Melatonin Alleviates Drought Stress by a Non-Enzymatic and Enzymatic Antioxidative System in Kiwifruit Seedlings

 ,
,
Abstract
:1. Introduction
2. Results
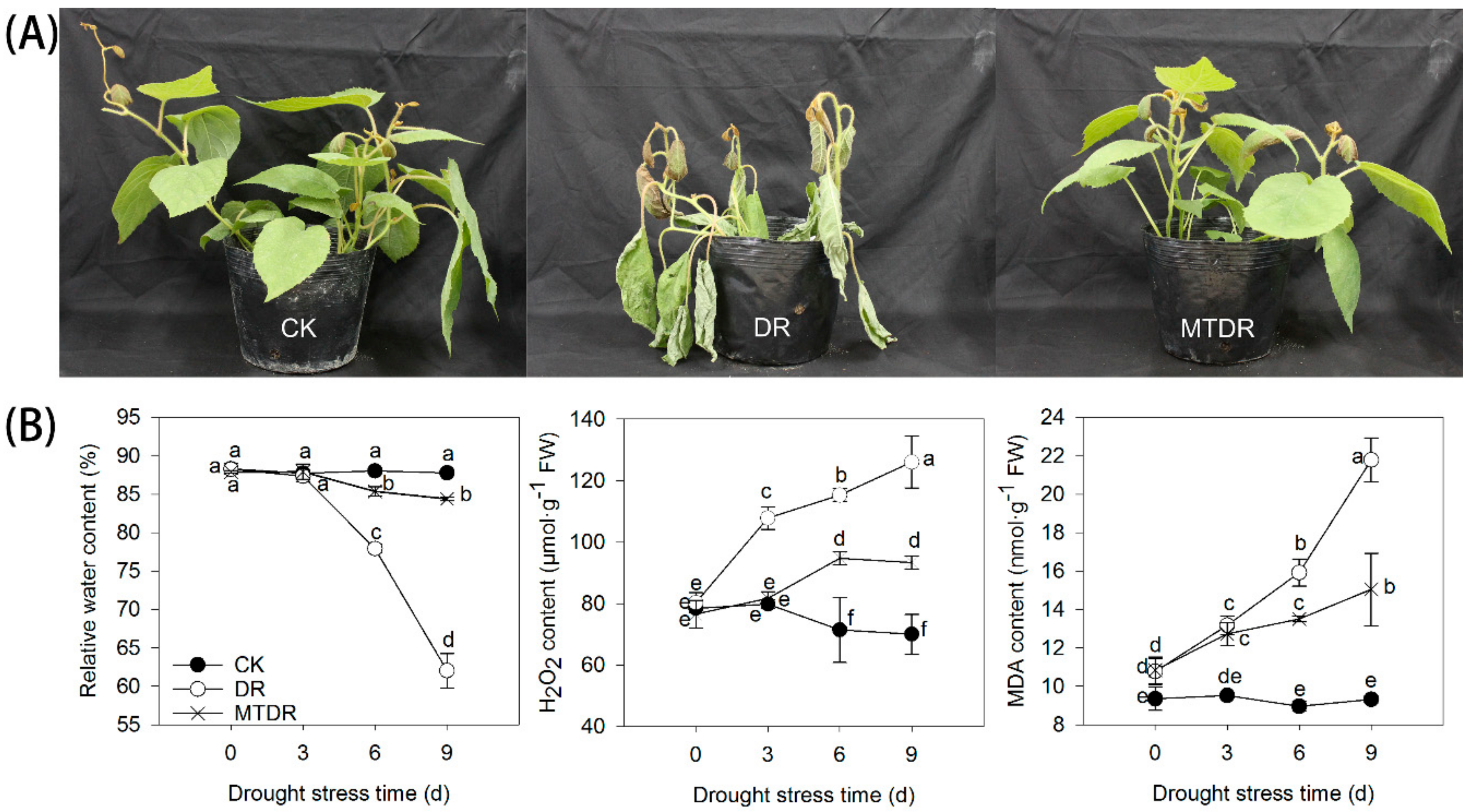
2.1. Morphologic and Physiologic Responses to Drought Treated by Melatonin
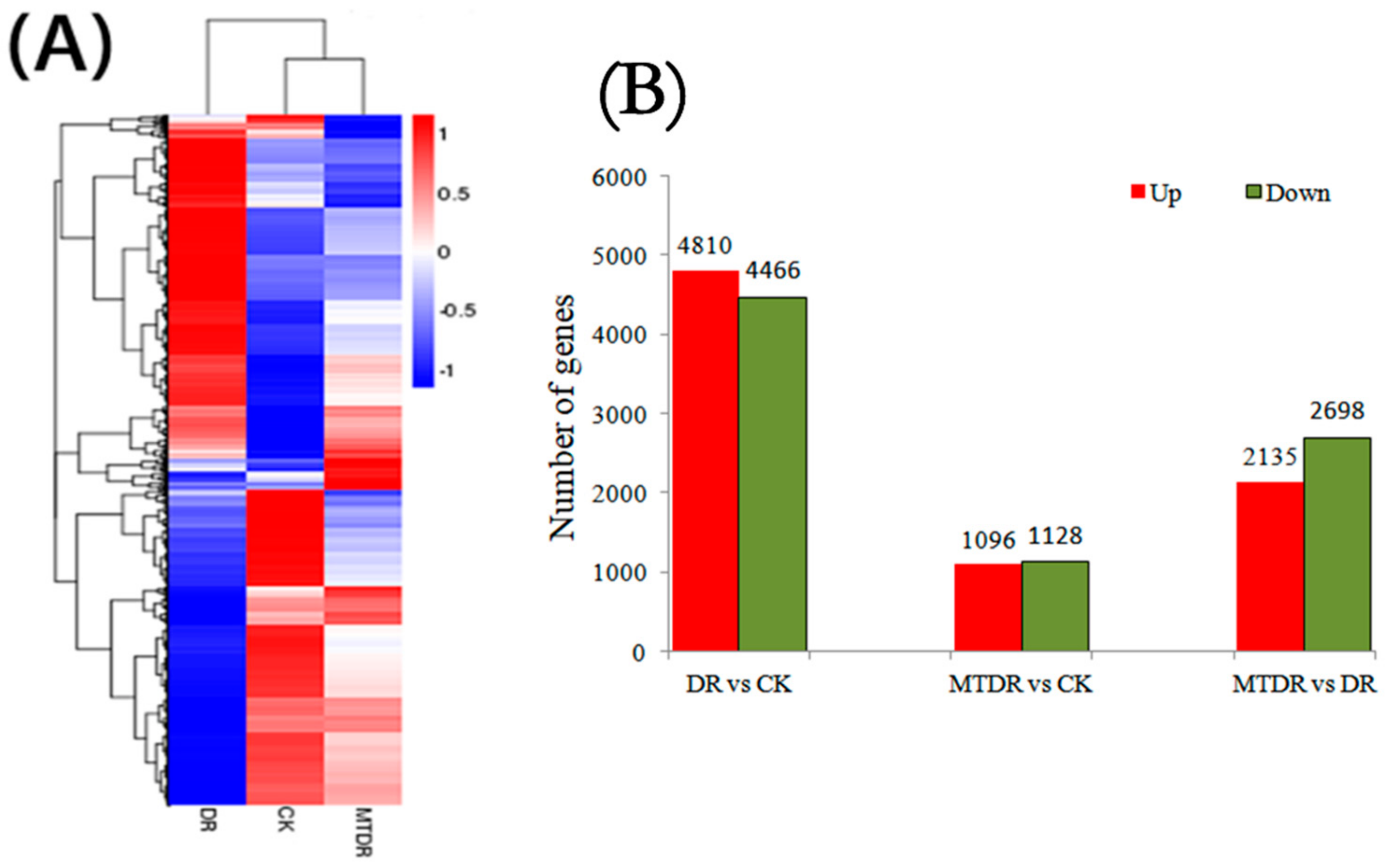
2.2. Transcriptome Profiling and Quality
2.3. Gene Expression Analysis
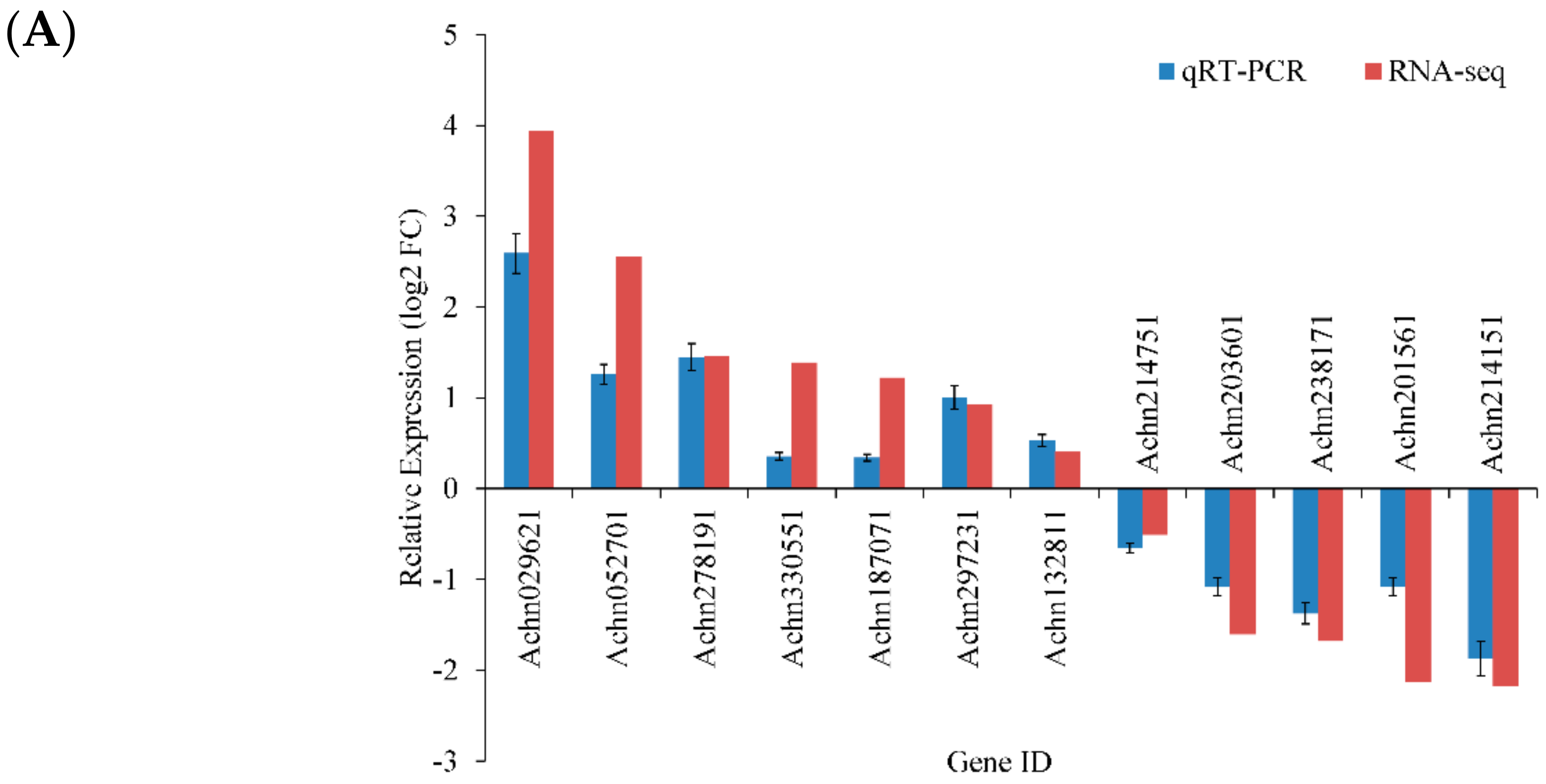
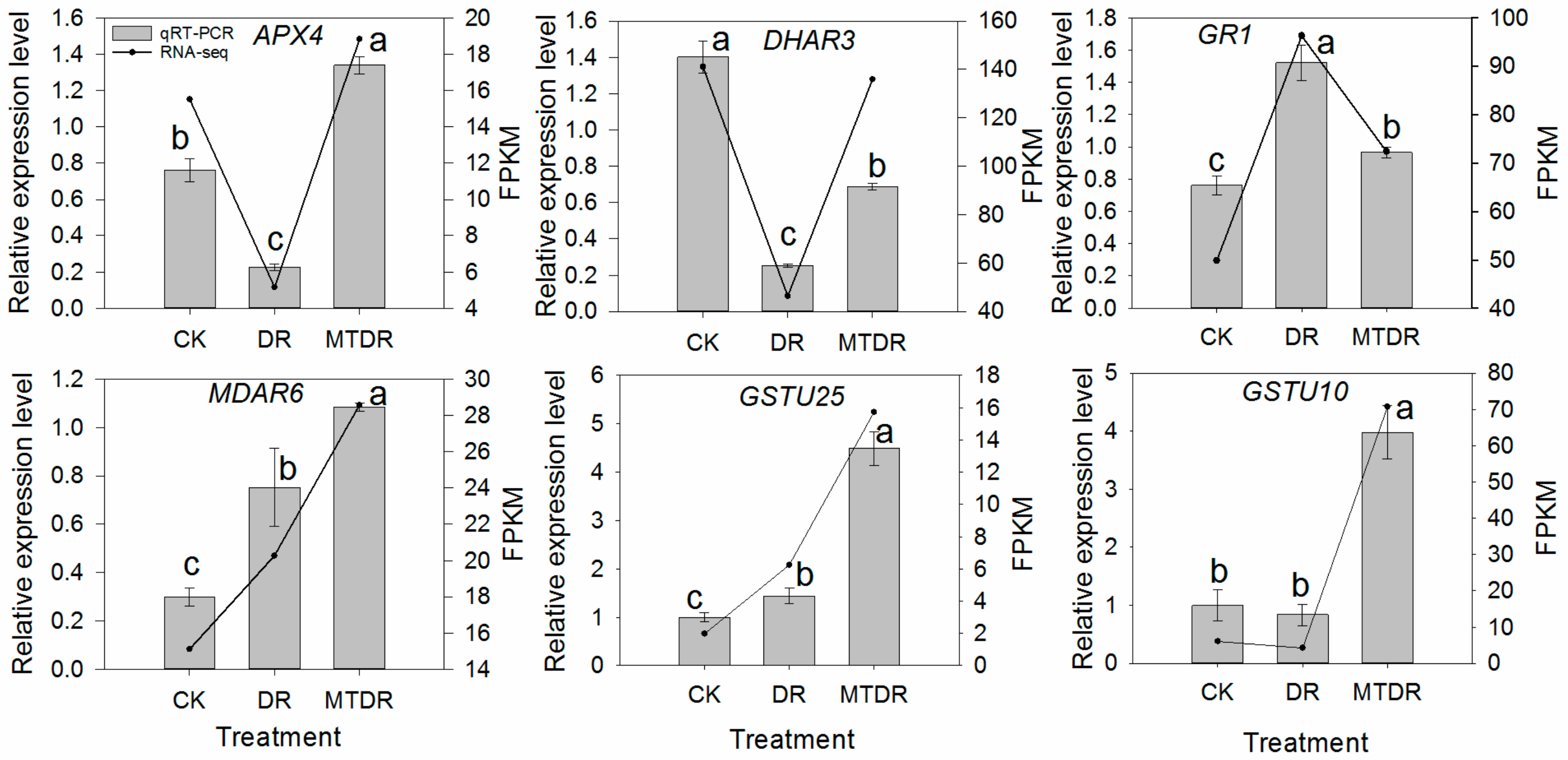
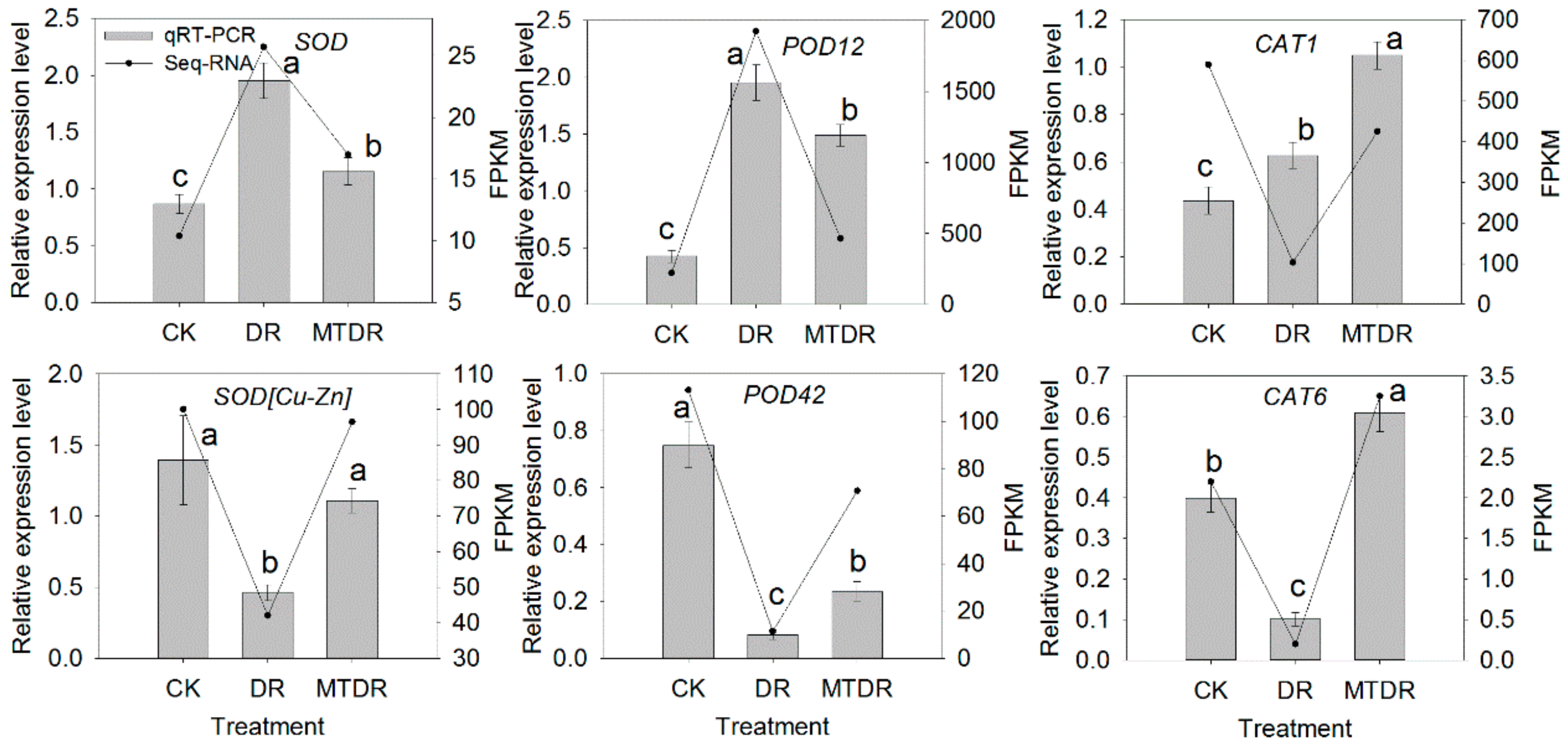
2.4. Verification of RNA-Seq Data by qRT-PCR
2.5. Function Classification of DEGs
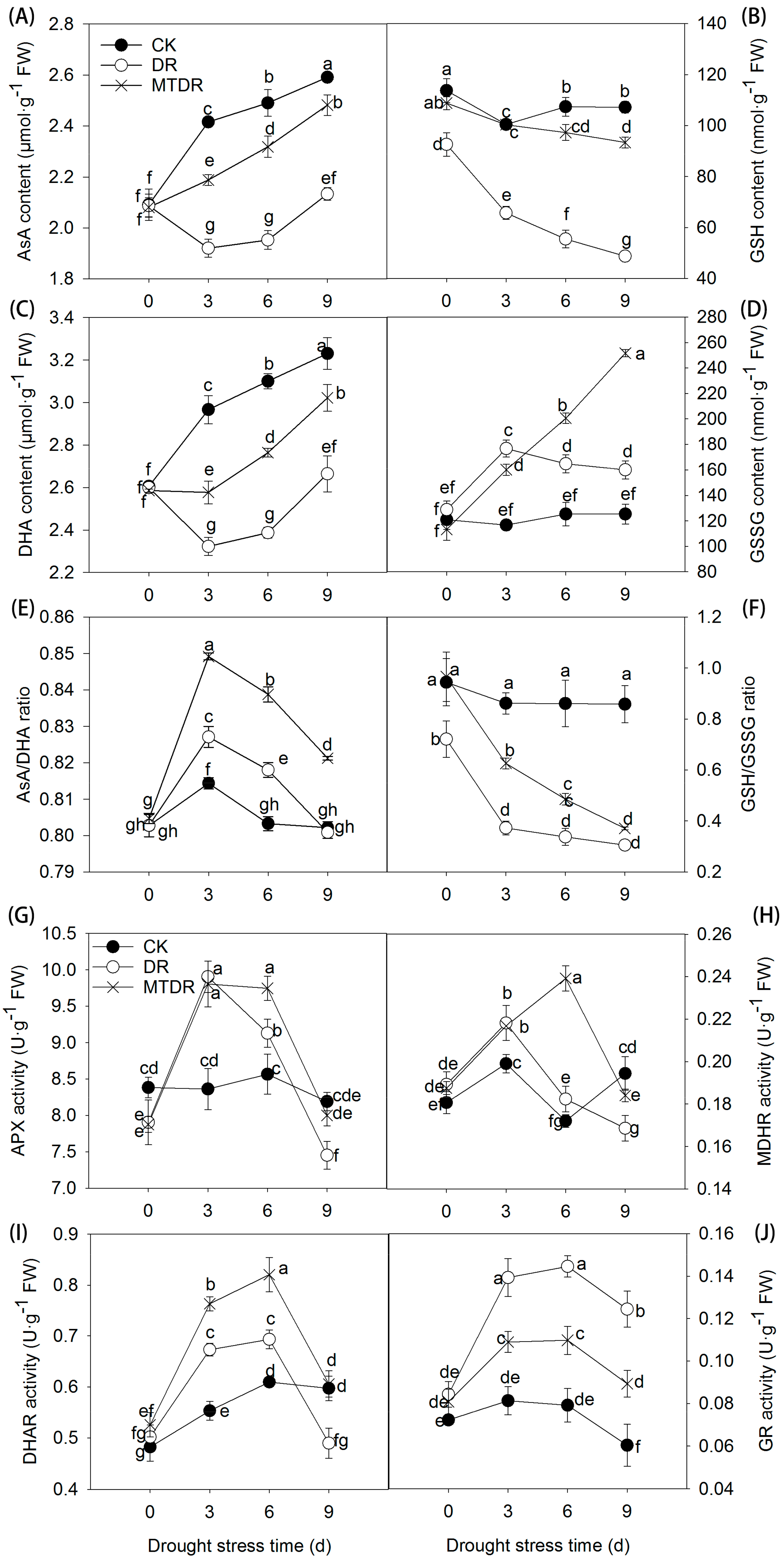
2.6. Effects of Melatonin Pretreatment on AsA Metabolism
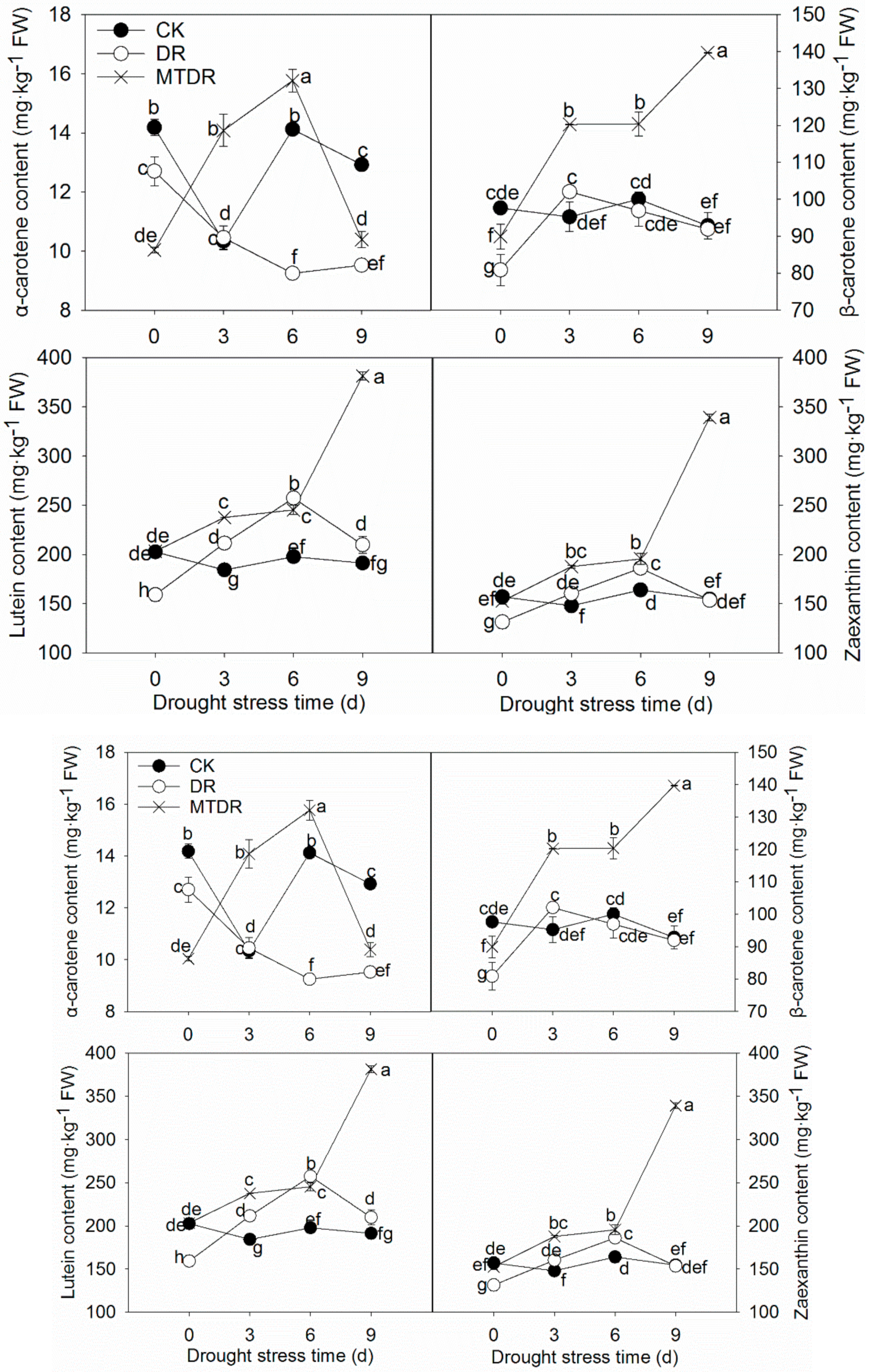
2.7. Effects of Melatonin Pretreatment on Carotenoid Metabolism
2.8. Effects of Melatonin Pretreatment on Antioxidant Enzymes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Experiment Design
4.3. Measurement of Leaf Relative Water Content
4.4. Determination of Hydrogen Peroxide (H2O2)
4.5. Assays for Lipid Peroxidation
4.6. Estimation Metabolin Content and Enzyme Activity of the AsA-GSH Cycle
4.7. Assays of Antioxidative Enzyme Activities
4.8. HPLC Analysis of Carotenoids
4.9. RNA Extraction and mRNA Sequencing
4.10. Validation by Quantitative Real-Time PCR (qRT-PCR)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, R.X.; Xin, L.F.; Zheng, H.F.; Li, L.L.; Ran, W.L.; Mao, J.; Yang, Q.H. Changes in chloroplast ultrastructure in leaves of drought-stressed maize inbred lines. Photosynthetica 2016, 54, 74–80. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Fang, J.; Chen, X.; Zhao, Z.; Li, Y.; Meng, Y.; Huang, L. Actinidia chinensis Planch.: A review of chemistry and pharmacology. Front. Pharmacol. 2019, 10, 1236. [Google Scholar] [CrossRef] [PubMed]
