Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates


1
Molecular Biology Section, Department of Cellular and Molecular Biology, Primate Research Institute, Kyoto University, Aichi 4848506, Japan
2
Department of Nutritional Science and Food Safety, Faculty of Applied Bioscience, Tokyo University of Agriculture, Tokyo 1568502, Japan
3
Structural Bioscience for Taste Molecular Recognition, Graduate School of Medicine, Kyoto University, Kyoto 6068507, Japan
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(3), 902; https://doi.org/10.3390/ijms21030902
Submission received: 20 December 2019
/
Revised: 27 January 2020
/
Accepted: 28 January 2020
/
Published: 30 January 2020
(This article belongs to the Section Molecular Biology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:(1) Background: Recent studies have investigated the expression of taste-related genes in the organs of various animals, including humans; however, data for additional taxa are needed to facilitate comparative analyses within and among species. (2) Methods: We investigated the expression of taste-related genes in the intestines of rhesus macaques, the non-human primates most commonly used in experimental models. (3) Results: Based on RNAseq and qRT-PCR, genes encoding bitter taste receptors and the G-protein gustducin were expressed in the gut of rhesus macaques. RNAscope analysis showed that one of the bitter receptors, TAS2R38, was expressed in some cells in the small intestine, and immunohistochemical analysis revealed the presence of T2R38-positive cells in the villi of the intestines. (4) Conclusions: These results suggest that bitter receptors are expressed in the gut of rhesus macaques, supporting the use of macaques as a model for studies of human taste, including gut analyses.
1. Introduction
Taste perception can provide important information about foods. In mammals, bitter taste in the oral cavity is detected by T2Rs, which are heterotrimeric G-protein-coupled receptors expressed on the surface of the tongue [1]. When bitter compounds bind to receptors, information is transmitted to the brain, enabling the recognition of bitter taste.
Non-human primates such as macaques and marmosets, which belong to the Old World and New World monkey families, respectively, are frequently used as models for humans in various types of research. Previous studies have shown that taste-related proteins (e.g., gustducin, TRPM5, and some T2Rs) are selectively expressed in the cecum of common marmosets [2,3]. These observations suggest that T2R expression patterns vary among species, even among primates. In addition, the functional properties of T2Rs are different even among orthologous species. For example, T2R38 in humans [4] and macaques [5] is responsive to phenylthiocarbamide (PTC) but the mouse orthologue T2R138 is not [6]. In humans, a polymorphism in the TAS2R38 gives rise to the difference in response to PTC between the PAV (taster) and AVI (non-taster) genotypes (phenotypes) [4]. Also, in Japanese macaques, there is a polymorphism in the TAS2R38 gene in which a mutation in the nucleotide position 2 of the starting methionine results in the non-taster phenotype [5].
A previous study using mouse and human organs suggested that T2R expression patterns vary among cell types. For example, Tas2R131 is expressed in Paneth cells and goblet cells in the mouse small and large intestines, respectively [7], T2R138 is expressed in the enteroendocrine cells of the mouse colon [8], and T2R38 is expressed in the enteroendocrine cells of the human colon [9]. To understand the general mechanism by which T2R expression is regulated in humans and rodents, it is necessary to use an experimental model that is intermediate between these species. In this study, we measured TAS2R mRNA levels and corresponding protein expression levels in rhesus macaques. This non-human primate could be a suitable model for studying human taste perception by focusing in particular on the TAS2R38 gene and T2R38 protein for which appropriate antibodies are readily available.
2. Results
2.1. RNAseq and RT-PCR
We first used RNAseq to screen for TAS2R mRNAs in various organs of rhesus macaques, which have orthologues to human TAS2Rs (Table S1 and Figure 1).
