Effect of Baicalin-Aluminum Complexes on Fecal Microbiome in Piglets

Abstract
:1. Introduction
2. Results
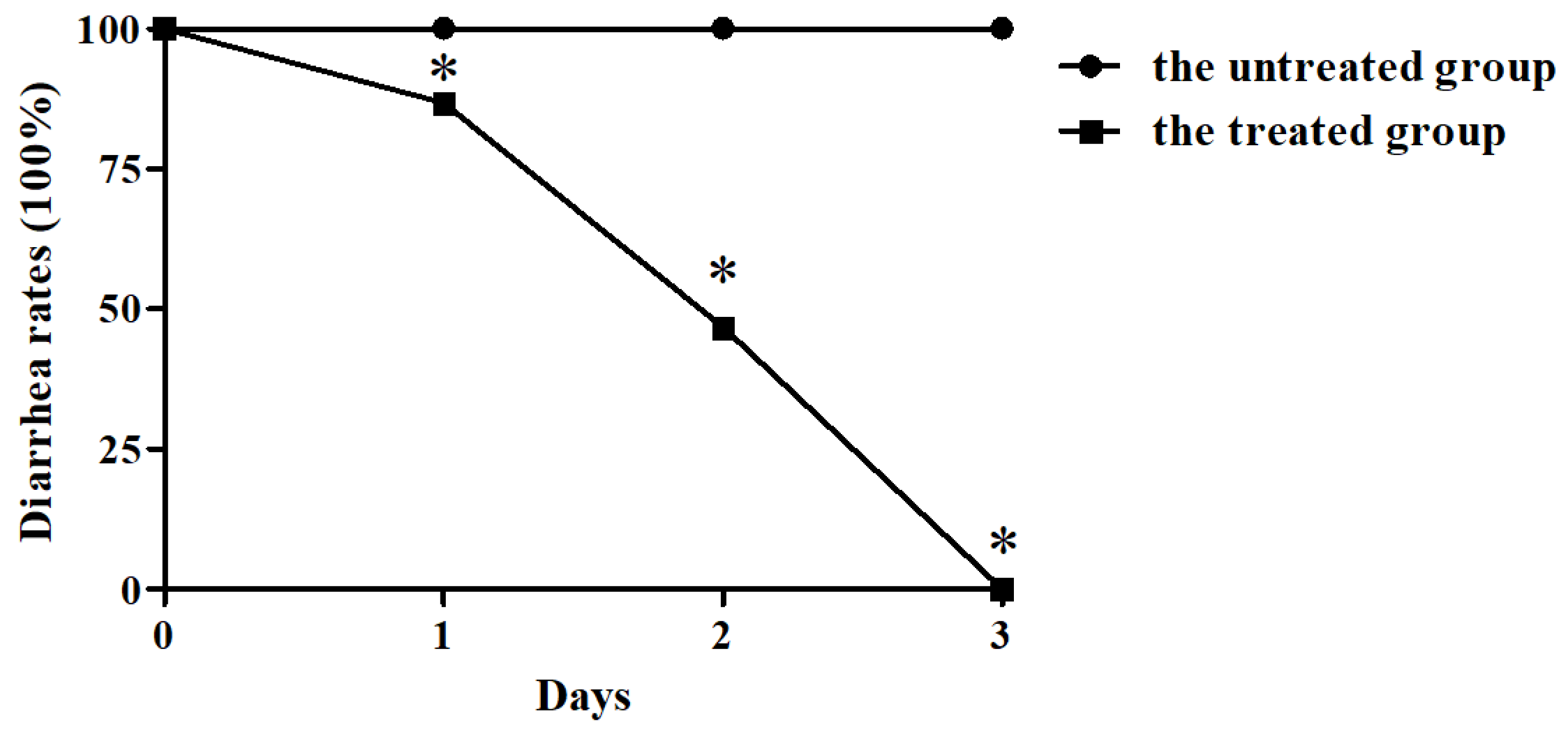
2.1. BBA Significantly Reduced the Diarrhea Rates of Piglets
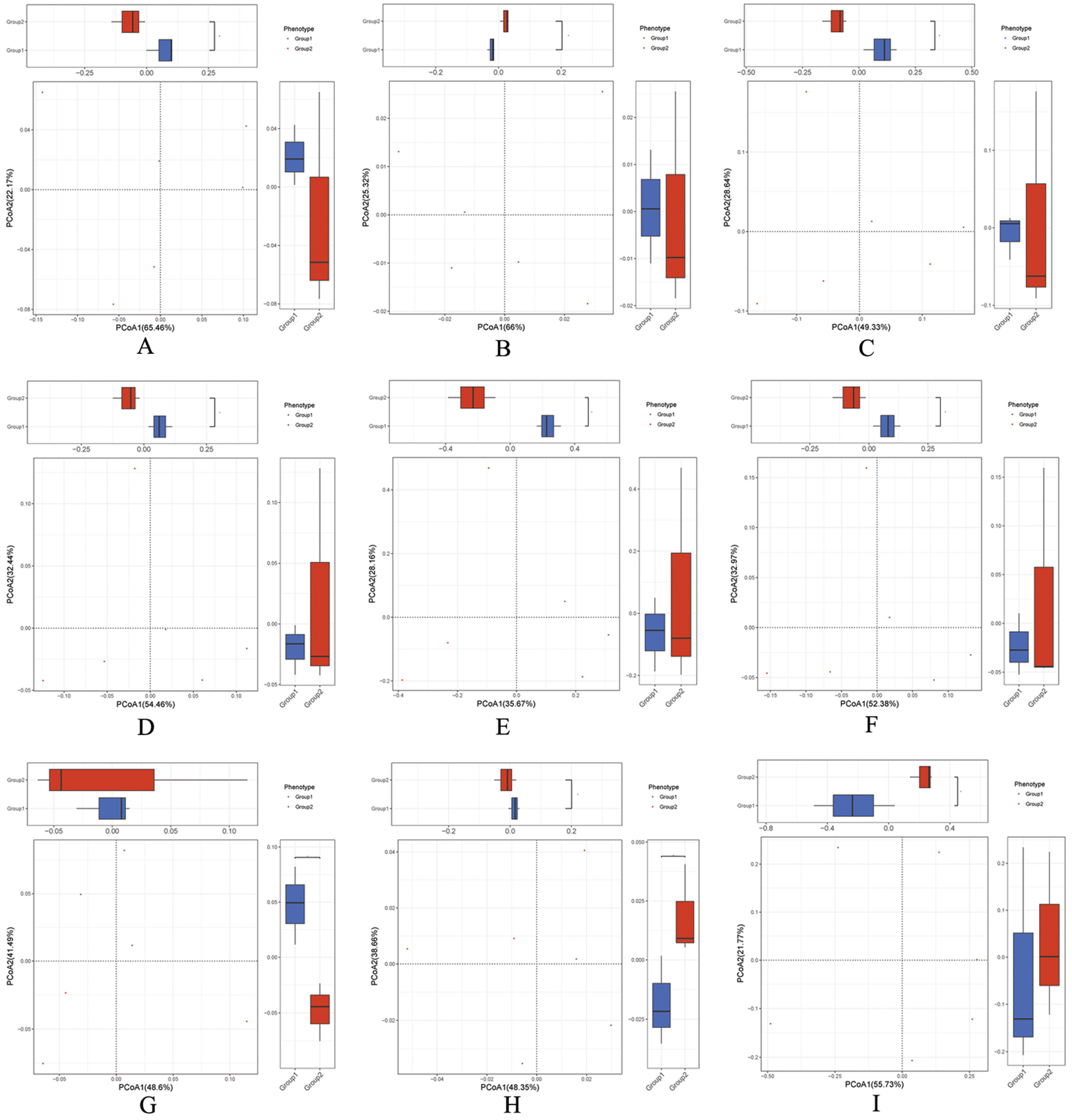
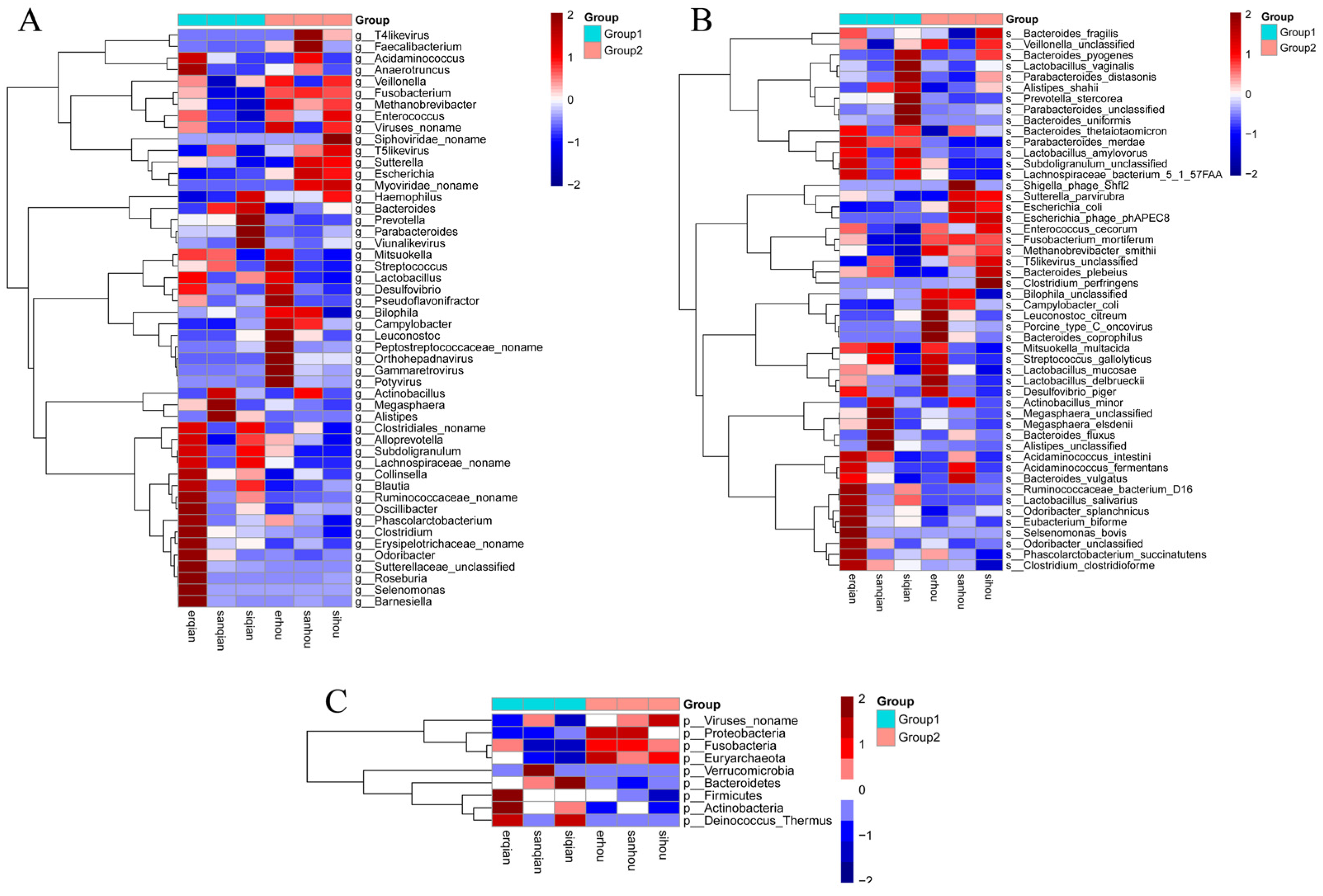
2.2. Overall Structural Modulation of Gut Microbiome Following Treatment with BBA
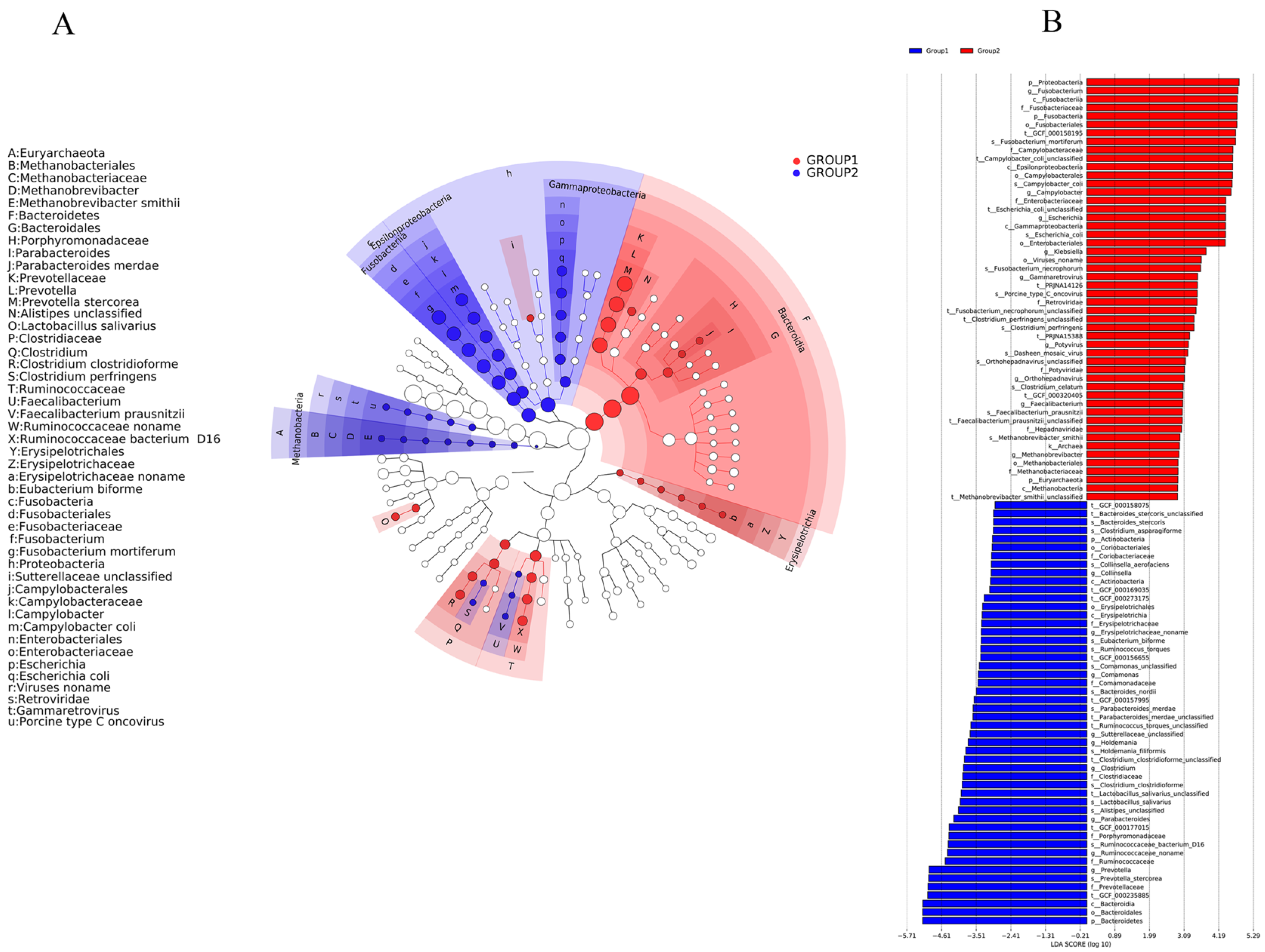
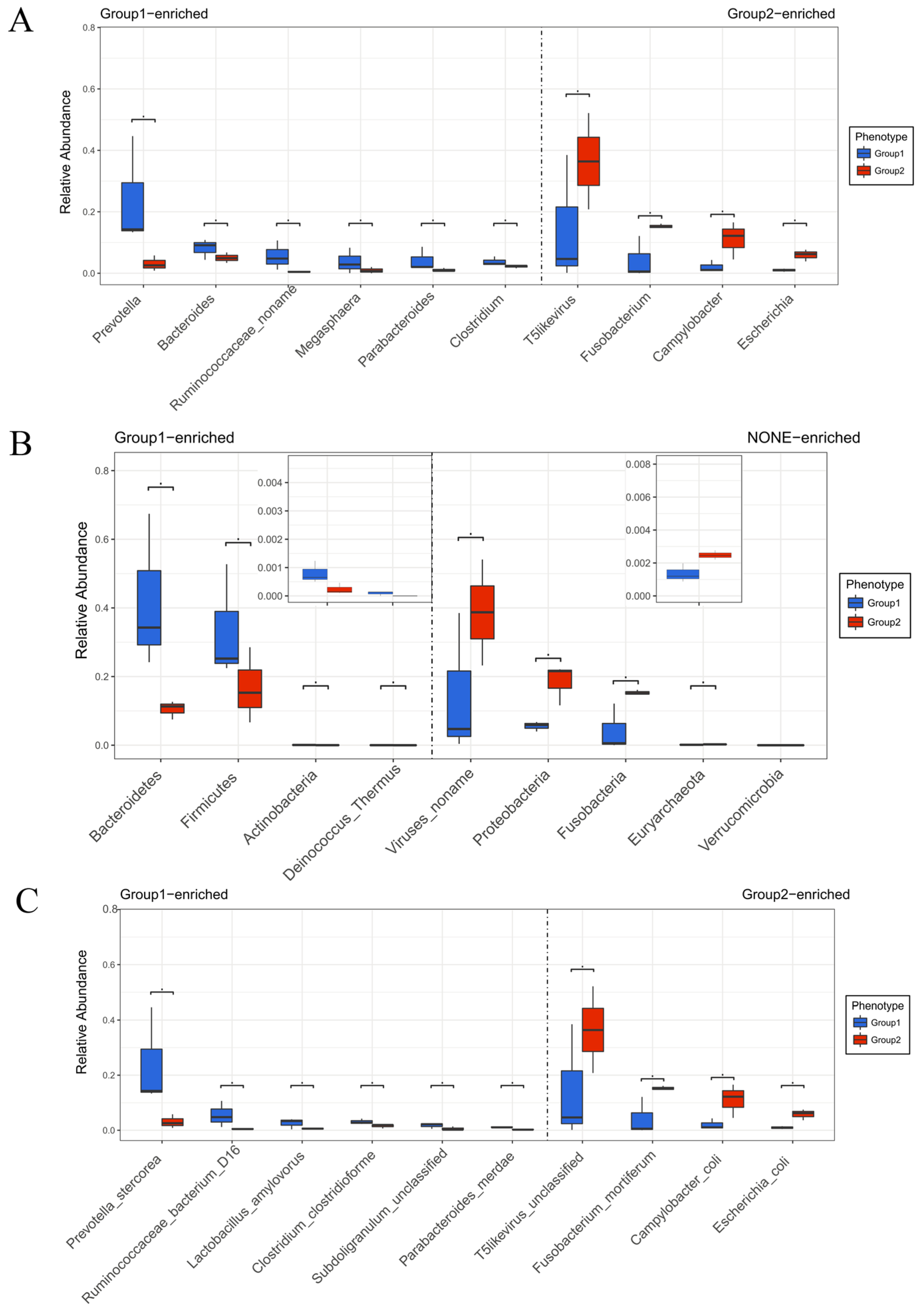
2.3. The Effect of BBA on Modulation of Important Phylotypes of Gut Microbiome
2.4. The Effect of BBA on Modulation of Genes Expression of Gut Microbiome
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Collection of Stool
4.3. DNA Extraction
4.4. DNA Library Construction and Sequencing
4.5. Metagenomic Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Mohamed, R.; John, M.F.; Francis, B.; Ann, L. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar]