- Mills, T.M.; Li, J.; Behboudian, M.H. Physiological responses of gold kiwifruit (Actinidia chinensis) to reduced irrigation. J. Am. Soc. Hortic. Sci. 2009, 134, 677–683. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, F.; Li, M.; Liang, D.; Zou, J. Physiological responses of kiwifruit plants to exogenous ABA under drought conditions. Plant Growth Regul. 2011, 64, 63–74. [Google Scholar] [CrossRef]
- Dichio, B.; Montanaro, G.; Sofo, A.; Xiloyannis, C. Stem and whole-planthydraulics in olive (Olea europaea) and kiwifruit (Actinidia deliciosa). Trees 2013, 27, 183–191. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Huang, Y.; Bie, Z.L.; Ahmed, W.; Reiter, R.J.; Niu, M.L.; Hameed, S. Melatonin: Current status and future perspectives in plant science. Front. Plant Sci. 2016, 6, 1230. [Google Scholar] [CrossRef] [Green Version]
- Sharif, R.; Xie, C.; Zhang, H.Q.; Arnao, M.B.; Ali, M.; Li, Y.H. Melatonin and its effects on plant systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A new plant hormone and/or a plant master regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zhang, Y.; Feng, Z.; Bai, Q.; He, J.; Wang, Y. Effects of melatonin on antioxidant capacity in naked oat seedlings under drought stress. Molecules 2018, 23, 1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Sun, X.; Li, C.; Wei, Z.; Liang, D.; Ma, F. Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J. Pineal Res. 2013, 54, 292–302. [Google Scholar] [CrossRef]
