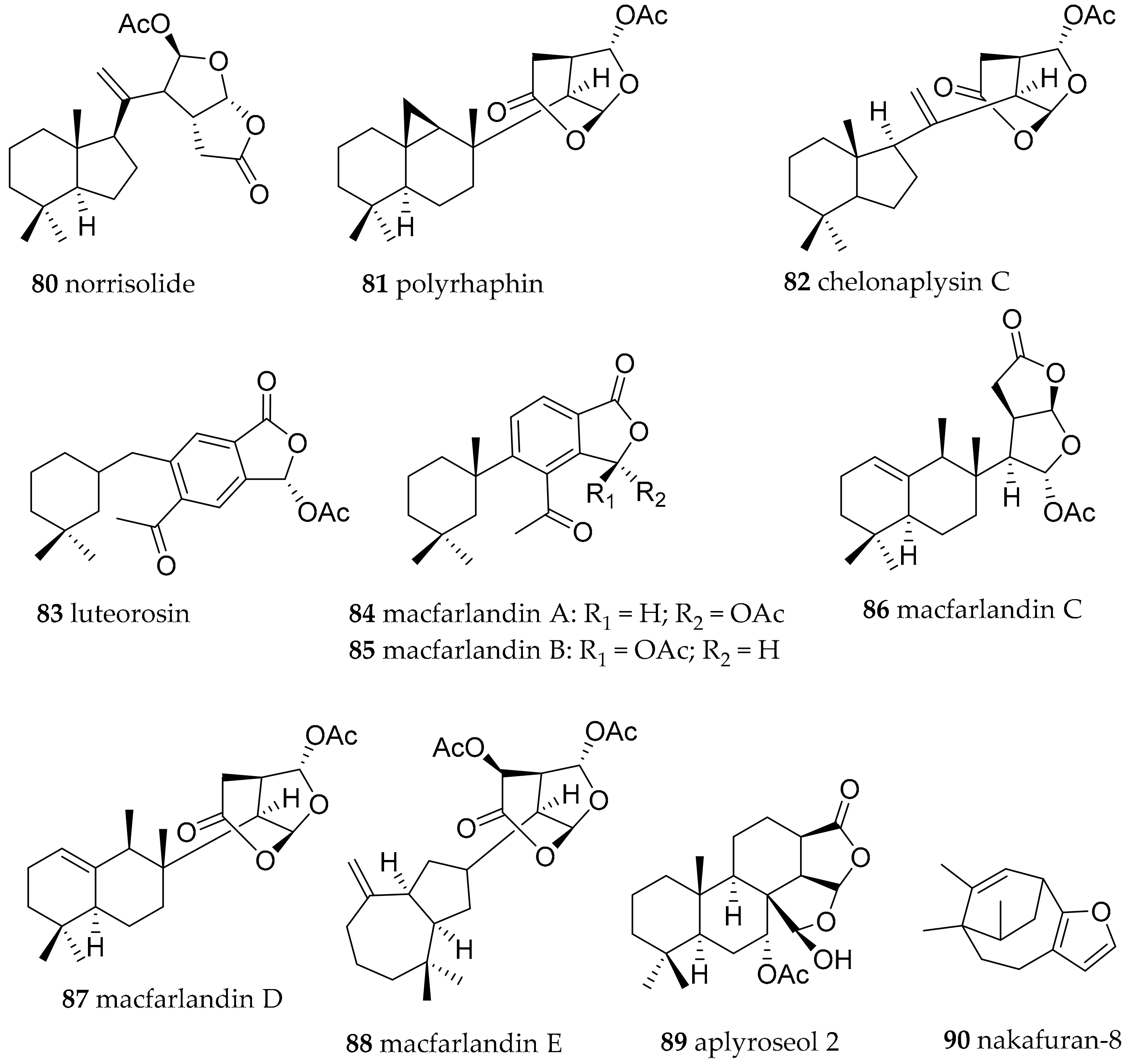
| Chromodoris luteorosea (Rapp, 1827) (accepted as Felimida luteorosea (Rapp, 1827)) | Spain | norrisolide (80), polyrhaphin C (81), chelonaplysin C (82), luterosin (83), macfarlandin A (84), | dietary (sponge) | [111] | ||
| Chromodoris macfarlandi (Cockerell, 1901) (accepted as Felimida macfarlandi (Cockerell, 1901)) | California, USA | macfarlandines A–E (84–88) | dietary (sponge, structures related to compounds from Aplysilla sulphurea) | [111,112,113] | ||
| Chromodoris sinensis (Rudman, 1985) (accepted as Goniobranchus sinensis (Rudman, 1985)) | South China Sea | Aplyroseol-2 (89) | dietary (sponge, structures related to compounds from Aplysilla sp.) | [97] | ||
| Chromodoris reticulata (Quoy and Gaimard, 1832) (accepted as Goniobranchus reticulatus (Quoy and Gaimard, 1832)) | Australia | Aplyroseol-2 (89) and other diterpenes | dietary (sponge, structures related to compounds from Aplysilla sp.) | [115] | ||
| Chromodoris maridadilus (Rudman, 1977) (accepted as Hypselodoris maridadilus (Rudman, 1977)) | Hawaii | nakafuran-9 (71), nakafuran-8 (90) | dietary (sponge Dysidea fragilis) | [116] | ||
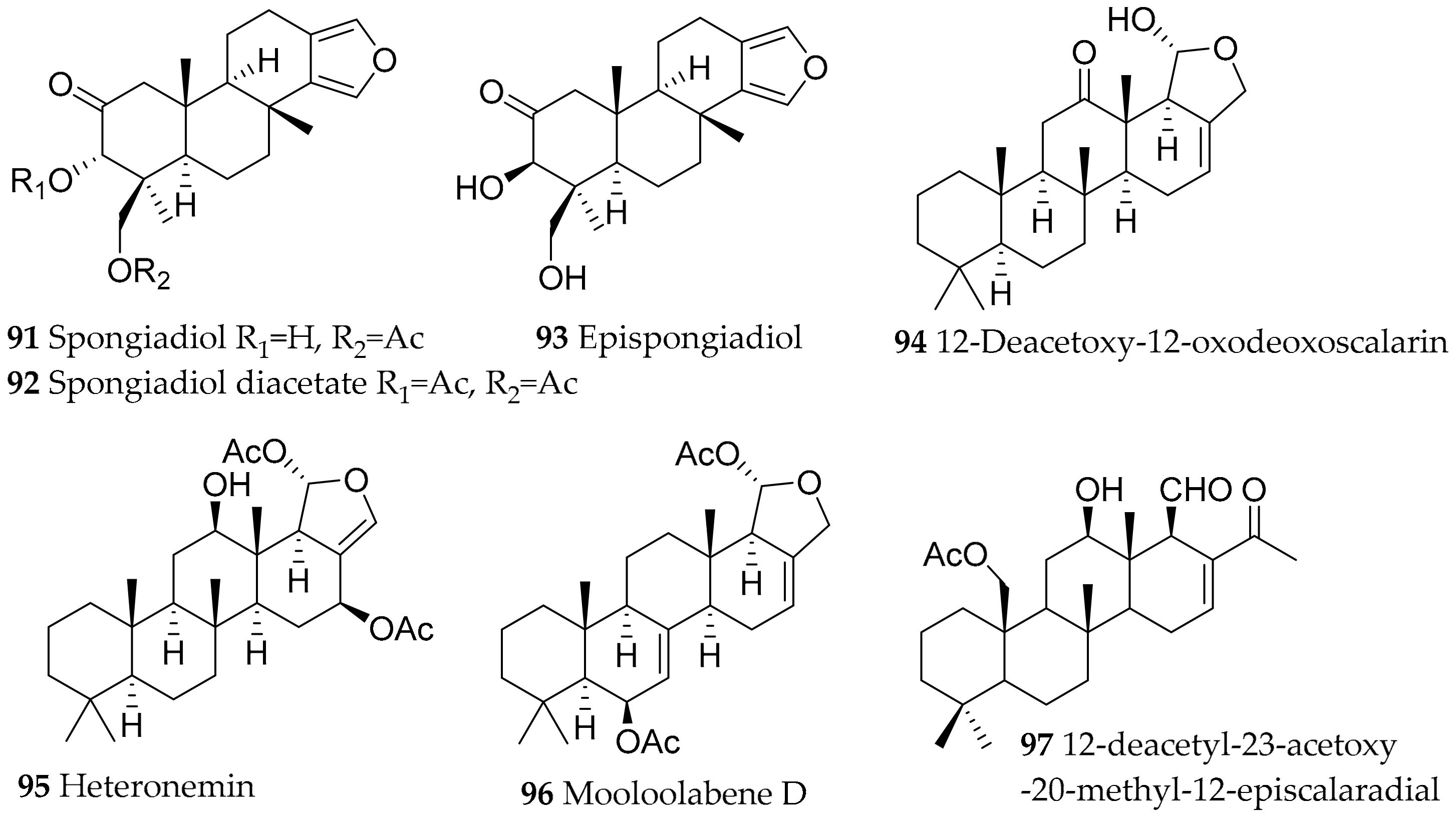
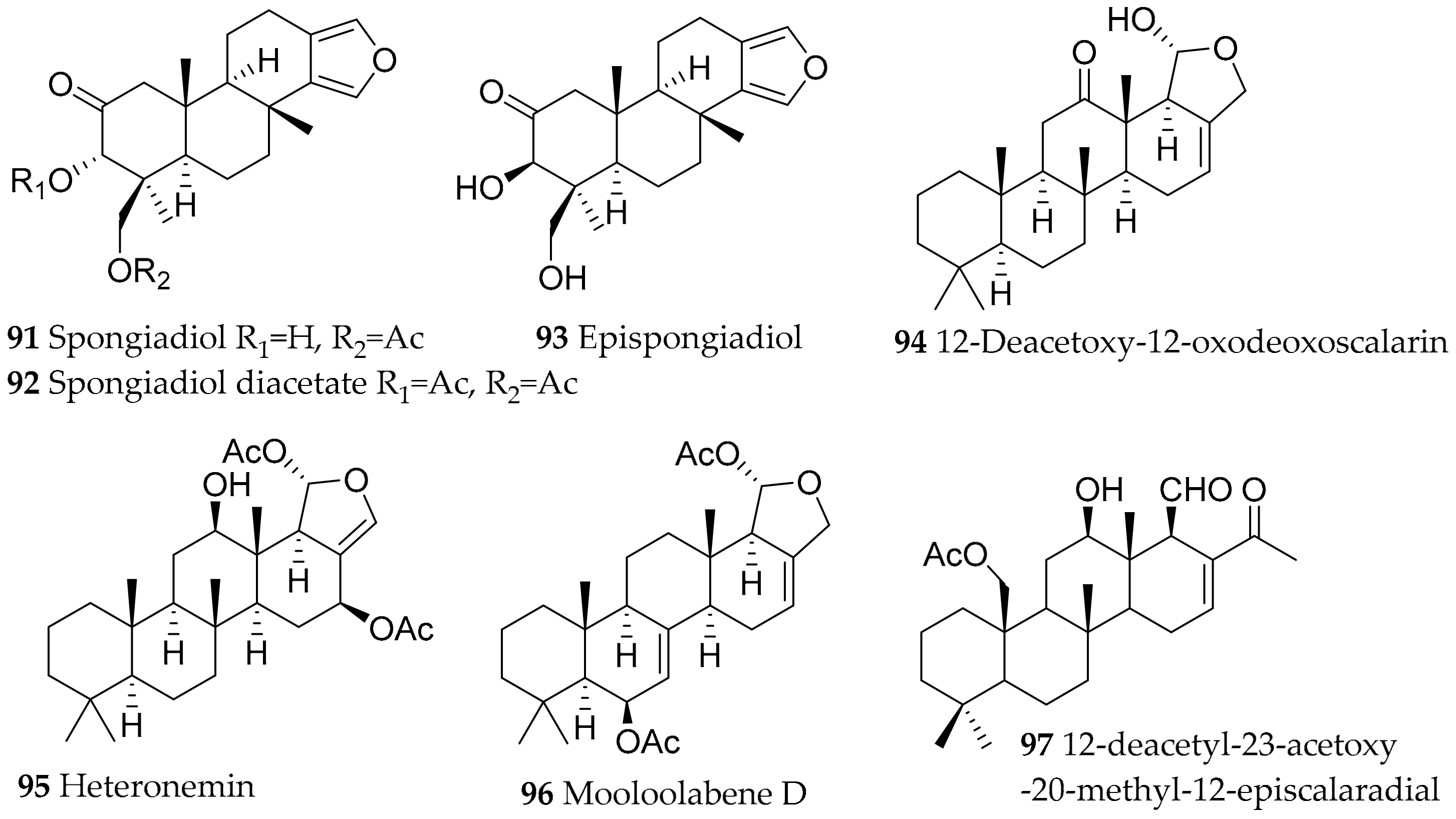