- Pu, J.; Chen, D.; Tian, G.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Huang, Z.; Zhu, L.; Luo, J.; Luo, Y.; Yu, B. Protective Effects of Benzoic Acid, Bacillus Coagulans, and Oregano Oil on Intestinal Injury Caused by Enterotoxigenic Escherichia coli in Weaned Piglets. BioMed Res. Int. 2018, 2018, 1829632. [Google Scholar] [CrossRef]
- Farshim, P.; Walton, G.; Chakrabarti, B.; Givens, I.; Saddy, D.; Kitchen, I.; Swann, J.S.; Bailey, A. Maternal Weaning Modulates Emotional Behavior and Regulates the Gut-Brain Axis. Sci. Rep. 2016, 6, 21958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Beaudry, F.; Theriault, W.; Bergeron, N.; Beauchamp, G.; Laurent-Lewandowski, S.; Fairbrother, J.M.; Letellier, A. In vivo therapeutic efficacy and pharmacokinetics of colistin sulfate in an experimental model of enterotoxigenic Escherichia coli infection in weaned pigs. Vet. Res. 2016, 47, 58. [Google Scholar] [CrossRef] [PubMed]
- Xiaohua, L.; Wei, H.; Yanxiang, G. Antibiotics use and drug resistance analysis of neurology patients’ Infection. Pak. J. Pharm. Sci. 2018, 31, 2235–2240. [Google Scholar]
