Identification and Biochemical Characterization of Halisulfate 3 and Suvanine as Novel Inhibitors of Hepatitis C Virus NS3 Helicase from a Marine Sponge

 , and
, and
Abstract
:1. Introduction
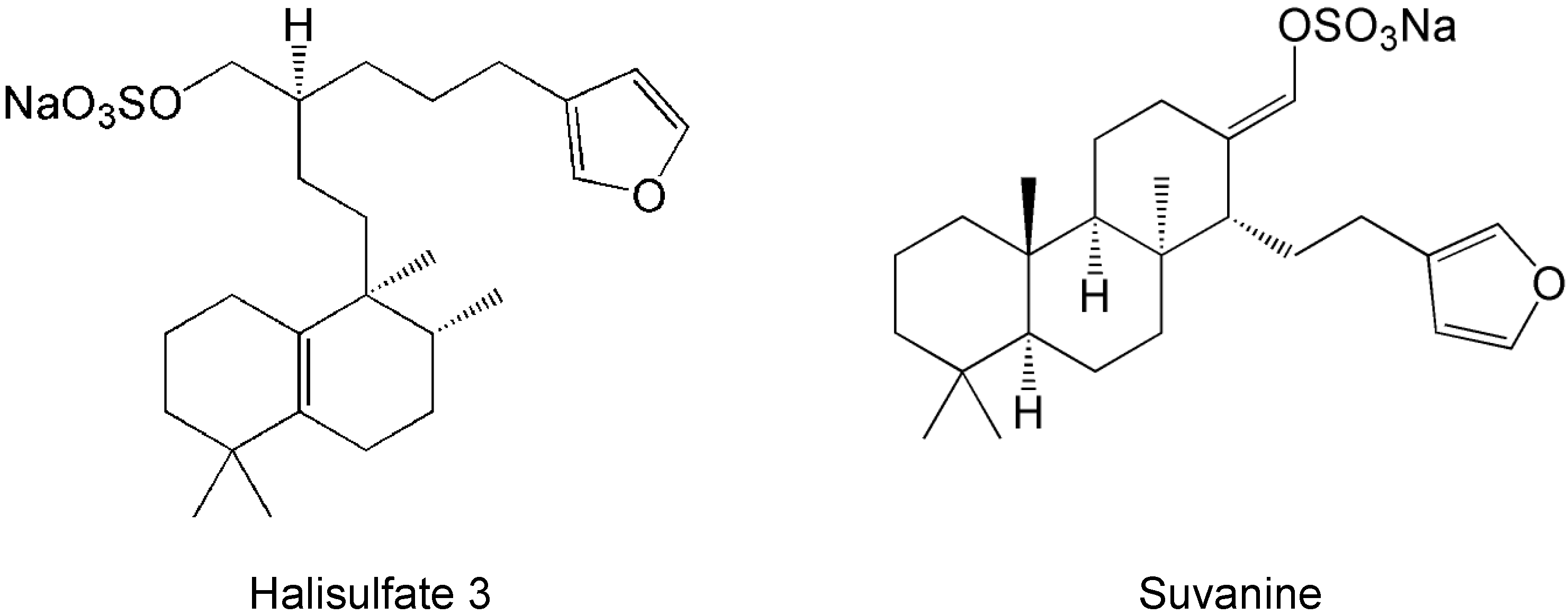
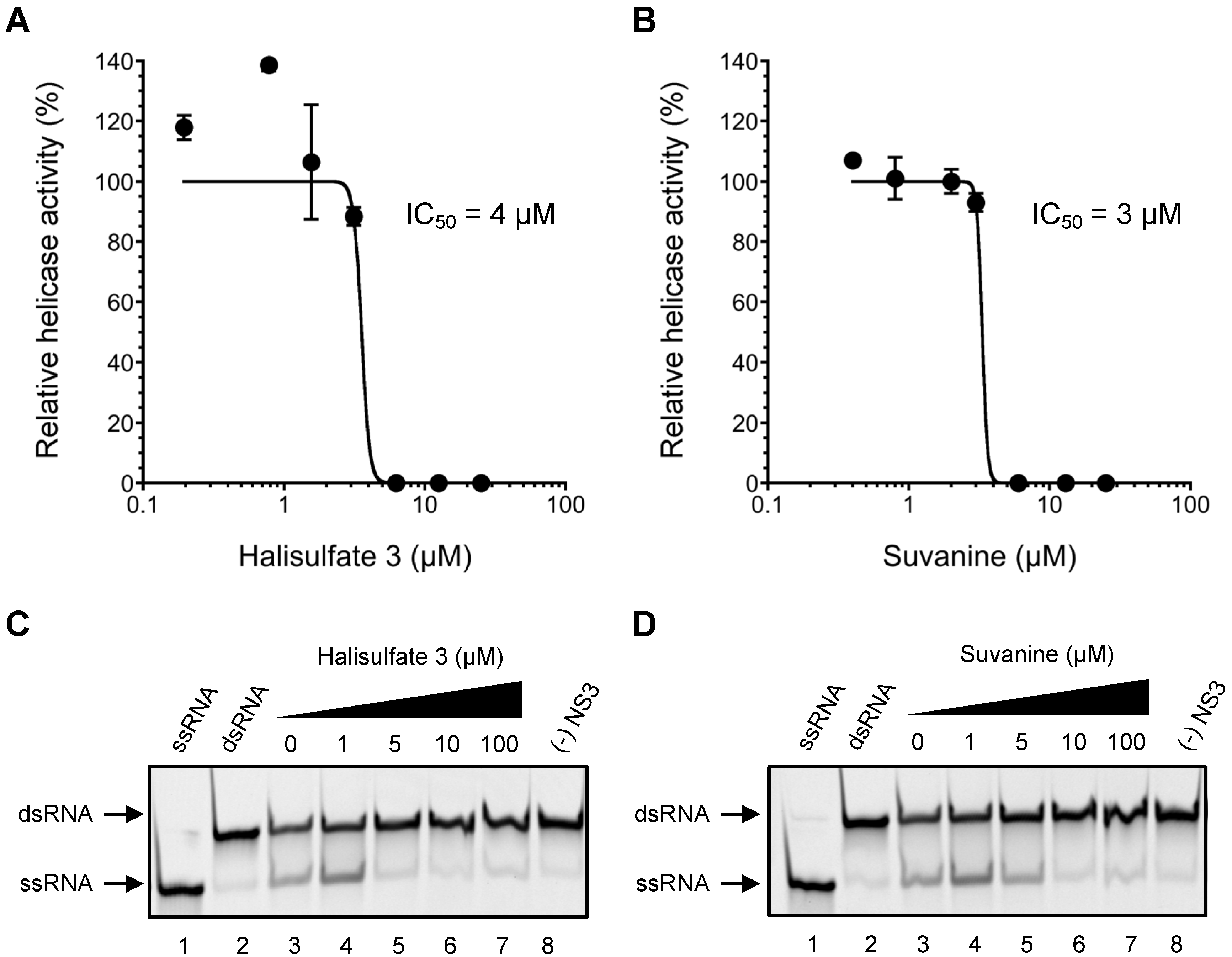
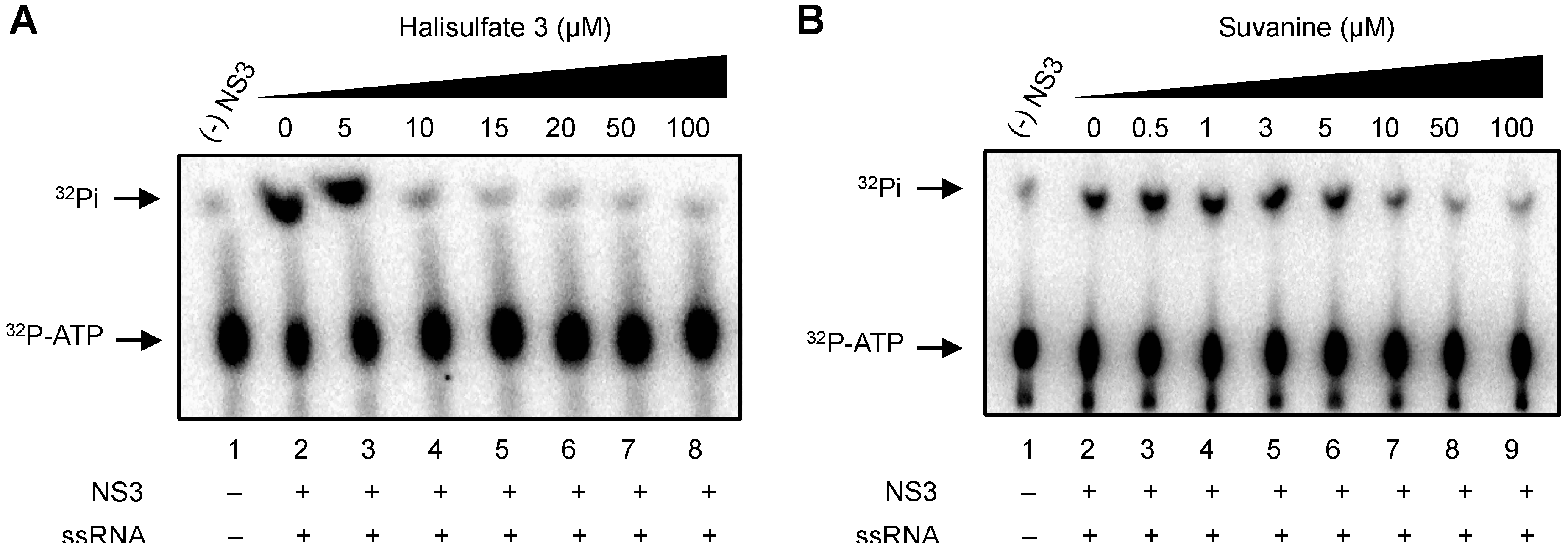
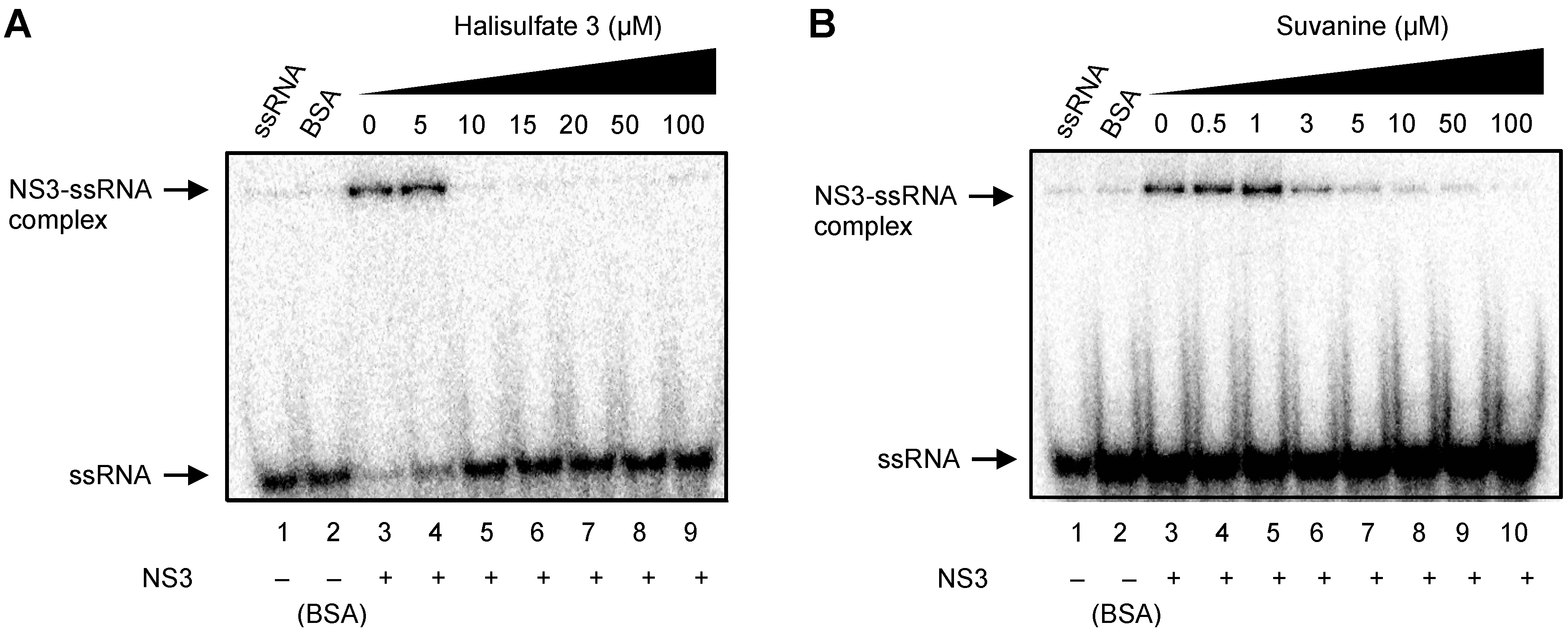
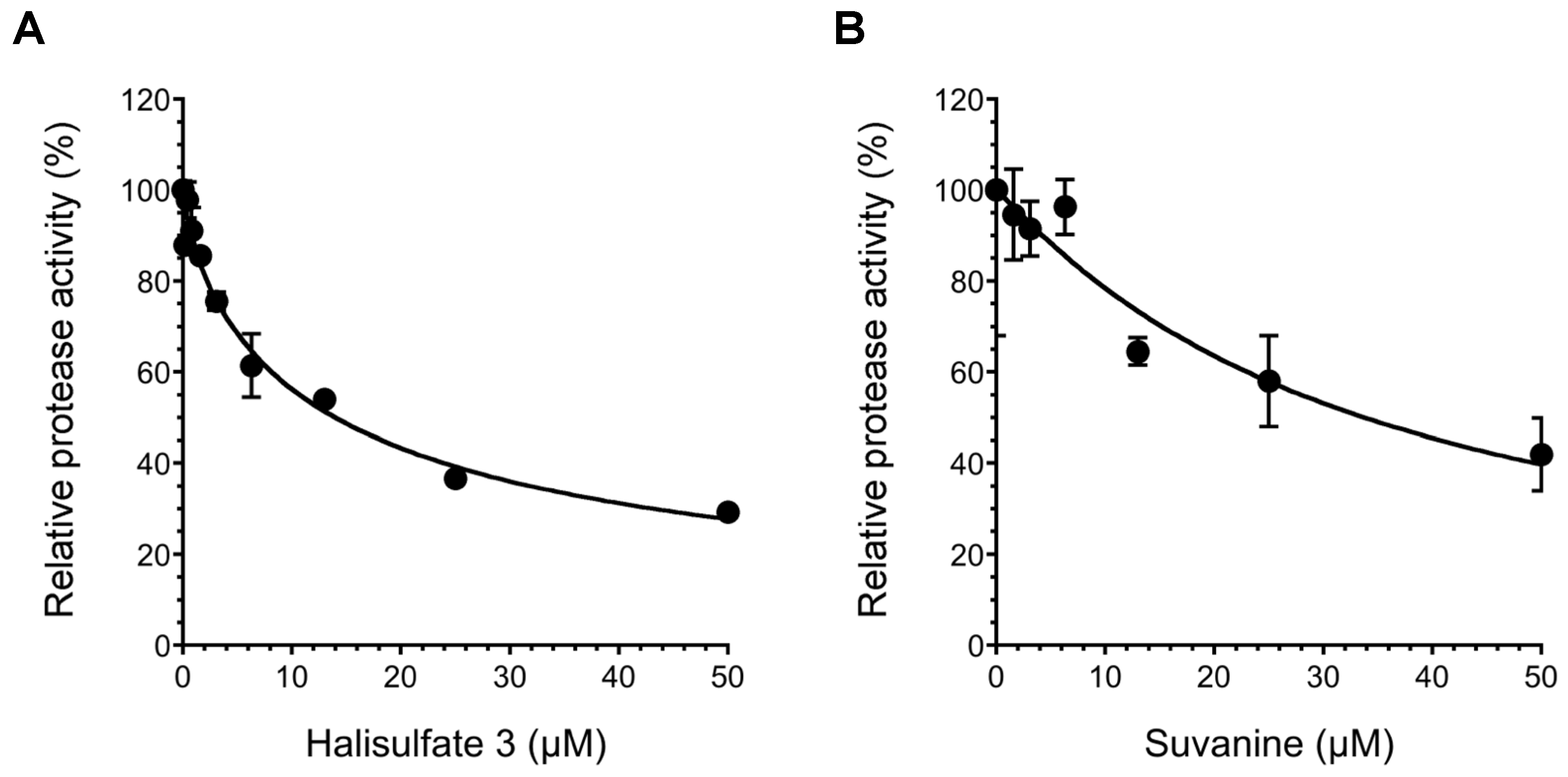
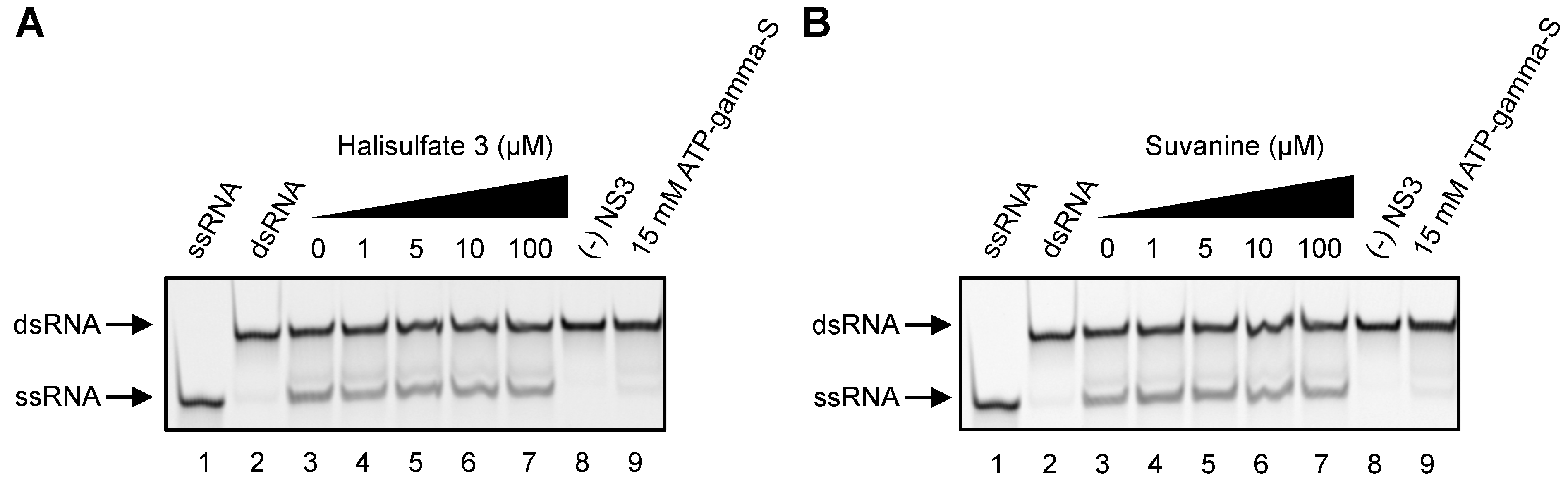
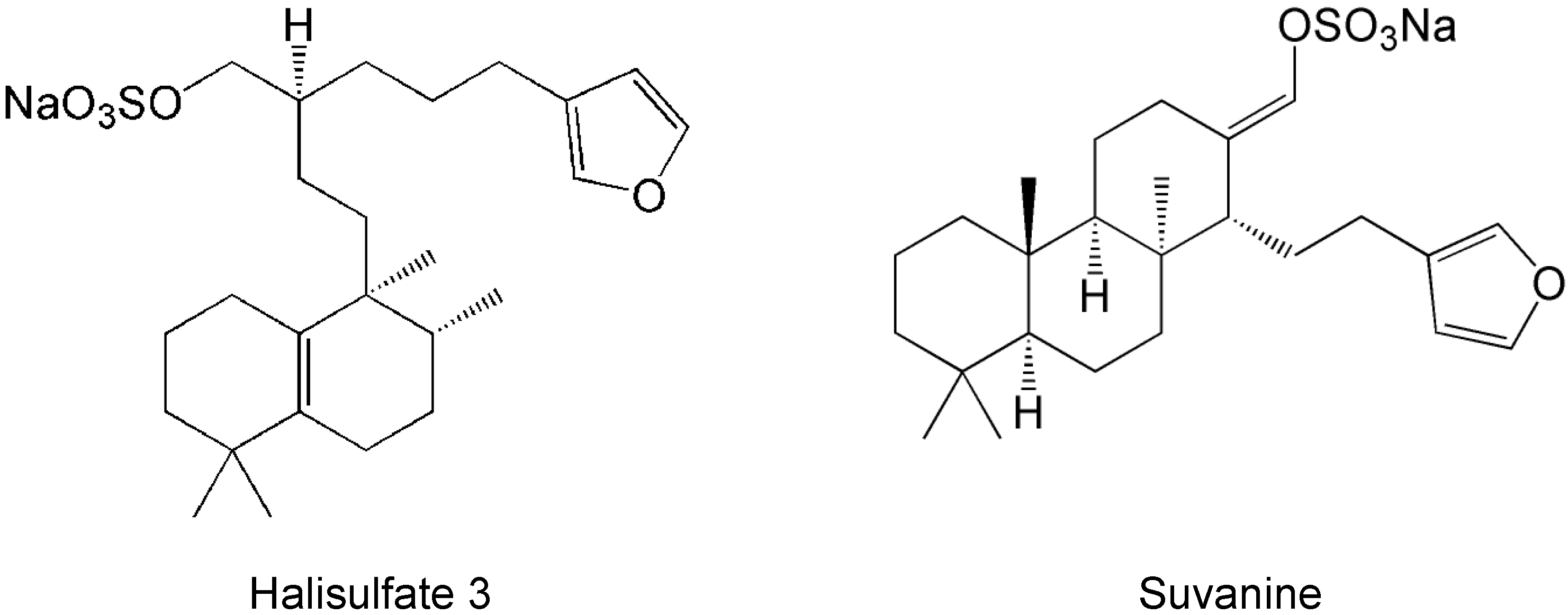
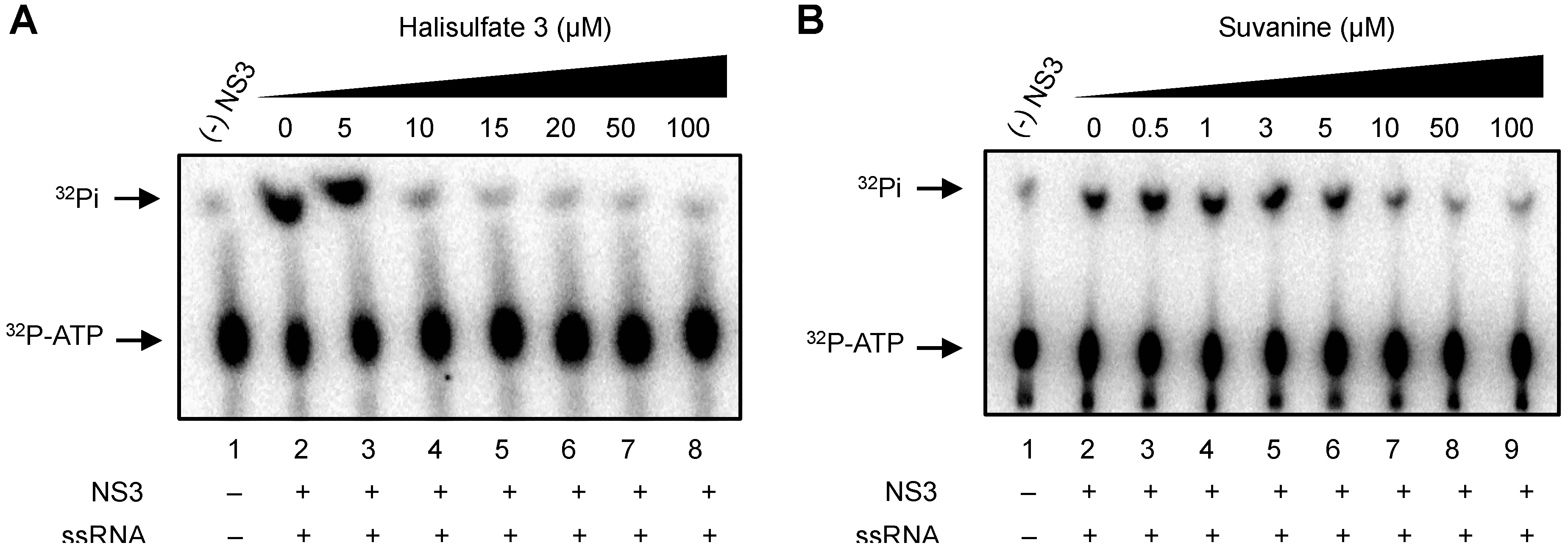
2. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | NS3 Helicase Activity (% of Control) * | Marine Organism | Species |
|---|---|---|---|
| 1 | 92 | Sponge | Unidentified |
| 2 | 74 | Soft coral | Briareum |
| 3 | 57 | Tunicate | Unidentified |
| 4 | 36 | Sponge | Liosina |
| 5 | 54 | Sponge | Unidentified |
| 6 | 71 | Sponge | Xestospongia |
| 7 | 77 | Sponge | Epipolasis |
| 8 | 110 | Sponge | Unidentified |
| 9 | 86 | Sponge | Strongylophora |
| 10 | 0 | Sponge | Unidentified |
| 11 | 83 | Sponge | Stylotella aurantium |
| 12 | 78 | Sponge | Epipolasis |
| 13 | 25 | Sponge | Unidentified |
| 14 | 43 | Sponge | Hippospongia |
| 15 | 75 | Sponge | Unidentified |
| 16 | 85 | Sponge | Unidentified |
| 17 | 49 | Sponge | Xestospongia testudinaria |
| 18 | 69 | Sponge | Unidentified |
| 19 | 40 | Sponge | Theonella |
| 20 | 64 | Sponge | Unidentified |
| 21 | 44 | Sponge | Unidentified |
| 22 | 46 | Sponge | Petrosia |
| 23 | 72 | Tunicate | Unidentified |
| 24 | 61 | Sponge | Unidentified |
| 25 | 50 | Tunicate | Didemnum molle |
| 26 | 33 | Sponge | Unidentified |
| 27 | 67 | Sponge | Unidentified |
| 28 | 87 | Soft coral | Unidentified |
| 29 | 62 | Sponge | Unidentified |
