Frequent Alteration of Annexin A9 and A10 in HPV-Negative Head and Neck Squamous Cell Carcinomas: Correlation with the Histopathological Differentiation Grade

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Method
2.1. Patients and Tissue Specimens
2.2. Tissue Microarray (TMA) Construction
2.3. Immunohistochemical Study
3. Results
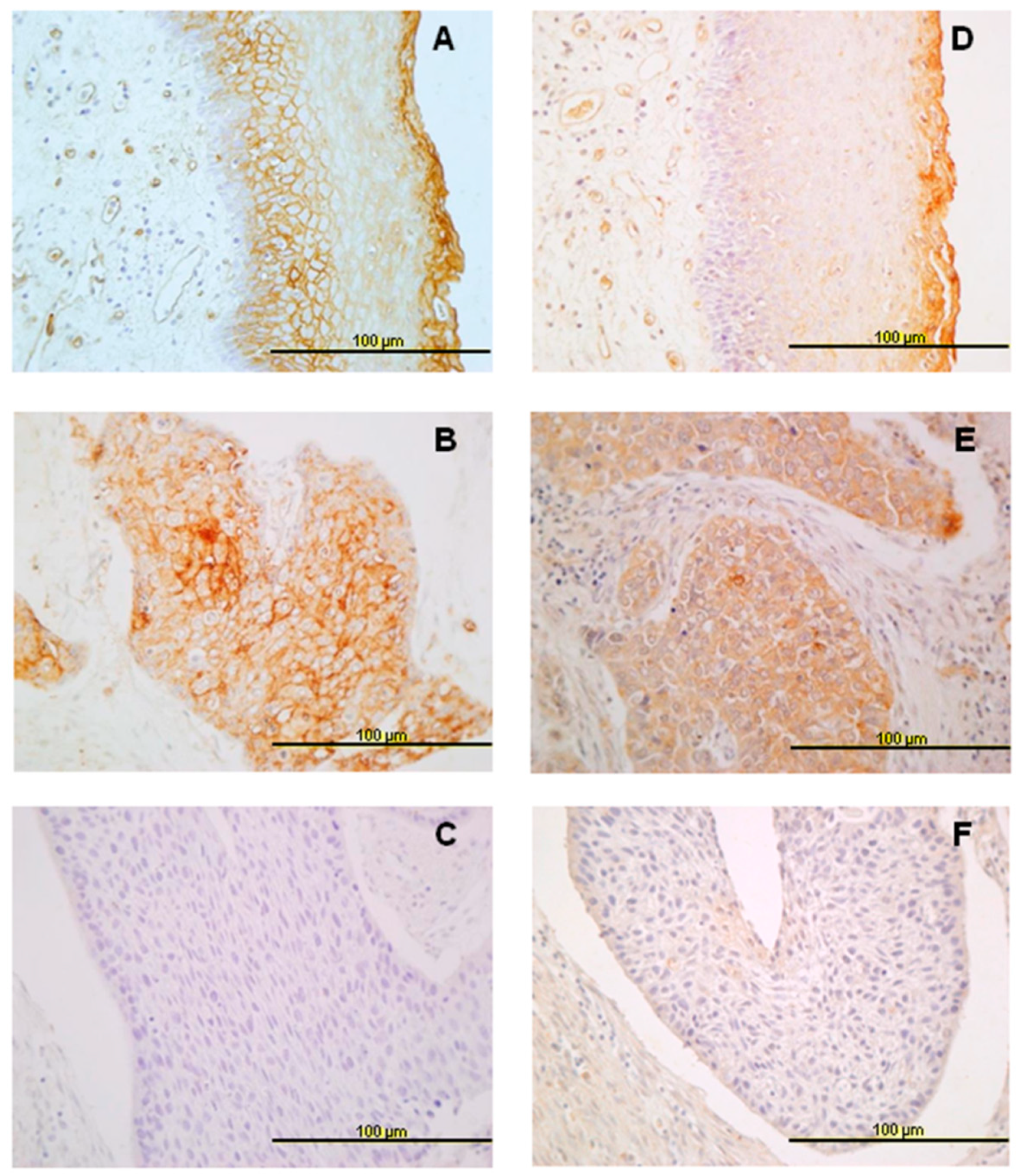
3.1. Expression of ANXA9 and ANX10 in Normal Epithelia
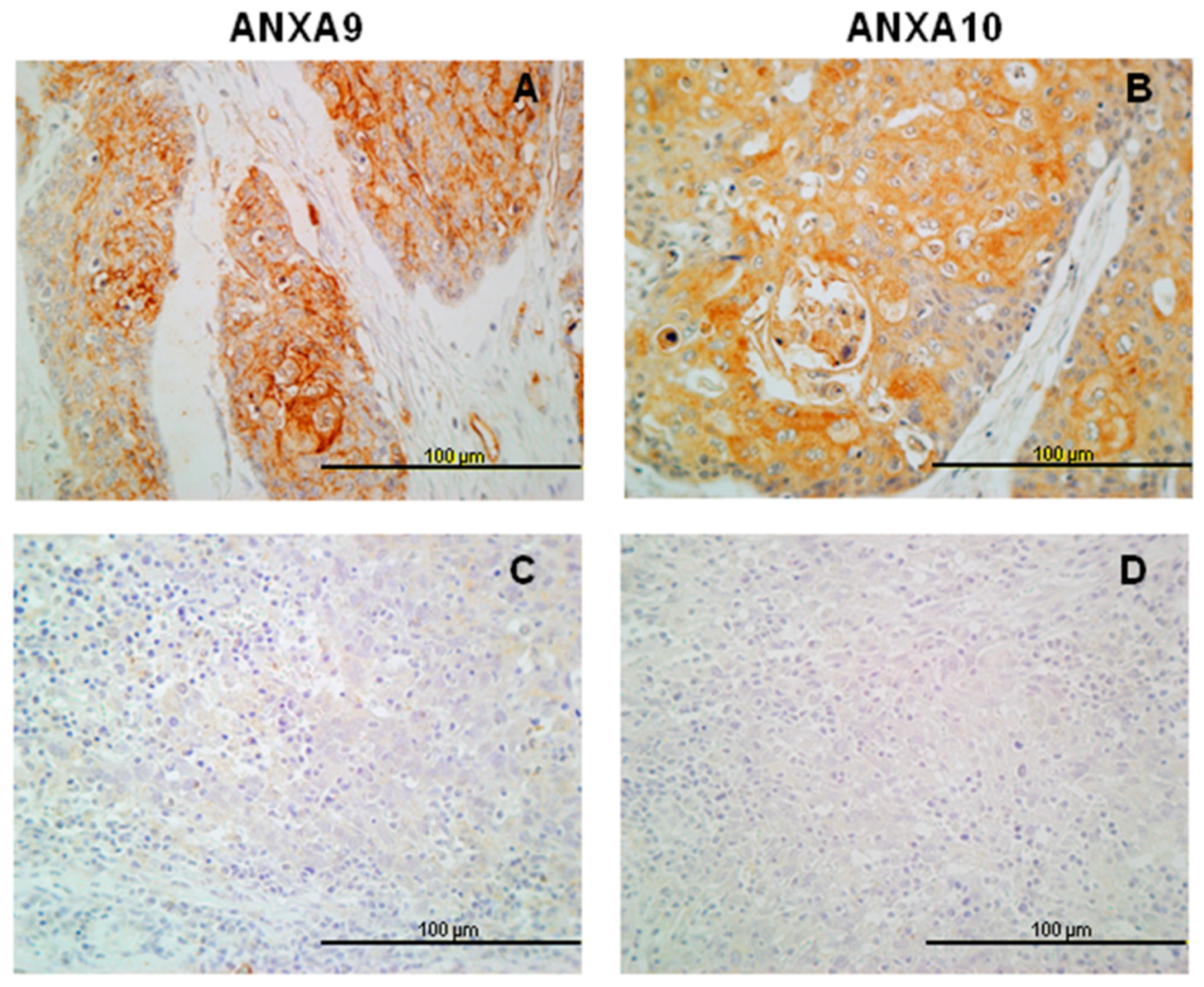
3.2. Expression of ANXA9 in HNSCC Tissue Specimens
3.3. Expression of ANXA10 in HNSCC Specimens
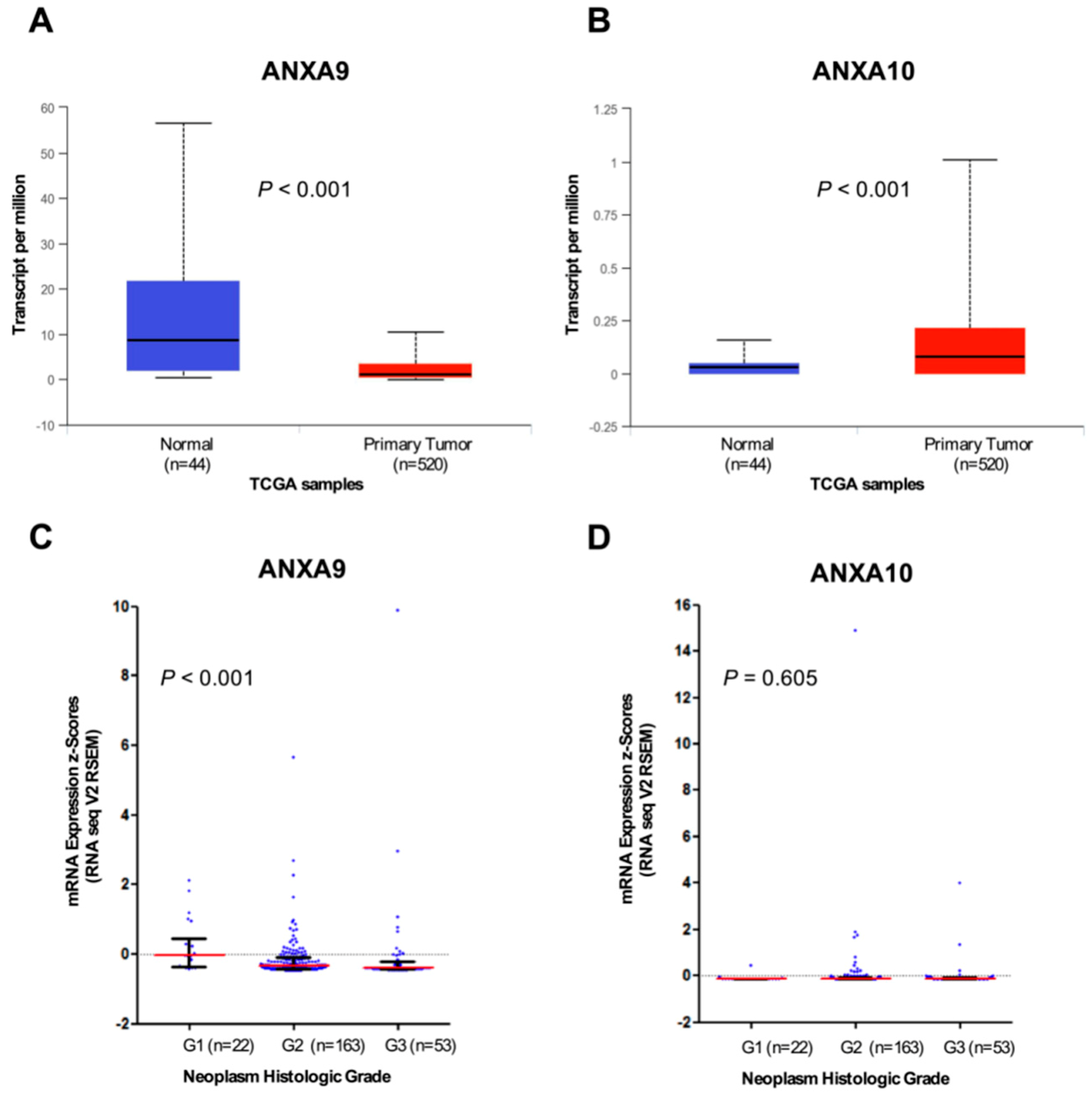
3.4. In Silico Analysis of ANXA9 and ANXA10 mRNA Expression Using The Cancer Genome Atlas (TCGA) HNSCC Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moss, S.E.; Morgan, R.O. The annexins. Genome Biol. 2004, 5, 219. [Google Scholar] [CrossRef] [PubMed]
- Gerke, V.; Moss, S.E. Annexins: From Structure to Function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.J.; Perez, F. Plasma membrane repair: The adaptable cell life-insurance. Curr. Opin. Cell Biol. 2017, 47, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.P.; Garcia, M.; Martin-Almedina, S.; Morgan, R.O. Novel domain architectures and functional determinants in atypical annexins revealed by phylogenomic analysis. Biol. Chem. 2017, 398, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Rescher, U.; Gerke, V. Annexins—Unique membrane binding proteins with diverse functions. J. Cell Sci. 2004, 117, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.B.; Morgan, R.O.; Fernandez, M.P.; Roux, S.J. Evolutionary adaptation of plant annexins has diversified their molecular structures; interactions and functional roles. New Phytol. 2012, 196, 695–712. [Google Scholar] [CrossRef] [PubMed]
