Ontogenetic Pattern Changes of Nucleobindin-2/Nesfatin-1 in the Brain and Intestinal Bulb of the Short Lived African Turquoise Killifish

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protocols and Ethics Statement
2.2. Animals and Tissue Preparation
2.3. Sequence Analysis
2.4. RNA Extraction and Reverse Transcription of cDNA Synthesis
2.5. Quantitative Real Time PCR
2.6. Statistical Analysis
2.7. Riboprobes Synthesis
2.8. In Situ Hybridization
2.9. Western Blot
2.10. Immunohistochemistry
2.11. Controls of Specificity
2.12. Image Acquisition
3. Results
3.1. Sequence Analysis and Antibody Specificity
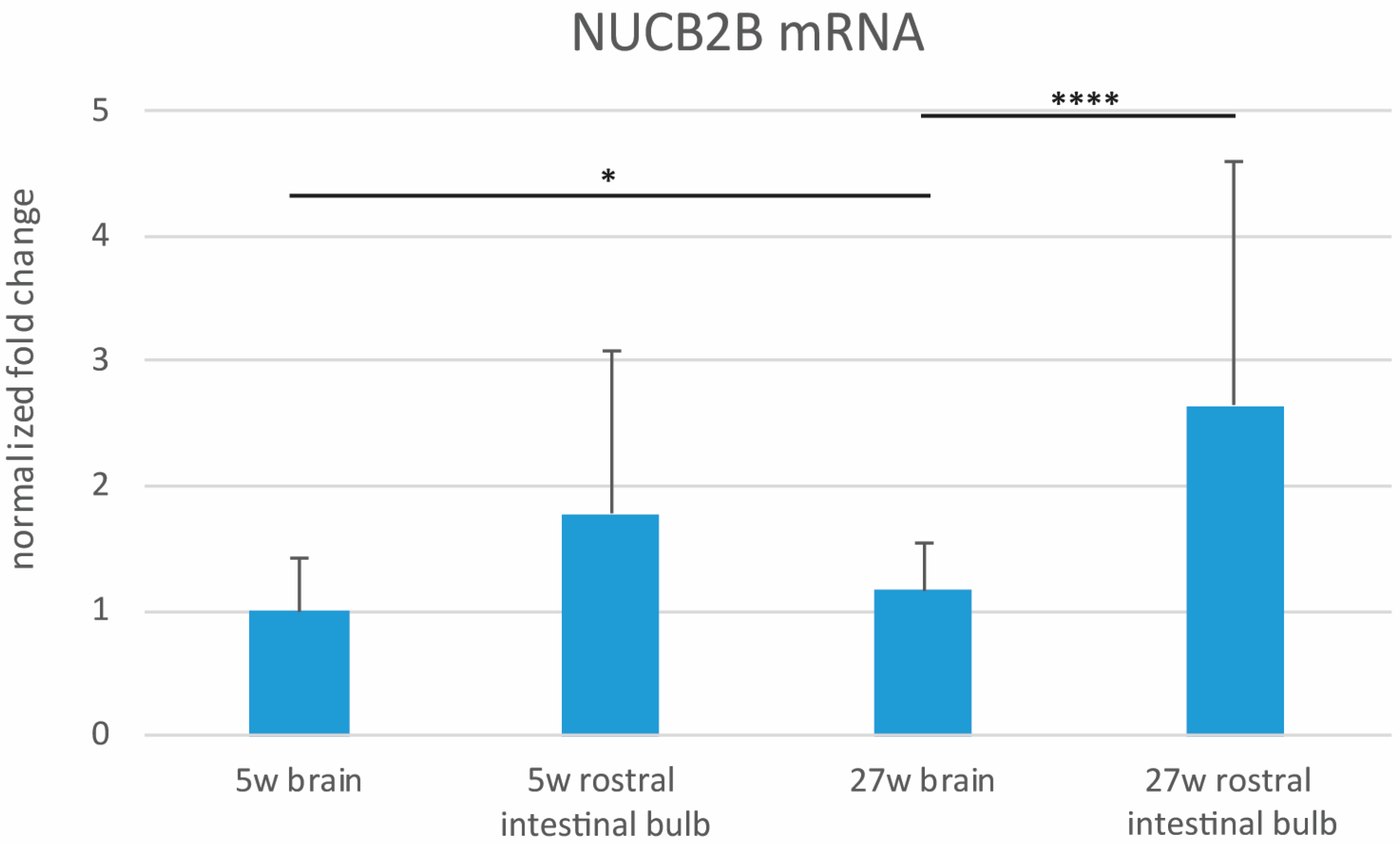
3.2. Expression Levels of NUCB2B mRNA in the Whole Brain and in the Rostral Intestinal Bulb of Young and Old Animals
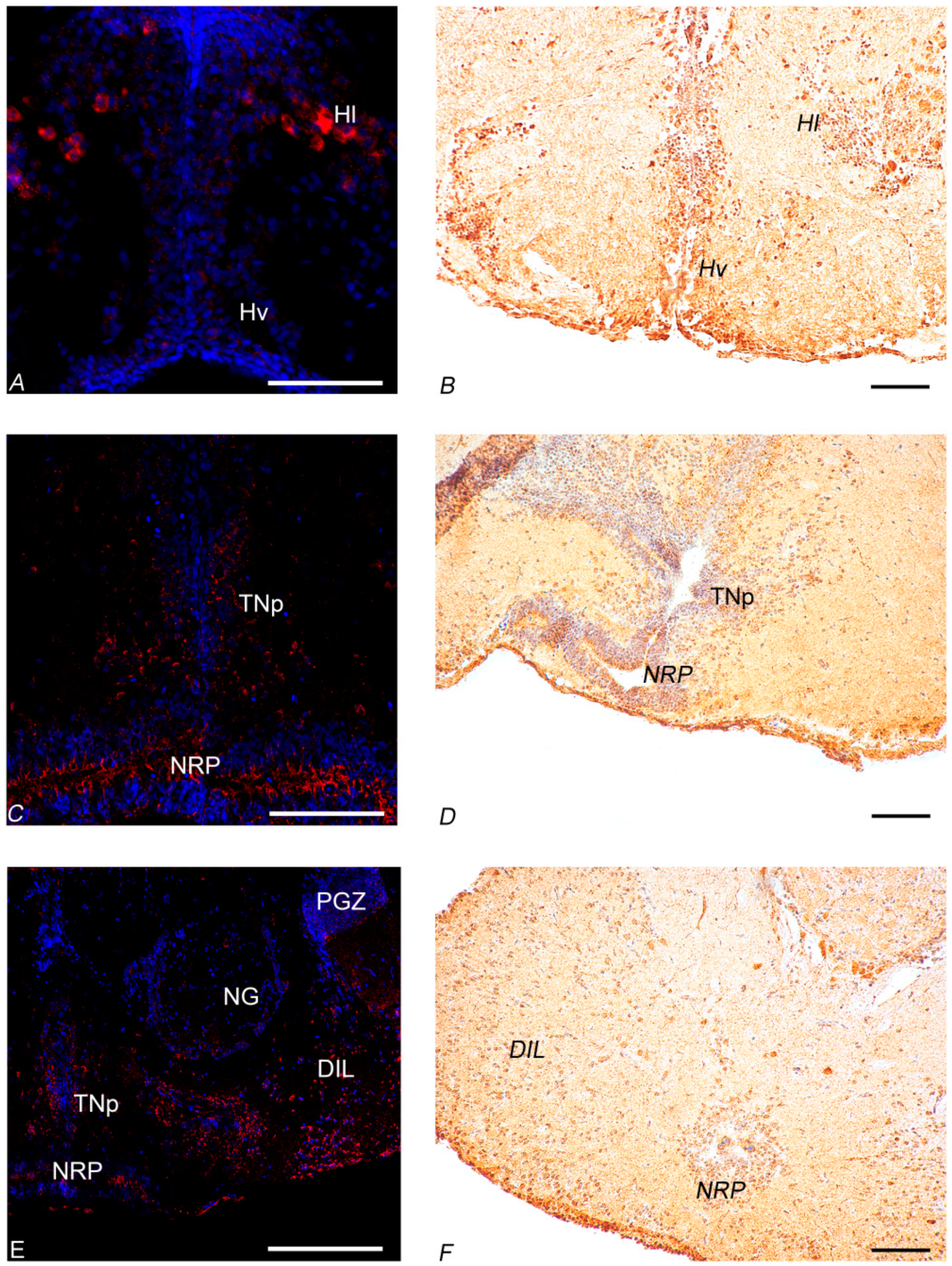
3.3. Morphological Studies of the Whole Brain of Young and Old Animals
3.3.1. In Situ Hybridization
3.3.2. Immunohistochemistry
3.4. Morphological Studies of the Intestine of Young and Old Animals
3.4.1. In Situ Hybridization
3.4.2. Immunohistochemistry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Dc | central zone of dorsal telencephalon |
| Dld | dorso-lateral zone of dorsal telencephalon |
| Dll | latero-lateral zone of dorsal telencephalon |
| Dlv | ventro-lateral zone of dorsal telencephalon |
| Dm | medial zone of dorsal telencephalon |
| DIL | inferior lobe of hypothalamus |
| Hd | dorsal hypothalamus |
| Hl | lateral hypothalamus |
| Hv | ventral hypothalamus |
| NG | glomerular nucleus |
| NRP | nucleus of posterior recess |
| OT | optic tectum |
| PGZ | periventricular grey zone of Optic Tectum |
| PPa | anterior preoptic nucleus |
| TNp | posterior tuberal nucleus |
| TS | semicircular tori |
| VM | ventro-medial thalamic nucleus |
| Vp | posterior zone of ventral telencephalon |
| Vs | supracommisural zone of ventral telencephalon |
References
- Gonzalez, R.; Mohan, H.; Unniappan, S. Nucleobindins: Bioactive precursor proteins encoding putative endocrine factors? Gen. Com. Endocrin. 2012, 176, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Brailoiu, G.C.; Deliu, E.; Tica, A.A.; Rabinowitz, J.E.; Tilley, D.G.; Benamar, K.; Koch, W.J.; Brailoiu, E. Nesfatin-1 activates cardiac vagal neurons of nucleus ambiguus and elicits bradycardia in conscious rats. J. Neurochem. 2013, 126, 739–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh-I, S.; Shimizu, H.; Satoh, T.; Okada, S.; Adachi, S.; Inoue, K.; Eguchi, H.; Yamamoto, M.; Imaki, T.; Hashimoto, K.; et al. Identification of nesfatin-1 as a satiety molecule in the hypothalamus. Nature 2006, 443, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Mohan, H.; Unniappan, S. Phylogenetic aspects of nucleobindin-2/nesfatin-1. Curr. Pharm. Des. 2013, 19, 6929–6934. [Google Scholar] [CrossRef] [PubMed]