| 30 | 60 | Sponge | Unidentified |
| 31 | 85 | Sponge | Cinachyra |
| 32 | 70 | Sponge | Liosina |
| 33 | 68 | Sponge | Unidentified |
| 34 | 58 | Sponge | Unidentified |
| 35 | 72 | Sponge | Stylotella |
| 36 | 57 | Sponge | Unidentified |
| 37 | 39 | Sponge | Unidentified |
| 38 | 72 | Tunicate | Didemnum |
| 39 | 62 | Sponge | Unidentified |
| 40 | 71 | Jellyfish | Unidentified |
| 41 | 74 | Sponge | Unidentified |
| 42 | 52 | Tunicate | Unidentified |
| 43 | 67 | Annelid | Unidentified |

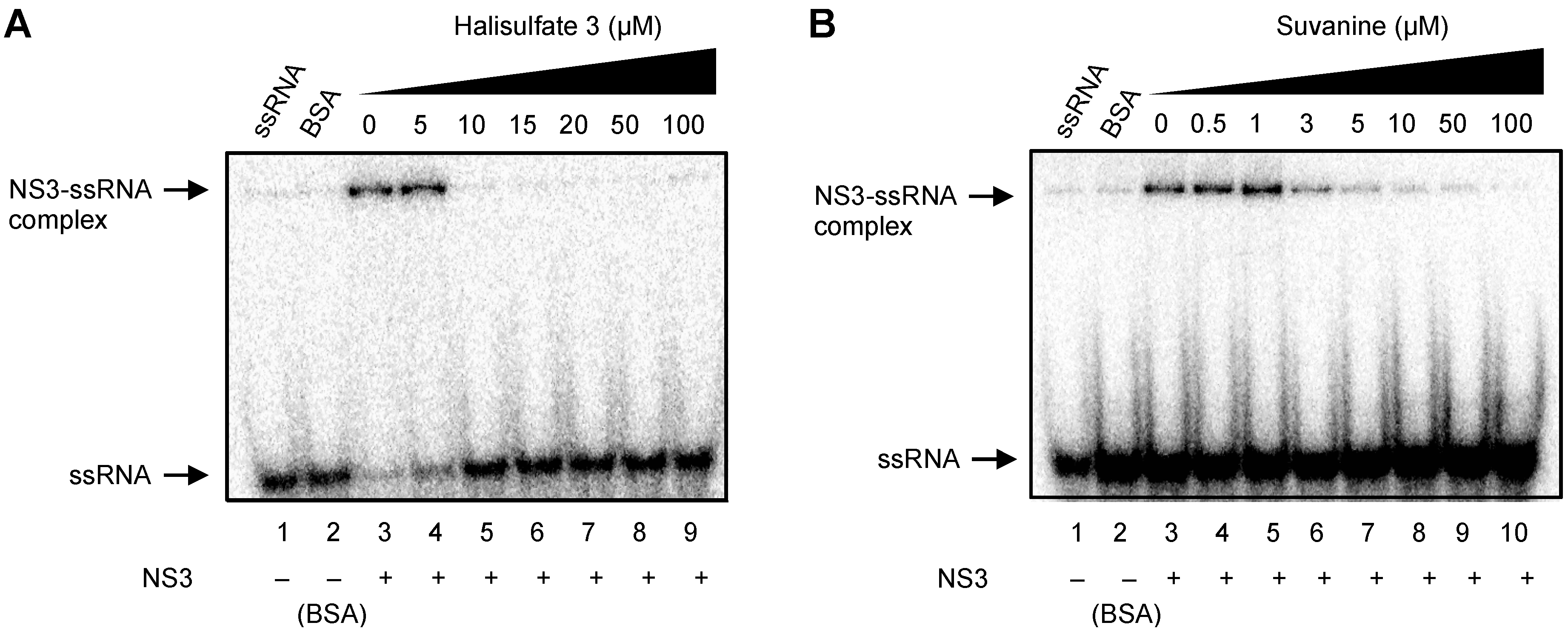
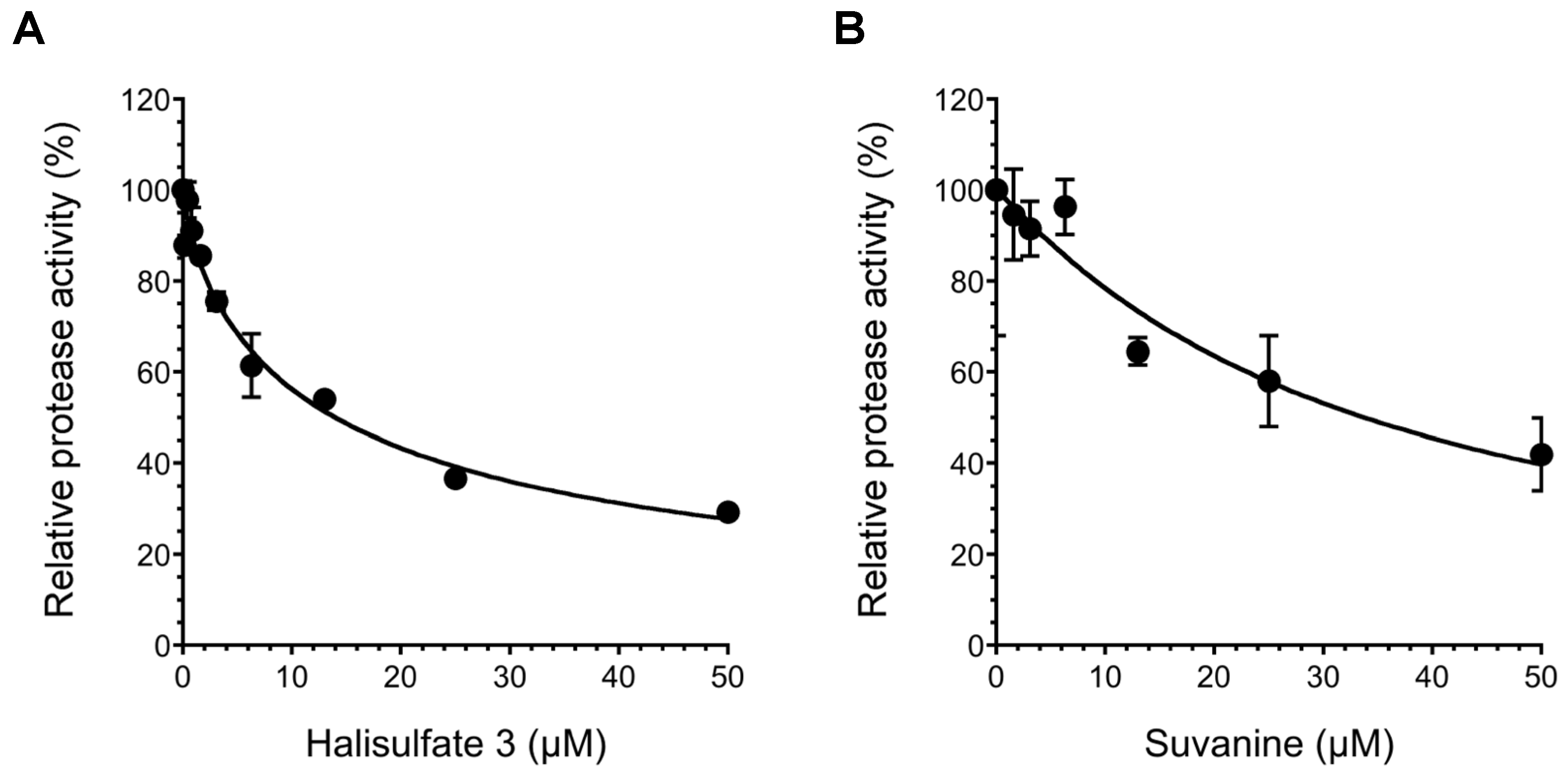
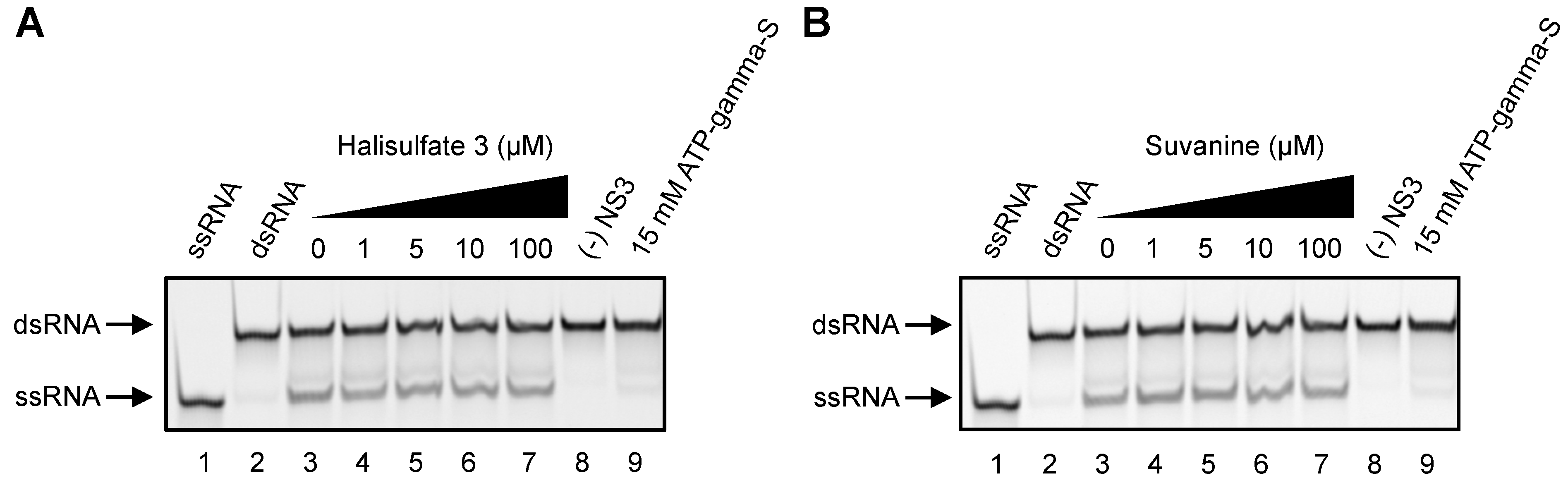
3. Experimental Section
3.1. Preparation of Extracts from Marine Organisms
3.2. Screening for HCV NS3 Helicase Inhibitors
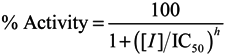
3.3. Gel-Based HCV NS3 Helicase Assay
3.4. ATPase Assay
3.5. RNA Binding Assay
3.6. Serine Protease Assay
3.7. Gel-Based DENV NS3 Helicase Assay
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ghany, M.G.; Nelson, D.R.; Strader, D.B.; Thomas, D.L.; Seeff, L.B. An update on treatment of genotype 1 chronic hepatitis C virus infection: 2011 practice guideline by the American Association for the Study of Liver Diseases. Hepatology 2011, 54, 1433–1444. [Google Scholar] [CrossRef]
- Liang, T.J.; Ghany, M.G. Current and future therapies for hepatitis C virus infection. N. Engl. J. Med. 2013, 368, 1907–1917. [Google Scholar] [CrossRef]
- Sarrazin, C.; Hézode, C.; Zeuzem, S.; Pawlotsky, J.-M. Antiviral strategies in hepatitis C virus infection. J. Hepatol. 2012, 56, S88–S100. [Google Scholar]
- Scheel, T.K.H.; Rice, C.M. Understanding the hepatitis C virus life cycle paves the way for highly effective therapies. Nat. Med. 2013, 19, 837–849. [Google Scholar] [CrossRef]