| Glossodoris atromarginata (Cuvier, 1804) (accepted as Doriprismatica stellata (Cuvier, 1804)) | Sri Lanka, Australia, India | furanoditerpenoid and scalarane type, structural variants of these metabolites (differences due to diff. sponge prey); spongiadiol (91), spongiadiol diacetate (92), epispongiadiol (93), 12-deacetoxy-12-oxodeoxoscalarin (94), heteronemin (95), mooloolabene D (96) | dietary (sponge, e.g., Spongia sp. (former Hyatella intestinales (Lamarck, 1814)), Hyrtios erectus (Keller, 1889) and Hyrtios sp.) | [117,118,119,120,121,122,123,124,125,126,127,128,129,130,131] | ||
| Glossodoris dalli (Bergh, 1879) (accepted as Felimida dalli (Bergh, 1879)) | Natural Park of Osa Ballena (Costa Rica) | homoscalarane and scalarane compounds | probably dietary from sponges | [132] | ||
| Glossodoris sedna (Ev. Marcus and Er. Marcus, 1967) (accepted as Doriprismatica sedna (Ev. Marcus and Er. Marcus, 1967)) | Natural Park of Osa Ballena (Costa Rica) | 12-deacetyl-23-acetoxy-20-methyl-12-episcalaradial (97) | probably dietary from sponges | [132] | ||
| Glossodoris rufomarginata (Bergh, 1890) | Hainan Island in the South China Sea | homoscalarane and scalarane compounds | probably dietary from sponges | [124] | ||
| Glossodoris pallida (Rüppell and Leuckart, 1830) | China and Guam | homoscalarane and scalarane compounds, different pattern at different location | probably dietary from sponges | [119] | ||
| Glossodoris vespa (Rudman, 1990) | Eastern Australia | homoscalarane and scalarane compounds | probably dietary from sponges | [119] | ||