- Ortelli, D.; Sporri, A.S.; Edder, P. Veterinary Drug Residue in Food of Animal Origin in Switzerland: A Health Concern? Chimia 2018, 72, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Kan, L.; Wu, L.; Zhu, Y.; Wang, Q. Effect of baicalin on renal function in patients with diabetic nephropathy and its therapeutic mechanism. Exp. Ther. Med. 2019, 17, 2071–2076. [Google Scholar] [CrossRef]
- Wu, S.C.; Chu, X.L.; Su, J.Q.; Cui, Z.Q.; Zhang, L.Y.; Yu, A.J.; Wu, Z.M.; Cai, M.L.; Li, H.X.; Zhang, Z.J. Baicalin protects mice against Salmonella typhimurium infection via the modulation of both bacterial virulence and host response. Phytomedicine 2018, 48, 21–31. [Google Scholar] [CrossRef]
- Fu, S.; Liu, H.; Chen, X.; Qiu, Y.; Ye, C.; Liu, Y.; Wu, Z.; Guo, L.; Hou, Y.; Hu, C.A. Baicalin Inhibits Haemophilus Parasuis-Induced High-Mobility Group Box 1 Release during Inflammation. Int. J. Mol. Sci. 2018, 19, 1307. [Google Scholar] [CrossRef]