- Liang, B.; Ma, C.; Zhang, Z.; Wei, Z.; Gao, T.; Zhao, Q.; Ma, F.; Li, C. Long-term exogenous application of melatonin improves nutrient uptake fluxes in apple plants under moderate drought stress. Environ. Exp. Bot. 2018, 155, 650–661. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, B.; Zhang, H.J.; Weeda, S.; Yang, C.; Yang, Z.C.; Ren, S.; Guo, Y.D. Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.). J. Pineal Res. 2013, 54, 15–23. [Google Scholar] [CrossRef]
- Meng, J.F.; Xu, T.F.; Wang, Z.Z.; Fang, Y.L.; Xi, Z.M.; Zhang, Z.W. The ameliorative effects of exogenous melatonin on grape cuttings under water-deficient stress: Antioxidant metabolites, leaf anatomy, and chloroplast morphology. J. Pineal Res. 2014, 57, 200–212. [Google Scholar] [CrossRef]
- Li, C.; Tan, D.X.; Liang, D.; Chang, C.; Jia, D.; Ma, F. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 2015, 66, 669–680. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, W.; Wang, L.; Sun, Y. Exogenous melatonin improves seedling health index and drought tolerance in tomato. Plant Growth Regul. 2015, 77, 317–326. [Google Scholar] [CrossRef]
- Ding, F.; Wang, G.; Wang, M.; Zhang, S. Exogenous melatonin improves tolerance to water deficit by promoting cuticle formation in tomato plants. Molecules 2018, 23, 1605. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Wang, S.; Deng, X.; Yin, L.; Xiong, B.; Wang, X. Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage. Acta Physiol. Plant. 2016, 38, 48. [Google Scholar] [CrossRef]