| Glossodoris averni (Rudman, 1985) (accepted as Ardeadoris averni (Rudman, 1985)) | Eastern Australia | homoscalarane and scalarane compounds | probably dietary from sponges | [119] | ||
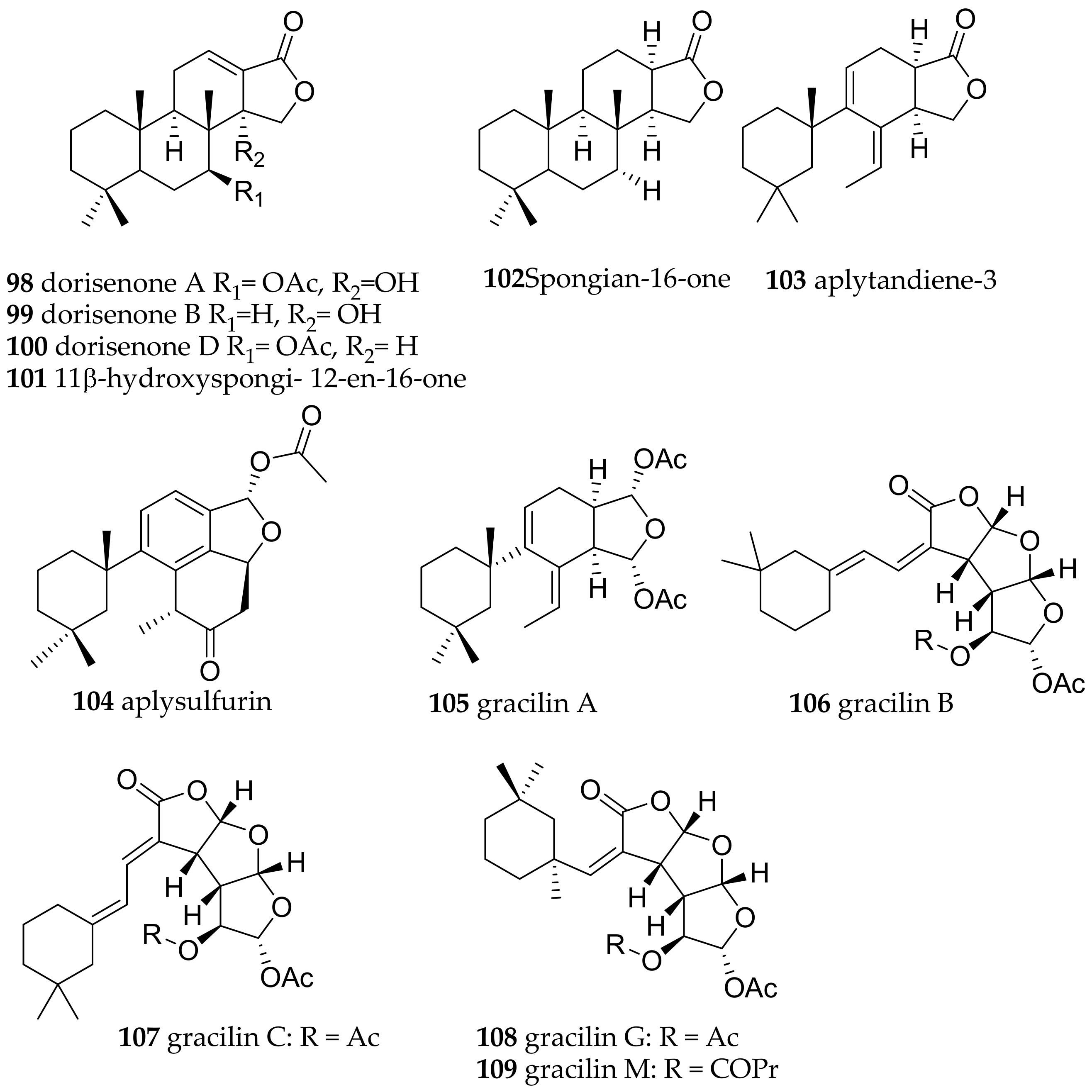
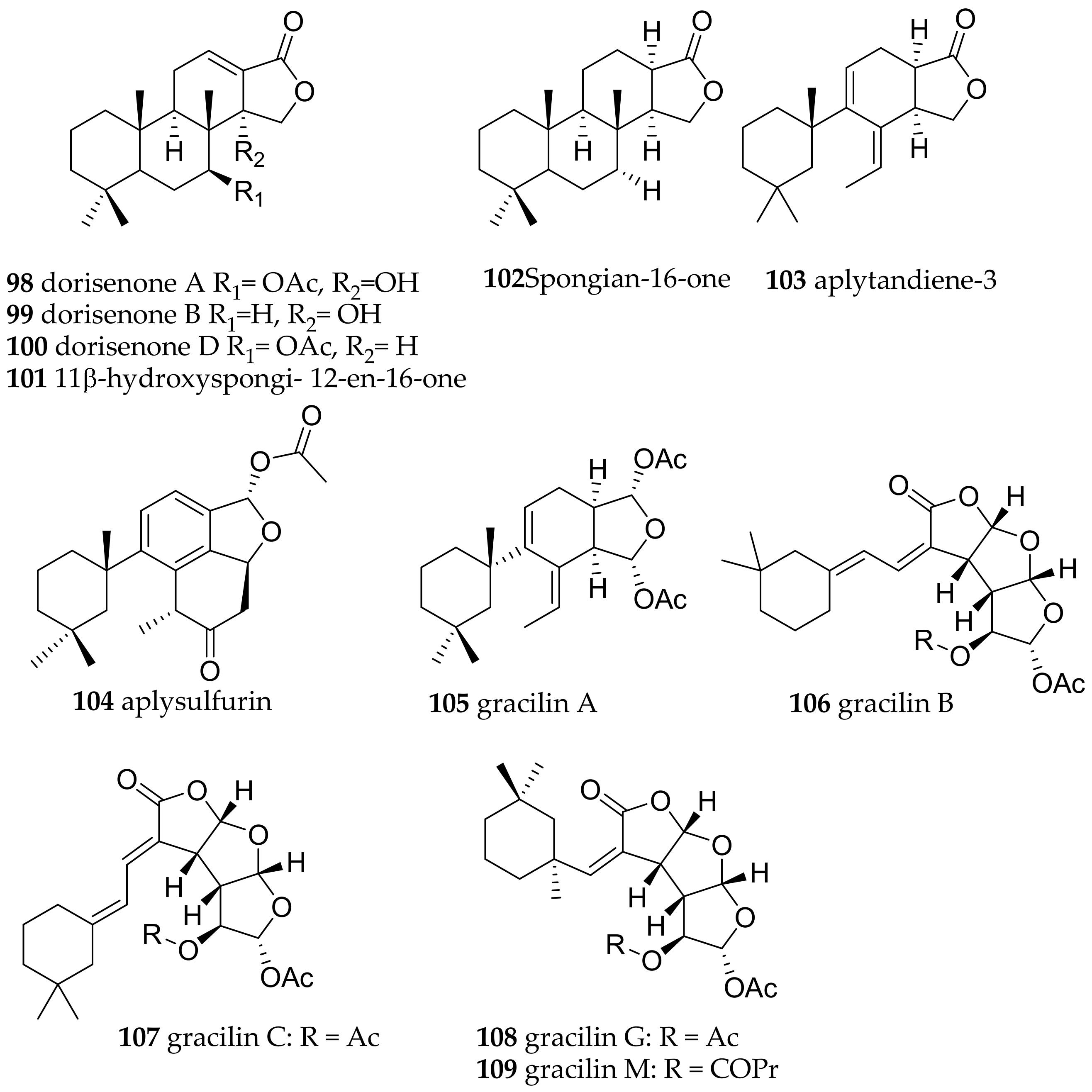
| Goniobranchus obsoletus (Rüppell and Leuckart, 1830) | Japan | most bioactive: dorisenones A (98), B (99), D (100), 11β-hydroxyspongi-12-en-16-one (101), spongian-16-one (102) | dietary (sponge Spongionella sp.) | [133] | ||
| Goniobranchus splendidus (Angas, 1864) | Australia | spongian-16-one (102), aplytandiene-3 (103), aplysulfurin (104) and aplyroseol-2 (89), the gracilins A (105), B (106), C (107), G (108), M (109)) | dietary (sponge Spongionella sp.) | [134,135,203] | ||
| Hypselodoris infucata (Rüppell and Leuckart, 1830) | Hawaii | nakafuran-8 (90) and nakafuran-9 (71) | dietary (sponge Dysidea fragilis (Montagu, 1814)) | [116,138] | ||
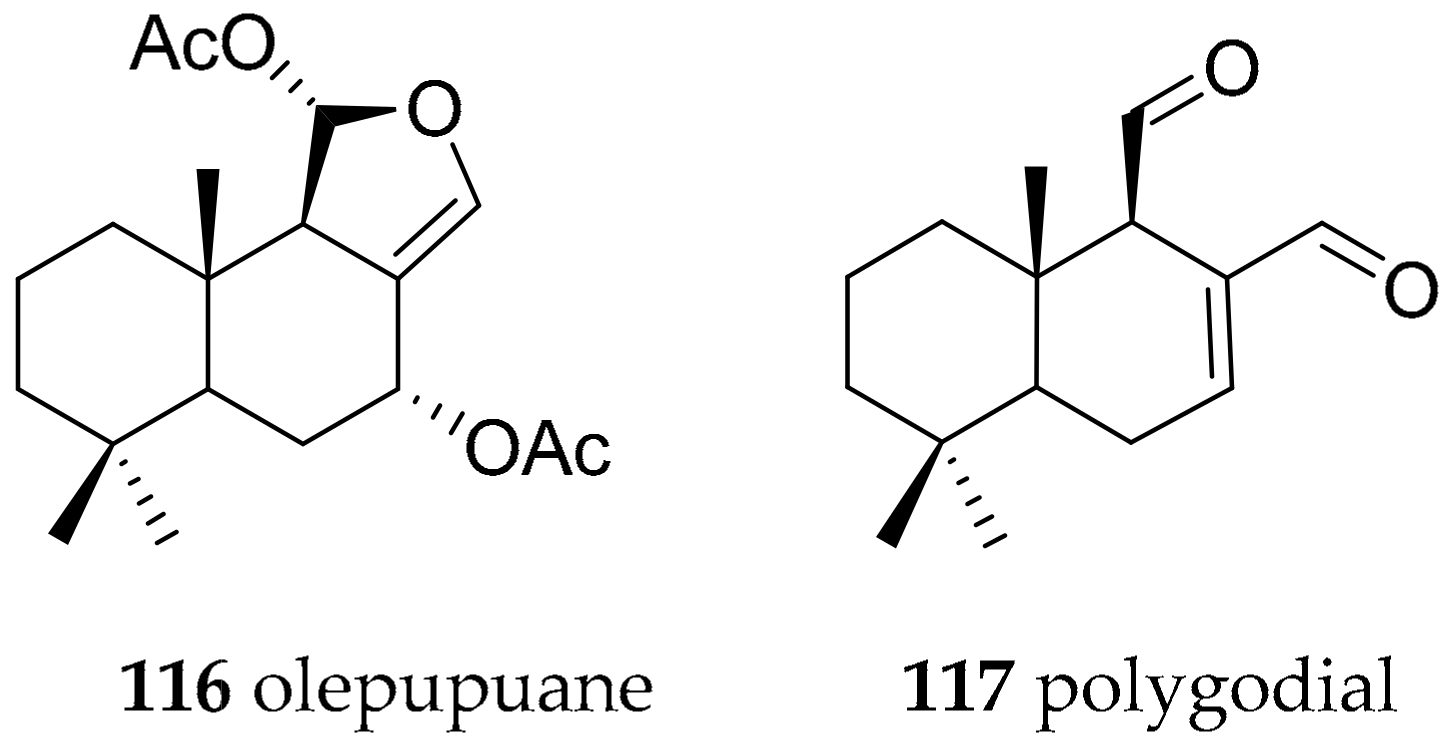
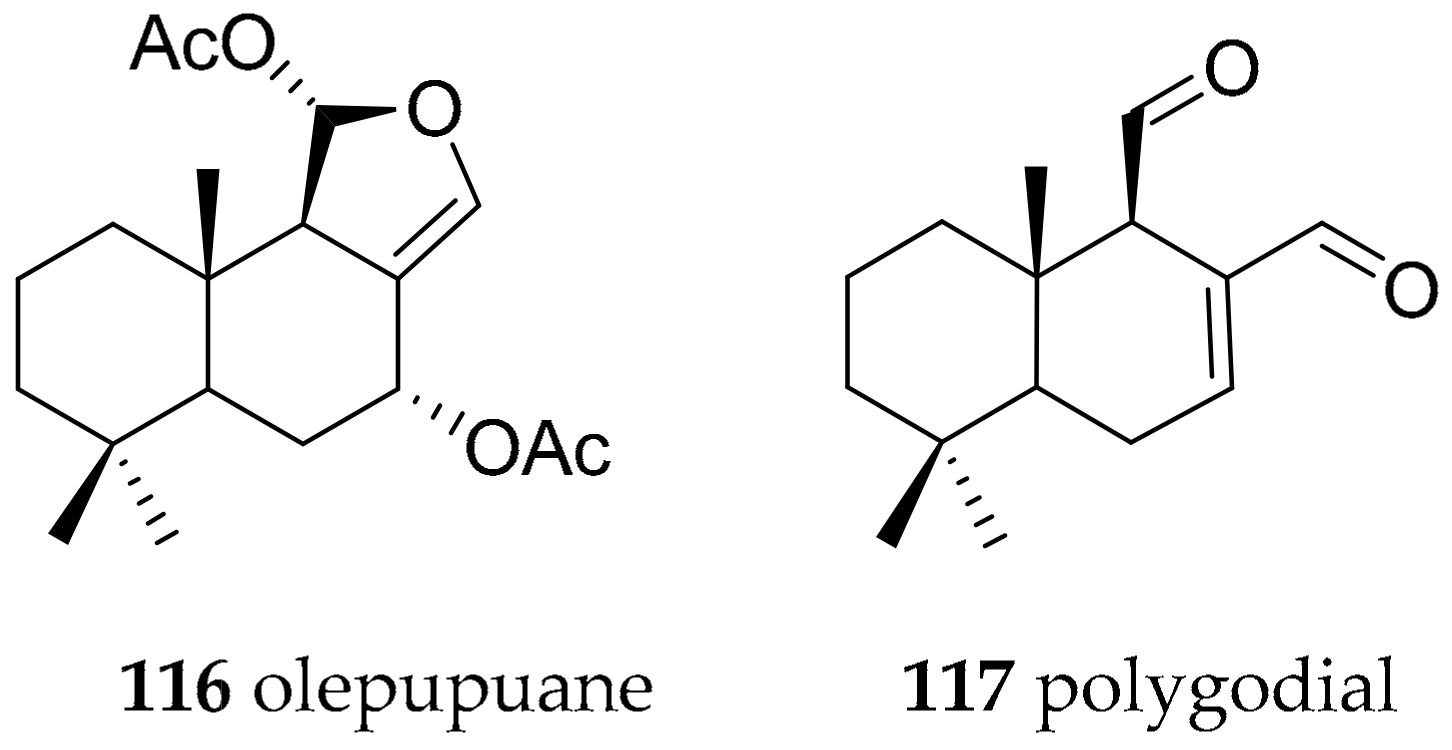
| Dendrodorididae O’Donoghue, 1924 (1864) | Dendrodoris limbata (Cuvier, 1804) | olepupuane (116) and polygodial (117) | de novo | [147,150,151,152] | ||
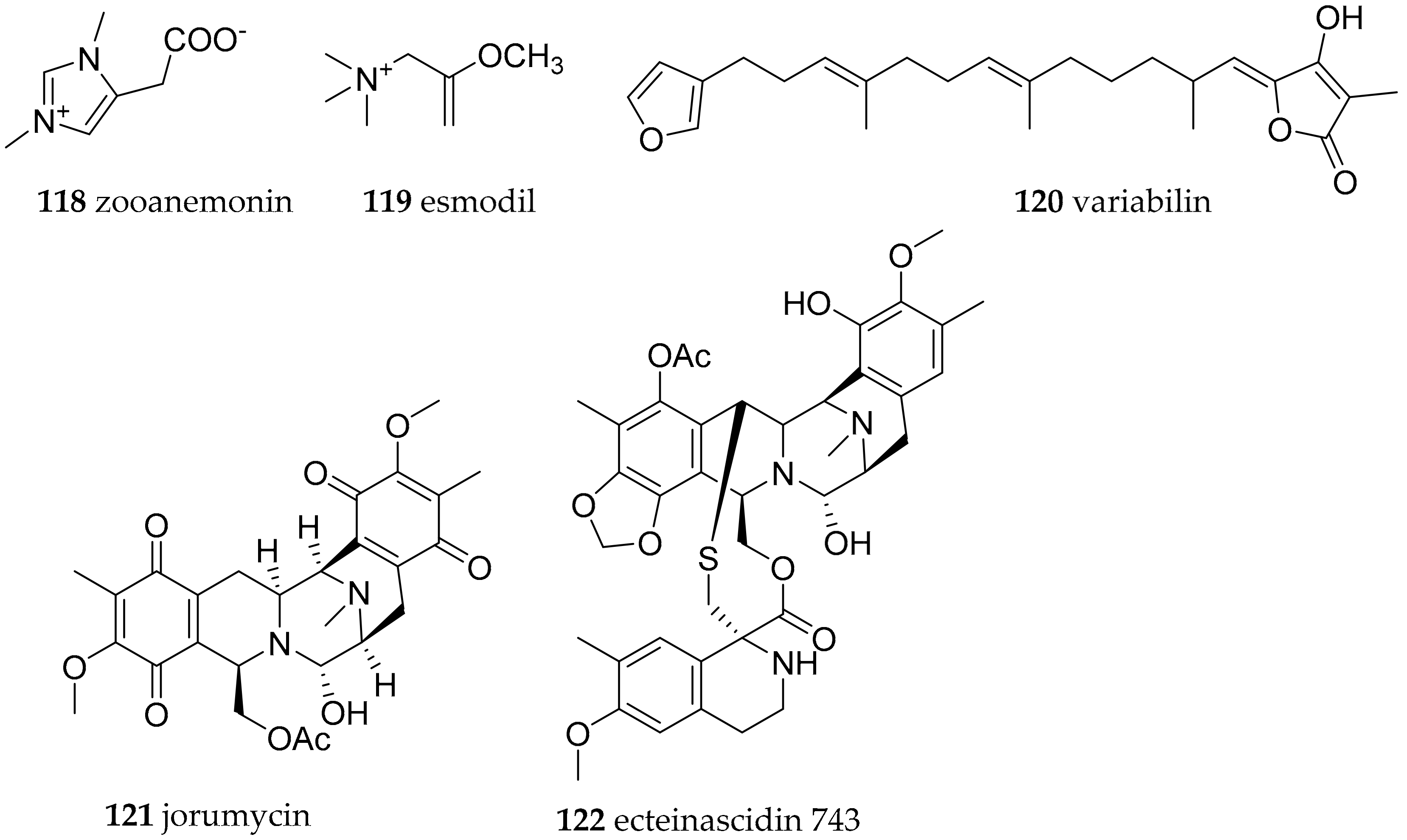
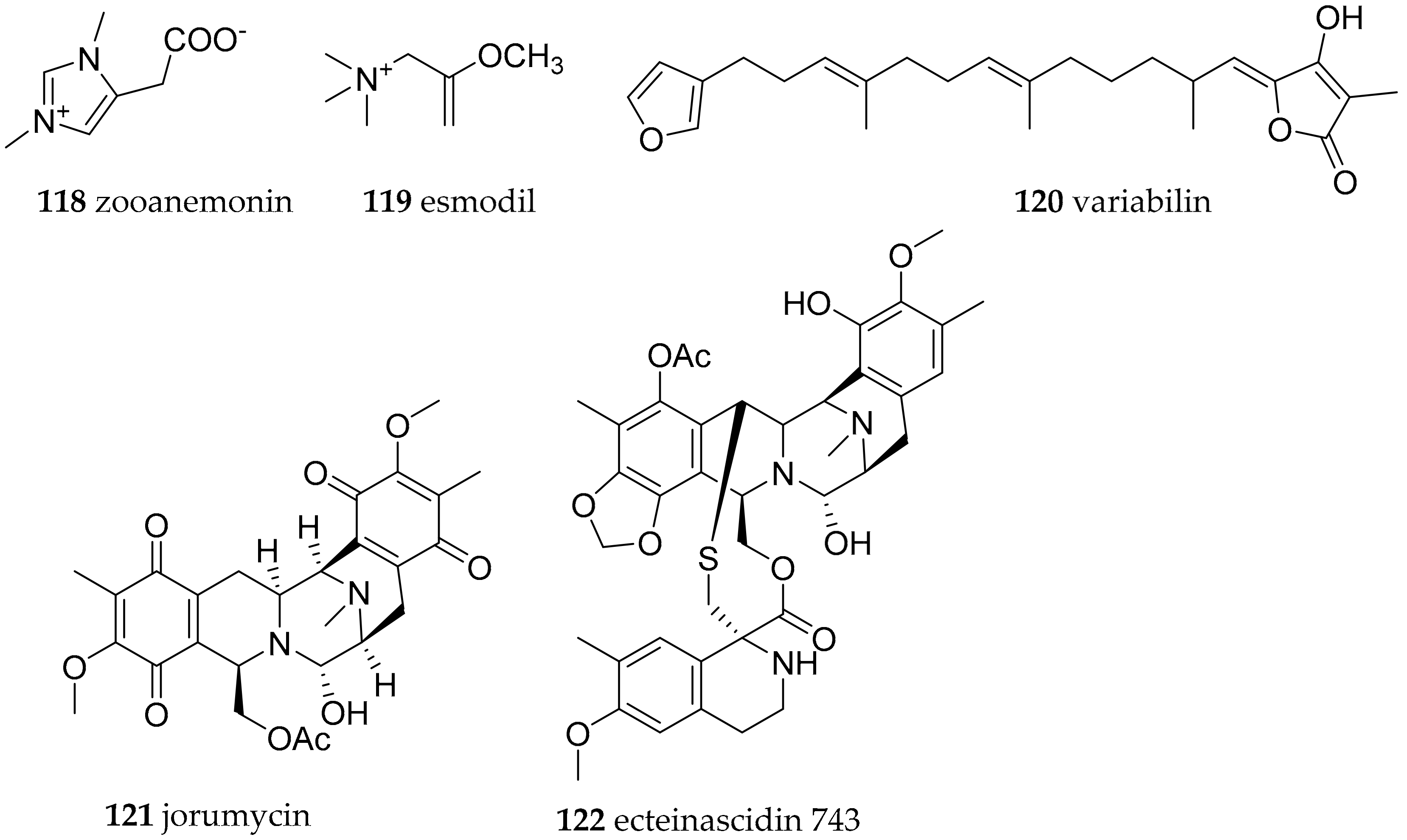
| Discodorididae Bergh, 1891 | Halgerda aurantiomaculata (Allan, 1932) | Japan, Australia | zooanemonin (118) and esmodil (119) | dietary (sponge, anemone Anemonia sulcate) | [153] | |
