
A Comparison of Deformed Wing Virus in Deformed and Asymptomatic Honey Bees


 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Honeybee Samples
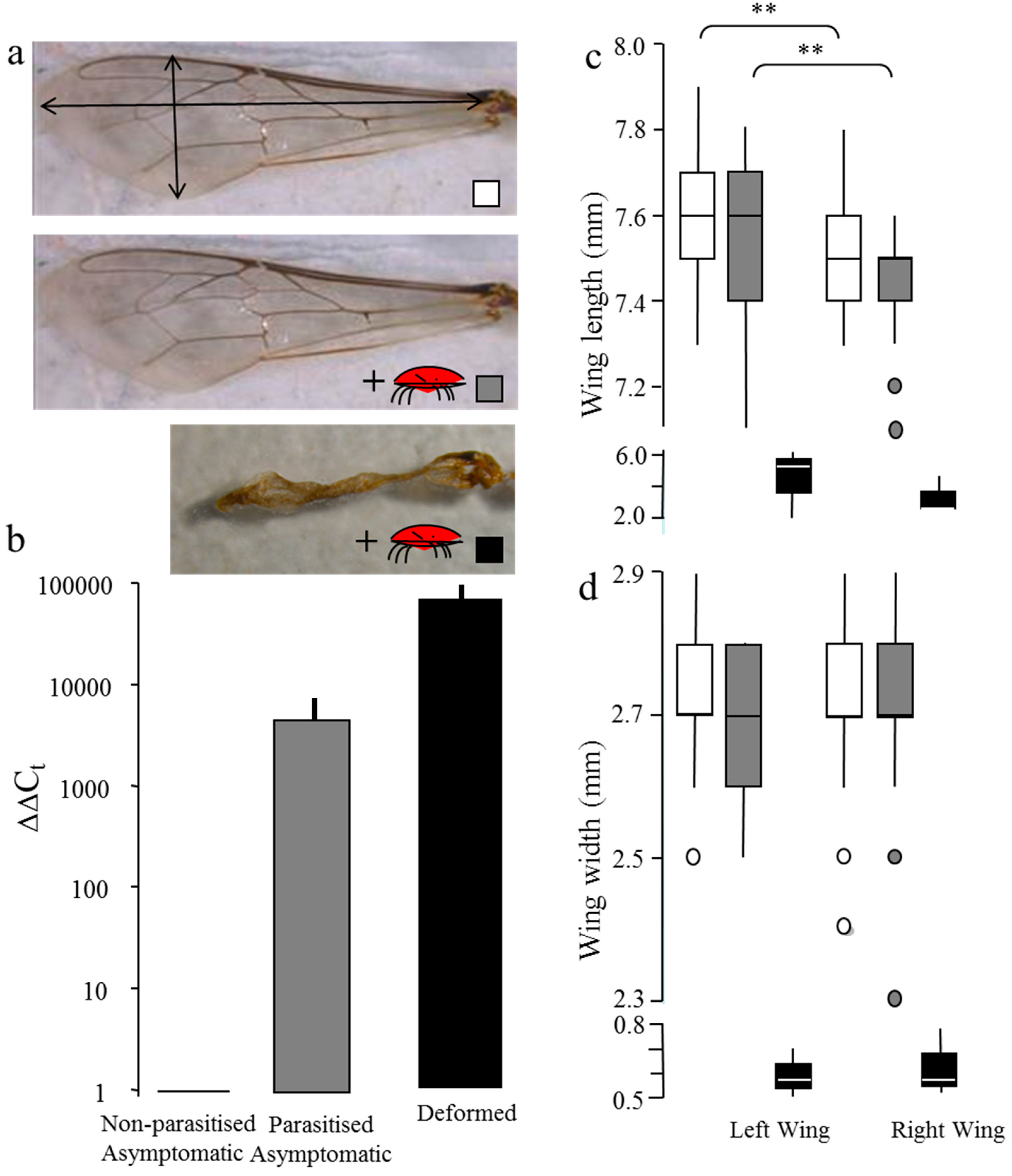
2.2. Effect of Viral Load on Wing Deformity
2.3. Next Generation Sequencing, Assembly and Data Normalisation
3. Results
3.1. Viral Quantification and Wing Deformity in Honey Bees
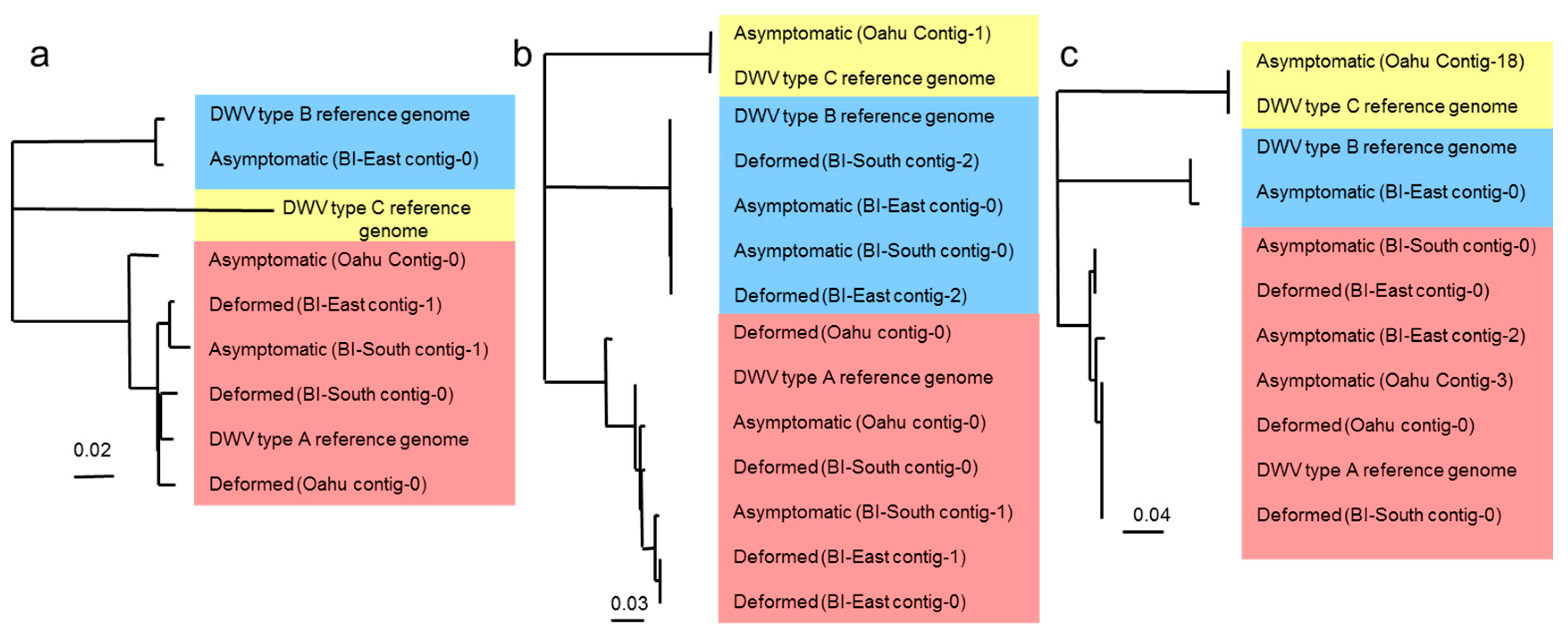
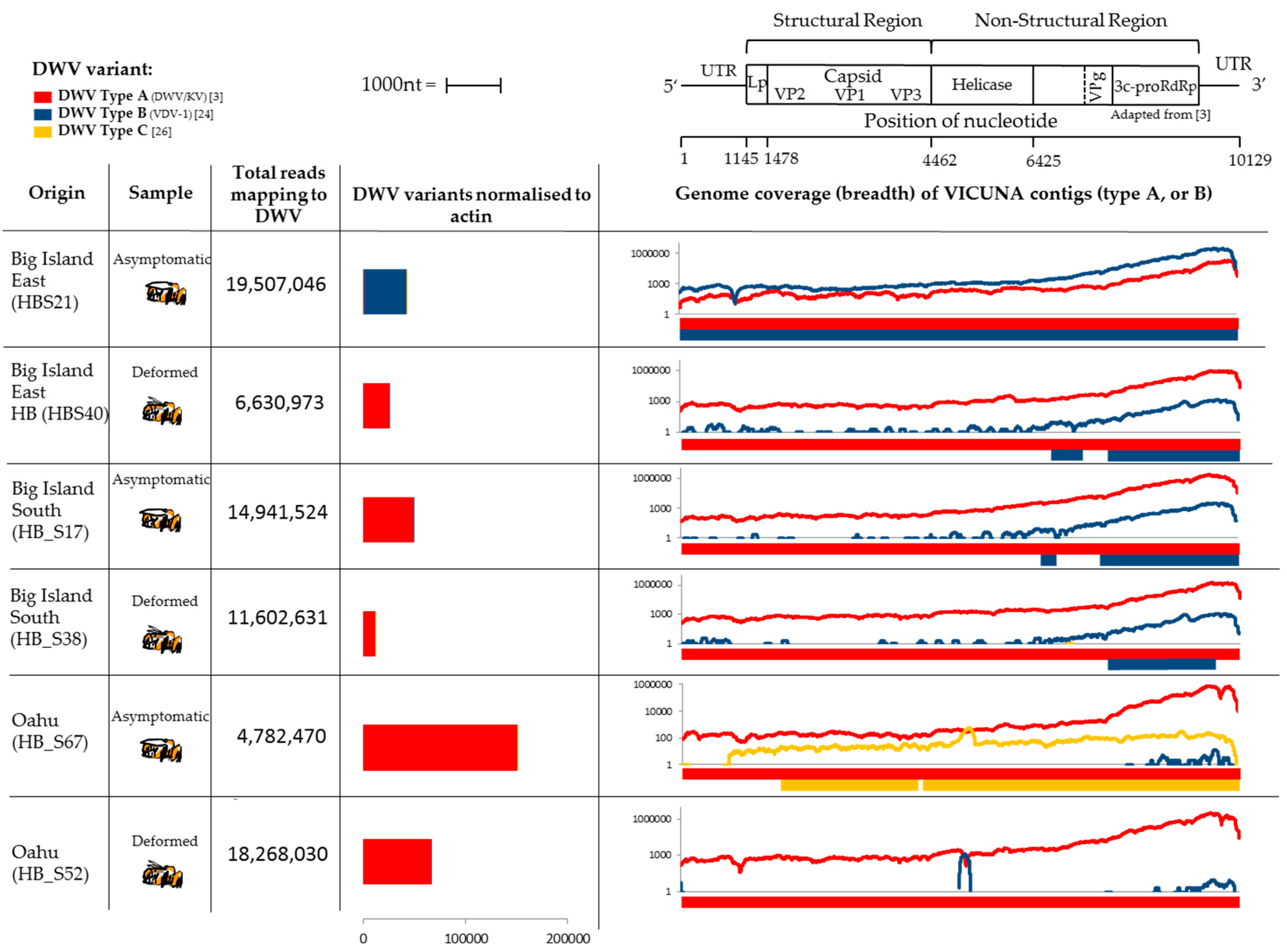
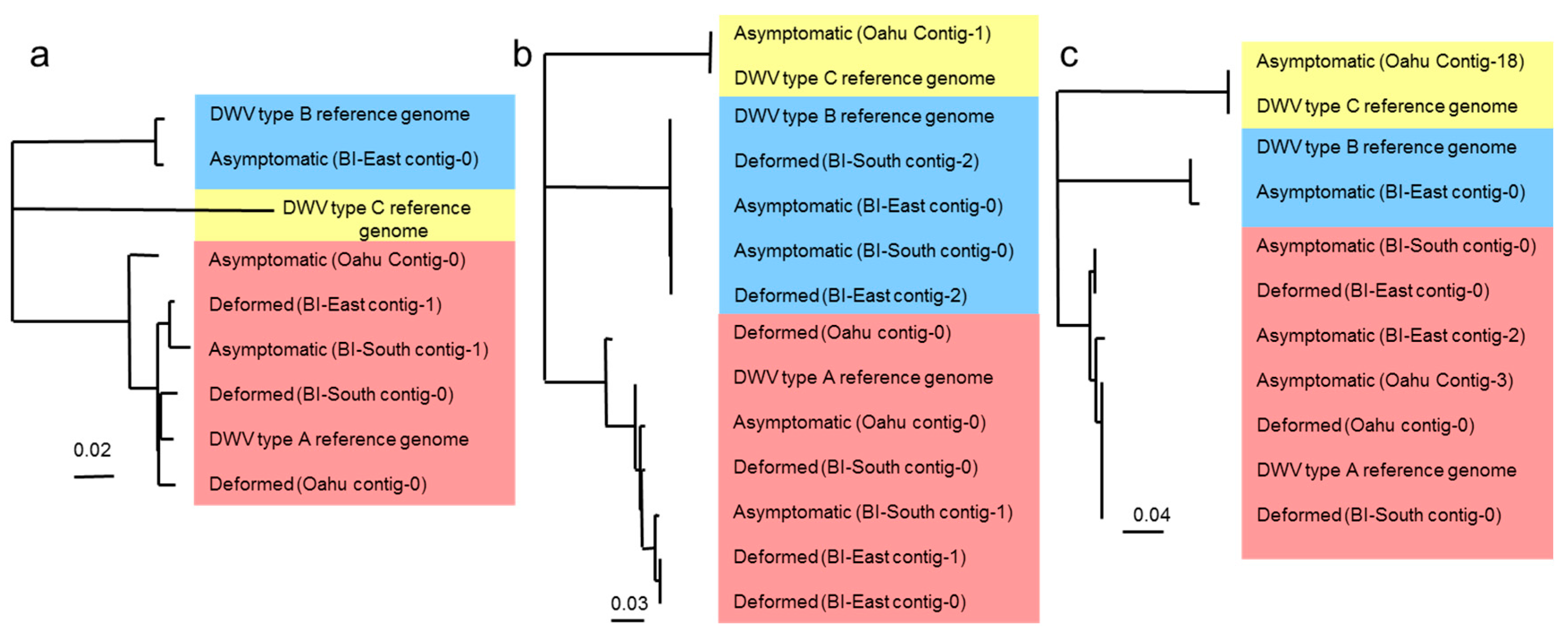
3.2. Next Generation Illumina Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tantillo, G.; Bottaro, M.; Di Pinto, A.; Martella, V.; Di Pinto, P.; Terio, V. Virus Infections of Honeybees Apis mellifera. Ital. J. Food Saf. 2015. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D. Global Honey Bee Viral Landscape Altered by a Parasitic Mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Lanzi, G.; de Miranda, J.R.; Boniotti, M.B.; Cameron, C.E.; Lavazza, A.; Capucci, L.; Camazine, S.M.; Rossi, C. Molecular and biological characterization of deformed wing virus of Honeybees (Apis mellifera L.). J. Virol. 2006, 80, 4998–5009. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cox-Foster, D. Effects of parasitization by Varroa destructor on survivorship and physiological traits of Apis mellifera in correlation with viral incidence and microbial challenge. J. Parasitol. 2007, 134, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Nordström, S.; Fries, I.; Aarhus, A.; Hansen, H.; Korpela, S. Virus infections in Nordic honey bee colonies with no, low or severe Varroa jacobsoni infestations. Apidologie 1999, 30, 475–484. [Google Scholar] [CrossRef]