- Lam, A.M.I.; Frick, D.N. Hepatitis C virus subgenomic replicon requires an active NS3 RNA helicase. J. Virol. 2006, 80, 404–411. [Google Scholar] [CrossRef]
- Kwong, A.D.; Rao, B.G.; Jeang, K.-T. Viral and cellular RNA helicases as antiviral targets. Nat. Rev. Drug Discov. 2005, 4, 845–853. [Google Scholar] [CrossRef]
- Bartenschlager, R.; Penin, F.; Lohmann, V.; André, P. Assembly of infectious hepatitis C virus particles. Trends Microbiol. 2011, 19, 95–103. [Google Scholar] [CrossRef]
- Bartenschlager, R.; Lohmann, V.; Penin, F. The molecular and structural basis of advanced antiviral therapy for hepatitis C virus infection. Nat. Rev. Microbiol. 2013, 11, 482–496. [Google Scholar] [CrossRef]
- Lohmann, V.; Körner, F.; Koch, J.-O.; Herian, U.; Theilmann, L.; Bartenschlager, R. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science 1999, 285, 110–113. [Google Scholar] [CrossRef]
- Gallinari, P.; Brennan, D.; Nardi, C.; Brunetti, M.; Tomei, L.; Steinkühler, C.; de Francesco, R. Multiple enzymatic activities associated with recombinant NS3 protein of hepatitis C virus. J. Virol. 1998, 72, 6758–6769. [Google Scholar]
- Kim, D.W.; Gwack, Y.; Han, J.H.; Choe, J. C-terminal domain of the hepatitis C virus NS3 protein contains an RNA helicase activity. Biochem. Biophys. Res. Commun. 1995, 215, 160–166. [Google Scholar] [CrossRef]
- Tai, C.-L.; Chi, W.-K.; Chen, D.-S.; Hwang, L.-H. The helicase activity associated with hepatitis C virus nonstructural protein 3 (NS3). J. Virol. 1996, 70, 8477–8484. [Google Scholar]
- Gwack, Y.; Kim, D.W.; Han, J.H.; Choe, J. Characterization of RNA binding activity and RNA helicase activity of the hepatitis C virus NS3 protein. Biochem. Biophys. Res. Commun. 1996, 225, 654–659. [Google Scholar] [CrossRef]