| Halgerda gunnessi Fahey and Gosliner, 2001 | Japan, Australia | investigated, but no compounds found | [153] | |||
| Halgerda rubicunda (Baba, 1949) (accepted as Sclerodoris rubicunda (Baba, 1949)) | Japan, Australia | investigated, but no compounds found | [153] | |||
| Halgerda theobroma (Fahey and Gosliner, 2001) | Japan, Australia | investigated, but no compounds found | [153] | |||
| Halgerda willeyi (Eliot, 1904) | Japan, Australia | investigated, but no compounds found | [153] | |||
| Paradoris indecora (Bergh, 1881) | Spain, Italy | variabilin (120) | dietary (sponge, e.g., Ircinia sp.) | [154] | ||
| Jorunna funebris (Kelaart, 1859) | India | jorumycin (121) | from structural similarity to ecteinascidin 743 (122) bacterial origin | [155,156,157] | ||
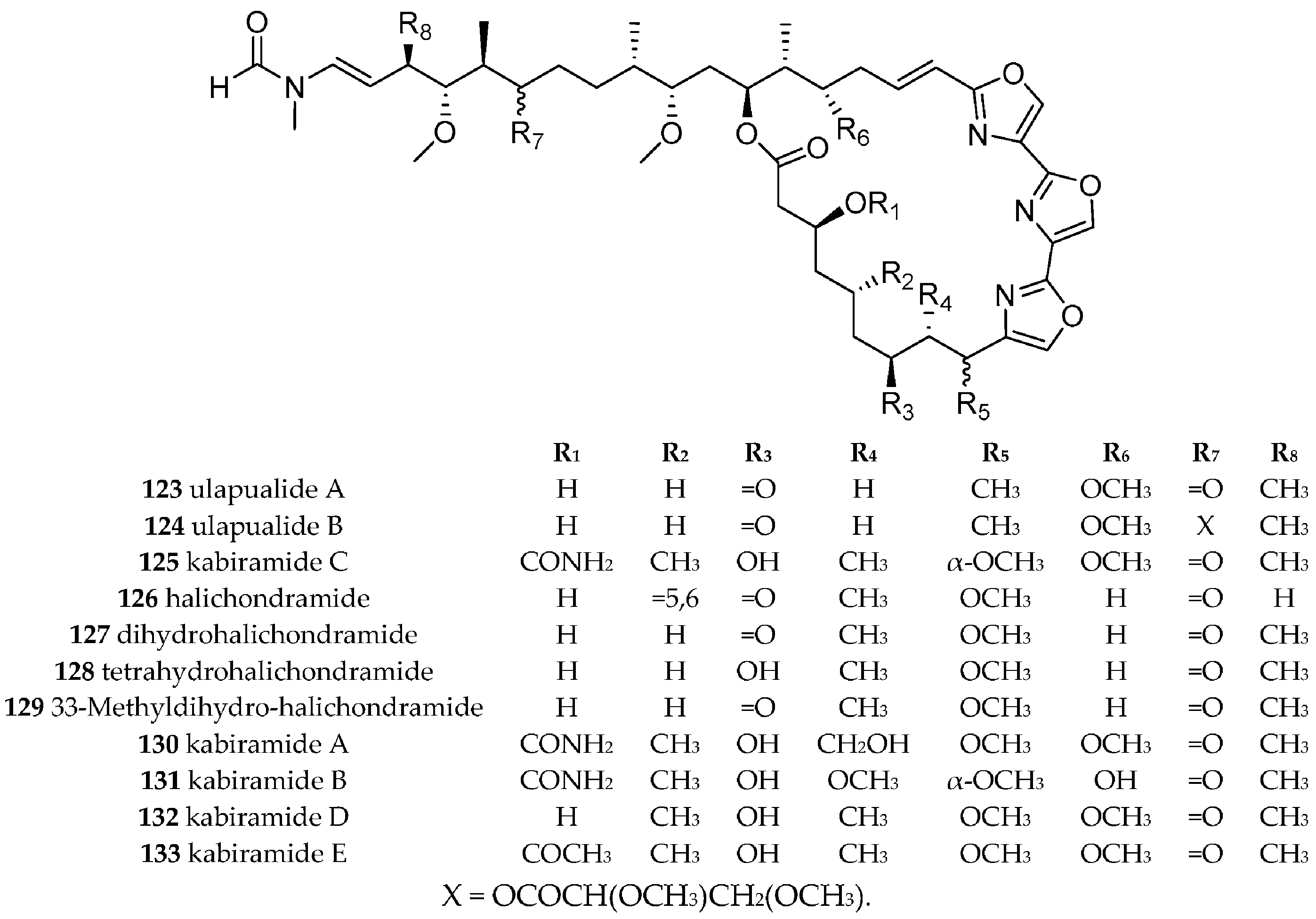
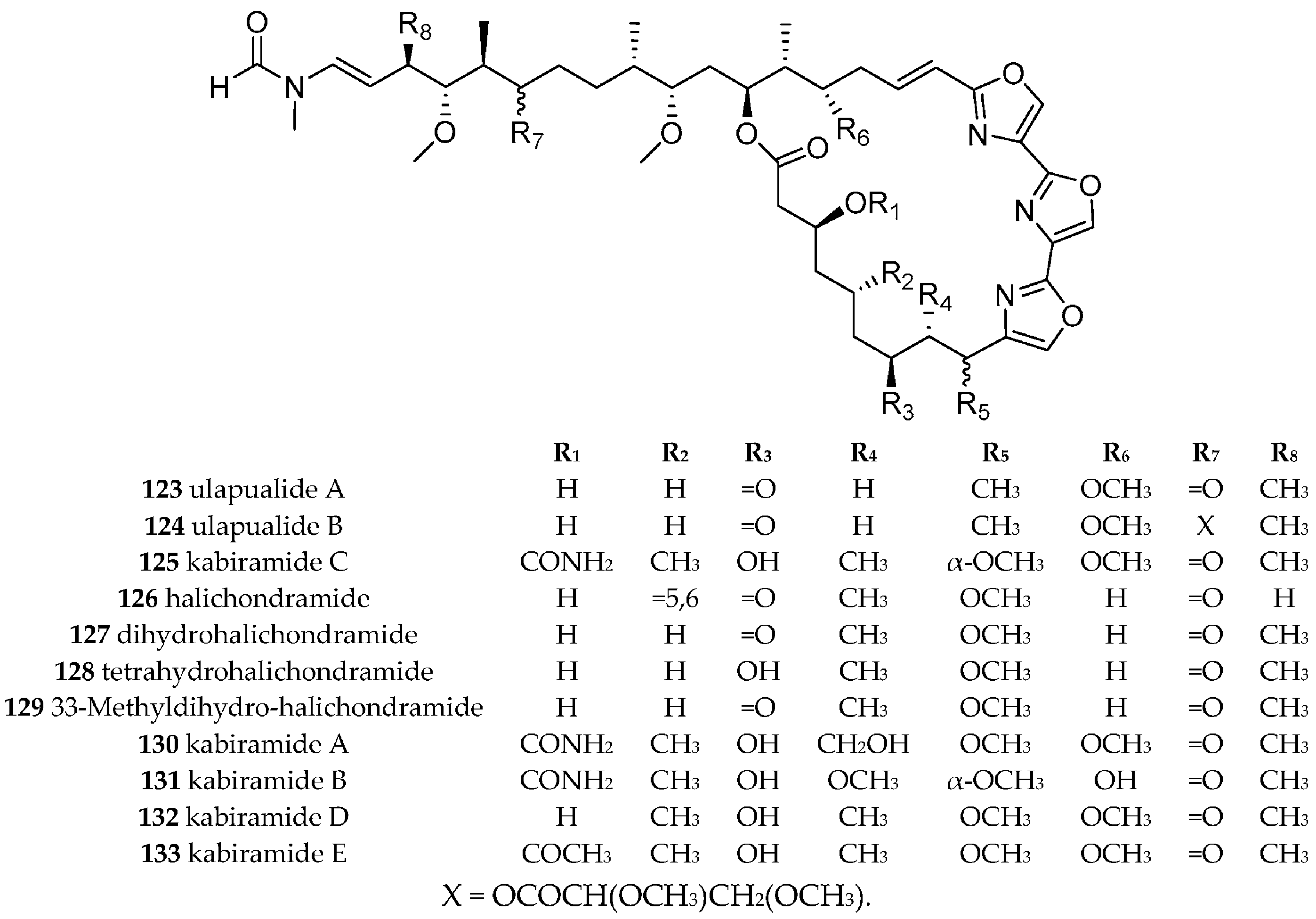
| Hexabranchidae Bergh, 1891 | Hexabranchus sanguineus (Rüppell and Leuckart, 1830) | Hawaii, Japan | Trisoxazole macrolides, i.e., ulapualide A (123) and B (124), halichondramides (126–129), kabiramide A–E (125, 130–133), | dietary, trisoxazole macrolides isolated from different sponges from the genera Halichondria, Mycale, Jaspis and Pachastrissa. halichondramides (126–129) isolated from sponge Halichondria sp. | [156,157,158,159,160,161,162,163,164,165,166,167,168] | |
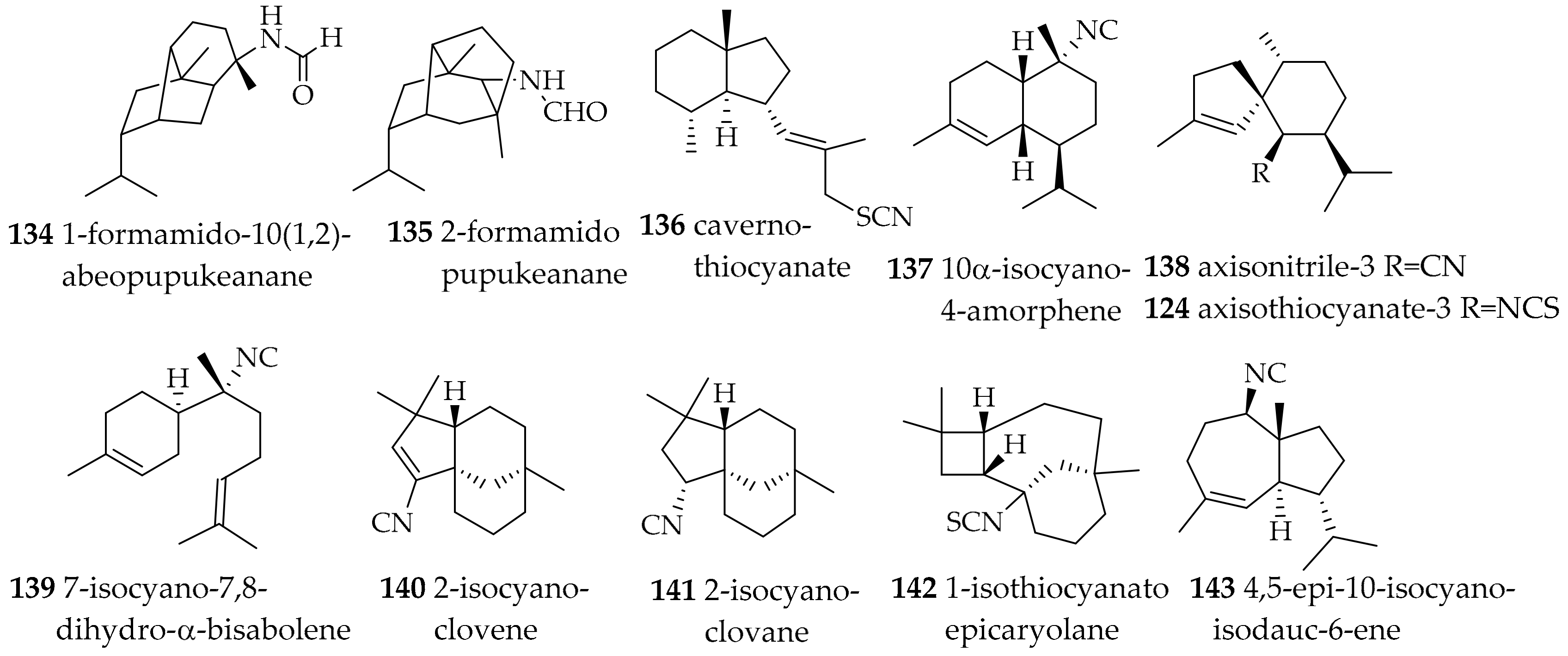
| Phyllidiidae Rafinesque, 1814 | Phyllidia coelestis (Bergh, 1905) | Thailand | 1-formamido-10(1,2)-abeopupukeanane (134), 2-formamidopupukeanane (135) | dietary (sponge) | [176,177] | |