- DeJong, D.; DeJong, P.H.; Goncalves, L.S. Weight loss and other damage to developing worker honeybees from infestation with Varroa jacobsoni. J. Apic. Res. 1982, 21, 165–167. [Google Scholar] [CrossRef]
- Bowen-Walker, P.L.; Martin, S.J.; Gunn, A. The transmission of deformed wing virus between Honeybees (Apis mellifera L.) by the ectoparasitic mite Varroa jacobsoni Oud. J. Invertebr. Pathol. 1999, 73, 101–106. [Google Scholar] [CrossRef] [PubMed]
- De Jong, D.; Soares, A. An isolated population of Italian bees that has survived Varroa jacobsoni infestation without treatment for over 12 years. Am. Bee J. 1997, 137, 742–745. [Google Scholar]
- De Mattos, I.M.; de Jong, D.; Soares, A.E.E. Island population of European honey bees in Northeastern Brazil that have survived Varroa infestations for over 30 years. Apidologie 2016, 47, 818–827. [Google Scholar] [CrossRef]
- Brettell, L.E.; Martin, S.J. Oldest Varroa tolerant honey bee population provides insight into the origins of the global decline of honey bees. Sci. Rep. 2017. (under review). [Google Scholar]
- Highfield, A.C.; El Nagar, A.; Mackinder, L.C.M.; Noel, L.M.-L.J.; Hall, M.J.; Martin, S.J.; Schroeder, D.C. Deformed wing virus implicated in overwintering honeybee colony losses. Appl. Environ. Microbiol. 2009, 75, 7212–7220. [Google Scholar] [CrossRef] [PubMed]
- Tentcheva, D.; Gauthier, L.; Jouve, S.; Canabady-Rochelle, L.; Dainat, B.; Cousserans, F.; Colin, M.E.; Ball, B.V.; Bergoin, M. Polymerase Chain Reaction detection of deformed wing virus (DWV) in Apis mellifera and Varroa destructor. Apidologie 2004, 35, 431–439. [Google Scholar] [CrossRef]