- Zhang, Q.; Sun, J.; Wang, Y.; He, W.; Wang, L.; Zheng, Y.; Wu, J.; Zhang, Y.; Jiang, X. Antimycobacterial and Anti-inflammatory Mechanisms of Baicalin via Induced Autophagy in Macrophages Infected with Mycobacterium tuberculosis. Front. Microbiol. 2017, 8, 2142. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhang, Y.; Chen, Y.; Niu, X.; Zhang, Y.; Yang, C.; Wang, Q.; Li, X.; Deng, X. Baicalin inhibits the lethality of Shiga-like toxin 2 in mice. Antimicrob. Agents Chemother. 2015, 59, 7054–7060. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Qi, Z.; Liu, Y.; He, W.; Yang, C.; Wang, Q.; Dong, J.; Deng, X. Baicalin Protects Mice from Lethal Infection by Enterohemorrhagic Escherichia coli. Front. Microbiol. 2017, 8, 395. [Google Scholar] [CrossRef]
- Zhao, Q.Y.; Yuan, F.W.; Liang, T.; Liang, X.C.; Luo, Y.R.; Jiang, M.; Qing, S.Z.; Zhang, W.M. Baicalin inhibits Escherichia coli isolates in bovine mastitic milk and reduces antimicrobial resistance. J. Dairy Sci. 2018, 101, 2415–2422. [Google Scholar] [CrossRef]