- Palasz, A.; Krzystanek, M.; Worthington, J.; Czajkowska, B.; Kostro, K.; Wiaderkiewicz, R.; Bajor, G. Nesfatin-1, a unique regulatory neuropeptide of the brain. Neuropeptides 2012, 46, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Kohno, D.; Nakata, M.; Maejima, Y.; Shimizu, H.; Sedbazar, U.; Yoshida, N.; Dezaki, K.; Onaka, T.; Mori, M.; Yada, T. Nesfatin-1 neurons in paraventricular and supraoptic nuclei of the rat hypothalamus coexpress oxytocin and vasopressin and are activated by refeeding. Endocrinology 2008, 149, 1295–1301. [Google Scholar] [CrossRef]
- Brunner, L.; Nick, H.P.; Cumin, F.; Chiesi, M.; Baum, H.P.; Whitebread, S.; Stricker-Krongrad, A.; Levens, N. Leptin is a physiologically important regulator of food intake. Int. J. Obes. Relat. Metab. Disord. 1997, 21, 1152–1160. [Google Scholar] [CrossRef] [Green Version]
- Ramanjaneya, M.; Chen, J.; Brown, J.E.; Tripathi, G.; Hallschmid, M.; Patel, S.; Kern, W.; Hillhouse, E.W.; Lehnert, H.; Tan, B.K.; et al. Identification of nesfatin-1 in human and murine adipose tissue: A novel depot-specific adipokine with increased levels in obesity. Endocrinology 2010, 151, 3169–3180. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.Q.; Li, X.L.; Jiang, C.Y.; Lin, L.; Shi, R.H.; Chen, J.D.; Oomura, Y. Expression of nesfatin-1/NUCB2 in rodent digestive system. World J. Gastroenterol. 2010, 6, 1735–1741. [Google Scholar] [CrossRef]
- Gatta, C.; De Felice, E.; D’Angelo, L.; Maruccio, L.; Leggieri, A.; Lucini, C.; Palladino, A.; Paolucci, M.; Scocco, P.; Varricchio, E.; et al. The case study of Nesfatin-1 in the pancreas of Tursiops truncatus. Front. Physiol. 2018, 9, 1845. [Google Scholar] [CrossRef]
- Stengel, A.; Goebel, M.; Yakubov, I.; Wang, L.; Witcher, D.; Coskun, T.; Taché, Y.; Sachs, G.; Lambrecht, N.W. Identification and characterization of nesfatin-1immunoreactivity in endocrine cell types of the rat gastric oxyntic mucosa. Endocrinology 2009, 150, 232–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stengel, A.; Goebel-Stengel, M.; Jawien, J.; Kobelt, P.; Taché, Y.; Lambrecht, N.W. Lipopolysaccharide increases gastric and circulating NUCB2/nesfatin-1 concentrations in rats. Peptides 2011, 32, 1942–1947. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Mulholland, M.; Zhang, W. Regulation of gastric nesfatin-1/NUCB2. Curr. Pharm. Des. 2013, 19, 6981–6985. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Kerbel, B.; Chun, A.; Unniappan, S. Molecular, cellular and physiological evidences for the anorexigenic actions of nesfatin-1 in goldfish. PLoS ONE 2010, 5, e15201. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zhou, C.; Chen, H.; Wu, H.; Xin, Z.; Liu, J.; Gao, Y.; Yuan, D.; Wang, T.; Wei, R.; et al. Molecular characterization, tissue distribution and feeding related changes of NUCB2A/nesfatin-1 in Ya-fish (Schizothoraxprenanti). Gene 2014, 536, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Hatef, A.; Shajan, S.; Unniappan, S. Nutrient status modulates the expression of nesfatin-1 encoding nucleobindin 2A and 2B mRNAs in zebrafish gut, liver and brain. Gen. Comp. Endocrin. 2015, 215, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Kerbel, B.; Unniappan, S. Nesfatin-1 suppresses energy intake, co-localises ghrelin in the brain and gut, and alters ghrelin, cholecystokinin and orexin mRNA expression in goldfish. J. Neuroendocr. 2011, 24, 366–377. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Canosa, L.F.; Unniappan, S. Glucose, amino acids and fatty acids directly regulate ghrelin and NUCB2/nesfatin-1 in the intestine and hepatopancreas of goldfish (Cariassus auratus) in vitro. Comp. Biochem. Physiol. Part A 2017, 206, 24–35. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Sanchez-Bretano, A.; Unniappan, S.; Canosa, L.F. Ghrelin and NUCB2/Nesfatin-1 co-localization with digestive enzymes in the intestine of Pejerrey (Odontesthes bonariensis). Anat. Rec. 2019, 302, 973–982. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, J.; Tang, N.; Wang, S.; Wu, Y.; Chen, H.; Tian, Z.; Wang, B.; Chen, D.; Li, Z. Intraperitoneal injection of nesfatin-1 primarily through the CCK-CCK1R signal pathway affects expression of appetite factors to inhibit the food intake of Siberian sturgeon (Acipenser baerii). Peptides 2018, 109, 14–22. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, S.; Chen, H.; Tang, N.; Qi, J.; Wu, Y.; Hao, J.; Tian, Z.; Wang, B.; Chen, D.; et al. The inhibitory effect of NUCB2/nesfatin-1 on appetite regulation of Siberian sturgeon (Acipenser baerii Brandt). Hormon. Behav. 2018, 103, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Mohan, H.; Unniappan, S. Ontogenic pattern of nucleobindin-2/nesfatin-1 expression in the gastroenteropancreatic tissues and serum of Sprague Dawley rats. Reg. Pept. 2012, 175, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Senin, L.; Al-Massadi, O.; Barja-Fernandez, S.; Folgueira, C.; Castelao, C.; Tovar, S.A.; Leis, R.; Lago, F.; Baltar, J.; Baamonde, I.; et al. Regulation of NUCB2/nesfatin-1production in rat’stomach and adipose tissue is dependent on age, testosterone levels and lactating status. Mol. Cell. Endocrinol. 2015, 411, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Terzibasi, E.; Valenzano, D.R.; Benedetti, M.; Roncaglia, P.; Cattaneo, A.; Domenici, L.; Cellerino, A. Large differences in aging phenotype between strains of the short-lived annual fish Nothobranchius furzeri. PLoS ONE 2008, 3, e3866. [Google Scholar] [CrossRef] [Green Version]