- Chen, Y.P.; Higgins, J.A.; Feldlaufer, M.F. Quantitative real-time reverse transcription-PCR analysis of deformed wing virus infection in the honeybee (Apis mellifera L.). Appl. Environ. Microbiol. 2005, 71, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Forsgren, E.; Ingemar, F.; de Miranda, J.R. Adult honey bees (Apis mellifera) with deformed wings discovered in confirmed varroa-free colonies. J. Apic. Res. 2012, 51, 136–138. [Google Scholar] [CrossRef]
- Gisder, S.; Aumeier, P.; Genersch, E. Deformed wing virus: replication and viral load in mites (Varroa destructor). J. Gen. Virol. 2009, 90, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Genersch, E. RT-PCR analysis of deformed wing virus in honeybees (Apis mellifera) and mites (Varroa destructor). J. Gen. Virol. 2005, 86, 3419–3424. [Google Scholar] [CrossRef] [PubMed]
- Fujiyuki, T.; Takeuchi, H.; Ono, M.; Ohka, S.; Sasaki, T.; Nomoto, A.; Kubo, T. Novel insect picornalike virus identified in the brains of aggressive worker honey bees. J. Virol. 2004, 78, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Mueller, U. Virus infection causes specific learning deficits in honeybee foragers. Proc. R. Soc. B 2007, 274, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [PubMed]
- Glinski, Z.; Jarosz, J. Varroa jacobsoni as a carrier of bacterial infections to a recipient bee host. Apidologie 1992, 23, 25–31. [Google Scholar] [CrossRef]