- Gwack, Y.; Kim, D.W.; Han, J.H.; Choe, J. DNA helicase activity of the hepatitis C virus nonstructural protein 3. Eur. J. Biochem. 1997, 250, 47–54. [Google Scholar]
- Kolykhalov, A.A.; Mihalik, K.; Feinstone, S.M.; Rice, C.M. Hepatitis C virus-encoded enzymatic activities and conserved RNA elements in the 3′ nontranslated region are essential for virus replication in vivo. J. Virol. 2000, 74, 2046–2051. [Google Scholar] [CrossRef]
- Hall, M.C.; Matson, S.W. Helicase motifs: The engine that powers DNA unwinding. Mol. Microbiol. 1999, 34, 867–877. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Koonin, E.V. Helicases: Amino acid sequence comparisons and structure-function relationships. Curr. Opin. Struct. Biol. 1993, 3, 419–429. [Google Scholar] [CrossRef]
- Jankowsky, E. RNA Helicases; Royal Society of Chemistry: London, UK, 2010; pp. 168–188. [Google Scholar]
- Frick, D.N.; Lam, A.M.I. Understanding helicases as a means of virus control. Curr. Pharm. Des. 2006, 12, 1315–1338. [Google Scholar] [CrossRef]
- Borowski, P.; Deinert, J.; Schalinski, S.; Bretner, M.; Ginalski, K.; Kulikowski, T.; Shugar, D. Halogenated benzimidazoles and benzotriazoles as inhibitors of the NTPase/helicase activities of hepatitis C and related viruses. Eur. J. Biochem. 2003, 270, 1645–1653. [Google Scholar] [CrossRef]
- Zhang, N.; Chen, H.-M.; Koch, V.; Schmitz, H.; Liao, C.-L.; Bretner, M.; Bhadti, V.S.; Fattom, A.I.; Naso, R.B.; Hosmane, R.S.; et al. Ring-expanded (“fat”) nucleoside and nucleotide analogues exhibit potent in vitro activity against Flaviviridae NTPases/helicases, including those of the West Nile virus, hepatitis C virus, and Japanese encephalitis virus. J. Med. Chem. 2003, 46, 4149–4164. [Google Scholar] [CrossRef]