- Cui, G.B.; Zhao, X.X.; Liu, S.D.; Sun, F.L.; Zhang, C.; Xi, Y.J. Beneficial effects of melatonin in overcoming drought stress in wheat seedlings. Plant Physiol Bioch. 2017, 118, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhang, J.; Burgess, P.; Rossi, S.; Huang, B. Interactive effects of melatonin and cytokinin on alleviating drought-induced leaf senescence in creeping bentgrass (Agrostis stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Liang, D.; Ni, Z.; Xia, H.; Xie, Y.; Lv, X.; Wang, J.; Lin, L.; Deng, Q.; Luo, X. Exogenous melatonin promotes biomass accumulation and photosynthesis of kiwifruit seedlings under drought stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Campos, C.N.; Ávila, R.G.; de Souza, K.R.D.; Azevedo, L.M.; Alves, J.D. Melatonin reduces oxidative stress and promotes drought tolerance in young Coffea arabica L. plants. Agric. Water Manag. 2019, 211, 37–47. [Google Scholar] [CrossRef]
- Yang, W.J.; Du, Y.T.; Zhou, Y.B.; Chen, J.; Xu, Z.S.; Ma, Y.Z.; Chen, M.; Min, D.H. Overexpression of TaCOMT improves melatonin production and enhances drought tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2019, 20, 652. [Google Scholar] [CrossRef] [Green Version]
- Zuo, B.; Zheng, X.; He, P.; Wang, L.; Lei, Q.; Feng, C.; Zhou, J.; Li, Q.; Han, Z.; Kong, J. Overexpression of MzASMT improves melatonin production and enhances drought tolerance in transgenic Arabidopsis thaliana plants. J. Pineal Res. 2014, 57, 408–417. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Wei, W.; Li, Q.T.; Chu, Y.N.; Reiter, R.J.; Yu, X.M.; Zhu, D.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, J.S.; et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2014, 66, 695–707. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Li, D.X.; Zhang, J.R.; Shan, C.; Rengel, Z.; Song, Z.B.; Chen, Q. Phytomelatonin Receptor PMTR1-Mediated Signaling Regulates Stomatal Closure in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12500. [Google Scholar] [CrossRef] [PubMed]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mibei, E.K.; Ambuko, J.; Giovannoni, J.J.; Onyango, A.N.; Owino, W.O. Carotenoid profiling of the leaves of selected African eggplant accessions subjected to drought stress. Food Sci. Nutrit. 2017, 5, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Okunlola, G.O.; Olatunji, O.A.; Akinwale, R.O.; Tariq, A.; Adelusi, A.A. Physiological response of the three most cultivated pepper species (Capsicum spp.) in Africa to drought stress imposed at three stages of growth and development. Sci. Hortic. 2017, 224, 198–205. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Terron, M.P.; Flores, L.J.; Czarnocki, Z. Melatonin and its metabolites: New findings regarding their production and their radical scavenging actions. Acta Biochim. Pol. 2007, 54, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Antoniou, C.; Chatzimichail, G.; Xenofontos, R.; Pavlou, J.J.; Panagiotou, E.; Christou, A.; Fotopoulos, V. Melatonin systemically ameliorates drought stress-induced damage in Medicago sativa plants by modulating nitro-oxidative homeostasis and proline metabolism. J. Pineal Res. 2017, 62, e12401. [Google Scholar] [CrossRef]
- Zou, Q. Experimental Manual on Plant Physiology; Chinese Agriculture Press: Beijing, China, 2000; pp. 127–130. [Google Scholar]