- Koch, W.; Ritter, W. Experimental examinations concerning the problem of deformed emerging bees after infestation with Varroa jacobsoni. J. Vet. Med. B 1991, 38, 337–344. [Google Scholar] [CrossRef]
- Alippi, A.M.; Albo, G.N.; Marcangeli, J.; Leniz, D.; Noriega, A. The mite Varroa jacobsoni does not transmit American foulbrood from infected to healthy colonies. Exp. Appl. Acarol. 1995, 19, 607–613. [Google Scholar] [CrossRef]
- De Miranda, J.R.; Genersch, E. Deformed wing virus. J. Invertebr. Pathol. 2010, 103, S48–S61. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Holland, J.J. RNA virus mutations and fitness for survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef] [PubMed]
- Ongus, J.R.; Peters, D.; Bonmatin, J.M.; Bengsch, E.; Vlak, J.M.; van Oers, M.M. Complete sequence of a picorna-like virus of the genus Iflavirus replicating in the mite Varroa destructor. J. Gen. Virol. 2004, 85, 3747–3755. [Google Scholar] [CrossRef] [PubMed]
- Zioni, N.; Soroker, V.; Chejanovsky, N. Replication of Varroa destructor virus 1 (VDV-1) and a Varroa destructor virus 1–deformed wing virus recombinant (VDV-1–DWV) in the head of the honey bee. Virology 2011, 417, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Mordecai, G.J.; Wilfert, L.; Martin, S.J.; Jones, I.M.; Schroeder, D.C. Diversity in a honey bee pathogen: First report of a third master variant of the Deformed Wing Virus quasispecies. ISME J. 2016, 10, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Mcmahon, D.P.; Natsopoulou, M.E.; Doublet, V.; Fürst, M.; Weging, S.; Brown, M.J.; Gogol-Döring, A.; Paxton, R.J. Elevated virulence of an emerging viral genotype as a driver of honeybee loss. Proc. R. Soc. B 2016. [Google Scholar] [CrossRef] [PubMed]