- Mirsaeidi, M.; Gidfar, S.; Vu, A.; Schraufnagel, D. Annexins family: Insights into their functions and potential role in pathogenesis of sarcoidosis. J. Transl. Med. 2016, 14, 89. [Google Scholar] [CrossRef]
- Chlystun, M.; Markoff, A.; Gerke, V. Structural and functional characterisation of the mouse annexin A9 promoter. Biochim. Biophys. Acta 2004, 1742, 141–149. [Google Scholar] [CrossRef]
- Rand, J.H. The annexinopathies: A new category of diseases. Biochim. Biophys. Acta 2000, 149, 169–173. [Google Scholar] [CrossRef]
- Mussunoor, S.; Murray, G.I. The role of annexins in tumour development and progression. J. Pathol. 2008, 216, 131–140. [Google Scholar] [CrossRef]
- Hayes, M.J.; Longbottom, R.E.; Evans, M.A.; Moss, S.E. Annexinopathies. Subcell Biochem. 2007, 45, 1–28. [Google Scholar]
- Cosphiadi, I.; Atmakusumah, T.D.; Siregar, N.C.; Muthalib, A.; Harahap, A.; Mansyur, M. Bone metastasis in advanced breast cancer: Analysis of gene expression microarray. Clin. Breast Cancer 2018, 17, 30777. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Bian, H.; Gao, X.; Gui, L. Annexin A9 promotes invasion and metastasis of colorectal cancer and predicts poor prognosis. Int. J. Mol. Med. 2018, 41, 2185–2192. [Google Scholar] [CrossRef] [PubMed]
- Amos, C.I.; Wang, L.E.; Lee, J.E.; Gershenwald, J.E.; Chen, W.V.; Fang, S.; Kosoy, R.; Zhang, M.; Qureshi, A.A.; Vattathil, S.; et al. Genome-wide association study identifies novel loci predisposing to cutaneous melanoma. Hum. Mol. Genet. 2011, 20, 5012–5023. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.T.; Ndoye, A.; Grando, S.A. Pemphigus vulgaris antibody identifies pemphaxin. A novel keratinocyte annexin-like molecule binding acetylcholine. J. Biol. Chem. 2000, 275, 29466–29476. [Google Scholar] [CrossRef] [PubMed]