- Liang, D.; Shen, Y.; Ni, Z.; Wang, Q.; Lei, Z.; Xu, N.; Deng, Q.; Lin, L.; Wang, J.; Lv, X.L.; et al. Exogenous melatonin application delays senescence of kiwifruit leaves by regulating the antioxidant capacity and biosynthesis of flavonoids. Front. Plant Sci. 2018, 9, 426. [Google Scholar] [CrossRef]
- Liang, D.; Gao, F.; Ni, Z.Y.; Lin, L.J.; Deng, Q.X.; Tang, Y.; Wang, X.; Luo, X.; Xia, H. Melatonin improves heat tolerance in kiwifruit seedlings through promoting antioxidant enzymatic activity and glutathione S-transferase transcription. Molecules 2018, 23, 584. [Google Scholar] [CrossRef] [Green Version]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases, II. purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol. 1977, 59, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Scebba, F.; Sebastiani, L.; Vitagliano, C. Activities of antioxidant enzymes during senescence of Prunus armeniaca leaves. Biol. Plant. 2001, 44, 41–46. [Google Scholar] [CrossRef]
- Yan, J.; Cai, Z.; Shen, Z.; Zhang, B.; Ma, R.; Yu, M. Extraction and analytical methods of carotenoids in fruit of yellow flesh peach. J. Fruit Sci. 2015, 32, 1267–1274. [Google Scholar]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-a python framework to work with high throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Ferradás, Y.; Rey, L.; Martínez, Ó.; Rey, M.; González, M.V. Identification and validation of reference genes for accurate normalization of real-time quantitative PCR data in kiwifruit. Plant Physiol. Biochem. 2016, 102, 27–36. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads | Clean Reads | Clean Bases (G) | Q20 (%) | GC Content (%) | Total Mapped (%) | Uniquely Mapped (%) |