- Wilfert, L.; Long, G.; Leggett, H.C.; Schmid-Hempel, P.; Butlin, R.; Martin, S.J.; Boots, M. Deformed wing virus is a recent global epidemic in honeybees driven by Varroa mites. Science 2016, 351, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Mordecai, G.J.; Brettell, L.E.; Martin, S.J.; Dixon, D.; Jones, I.M.; Schroeder, D.C. Superinfection exclusion and the long-term survival of honey bees in Varroa-infested colonies. ISME J. 2016, 10, 1182–1191. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Wood, G.R.; Fannon, J.M.; Moore, J.D.; Bull, J.C.; Chandler, D.; Mead, A.; Burroughs, N.; Evans, D.J. A Virulent strain of deformed wing virus (DWV) of Honeybees (Apis mellifera) prevails after Varroa destructor-Mediated, or in vitro, transmission. PLoS Pathog. 2014, 10, e1004230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingo, E.; Sheldon, J.; Perales, C. Viral quasispecies evolution. Microbiol. Mol. Biol. Rev. 2012, 76, 159–216. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Carreck, N.L.; Ball, B.V. The role of Deformed wing virus in the collapse of honeybee colonies. J. Apic. Res. 2013, 52, 251–258. [Google Scholar] [CrossRef]
- Marcangeli, J.; Monetti, L.; Fernandez, N. Malformations produced by Varroa jacobsoni on Apis mellifera in the province of Buenos Aires, Argentina. Apidologie 1992, 23, 399–402. [Google Scholar] [CrossRef]
- Schneider, S.S.; Leamy, L.J.; Lewis, L.A.; Hoffman, G.D. The influence of hybridization between African and European Honeybees, Apis mellifera, on asymmetry in wing size and shape. Evolution 2003, 57, 2350–2364. [Google Scholar] [CrossRef] [PubMed]