- Boczonadi, V.; Määttä, A. Annexin A9 is a periplakin interacting partner in membrane-targeted cytoskeletal linker protein complexes. FEBS Lett. 2012, 586, 3090–3096. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kasamatsu, A.; Yamamoto, A.; Koike, K.; Ishige, S.; Takatori, H.; Sakamoto, Y.; Ogawara, K.; Shiiba, M.; Tanzawa, H.; et al. Annexin A10 in Human Oral Cancer: Biomarker for Tumoral Growth via G1/S Transition by Targeting MAPK Signaling Pathways. PLoS ONE 2012, 7, e45510. [Google Scholar] [CrossRef]
- Rodrigo, J.P.; Heideman, D.A.; García-Pedrero, J.M.; Fresno, M.F.; Brakenhoff, R.H.; Díaz Molina, J.P.; Snijders, P.J.; Hermsen, M.A. Time trends in the prevalence of HPV in oropharyngeal squamous cell carcinomas in northern Spain (1990–2009). Int. J. Cancer 2014, 134, 487–492. [Google Scholar] [CrossRef]
- Rodrigo, J.P.; Hermsen, M.A.; Fresno, M.F.; Brakenhoff, R.H.; García-Velasco, F.; Snijders, P.J.; Heideman, D.A.; García-Pedrero, J.M. Prevalence of human papillomavirus in laryngeal and hypopharyngeal squamous cell carcinomas in northern Spain. Cancer Epidemiol. 2015, 39, 37–41. [Google Scholar] [CrossRef]
- Menéndez, S.T.; Rodrigo, J.P.; Alvarez-Teijeiro, S.; Villaronga, M.Á.; Allonca, E.; Vallina, A.; Astudillo, A.; Barros, F.; Suárez, C.; García-Pedrero, J.M. Role of HERG1 potassium channel in both malignant transformation and disease progression in head and neck carcinomas. Mod. Pathol. 2012, 25, 1069–1078. [Google Scholar] [CrossRef]
- Rodrigo, J.P.; Menéndez, S.T.; Hermida-Prado, F.; Álvarez-Teijeiro, S.; Villaronga, M.Á.; Alonso-Durán, L.; Vallina, A.; Martínez-Camblor, P.; Astudillo, A.; Suárez, C.; et al. Clinical significance of Anoctamin-1 gene at 11q13 in the development and progression of head and neck squamous cell carcinomas. Sci. Rep. 2015, 5, 15698. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Network. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Rodriguez, I.P.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Fatimathas, L.; Moss, S.E. Annexins as disease modifiers. Histol. Histopathol. 2010, 25, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Goebeler, V.; Ruhe, D.; Gerke, V.; Rescher, U. Atypical properties displayed by annexin A9; a novel member of the annexin family of Ca2+ and lipid binding proteins. FEBS Lett. 2003, 546, 359–364. [Google Scholar] [CrossRef]
- Morgan, R.O.; Martin-Almedina, S.; Iglesias, J.M.; Gonzalez-Florez, M.I.; Fernandez, M.P. Evolutionary perspective on annexin calcium-binding domains. Biochim. Biophys. Acta 2004, 1742, 133–140. [Google Scholar] [CrossRef]
- Garcia-Pedrero, J.M.; Fernandez, M.P.; Morgan, R.O.; Herrero Zapatero, A.; Gonzalez, M.V.; Suarez Nieto, C.; Rodrigo, J.P. Annexin A1 down-regulation in head and neck cancer is associated with epithelial differentiation status. Am. J. Pathol. 2004, 164, 73–79. [Google Scholar] [CrossRef]
- Álvarez-Teijeiro, S.; Menéndez, S.T.; Villaronga, M.A.; Pena-Alonso, E.; Rodrigo, J.P.; Morgan, R.O.; Granda-Díaz, R.; Salom, C.; Fernandez, M.P.; García-Pedrero, J.M. Annexin A1 down-regulation in head and neck squamous cell carcinoma is mediated via transcriptional control with direct involvement of miR-196a/b. Sci. Rep. 2017, 7, 67–90. [Google Scholar] [CrossRef]
- Pena-Alonso, E.; Rodrigo, J.P.; Parra, I.C.; Pedrero, J.M.; Meana, M.V.; Nieto, C.S.; Fresno, M.F.; Morgan, R.O.; Fernandez, M.P. Annexin A2 localizes to the basal epithelial layer and is down-regulated in dysplasia and head and neck squamous cell carcinoma. Cancer Lett. 2008, 263, 89–98. [Google Scholar] [CrossRef]