| Phyllidia ocellata (Cuvier, 1804) | Japan | cavernothiocyanate (136), 10α-isocyano-4-amorphene (137), axisonitrile-3 (138), and 7-isocyano-7,8-dihydro-α-bisabolene (139) | dietary ((136–139) from sponge Acanthella cf. cavernosa) | [179] | ||
| Phyllidia ocellata (Cuvier, 1804) | Australia | 2-isocyanoclovene (140), 2-isocyanoclovane (141), 1-isothiocyanatoepicaryolane (142), 4,5-epi-10-isocyanoisodauc-6-ene (143) | dietary (sponges) | [180] | ||
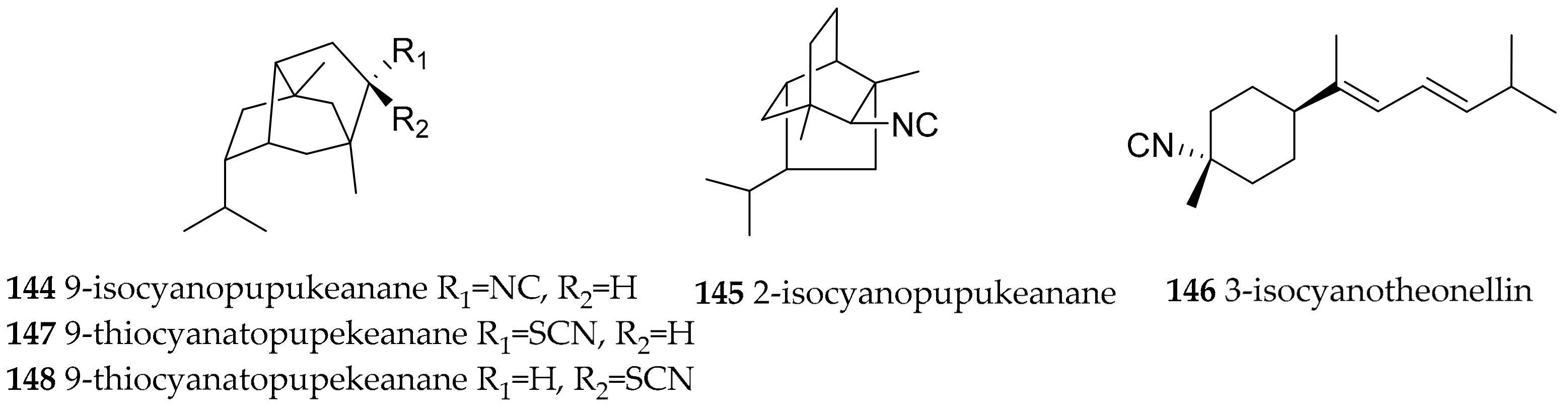
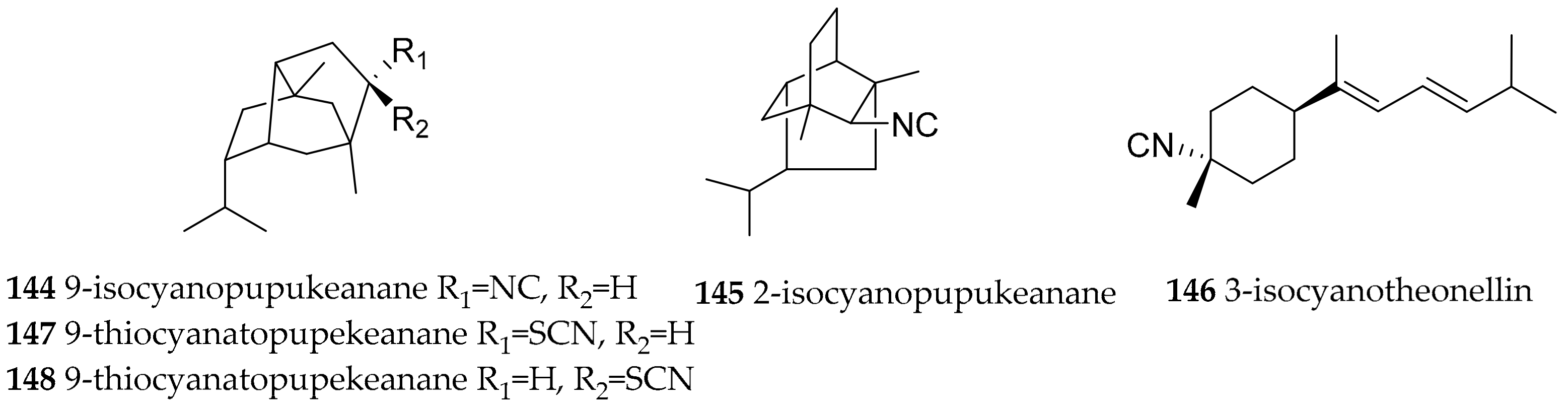
| Phyllidia varicosa (Lamarck, 1801) | Hawaii | 9-isocyanopupukeanane (144), 2-isocyanopupukeanane (145) | dietary, sponge Ciocalypta sp. (ex. Hymeniacidon sp.) | [182,183] | ||
| Phyllidia varicosa (Lamarck, 1801) | Indonesia | 9-isocyanopupukeanane (144), epimeric 9-thiocyanato-pupekeanane (147, 148) | dietary, sponge Ciocalypta sp. (ex. Hymeniacidon sp.) | [186] | ||
| Phyllidia varicosa (Lamarck, 1801) | Sri Lanka | 3-isocyanotheonellin (146) | dietary (sponge) | [185] | ||
| Phyllidiella rosans (Bergh, 1873) | Japan | 9-isocyanopupukeanane (144) | dietary (sponge) | [184] | ||
| Phyllidiella pustulosa (Cuvier, 1804) | Japan | axisonitrile-3 (138), unnamed molecule with isocyano group (155) and substituted axinisothiocyanate K derivative | dietary (sponge) | [190] | ||