- Cellerino, A.; Valenzano, D.R.; Reichard, M. From the bush to the bench: The annual Nothobranchius fishes as a new model system in biology. Biol. Rev. Camb. Philos. Soc. 2016, 91, 511–533. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, L.; Lossi, L.; Merighi, A.; de Girolamo, P. Anatomical features for the adequate choice of experimental animal models in biomedicine: I. Fishes. Ann. Anat. Anat. Anz. 2016, 205, 75–84. [Google Scholar] [CrossRef]
- Genade, T.; Benedetti, M.; Terzibasi, E.; Roncaglia, P.; Valenzano, D.R.; Cattaneo, A.; Cellerino, A. Annual fishes of the genus Nothobranchius as a model system for aging research. Aging Cell 2005, 4, 223–233. [Google Scholar] [CrossRef]
- Montesano, A.; Baumgart, M.; Avallone, L.; Castaldo, L.; Lucini, C.; Tozzini, E.T.; Cellerino, A.; D’Angelo, L.; de Girolamo, P. Age-related central regulation of orexin and NPY in the short lived African killifish Notobranchius furzeri. J. Comp. Neurol. 2019, 527, 1508–1526. [Google Scholar] [CrossRef]
- Arcamone, N.; Neglia, S.; Gargiulo, G.; Esposito, V.; Varricchio, E.; Battaglini, P.; de Girolamo, P.; Russo, F. Distribution of ghrelin peptide in the gastrointestinal tract of stomachless and stomach-containing teleosts. Microsc. Res. Tech. 2009, 72, 525–533. [Google Scholar] [CrossRef]
- Russo, F.; de Girolamo, P.; Neglia, S.; Gargiulo, A.; Arcamone, N.; Gargiulo, G.; Varricchio, E. Immunohistochemical and immunochemical characterization of the distribution of leptin-like proteins in the gastroenteric tract of two teleosts (Dicentrarchus labrax and Carassius auratus L.) with different feeding habits. Microsc. Res. Tech. 2011, 74, 714–719. [Google Scholar] [CrossRef]
- D’Angelo, L.; Castaldo, L.; de Girolamo, P.; Lucini, C.; Paolucci, M.; Pelagalli, A.; Varricchio, E.; Arcamone, N. Orexins and receptor OX2R in the gastroenteric apparatus of two teleostean species: Dicentrarchu sLabrax and Carassius Auratus. Anat. Rec. 2016, 299, 1121–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkoff, H. Fish as models for understanding the vertebrate endocrine regulation of feeding and weight. Mol. Cell. Endocrinol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, M.; Groth, M.; Priebe, S.; Savino, A.; Testa, G.; Dix, A.; Ripa, R.; Spallotta, F.; Gaetano, C.; Ori, M.; et al. RNA-seq of the aging brain in the short-lived fish N. furzeri—conserved pathaways and novel genes associated with neurogenesis. Aging Cell 2014, 13, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Reichwald, K.; Petzold, A.; Koch, P.; Downie, B.R.; Hartmann, N.; Pietsch, S.; Baumgart, M.; Chalopin, D.; Felder, M.; Bens, M.; et al. Insights into sex chromosome evolution and aging from the genome of a short-lived fish. Cell 2015, 163, 1527–1538. [Google Scholar] [CrossRef] [Green Version]