- Smid, M.; Wang, Y.; Klijn, J.G.; Sieuwerts, A.M.; Zhang, Y.; Atkins, D.; Martens, J.W.; Foekens, J.A. Genes associated with breast cancer metastatic to bone. J. Clin. Oncol. 2006, 24, 2261–2267. [Google Scholar] [CrossRef]
- Miyoshi, N.; Yamamoto, H.; Mimori, K.; Yamashita, S.; Miyazaki, S.; Nakagawa, S.; Ishii, H.; Noura, S.; Ohue, M.; Yano, M.; et al. Anxa9 gene expression in colorectal cancer: A novel marker for prognosis. Oncol. Lett. 2014, 8, 2313–2317. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Rhee, Y.Y.; Kim, K.J.; Cho, N.Y.; Lee, H.S.; Kang, G.H. Annexin A10 expression correlates with serrated pathway features in colorectal carcinoma with microsatellite instability. Apmis 2014, 122, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Lin, C.Y.; Peng, S.Y.; Jeng, Y.M.; Pan, H.W.; Lai, P.L.; Liu, C.L.; Hsu, H.C. Down-regulation of annexin A10 in hepatocellular carcinoma is associated with vascular invasion; early recurrence; and poor prognosis in synergy with p53 mutation. Am. J. Pathol. 2002, 160, 1831–1837. [Google Scholar] [CrossRef]
- Peng, S.Y.; Ou, Y.H.; Chen, W.J.; Li, H.Y.; Liu, S.H.; Pan, H.W.; Lai, P.L.; Jeng, Y.M.; Chen, D.C.; Hsu, H.C. Aberrant expressions of annexin A10 short isoform; osteopontin and alpha-fetoprotein at chromosome 4q cooperatively contribute to progression and poor prognosis of hepatocellular carcinoma. Int. J. Oncol. 2005, 26, 1053–1061. [Google Scholar] [PubMed]
- Kim, J.K.; Kim, P.J.; Jung, K.H.; Noh, J.H.; Eun, J.W.; Bae, H.J.; Xie, H.J.; Shan, J.M.; Ping, W.Y.; Park, W.S.; et al. Decreased expression of annexin A10 in gastric cancer and its overexpression in tumor cell growth suppression. Oncol. Rep. 2010, 24, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Munksgaard, P.P.; Mansilla, F.; Brems