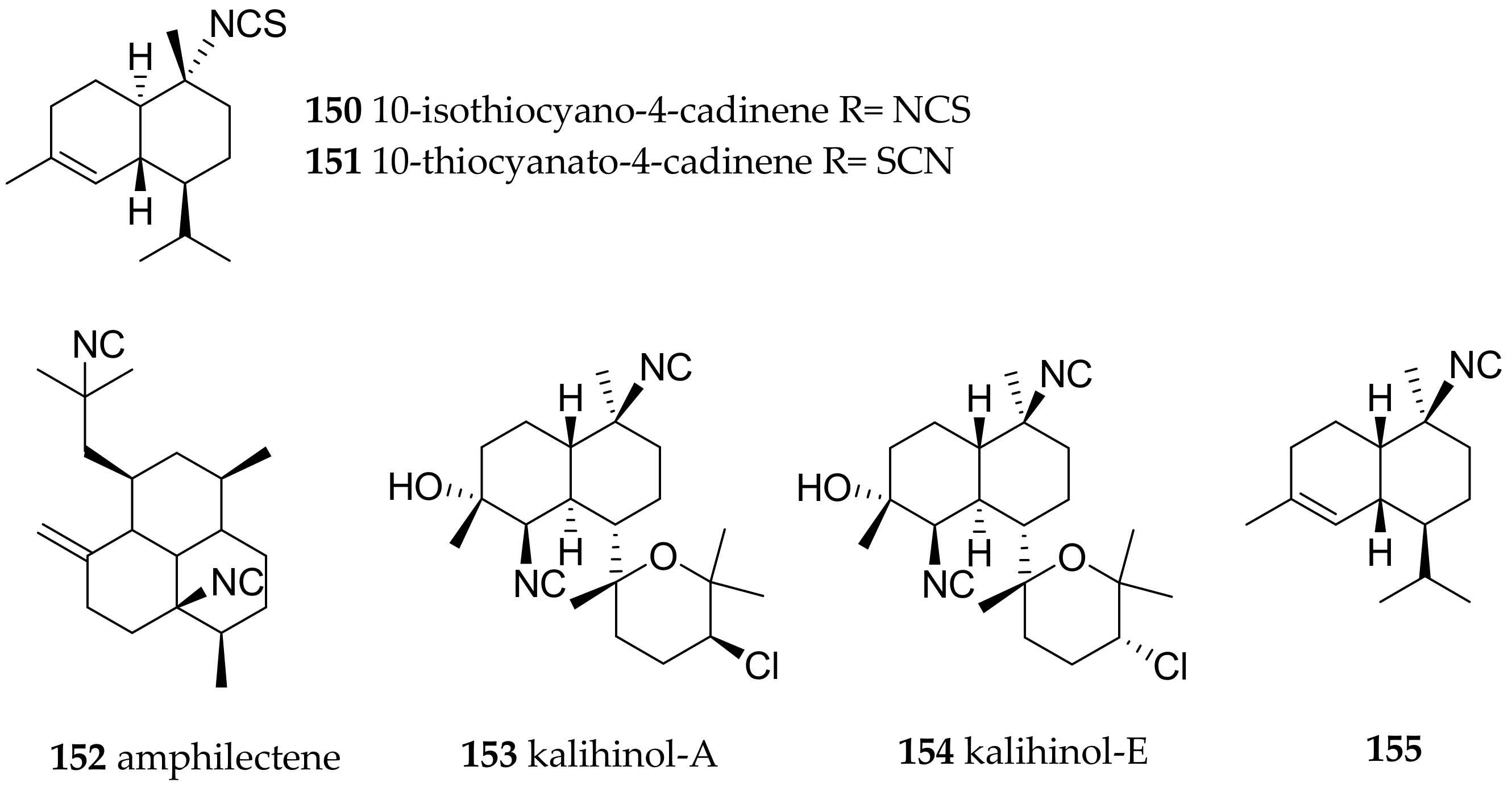
| Phyllidiella pustulosa (Cuvier, 1804) | China | 3-isocyanotheonellin (146), amphilectene (152), kalihinol-A (153), kalihinol-E (154) | dietary (sponges, due to very similar compounds present in both samples) | [175] | ||
| Phyllidiella pustulosa (Cuvier, 1804) | Vietnam | 9-isocyanopupukeanane (144) and its C-9 epimer | dietary, based on the terpenes obtained the authors concluded that the mollusk feeds on sponges of the genera Acanthella, Halichondria, Axinella and Axinyssa | [189] | ||
| Phyllidiella pustulosa (Cuvier, 1804) | Fiji | axisonitrile-3 (138), 10-isothiocyano-4-cadinene (150)/10-thiocyanato-4-cadinene (151) | sponge Phakellia carduus, due to very similar secondary metabolites present in both samples | [187,188] | ||
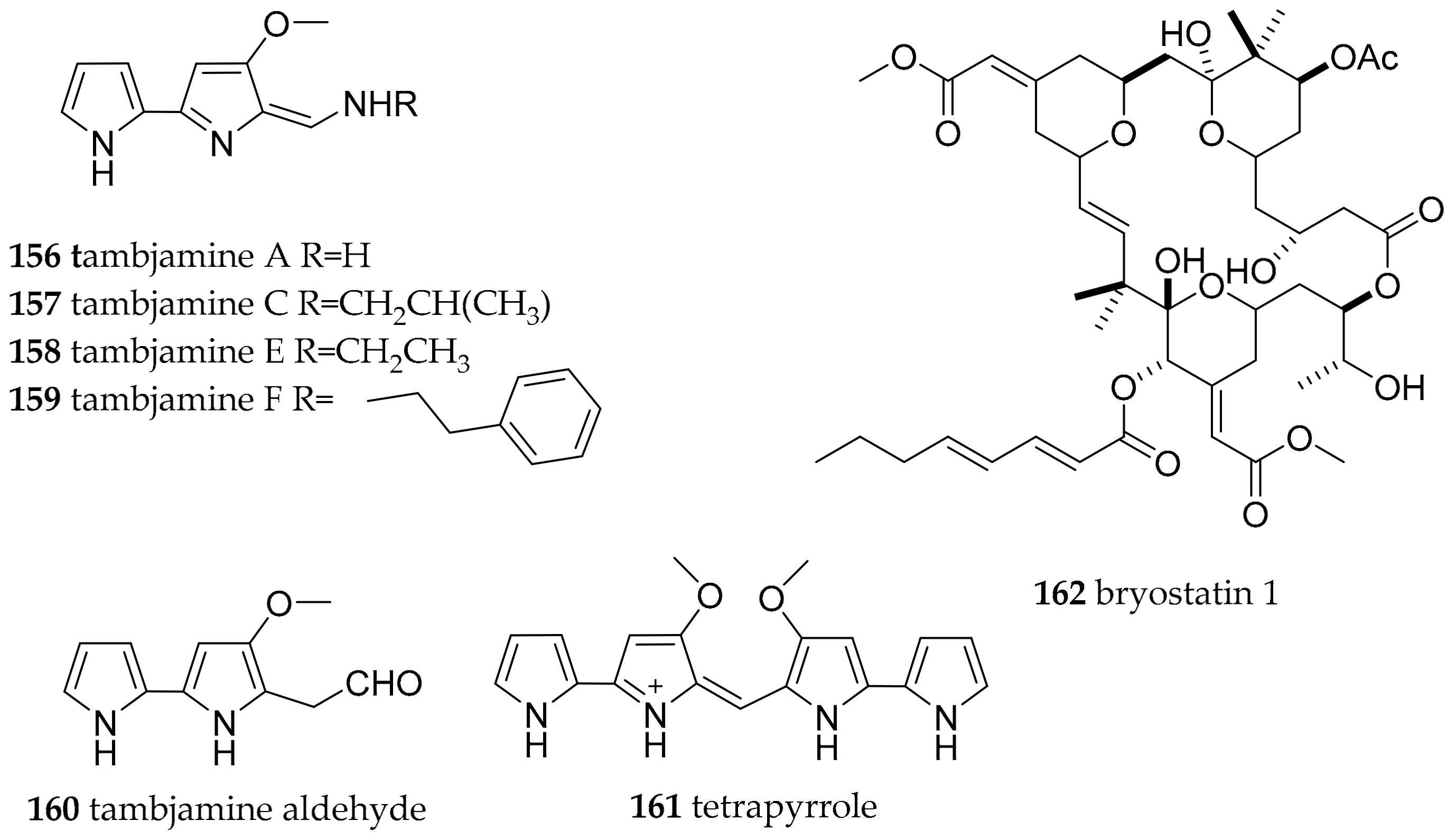
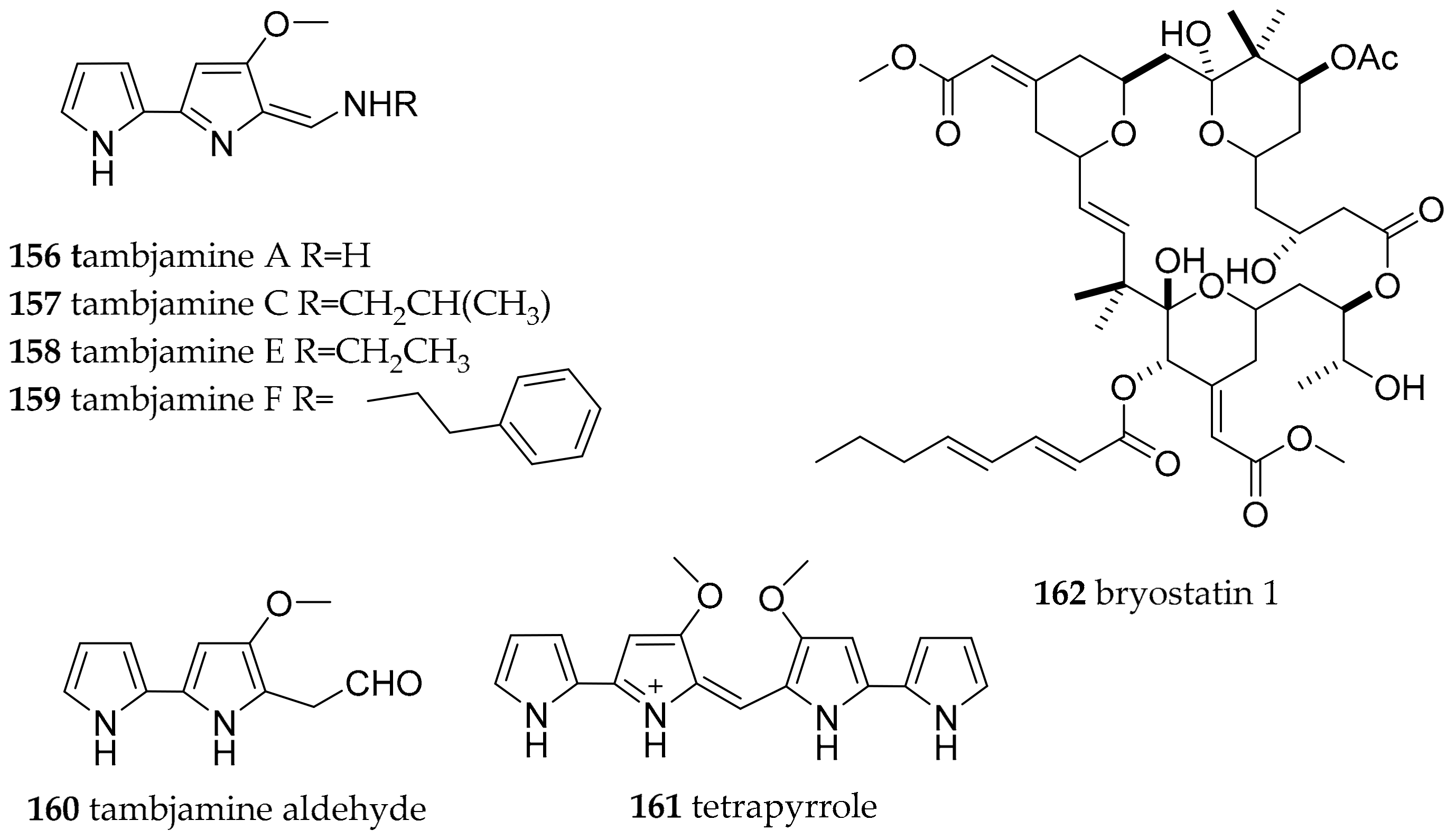
| Polyceridae Alder and Hancock, 1845 | Tambja abdere (Farmer, 1978) | West coast of America | tambjamines A–D, tambjamines A (156), C (157) | dietary (bryozoan Sessibugula translucens) | [191] | |
| Tambja eliora (Er. Marcus and Ev. Marcus, 1967) | West coast of America | tambjamines A-D, tambjamines A (156), C (157) | dietary (bryozoan Sessibugula translucens) | [191] | ||
| Roboastra tigris (Farmer, 1978) | Gulf of California | tambjamines A-D, tambjamines A (156), C (157) | dietary (bryozoan Bugula neritida) | [191] | ||