|---|---|---|---|---|---|---|---|
| CK | 47,300,498 | 45,707,847 | 6.86 | 97.15 | 45.90 | 68.56 | 66.16 |
| DR | 49,237,617 | 46,895,636 | 7.04 | 97.12 | 46.30 | 67.71 | 65.18 |
| MTDR | 54,138,373 | 51,348,163 | 7.71 | 97.26 | 45.86 | 68.33 | 65.74 |
| KEGG ID | Pathway | Number of Transcripts | p Value |
|---|---|---|---|
| ath00480 | Glutathione metabolism | 24 | 0.0001 |
| ath00053 | Ascorbate and aldarate metabolism | 12 | 0.0023 |
| ath00906 | Carotenoid metabolism | 9 | 0.0057 |
| Gene Name | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| SOD | AAAGGCGGGCTAGGGTTAGG | TGGAAGATCCGGGAGCGATA |
| SOD[Cu-Zn] | GCGGGTGACCTGGGAAACAT | AGGCTCTGCCGACGACTGAA |
| POD12 | CTGCCCAGCACTAGACACAA | GTCCTGGTCGGACGTAAAAA |
| POD42 | CCGAACGCGGTCCAGTATGT | TTGGTCCTCTTGTCGGTGGC |
| CAT1 | ACCTGAGTGCCCTTTAAGCC | TTTGGGTATGAACGAGTTGG |
| CAT6 | CTACATCCGCATCACCTTCG | TATCAGATTCGCTCCCGTCA |
| APX4 | GCTGTCATGTCCGCATTCTT | GGTTTCTCGGCTTTGTTGGT |
| APX6 | TCTTCACAGCTTTCGCATCT | AGCATTAGCACGGTATCCTT |
| MDAR4 | GACTACCTGCCGTTCTTCTA | CTATCTCGCCTACACCATCT |
| MDAR6 | CGCTGGAGAAAGCACAGAAA | GCGAGGGAAGGAGTAAACAA |
| DHAR3 | AAAACATCTCCCTTACGACA | CTTTACCTTCTGGGCTTATT |
| GR1 | CCTAATGAAGTCGAGGTGAC | GCCAGTTGCGATGAGTATGT |
| AO | CTCTACTCCAAGCCATTTCG | CTCCCTCTGACACTTACCG |
| GME | TTTGGCATTGAATGTAGG | AGGGTGTTGTCCGAGTTT |
| VTC2 | CGCTACGATGTCACTGCT | CCTTGTCAACACGAAACTC |
| MIOX4 | ATTTATGCGGAAGGCTGTG | GCCGAAGGTAGGGTGGTT |
| MIOX5 | ATGATTGTATGGGTTGGA | GATGTCTGGGTTCTGCTC |