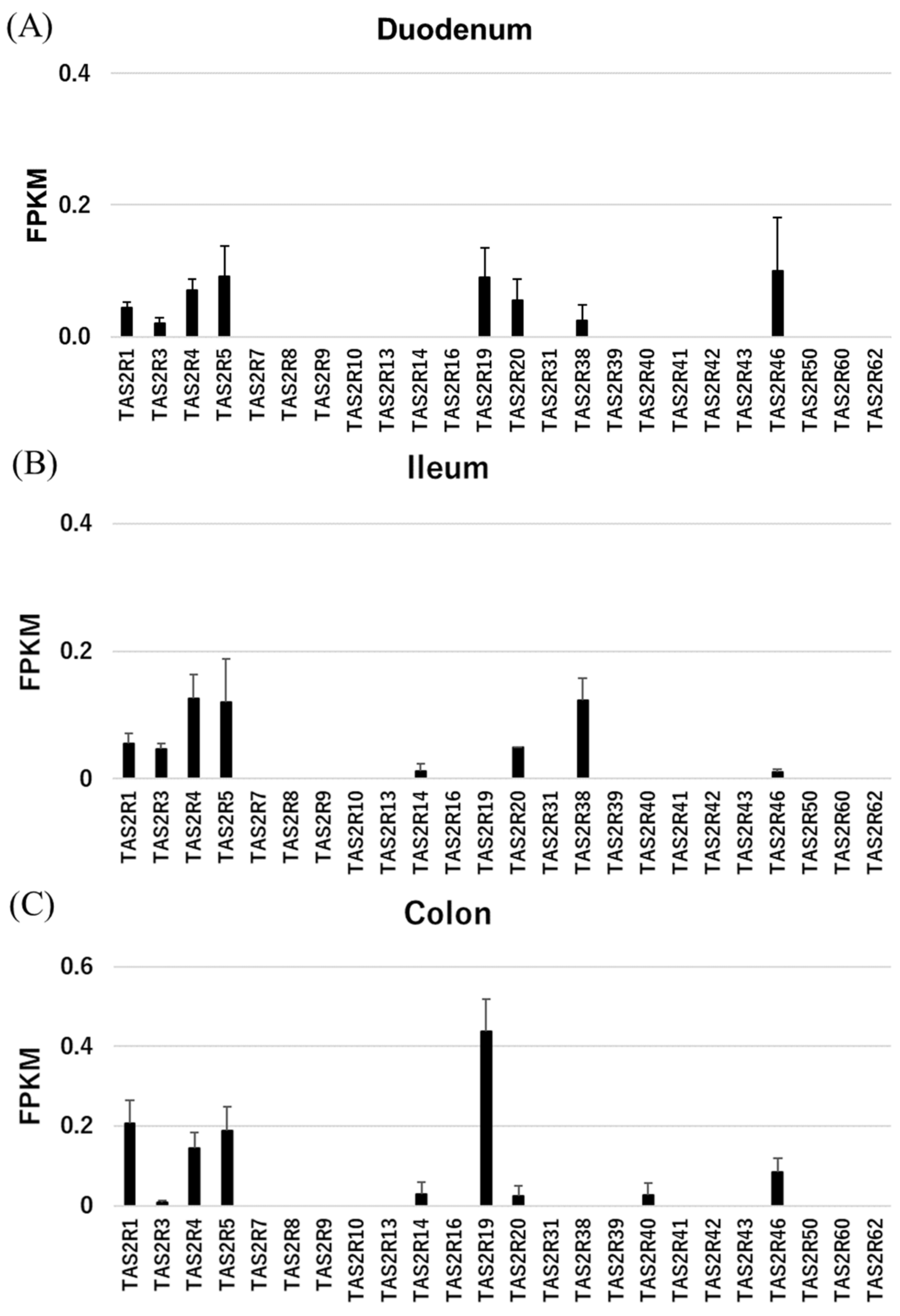
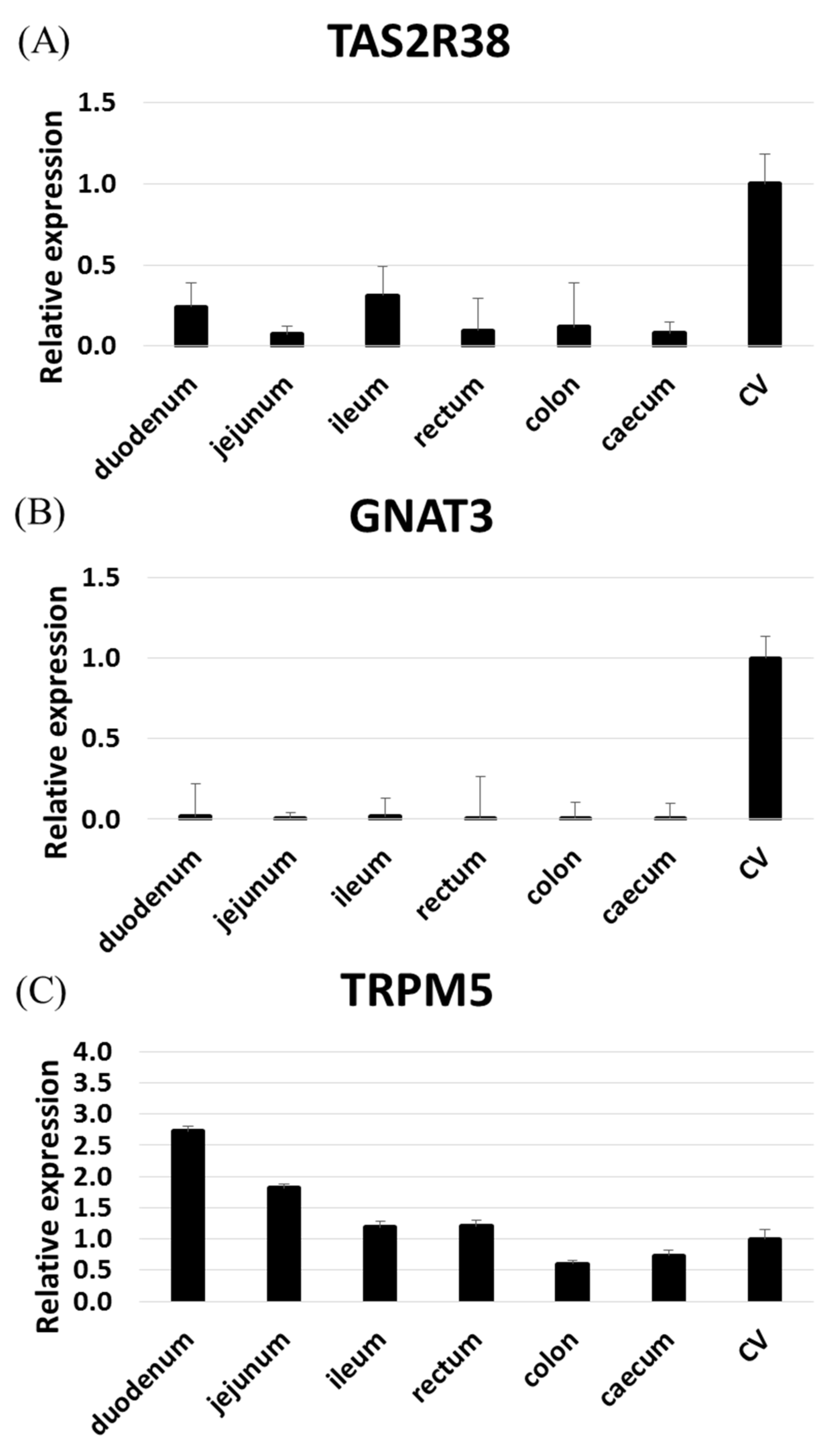
We found that some TAS2Rs are highly expressed in the rhesus macaque gut. In particular, TAS2R1, 3, 4, 5, and 46 were expressed in nearly all gut samples, while TAS2R14, 19, 20, and 38 were expressed in only certain tissues. RNAseq failed to detect other TAS2Rs. Of note, TAS2R38 expression in the intestine is related to the risk of intestinal cancers associated with the taster/non-taster phenotype [10]. To confirm the expression pattern of TAS2R38 in the intestine, we performed RT-qPCR analyses of TAS2R38, GNAT3 (gustducin), and TRPM5 (Tables S2, S3 and Figure 2). TAS2R38 and GNAT3 were both highly expressed in the circumvallate papillae (CV), but TAS2R38 was moderately expressed in the ileum and duodenum as compared with GNAT3 [3]. Meanwhile, TRPM5 was highly expressed in the upper digestive system. These results revealed an expression pattern similar to that in human tissues.
2.2. RNAscope
2.3. Immunohistochemistry
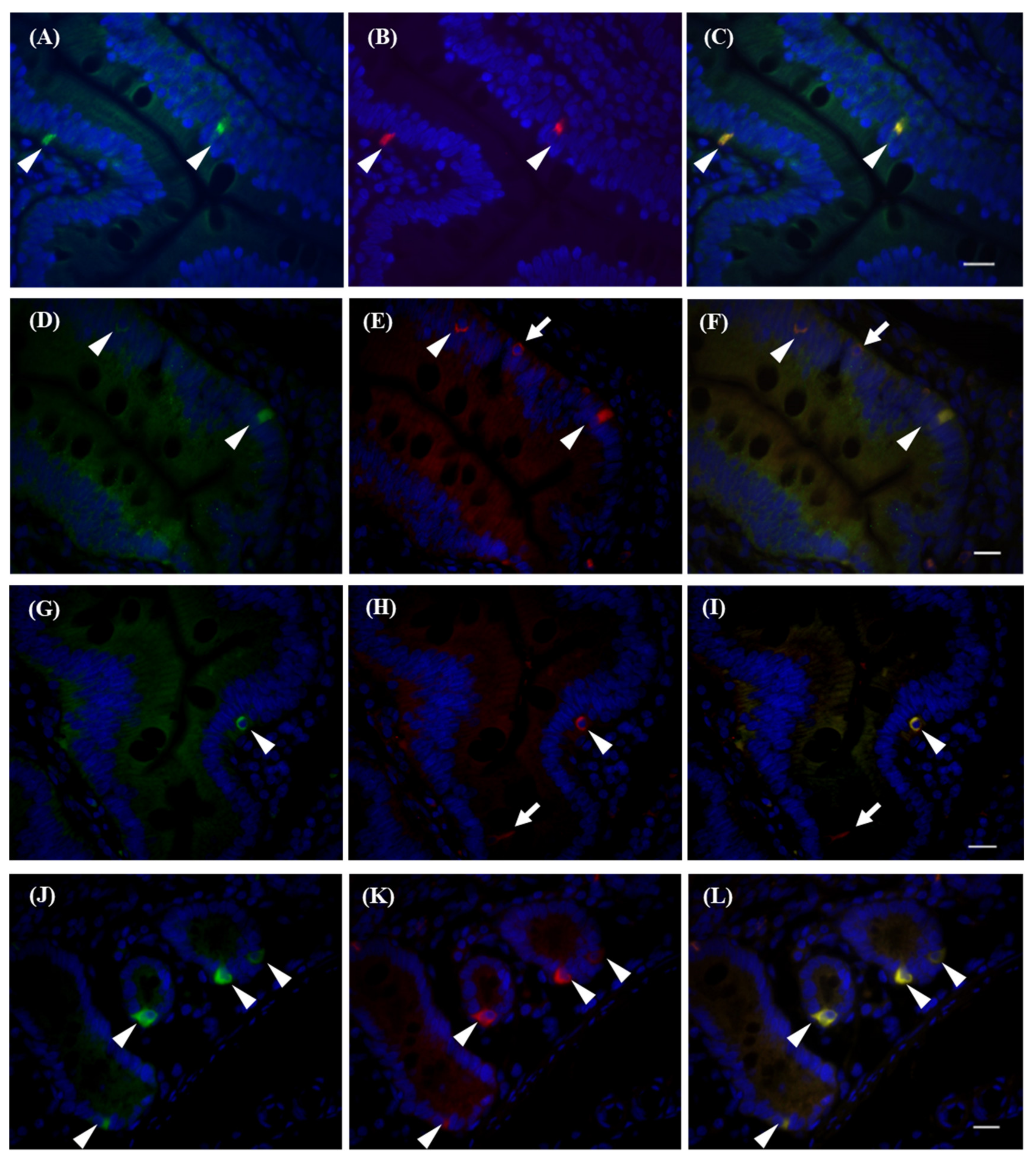