Eskildsen, A.S.; Fristrup, N.; Birkenkamp-Demtröder, K.; Ulhøi, B.P.; Borre, M.; Agerbæk, M.; Hermann, G.G.; Orntoft, T.F.; et al. Low ANXA10 expression is associated with disease aggressiveness in bladder cancer. Br. J. Cancer 2011, 105, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Sajanti, S.A.; Väyrynen, J.P.; Sirniö, P.; Klintrup, K.; Mäkelä, J.; Tuomisto, A.; Mäkinen, M.J. Annexin A10 is a marker for the serrated pathway of colorectal carcinoma. Virchows Arch. 2015, 466, 5–12. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | No. Cases (%) |
|---|---|
| Age, mean (range) | 58.6 (30–86 years) |
| Location | |
| Oropharynx | 241 (65) |
| Hypopharynx | 64 (17) |
| Larynx | 67 (18) |
| pathologic T classification | |
| T1 | 38 (10) |
| T2 | 77 (21) |
| T3 | 125 (34) |
| T4 | 132 (35) |
| pathologic N classification | |
| N0 | 103 (28) |
| N1 | 46 (12) |
| N2 | 183 (49) |
| N3 | 40 (11) |
| Stage | |
| I | 20 (5) |
| II | 24 (6) |
| III | 64 (17) |
| IV | 264 (71) |
| Degree of differentiation | |
| Well-differentiated | 147 (39) |
| Moderately-differentiated | 148 (40) |
| Poorly-differentiated | 77 (21) |
| Total | 372 |
| Characteristic | No. Cases for ANXA9 | Positive ANXA9 Expression (%) | p | No. Cases for ANXA10 | Positive ANXA10 Expression (%) | p |
|---|---|---|---|---|---|---|
| Location | ||||||
| Oropharynx | 234 | 166 (71) | 231 | 160 (69) | ||
| Hypopharynx | 58 | 17 (29) | 0.000 # | 55 | 28 (51) | 0.019 # |
| Larynx | 54 | 17 (31) | 54 | 31 (57) | ||
| pT Classification | ||||||
| T1-T2 | 100 | 52 (52) | 95 | 58 (61) | ||
| T3 | 120 | 73 (61) | 0.377 # | 119 | 75 (63) | 0.591 # |
| T4 | 126 | 73 (59) | 123 | 83 (67) | ||
| pN Classification | ||||||
| N0 | 87 | 48 (55) | 0.616 † | 87 | 53 (61) | 0.439 † |
| N1-3 | 259 | 152 (59) | 253 | 166 (66) | ||
| Stage | ||||||
| I-II | 33 | 14 (42) | 32 | 19 (59) | ||