| GSTL2 | GCCTGGTACAAGGAGAAA | CCAACAATAAGGGATGAAAT |
| GSTU10 | TACAACCCAGTCCACAAA | CATTCCCTCTTCCACCAC |
| GSTU18 | GTCCCTCCATCCTCCCTT | TCTCCTTGTGCCGTCCTA |
| GSTU19 | TATGTTTGGGATGAGGGT | CTGGTAAGGGTCAGAGGG |
| GSTU25 | GATACATAACGGAAAGCC | ACATAATCAGCCCAGAAC |
| PSY | TATAACGCTGCCTTGGCTCT | TTTCCGGCAAATATGTCCTC |
| PDS | CTTTGCATGCCAATAGCAGA | GTCGGACTTCACCACCAAGT |
| LYC | AGGAAGCAGCTGAAATCCAA | AGCCAGTCGCATCAAGAACT |
| NPQ | TTTTGCCTCTGAGCATTGTG | TCTACAAGGGGTGGTTCAGG |
| CYP | TAGCAAGCAACTCCGTGATG | ACTGCAGCTGATGTTTCGTG |
| ZDS | AGGAAGCAGCTGAAATCCAA | AGCCAGTCGCATCAAGAACT |
| Actin | TGCATGAGCGATCAAGTTTCAAG | TGTCCCATGTCTGGTTGATGACT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, H.; Ni, Z.; Hu, R.; Lin, L.; Deng, H.; Wang, J.; Tang, Y.; Sun, G.; Wang, X.; Li, H.; et al. Melatonin Alleviates Drought Stress by a Non-Enzymatic and Enzymatic Antioxidative System in Kiwifruit Seedlings. Int. J. Mol. Sci. 2020, 21, 852. https://doi.org/10.3390/ijms21030852
Xia H, Ni Z, Hu R, Lin L, Deng H, Wang J, Tang Y, Sun G, Wang X, Li H, et al. Melatonin Alleviates Drought Stress by a Non-Enzymatic and Enzymatic Antioxidative System in Kiwifruit Seedlings. International Journal of Molecular Sciences. 2020; 21(3):852. https://doi.org/10.3390/ijms21030852
Chicago/Turabian StyleXia, Hui, Zhiyou Ni, Rongping Hu, Lijin Lin, Honghong Deng, Jin Wang, Yi Tang, Guochao Sun, Xun Wang, Huanxiu Li, and et al. 2020. "Melatonin Alleviates Drought Stress by a Non-Enzymatic and Enzymatic Antioxidative System in Kiwifruit Seedlings" International Journal of Molecular Sciences 21, no. 3: 852. https://doi.org/10.3390/ijms21030852