We evaluated T2R38 signals by immunohistochemistry (Figure S1). Using T2R38 antibodies, we detected signals in the CV, and these signals overlapped with GNAT3 signals (Figure S1B,C). To confirm antibody reactivity, we used HEK293 cells transiently transfected with rhesus macaque TAS2R38 cDNA. Transfected cells were then stained using each antibody (Table S4). As shown in Figure S2, T2R38 protein was detected with response to PTC. We confirmed that each antibody could specifically detect T2R38 protein in rhesus macaques. We further examined T2R expression in cells of the intestinal villi in the duodenum that were positive for T2R38. There are several types of cells in the intestinal epithelium and expression of T2Rs is limited to specific types: Paneth cells in the small intestine, [7] goblet cells in the large intestine [11], enteroendocrine cells [9], and tuft cells [12]. T2R38 antibody immunoreactivity was observed in both enteroendocrine cells and goblet cells marked by chromogranin A (arrowheads in Figure 4A–C) [13] and mucin2 (arrowheads in Figure 4D–F) [14], respectively. We also detected co-expression of GNAT3 and T2R38 (arrowheads in Figure 4J-L), making GNAT3-dependent signal transduction in these cells possible. We have previously observed that rhesus macaque T2R38 can activate G-protein with the C-terminus of GNAT3 (G16/gust44) [15] in a heterologous expression system [5], so this may also be possible in intestinal cells. Several DCLK1-positive tuft cells were negative for T2R38 (arrows in Figure 4H,I); however, some cells were double-stained by T2R38 and DCLK1 antibodies (Arrowheads in Figure 4G-I). We did not obtain evidence of T2R38 expression in Paneth cells, possibly due to the functionality of the Paneth cell marker antibody. Similar expression profiles have been observed in the ileum (Figure S3) and colon (Figure S4) of rhesus macaques, suggesting differences from mice. In mice, the Tas2R131 promoter functioned in Paneth cells in the small intestine [7] and in goblet cells in the large intestine [11]. These results indicate that bitter taste receptors, or at least T2R38, are expressed in the intestines of rhesus macaques, as in humans, and have similar functionality. Accordingly, rhesus macaques are suitable models for human research, owing to these similarities in receptivity and expression profiles, at least for T2R38 receptors.