- Borowski, P.; Heising, M.V.; Miranda, I.B.; Liao, C.-L.; Choe, J.; Baier, A. Viral NS3 helicase activity is inhibited by peptides reproducing the Arg-rich conserved motif of the enzyme (motif VI). Biochem. Pharmacol. 2008, 76, 28–38. [Google Scholar] [CrossRef]
- Frick, D.N.; Ginzburg, O.; Lam, A.M.I. A method to simultaneously monitor hepatitis C virus NS3 helicase and protease activities. Methods Mol. Biol. 2010, 587, 223–233. [Google Scholar] [CrossRef]
- Dahl, G.; Sandström, A.; Akerblom, E.; Danielson, U.H. Effects on protease inhibition by modifying of helicase residues in hepatitis C virus nonstructural protein 3. FEBS J. 2007, 274, 5979–5986. [Google Scholar] [CrossRef]
- Frick, D.N. The hepatitis C virus NS3 protein: A model RNA helicase and potential drug target. Curr. Issues Mol. Biol. 2007, 9, 1–20. [Google Scholar]
- Belon, C.A.; Frick, D.N. Helicase inhibitors as specifically targeted antiviral therapy for hepatitis C. Future Virol. 2009, 4, 277–293. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of marine natural products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Furuta, A.; Salam, K.A.; Akimitsu, N.; Tanaka, J.; Tani, H.; Yamashita, A.; Moriishi, K.; Nakakoshi, M.; Tsubuki, M.; Sekiguchi, Y.; et al. Cholesterol sulfate as a potential inhibitor of hepatitis C virus NS3 helicase. J. Enzym. Inhib. Med. Chem. 2013. [Google Scholar] [CrossRef]
- Müller, E.L.; Faulkner, D.J. Absolute configuration of halisulfate 3 from the sponge Ircinia sp. Tetrahedron 1997, 53, 5373–5378. [Google Scholar] [CrossRef]