| Nembrotha species | Micronesia | tambjamines A (156), C (157), E (158), F (159), the tambjamine aldehyde (160) and the blue tetrapyrrol (161) | dietary (ascidian Atapozoa sp.) | [138,192] | ||
| Nembrotha cristata (Bergh, 1877) | Ant Atoll | tambjamines A (156), C (157), E (158), F (159), the tambjamine aldehyde (160) and the blue tetrapyrrol (161) | dietary (ascidian Atapozoa sp.) | [138,192] | ||
| Nembrotha kubaryana (Bergh, 1877) | Sumilon Island, the Philippines | tambjamines C (157), E (158) and the blue tetrapyrrol (161) | dietary (ascidian Atapozoa sp.) | [138,192] | ||
| Nembrotha sp. | Apo Islands, the Philippines | tambjamines C (157), E (158), F (159), and the tambjamine aldehyde (160) | dietary (ascidian Atapozoa sp.) | [192] | ||
| Polycera atra (MacFarland, 1905) | Torrey Pines artificial reef | bryostatins, e.g., bryostatin 1 (162) | dietary (bryozoan Bugula neritina) | [193,194,195] | ||
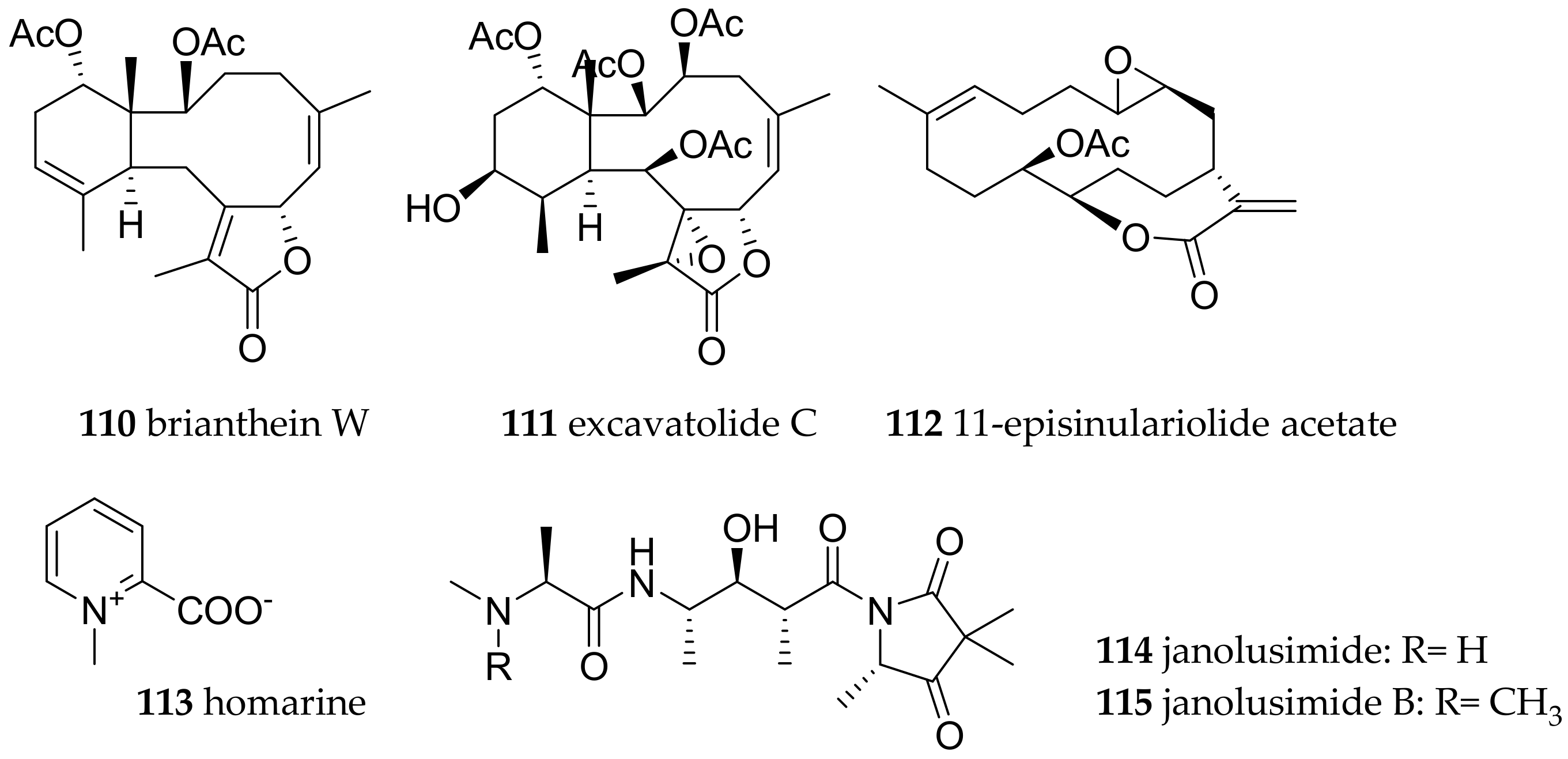
| Subclade Cladobranchia | Facelinidae Bergh, 1889 | Phyllodesmium briareum (Bergh, 1896) | not known | brianthein W (110) and excavatolide C (111) | dietary (coral Briareum sp.) | [1] |
| Phyllodesmium magnum (Rudman, 1991) | not known | cembrane diterpenes, e.g., 11-episinulariolide acetate (112) | dietary, e.g., Sinularia spp., Capnella sp. | [1] | ||
| Flabellinidae Bergh, 1889 | Flabellina exoptata (Gosliner and Willan, 1991) | not known | hormarin (113) | dietary | [139,141] | |