3. Discussion
3.1. Expression Profiles
We screened the expression profiles of TAS2Rs and related molecules in the gut of rhesus macaques. RNAseq revealed that TAS2R1, 3, 4, 5, 20, and 46 were expressed in the duodenum, ileum, and colon, consistent with observations using human cultured cell lines [16] and the RefEX and GTEx databases; the exception is TAS2R1, which was not expressed in humans but was substantially expressed in rhesus macaque tissues. The expression of TAS2R119, an orthologue of primate TAS2R1 in mice, has been reported in the mouse duodenum, jejunum, ileum, and colon [7], suggesting that TAS2R1 exhibits similar expression patterns to TAS2R119 in mice. However, the expression patterns of other genes were very similar to those in humans, suggesting that rhesus macaque is a good model for the human gut. In particular, the expression of TAS2R38 in rhesus macaques was similar to that in humans, with higher levels in the small intestine than in the large intestine, but was not similar to the expression of the mouse orthologue Tas2R138, which is more highly expressed in the large intestine than in the small intestine [7]. The expression patterns of TAS2R3, 4, and 5 were similar among species, while TAS2R5 expression was lost in mice. Owing to the numerous mouse orthologues of human and rhesus macaque TAS2R14 as well as the high number of orthologues of mouse TAS2R46 in humans and rhesus macaques, it is difficult to directly compare the expression levels of these genes between rodents and primates.
3.2. TAS2R-Positive Cells
We observed the expression of TAS2R38 with gustducin in enteroendocrine cells and goblet cells in the rhesus macaque intestine. These results are basically consistent with those of previous studies in mice and humans; mouse T2R131 is expressed in Paneth cells in the small intestine and in goblet cells in the large intestine [7,11]. T2R138, an orthologue of primate T2R38, is expressed in the enteroendocrine cells of the mouse ileum and colon [8]. T2R38 is also expressed in the enteroendocrine cells of the human colon [9]. We found that T2R38 is expressed in DCLK1-positive cells in the intestine. DCLK1 is a marker for tufted cells (reviewed in [17]). In mice, the expression of T2Rs in tuft cells is associated with parasitic infection [12]. T2Rs include T2R138, consistent with our data in rhesus macaques. Therefore, the basic expression patterns are similar to those in other species, although it is necessary to evaluate the expression patterns of each type of T2R. Our results indicate that rhesus macaques would be an appropriate model for analyses of specific phenotypes in the gut.
4. Materials and Methods
4.1. Materials
Samples of rhesus macaque gut were acquired from the stored tissue bank (consisting of tissues obtained from dead rhesus macaques) at the Primate Research Institute, Kyoto University (Permit Numbers: 2016-054 (1 April 2016); 2017-016 (1 April 2017); 2018-004 (1 April 2018)). For the gene expression analysis, we used tissues from six rhesus macaques. Small blocks of tissue were incubated at 4 °C in RNAlater (Agilent Technologies, Santa Clara, CA, USA) overnight and stored at −80 °C until use. For immunohistochemical analysis, we used tissues from three rhesus macaques. Small blocks of tissue were incubated in 4% paraformaldehyde at 4 °C before use. Additional chemicals were purchased from commercial suppliers.
4.2. RNAseq and RT-PCR
Total RNA was extracted from tissues using an RNeasy Plus Mini Kit (Qiagen GmbH, Hilden, Germany) and its quality was confirmed using a Bioanalyzer (Agilent Technologies). Genomic DNA removal and reverse transcription were performed using the PrimeScript RT Reagent Kit with gDNA Eraser (Takara, Kyoto, Japan) for 1 μg of total RNA with and without enzymes. RNAseq was performed at The Research Institute for Microbial Diseases, Osaka University, using a TruSeq Stranded mRNA Sample Prep Kit and HiSeq2500 as the sequencer. Analysis was performed by GeneBay to confirm the annotation of TAS2Rs of RheMac2, as shown in Table S1. Real-time PCR was performed using the StepOne Plus system (Applied Biosystems, Foster City, CA, USA) with Thunderbird SYBR qPCR Mix (Toyobo, Tokyo, Japan) and the primers listed in Supplemental Table S2. Standard conditions (95 °C for 30 s, 50 cycles of 95 °C for 15 s, and 60 °C for 45 s) were applied and analyses were performed in triplicate. Quantities of mRNA for TAS2R38, GNAT3 (gustducin), and TRPM5 were determined relative to the quantities of the internal markers GAPDH and β-actin [18,19]. Amplicons were also analyzed by 2% agarose gel electrophoresis and the presence of single bands was confirmed.