- Manes, L.V.; Crews, P.; Kernan, M.R.; Faulkner, D.J.; Fronczek, F.R.; Gandour, R.D. Chemistry and revised structure of suvanine. J. Org. Chem. 1988, 53, 570–575. [Google Scholar] [CrossRef]
- Kernan, M.R.; Faulkner, D.J. Sesterterpene sulfates from a sponge of the family Halichondriidae. J. Org. Chem. 1988, 53, 4574–4578. [Google Scholar] [CrossRef]
- Manes, L.V.; Naylor, S.; Crews, P.; Bakus, G.J. Suvanine, a novel sesterterpene from an Ircinia marine sponge. J. Org. Chem. 1985, 50, 284–286. [Google Scholar] [CrossRef]
- Kimura, J.; Ishizuka, E.; Nakao, Y.; Yoshida, W.Y.; Scheuer, P.J.; Kelly-Borges, M. Isolation of 1-methylherbipoline salts of halisulfate-1 and of suvanine as serine protease inhibitors from a marine sponge, Coscinoderma mathewsi. J. Nat. Prod. 1998, 61, 248–250. [Google Scholar] [CrossRef]
- Di Leva, F.S.; Festa, C.; D’Amore, C.; de Marino, S.; Renga, B.; D’Auria, M.V.; Novellino, E.; Limongelli, V.; Zampella, A.; Fiorucci, S. Binding mechanism of the farnesoid X receptor marine antagonist suvanine reveals a strategy to forestall drug modulation on nuclear receptors. Design, synthesis, and biological evaluation of novel ligands. J. Med. Chem. 2013, 56, 4701–4717. [Google Scholar] [CrossRef]
- Cassiano, C.; Monti, M.C.; Festa, C.; Zampella, A.; Riccio, R.; Casapullo, A. Chemical proteomics reveals heat shock protein 60 to be the main cellular target of the marine bioactive sesterterpene suvanine. ChemBioChem 2012, 13, 1953–1958. [Google Scholar] [CrossRef]
- Singleton, M.R.; Dillingham, M.S.; Wigley, D.B. Structure and mechanism of helicases and nucleic acid translocases. Annu. Rev. Biochem. 2007, 76, 23–50. [Google Scholar] [CrossRef]