- Mazeed, A.M.M. Anomalies and asymmetry of wing venation pattern in Carniolan and Egyptian bee populations in Egypt. Egypt Acad. J. Biolog. Sci. 2011, 4, 149–161. [Google Scholar]
- Brooks, E.M.; Sheflin, L.G.; Spaulding, S.W. Secondary structure in the 3’ UTR of EGF and the choice of reverse transcriptases affect the detection of message diversity by RT-PCR. Biotechniques 1995, 19, 806–812. [Google Scholar] [PubMed]
- Smith, D.R.; Crespi, B.J.; Bookstein, F.L. Fluctuating asymmetry in the honey bee, Apis mellifera: effects of ploidy and hybridization. J. Evol. Biol. 1997, 10, 551–574. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; McIntyre, G.S.; Zaklan, S.D. Leftright asymmetry of fly wings and the evolution of body axes. Proc. R. Soc. Lond. B 1998, 265, 1255–1259. [Google Scholar] [CrossRef] [PubMed]
- Bowen-Walker, P.L.; Gunn, A. Inter-host transfer and survival of Varroa jacobsoni under simulated and natural winter conditions. J. Apic. Res. 1998, 37, 199–204. [Google Scholar] [CrossRef]
- Yue, C.; Schröder, M.; Gisder, S.; Genersch, E. Vertical-transmission routes for deformed wing virus of honeybees (Apis mellifera). J. Gen. Virol. 2007, 88, 2329–2336. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, E.W.; Chen, Y.; Message, D.; Pettis, J.; Evans, J.D. Virus infections in Brazilian honey bees. J. Invertebr. Pathol. 2008, 99, 117–119. [Google Scholar] [CrossRef]