4.3. RNAscope
RNAscope experiments were conducted according to the manufacturer’s instructions (Advanced Cell Diagnostics, Newark, CA). For the manual single-plex RNAscope assays, tissue sections were baked for 1 h at 60 °C. Deparaffinized treatment was performed at room temperature. Target retrieval was performed for 15 min at 100 °C, followed by protease treatment for 30 min at 40 °C. Probes were then hybridized for 2 h at 40 °C. RNAscope amplification was performed by DAB chromogenic detection. Images were acquired by all-in-one microscopy (BZ-X710; Keyence, Osaka, Japan).
4.4. Immunohistochemistry
Stored samples were embedded in paraffin and sectioned at 10 μm. Sections were deparaffinized and rehydrated in a graded alcohol series followed by water. To unmask immunogenic epitopes, antigen retrieval was performed with RNAscope target retrieval reagents (Advanced Cell Diagnostics) diluted to 1:10 in distilled water at 98–100 °C for 15 min. Then, an antigen epitope was fixed in 100% ethanol for 5 min. Nonspecific binding was blocked with 10% normal horse serum (S-2000; Vector Laboratories, Burlingame, CA, USA) diluted in 0.01% Tween-20/0.01 M phosphate-buffered saline (PBS-T) and incubated at room temperature for 1 h. Sections were incubated overnight at 4 °C with primary antibodies (Table S4) diluted in blocking solution (10% normal horse serum/PBS-T). Sections were then incubated with secondary antibodies (Table S4) diluted in blocking solution at room temperature for 1 h. Nuclei were stained by a 4’,6-Diamidino-2-phenylindole (DAPI) solution (Dojindo Molecular Technologies, Tokyo, Japan) diluted at 1:1000 in PBS-T at room temperature for 1 min. Sections were then washed and embedded by Mountant Permafluor (TA-030-FM; Thermo Fisher Scientific, Waltham, MA, USA). Fluorescent images were acquired by a fluorescence microscope (LSM-510, Axioplan 2; Carl Zeiss Microscopy GmbH, Jena, Germany) mounted on a cooled charge-coupled device camera system (Cool SNAP HQ; Photometrics, Tucson, AZ, USA). Image acquisition and processing were performed using an Apple Macintosh Power Mac G4 computer running IPLab spectrum software concatenated with an auto-excitation filter wheel (Scanalytics Inc., Chicago, IL, USA) with channels for DAPI (blue), Cy3 (red), and FITC (green).
4.5. Immunocytochemistry and Calcium Imaging
HEK293 cells were transiently transfected using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol with cDNA constructs coding for the rhesus macaque bitter taste receptor TAS2R38 [5]. The coding regions of the receptors were fused N-terminally to the sst epitope, and C-terminally to a 1D4-tag. An empty pEAK10 vector was used as a negative control. Next, transfected cells were seeded into poly-D-lysine coated chambers and incubated overnight at 37 °C in 5% CO2 [20]. Then, cells were washed with PBS-T, cooled on ice for 30 min, and incubated with biotin-conjugated concanavalin A (5 μg/mL, Sigma) for 1 h at 4 °C. After washing with PBS-T, the cells were fixed by treatment with methanol/acetone (1:1, v/v) for 2 min. Cells were washed again with PBS-T and incubated for 1 h with 10% normal horse serum. Primary antibodies were added at several concentrations and incubated overnight at 4 °C (Table S4). After washing with PBS-T and blocking with 10% normal horse serum, cells were incubated with the secondary antibodies streptavidin and AMCA conjugate for 1 h at room temperature (Table S4). Finally, cells were washed with PBS-T and fluorescence was measured by fluorescence microscopy with channels for DAPI (blue), Cy3 (red), and FITC (green) (LCM-510; Carl Zeiss Microscopy GmbH). Calcium imaging was performed to check the response of T2R38 according to the methods previously reported [5].
Abbreviation
| T2R | Bitter taste receptor protein |
| TAS2R | Bitter taste receptor gene in primates |
| Tas2R | Bitter taste receptor gene in rodents |
| CV | Circumvallate papillae |
| PTC | Phenylthiocarbamide |
| DAPI | 4’,6-Diamidino-2-phenylindole |
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/1422-0067/21/3/902/s1.
Author Contributions
Conceptualization, H.I. and K.M.; methodology, M.H. (Miho Hakukawa); validation, H.I., K.I., and K.M.; investigation, M.H. (Miho Hakukawa); writing—original draft preparation, H.I.; writing—review and editing, K.M.; visualization, M.H. (Miho Hakukawa) and M.H. (Misa Hayashi) supervision, H.I.; project administration, K.M.; funding acquisition, K.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Grants-in-Aid from the Japan Society for the Promotion of Science (KAKENHI 19K21586, 18H04005,16H01338, 15H02421, 15H05242, and 25257409 to HI), and research grants from the Kobayashi International Scholarship Foundation, the Terumo Foundation for Life Sciences and Arts, the Umami Manufacturers Association of Japan, and the JSPS and DG-RSTHE or NSF under the Japan-Indonesia or Korea Research Cooperative Program and Core-to-Core Program to H.I.