- Wang, C.-C.; Huang, Z.-S.; Chiang, P.-L.; Chen, C.-T.; Wu, H.-N. Analysis of the nucleoside triphosphatase, RNA triphosphatase, and unwinding activities of the helicase domain of dengue virus NS3 protein. FEBS Lett. 2009, 583, 691–696. [Google Scholar] [CrossRef]
- Ndjomou, J.; Kolli, R.; Mukherjee, S.; Shadrick, W.R.; Hanson, A.M.; Sweeney, N.L.; Bartczak, D.; Li, K.; Frankowski, K.J.; Schoenen, F.J.; et al. Fluorescent primuline derivatives inhibit hepatitis C virus NS3-catalyzed RNA unwinding, peptide hydrolysis and viral replicase formation. Antivir. Res. 2012, 96, 245–255. [Google Scholar] [CrossRef]
- Saalau-Bethell, S.M.; Woodhead, A.J.; Chessari, G.; Carr, M.G.; Coyle, J.; Graham, B.; Hiscock, S.D.; Murray, C.W.; Pathuri, P.; Rich, S.J.; et al. Discovery of an allosteric mechanism for the regulation of HCV NS3 protein function. Nat. Chem. Biol. 2012, 8, 920–925. [Google Scholar]
- Ding, S.C.; Kohlway, A.S.; Pyle, A.M. Unmasking the active helicase conformation of nonstructural protein 3 from hepatitis C virus. J. Virol. 2011, 85, 4343–4353. [Google Scholar] [CrossRef]
- Tani, H.; Akimitsu, N.; Fujita, O.; Matsuda, Y.; Miyata, R.; Tsuneda, S.; Igarashi, M.; Sekiguchi, Y.; Noda, N. High-throughput screening assay of hepatitis C virus helicase inhibitors using fluorescence-quenching phenomenon. Biochem. Biophys. Res. Commun. 2009, 379, 1054–1059. [Google Scholar] [CrossRef]
- Copeland, R.A. Evaluation of Enzyme Inhibitors in Drug Discovery; John Wiley & Sons: New York, NY, USA, 2005; pp. 111–140. [Google Scholar]
- Salam, K.A.; Furuta, A.; Noda, N.; Tsuneda, S.; Sekiguchi, Y.; Yamashita, A.; Moriishi, K.; Nakakoshi, M.; Tsubuki, M.; Tani, H.; et al. Inhibition of hepatitis C virus NS3 helicase by manoalide. J. Nat. Prod. 2012, 75, 650–654. [Google Scholar] [CrossRef]