| III | 61 | 39 (64) | 0.124 # | 60 | 39 (65) | 0.822 # |
| IV | 252 | 147 (58) | 248 | 161 (65) | ||
| Degree of differentiation | ||||||
| Well-differentiated | 136 | 98 (72) | 134 | 103 (77) | ||
| Moderately-differentiated | 137 | 73 (53) | 0.000 # | 137 | 85 (62) | 0.000 # |
| Poorly-differentiated | 73 | 29 (40) | 69 | 31 (45) | ||
| Recurrence | ||||||
| No | 132 | 77 (58) | 0.91 † | 132 | 80 (61) | 0.248 † |
| Yes | 214 | 123 (57) | 208 | 139 (67) | ||
| Total | 346 | 200 (58) | 340 | 219 (64) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salom, C.; Álvarez-Teijeiro, S.; Fernández, M.P.; Morgan, R.O.; Allonca, E.; Vallina, A.; Lorz, C.; de Villalaín, L.; Fernández-García, M.S.; Rodrigo, J.P.; et al. Frequent Alteration of Annexin A9 and A10 in HPV-Negative Head and Neck Squamous Cell Carcinomas: Correlation with the Histopathological Differentiation Grade. J. Clin. Med. 2019, 8, 229. https://doi.org/10.3390/jcm8020229
Salom C, Álvarez-Teijeiro S, Fernández MP, Morgan RO, Allonca E, Vallina A, Lorz C, de Villalaín L, Fernández-García MS, Rodrigo JP, et al. Frequent Alteration of Annexin A9 and A10 in HPV-Negative Head and Neck Squamous Cell Carcinomas: Correlation with the Histopathological Differentiation Grade. Journal of Clinical Medicine. 2019; 8(2):229. https://doi.org/10.3390/jcm8020229
Chicago/Turabian StyleSalom, Cecilia, Saúl Álvarez-Teijeiro, M. Pilar Fernández, Reginald O. Morgan, Eva Allonca, Aitana Vallina, Corina Lorz, Lucas de Villalaín, M. Soledad Fernández-García, Juan P. Rodrigo, and et al. 2019. "Frequent Alteration of Annexin A9 and A10 in HPV-Negative Head and Neck Squamous Cell Carcinomas: Correlation with the Histopathological Differentiation Grade" Journal of Clinical Medicine 8, no. 2: 229. https://doi.org/10.3390/jcm8020229