Acknowledgments
The authors would like to thank Takashi Ueda, Yoshiro Ishimaru, Takumi Misaka, and Keiko Abe for providing the Ga16/gust44 and pEAK10 vectors, and Hiroaki Matsunami for providing HEK293T cells. We also wish to express our gratitude to the members of the molecular biology section of the Primate Research Institute, Kyoto University. This work was supported by the Cooperative Research Program of the Primate Research Institute, Kyoto University (H31-B17).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Behrens, M.; Meyerhof, W. Gustatory and extragustatory functions of mammalian taste receptors. Physiol. Behav. 2011, 105, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, Y.; Abe, M.; Asakura, T.; Imai, H.; Abe, K. Expression analysis of taste signal transduction molecules in the fungiform and circumvallate papillae of the rhesus macaque, Macaca mulatta. PloS ONE 2012, 7, e45426. [Google Scholar] [CrossRef] [PubMed]
- Gonda, S.; Matsumura, S.; Saito, S.; Go, Y.; Imai, H. Expression of taste signal transduction molecules in the caecum of common marmosets. Biol. Lett. 2013, 9, 20130409. [Google Scholar] [CrossRef] [PubMed]
- Bufe, B.; Breslin, P.A.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.K.; Drayna, D.; Meyerhof, W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashido, S.N.; Hayakawa, T.; Matsui, A.; Go, Y.; Ishimaru, Y.; Misaka, T.; Abe, K.; Hirai, H.; Satta, Y.; Imai, H. Rapid expansion of phenylthiocarbamide non-tasters among Japanese macaques. PloS ONE 2015, 10, e0132016. [Google Scholar] [CrossRef] [Green Version]
- Lossow, K.; Hubner, S.; Roudnitzky, N.; Slack, J.P.; Pollastro, F.; Behrens, M.; Meyerhof, W. Comprehensive analysis of mouse bitter taste receptors reveals different molecular receptive ranges for orthologous receptors in mice and humans. J. Biol. Chem. 2016, 291, 15358–15377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prandi, S.; Voigt, A.; Meyerhof, W.; Behrens, M. Expression profiling of Tas2r genes reveals a complex pattern along the mouse GI tract and the presence of Tas2r131 in a subset of intestinal Paneth cells. Cell. Mol. Life Sci. 2018, 75, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Vegezzi, G.; Anselmi, L.; Huynh, J.; Barocelli, E.; Rozengurt, E.; Raybould, H.; Sternini, C. Diet-induced regulation of bitter taste receptor subtypes in the mouse gastrointestinal tract. PloS ONE 2014, 9, e107732. [Google Scholar] [CrossRef] [PubMed]
- Latorre, R.; Huynh, J.; Mazzoni, M.; Gupta, A.; Bonora, E.; Clavenzani, P.; Chang, L.; Mayer, E.A.; De Giorgio, R.; Sternini, C. Expression of the bitter taste receptor, T2R38, in enteroendocrine cells of the colonic mucosa of overweight/obese vs. lean subjects. PloS ONE 2016, 11, e0147468. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, M.; Saito, H.; Isono, K.; Goto, T.; Shirakawa, H.; Shoji, N.; Satoh-Kuriwada, S.; Sasano, T.; Okada, R.; Kudoh, K.; et al. Genotyping analysis of bitter-taste receptor genes TAS2R38 and TAS2R46 in Japanese patients with gastrointestinal cancers. J. Nutr. Sci. Vitaminol. 2017, 63, 148–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prandi, S.; Bromke, M.; Hubner, S.; Voigt, A.; Boehm, U.; Meyerhof, W.; Behrens, M. A subset of mouse colonic goblet cells expresses the bitter taste receptor Tas2r131. PloS ONE 2013, 8, e82820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.C.; Chen, Z.H.; Xue, J.B.; Zhao, D.X.; Lu, C.; Li, Y.H.; Li, S.M.; Du, Y.W.; Liu, Q.; Wang, P.; et al. Infection by the parasitic helminth Trichinella spiralis activates a Tas2r-mediated signaling pathway in intestinal tuft cells. Proc. Natl. Acad. Sci. USA 2019, 116, 5564–5569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konno, T.; Kotani, T.; Setiawan, J.; Nishigaito, Y.; Sawada, N.; Imada, S.; Saito, Y.; Murata, Y.; Matozaki, T. Role of lysophosphatidic acid in proliferation and differentiation of intestinal epithelial cells. PloS ONE 2019, 14, e0215255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finch, A.J.; Soucek, L.; Junttila, M.R.; Swigart, L.B.; Evan, G.I. Acute overexpression of Myc in intestinal epithelium recapitulates some but not all the changes elicited by Wnt/beta-catenin pathway activation. Mol. Cell Biol. 2009, 29, 5306–5315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, T.; Ugawa, S.; Yamamura, H.; Imaizumi, Y.; Shimada, S. Functional interaction between T2R taste receptors and G-protein alpha subunits expressed in taste receptor cells. J. Neurosci. 