- Hunt, S.E.; McLaren, W.; Laurent, G.; Thormann, A.; Schuilenburg, H.; Sheppard, D.; Parton, A.; Armean, I.M.; Trevanion, S.J.; Flicek, P.; et al. Ensembl variation resources. Database 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Rzhetsky, A.; Nei, M. A simple method for estimating and testing minimum evolution trees. Mol. Biol. Evol. 1992, 9, 945–967. [Google Scholar]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. [Google Scholar]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Geer, L.Y.; Marchler-Bauer, A.; Geer, R.C.; Han, L.; He, J.; He, S.; Liu, C.; Shi, W.; Bryant, S.H. The NCBI BioSystems database. Nucleic Acids Res. 2010, 38, D492–D496. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Baumgart, M.; Groth, M.; Priebe, S.; Appelt, J.; Guthke, R.; Platzer, M.; Cellerino, A. Age-dependent regulation of tumor-related microRNA in the brain of the annual fish Nothobranchius furzeri. Mech. Ageing Dev. 2012, 133, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, L. Brain atlas of an emerging teleostean model: Nothobranchiusfurzeri. Anat. Rec. 2013, 296, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.; Ramesh, N.; Vogel, C.; Unniappan, S. Nucleobindins and encoded peptides: From cell signaling to physiology. Adv. Prot. Chem. Struct. Biol. 2019, 116, 91–133. [Google Scholar] [CrossRef]
- Wilson, J.M.; Castro, L.F.C. Morphological diversity of the gastrointestinal tract in fishes. Fish Physiol. 2010, 30, 1–55. [Google Scholar]
- Kapoor, B.G.; Smit, H.; Verighina, I.A. The alimentary canal and digestion in teleosts. Adv. Mar. Biol. 1976, 13, 109–239. [Google Scholar]
- Taylor, J.S.; Braasch, I.; Frickey, T.; Meyer, A.; Van de Peer, Y. Genome duplication, a trait shared by 22000 species of ray-finned fish. Genome Res. 2003, 13, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Brailoiu, G.C.; Dun, S.L.; Brailoiu, E.; Inan, S.; Yang, J.; Chang, J.K.; Dun, N.J. Nesfatin-1: Distribution and interaction with a G protein-coupled receptor in the rat brain. Endocrinology 2007, 148, 5088–5094. [Google Scholar] [CrossRef] [Green Version]
- Fort, P.; Salvert, D.; Hanriot, L.; Jego, S.; Shimizu, H.; Hashimoto, K.; Mori, M.; Luppi, P.H. The satiety molecule nesfatin-1 is co-expressed with melanin concentrating hormone in tuberal hypothalamic neurons of the rat. Neuroscience 2008, 155, 174–181. [Google Scholar] [CrossRef]