- Medina, L.M.; Martin, S.J.; Espinosa-Montaño, L.; Ratnieks, F.L. Reproduction of Varroa destructor in worker brood of Africanized honey bees (Apis mellifera). Exp. Appl. Acarol. 2002, 27, 79–88. [Google Scholar] [CrossRef]
- Erban, T.; Harant, K.; Hubalek, M.; Vitamvas, P.; Kamler, M.; Poltronieri, P.; Tyl, J.; Markovic, M.; Titera, D. In-depth proteomic analysis of Varroa destructor: Detection of DWV-complex, ABPV, VdMLV and honeybee proteins in the mite. Sci. Rep. 2015. [Google Scholar] [CrossRef] [PubMed]
- Berényi, O.; Bakonyi, T.; Derakhshifar, I.; Köglberger, H.; Topolska, G.; Ritter, W.; Pechhacker, H.; Nowotny, N. Phylogenetic analysis of deformed wing virus genotypes from diverse geographic origins indicates recent global distribution of the virus. Appl. Environ. Microbiol. 2007, 73, 3605–3611. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brettell, L.E.; Mordecai, G.J.; Schroeder, D.C.; Jones, I.M.; Da Silva, J.R.; Vicente-Rubiano, M.; Martin, S.J. A Comparison of Deformed Wing Virus in Deformed and Asymptomatic Honey Bees. Insects 2017, 8, 28. https://doi.org/10.3390/insects8010028
Brettell LE, Mordecai GJ, Schroeder DC, Jones IM, Da Silva JR, Vicente-Rubiano M, Martin SJ. A Comparison of Deformed Wing Virus in Deformed and Asymptomatic Honey Bees. Insects. 2017; 8(1):28. https://doi.org/10.3390/insects8010028
Chicago/Turabian StyleBrettell, Laura E., Gideon J. Mordecai, Declan C. Schroeder, Ian M. Jones, Jessica R. Da Silva, Marina Vicente-Rubiano, and Stephen J. Martin. 2017. "A Comparison of Deformed Wing Virus in Deformed and Asymptomatic Honey Bees" Insects 8, no. 1: 28. https://doi.org/10.3390/insects8010028