2003, 23, 7376–7380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaggupilli, A.; Singh, N.; Upadhyaya, J.; Sikarwar, A.S.; Arakawa, M.; Dakshinamurti, S.; Bhullar, R.P.; Duan, K.; Chelikani, P. Analysis of the expression of human bitter taste receptors in extraoral tissues. Mol. Cell. Biochem. 2017, 426, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Middelhoff, M.; Westphalen, C.B.; Hayakawa, Y.; Yan, K.S.; Gershon, M.D.; Wang, T.C.; Quante, M. Dclk1-expressing tuft cells: Critical modulators of the intestinal niche? Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G285–G299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034.1–0034.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, M.; Born, S.; Redel, U.; Voigt, N.; Schuh, V.; Raguse, J.D.; Meyerhof, W. Immunohistochemical detection of TAS2R38 protein in human taste cells. PloS ONE. 2012, 7, e40304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Expression levels of TAS2Rs as measured by RNAseq, which was performed using samples from the rhesus macaque duodenum (A), ileum (B), and colon (C). Each n = 3.
Figure 1.
Expression levels of TAS2Rs as measured by RNAseq, which was performed using samples from the rhesus macaque duodenum (A), ileum (B), and colon (C). Each n = 3.

Figure 2.
Expression levels of TAS2R38 (A), GNAT3 (B), and TRPM5 (C) in rhesus macaque tissue samples (n = 3) as determined by qRT-PCR. The expression levels of TAS2Rs were normalized against the expression in circumvallate papillae (CV) (set to 1) using GAPDH and beta-actin as reference.
Figure 2.
Expression levels of TAS2R38 (A), GNAT3 (B), and TRPM5 (C) in rhesus macaque tissue samples (n = 3) as determined by qRT-PCR. The expression levels of TAS2Rs were normalized against the expression in circumvallate papillae (CV) (set to 1) using GAPDH and beta-actin as reference.

Figure 3.
The mRNA localization in the CV and gut, as determined by RNAscope. Probes for TAS2R38 (blue) and TRPM5 (red) were hybridized and visualized in the rhesus macaque CV (A), duodenum (B), ileum (C), and colon (D). Arrowheads indicate TAS2R38+/TRPM5+ cells. Bar = 50 µm.
Figure 3.
The mRNA localization in the CV and gut, as determined by RNAscope. Probes for TAS2R38 (blue) and TRPM5 (red) were hybridized and visualized in the rhesus macaque CV (A), duodenum (B), ileum (C), and colon (D). Arrowheads indicate TAS2R38+/TRPM5+ cells. Bar = 50 µm.

Figure 4.
Immunohistochemical detection of T2R38-positive cells in the duodenum. T2R38-positive cells were visualized by antibodies against T2R38 (green in A, D, G, J), chromogranin A (red in B), Mucin2 (red in E), DCLK1 (red in H), gustducin (red in K), and DAPI (blue) with overlay (C, F, I, L). Arrowheads indicate T2R38+/marker+ cells. Arrows indicate T2R38−/marker+ cells. Bar = 10 µm.
Figure 4.
Immunohistochemical detection of T2R38-positive cells in the duodenum. T2R38-positive cells were visualized by antibodies against T2R38 (green in A, D, G, J), chromogranin A (red in B), Mucin2 (red in E), DCLK1 (red in H), gustducin (red in K), and DAPI (blue) with overlay (C, F, I, L). Arrowheads indicate T2R38+/marker+ cells. Arrows indicate T2R38−/marker+ cells. Bar = 10 µm.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Imai, H.; Hakukawa, M.; Hayashi, M.; Iwatsuki, K.; Masuda, K. Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates. Int. J. Mol. Sci. 2020, 21, 902. https://doi.org/10.3390/ijms21030902
AMA Style
Imai H, Hakukawa M, Hayashi M, Iwatsuki K, Masuda K. Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates. International Journal of Molecular Sciences. 2020; 21(3):902. https://doi.org/10.3390/ijms21030902
Chicago/Turabian StyleImai, Hiroo, Miho Hakukawa, Misa Hayashi, Ken Iwatsuki, and Katsuyoshi Masuda. 2020. "Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates" International Journal of Molecular Sciences 21, no. 3: 902. https://doi.org/10.3390/ijms21030902
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.