- Nonogaki, K.; Ohba, Y.; Sumii, M.; Oka, Y. Serotonin systems upregulate the expression of hypothalamic NUCB2 via 5-HT2C receptors and induce anorexia via a leptin-independent pathway in mice. Biochem. Biophys. Res. Commun. 2008, 372, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Foo, K.S.; Brismar, H.; Broberger, C. Distribution and neuropeptide coexistence of nucleobindin-2 mRNA/nesfatin-like immunoreactivity in the rat CNS. Neuroscience 2008, 156, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Senejani, A.G.; Gaupale, T.C.; Unniappan, S.; Bhargava, S. Nesfatin-1/nucleobindin-2 like immunoreactivity in the olfactory system, brain and pituitary of frog, Microhyla ornata. Gen. Comp. Endocrin. 2014, 202, 8–14. [Google Scholar] [CrossRef]
- Volkoff, H.; Canosa, L.F.; Unniappan, S.; Cerdá-Reverter, J.M.; Bernier, N.J.; Kelly, S.P.; Peter, R.E. Neuropeptides and the control of food intake in fish. Gen. Comp. Endocrin. 2005, 142, 3–19. [Google Scholar] [CrossRef]
- Demski, L.S.; Knigge, K.M. The telencephalon and hypothalamus of the bluegill (Lepomis macrochirus): Evoked feeding, aggressive and reproductive behavior with representative frontal sections. J. Comp. Neur. 1971, 143, 1–16. [Google Scholar] [CrossRef]
- Stacey, N.E.; Kyle, A.L. Effects of olfactory tract lesions on sexual and feeding behavior in the goldfish. Physiol. Behav. 1983, 30, 621–628. [Google Scholar] [CrossRef]
- Stengel, A. Nesfatin-1- More than a food intake regulatory peptide. Peptides 2015, 72, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Shami, G.J.; Morsch, M.; Chung, R.S.; Braet, F. Ultrastructural mapping of the zebrafish gastrointestinal system as a basis for experimental drug studies. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Smit, H. Gastric secretion in the lower vertebrates and birds. In Handbook of Physiology Section 6 Alimentary Canal Vol. V Bile, Digestion, Ruminal Physiology; Code, C.F., Ed.; American Physiological Society: Washington, DC, USA, 1968; pp. 2791–2805. [Google Scholar]
- Morley, J.E.; Silver, A.J. Anorexia in the elderly. Neurobiol. Aging 1988, 9, 9–16. [Google Scholar] [CrossRef]
- Ramanjaneya, M.; Tan, B.K.; Rucinski, M.; Kawan, M.; Hu, J.; Kaur, J.; Patel, V.H.; Malendowicz, L.K.; Komarowska, H.; Lehnert, H.; et al. Nesfatin-1 inhibits proliferation and enhances apoptosis of human adrenocortical H295R cells. J. Endocrinol. 2015, 226, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.G.; Bloom, S.R. Gut hormones and the regulation of energy homeostasis. Nature 2006, 444, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Ahima, R.S.; Antwi, D.A. Brain regulation of appetite and satiety. Endocrinol. Metab. Clin. N. Am. 2008, 37, 811–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Štimac, D.; Klobučar Majanović, S.; Franjić, N. Stomach—keyplayer in the regulation of metabolism. Dig. Dis. 2014, 32, 192–201. [Google Scholar] [CrossRef]