- Salam, K.A.; Furuta, A.; Noda, N.; Tsuneda, S.; Sekiguchi, Y.; Yamashita, A.; Moriishi, K.; Nakakoshi, M.; Tsubuki, M.; Tani, H.; et al. Psammaplin A inhibits hepatitis C virus NS3 helicase. J. Nat. Med. 2013, 67, 765–772. [Google Scholar] [CrossRef]
- Takahashi, H.; Takahashi, C.; Moreland, N.J.; Chang, Y.-T.; Sawasaki, T.; Ryo, A.; Vasudevan, S.G.; Suzuki, Y.; Yamamoto, N. Establishment of a robust dengue virus NS3-NS5 binding assay for identification of protein-protein interaction inhibitors. Antivir. Res. 2012, 96, 305–314. [Google Scholar] [CrossRef]
Supplementary Files
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Furuta, A.; Salam, K.A.; Hermawan, I.; Akimitsu, N.; Tanaka, J.; Tani, H.; Yamashita, A.; Moriishi, K.; Nakakoshi, M.; Tsubuki, M.; et al. Identification and Biochemical Characterization of Halisulfate 3 and Suvanine as Novel Inhibitors of Hepatitis C Virus NS3 Helicase from a Marine Sponge. Mar. Drugs 2014, 12, 462-476. https://doi.org/10.3390/md12010462
Furuta A, Salam KA, Hermawan I, Akimitsu N, Tanaka J, Tani H, Yamashita A, Moriishi K, Nakakoshi M, Tsubuki M, et al. Identification and Biochemical Characterization of Halisulfate 3 and Suvanine as Novel Inhibitors of Hepatitis C Virus NS3 Helicase from a Marine Sponge. Marine Drugs. 2014; 12(1):462-476. https://doi.org/10.3390/md12010462
Chicago/Turabian StyleFuruta, Atsushi, Kazi Abdus Salam, Idam Hermawan, Nobuyoshi Akimitsu, Junichi Tanaka, Hidenori Tani, Atsuya Yamashita, Kohji Moriishi, Masamichi Nakakoshi, Masayoshi Tsubuki, and et al. 2014. "Identification and Biochemical Characterization of Halisulfate 3 and Suvanine as Novel Inhibitors of Hepatitis C Virus NS3 Helicase from a Marine Sponge" Marine Drugs 12, no. 1: 462-476. https://doi.org/10.3390/md12010462