| Flabellina ischitana (Hirano and Thompson, 1990) | not known | hormarin (113) | dietary | [139,141] | ||
| Flabellina pedata (Montagu, 1816) | not known | hormarin (113) | dietary | [139,141] | ||
| Flabellina affinis (Gmelin, 1791) | not known | hormarin (113) | dietary | [139,141] | ||
| Proctonotidae Gray, 1853 | Janolus cristatus (Delle Chiaje, 1841) | Mediterranean | janolusimide (114) | dietary (bryozoa) | [139,141] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fisch, K.M.; Hertzer, C.; Böhringer, N.; Wuisan, Z.G.; Schillo, D.; Bara, R.; Kaligis, F.; Wägele, H.; König, G.M.; Schäberle, T.F. The Potential of Indonesian Heterobranchs Found around Bunaken Island for the Production of Bioactive Compounds. Mar. Drugs 2017, 15, 384. https://doi.org/10.3390/md15120384
Fisch KM, Hertzer C, Böhringer N, Wuisan ZG, Schillo D, Bara R, Kaligis F, Wägele H, König GM, Schäberle TF. The Potential of Indonesian Heterobranchs Found around Bunaken Island for the Production of Bioactive Compounds. Marine Drugs. 2017; 15(12):384. https://doi.org/10.3390/md15120384
Chicago/Turabian StyleFisch, Katja M., Cora Hertzer, Nils Böhringer, Zerlina G. Wuisan, Dorothee Schillo, Robert Bara, Fontje Kaligis, Heike Wägele, Gabriele M. König, and Till F. Schäberle. 2017. "The Potential of Indonesian Heterobranchs Found around Bunaken Island for the Production of Bioactive Compounds" Marine Drugs 15, no. 12: 384. https://doi.org/10.3390/md15120384