- Hussain, S.S.; Bloom, S.R. Theregulation of food intake by the gut-brain axis: Implications for obesity. Int. J. Obes. 2013, 37, 625–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young Brain | Old Brain | |||||
|---|---|---|---|---|---|---|
| mRNA | Protein | mRNA | Protein | |||
| Perikarya | Perikarya | Fibers | Perikarya | Perikarya | Fibers | |
| Telencephalon | ||||||
| Dorsal Telencephalon | ||||||
| Sopracommissural Zone of the Ventral Telencephalon (Vs) | + | + | ||||
| Central Part of the Ventral Telencephalon (Vc) | + | + | ||||
| Preoptic area | ||||||
| Preoptic Nucleus, Parvocellular Part (PPp) | + | + | ||||
| Cortical Nucleus (CN) | + | + | + | |||
| Lateral Preglomerular Nucleus (PGI) | + | + | ||||
| Medial Preglomerular Nucleus (PGm) | + | + | ||||
| Tuberal hypothalamus | ||||||
| Dorsal Hypothalamus (Hd) | + | + | + | + | + | + |
| Ventral Hypothalamus (Hv) | + | + | + | + | + | + |
| Caudal Hypothalamus (Hc) | + | + | + | + | ||
| Periventricular Nucleus of Posterior Tuberculum (TPp) | + | + | + | |||
| Glomerular Nucleus (NG) | + | + | + | + | ||
| Nucleus of Posterior Recess (NRP) | + | + | + | + | ||
| Diffuse Inferior Lobe of Hypothalamus (DIL) | + | + | + | + | + | |
| Posterior tubercle | ||||||
| Paraventricular Organ (PVO) | + | + | + | + | + | |
| Thalamus | ||||||
| Dorsal Posterior Talamic Nucleus (DP) | + | + | ||||
| Ventro-Medial Talamic Nucleus (VM) | + | + | + | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montesano, A.; De Felice, E.; Leggieri, A.; Palladino, A.; Lucini, C.; Scocco, P.; de Girolamo, P.; Baumgart, M.; D’Angelo, L. Ontogenetic Pattern Changes of Nucleobindin-2/Nesfatin-1 in the Brain and Intestinal Bulb of the Short Lived African Turquoise Killifish. J. Clin. Med. 2020, 9, 103. https://doi.org/10.3390/jcm9010103
Montesano A, De Felice E, Leggieri A, Palladino A, Lucini C, Scocco P, de Girolamo P, Baumgart M, D’Angelo L. Ontogenetic Pattern Changes of Nucleobindin-2/Nesfatin-1 in the Brain and Intestinal Bulb of the Short Lived African Turquoise Killifish. Journal of Clinical Medicine. 2020; 9(1):103. https://doi.org/10.3390/jcm9010103
Chicago/Turabian StyleMontesano, Alessia, Elena De Felice, Adele Leggieri, Antonio Palladino, Carla Lucini, Paola Scocco, Paolo de Girolamo, Mario Baumgart, and Livia D’Angelo. 2020. "Ontogenetic Pattern Changes of Nucleobindin-2/Nesfatin-1 in the Brain and Intestinal Bulb of the Short Lived African Turquoise Killifish" Journal of Clinical Medicine 9, no. 1: 103. https://doi.org/10.3390/jcm9010103