- Shahriari, R.; Salari, S.; Shahriari, S. In vitrostudy of concentration-effect and time-course pattern of white alum on Escherichia coli O157:H7 growth. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 311–318. [Google Scholar] [CrossRef]
- Yan, Y.B.; Yang, D.Y.; Li, X.Y.; Wang, H. Determination of baicalin in Huangqinsulv capsule by HPLC. Word Phytomed. 2007, 22, 206–207. [Google Scholar]
- Robino, P.; Ferrocino, I.; Rossi, G.; Dogliero, A.; Alessandria, V.; Grosso, L.; Galosi, L.; Tramuta, C.; Cocolin, L.; Nebbia, P. Changes in gut bacterial communities in canaries infected by Macrorhabdus ornithogaster. Avian Pathol. 2018. [Google Scholar] [CrossRef]
- Sun, X.; Jia, Z. Microbiome modulates intestinal homeostasis against inflammatory diseases. Vet. Immunol. Immunopathol. 2018, 205, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Masuda, T.; Kurosawa, D.; Tsukahara, T. Dietary administration of probiotics to sows and/or their neonates improves the reproductive performance, incidence of post-weaning diarrhea and histopathological parameters in the intestine of weaned piglets. Anim. Sci. J. 2016, 87, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Zundler, S.; Neurath, M.F. Pathogenic T cell subsets in allergic and chronic inflammatory bowel disorders. Immunol. Rev. 2017, 278, 263–276. [Google Scholar] [CrossRef]
- Feng, W.; Wang, H.; Zhang, P.; Gao, C.; Tao, J.; Ge, Z.; Zhu, D.; Bi, Y. Modulation of gut microbiota contributes to curcumin-mediated attenuation of hepatic steatosis in rats. Biochim. Biophys. Acta 2017, 1861, 1801–1812. [Google Scholar] [CrossRef]
- Abia, A.L.K.; Alisoltani, A.; Ubomba-Jaswa, E.; Dippenaar, M.A. Microbial life beyond the grave: 16S rRNA gene-based metagenomic analysis of bacteria diversity and their functional profiles in cemetery environments. Sci. Total Environ. 2019, 655, 831–841. [Google Scholar] [CrossRef]
- Wang, Y.; Fei, Y.; Liu, L.; Xiao, Y.; Pang, Y.; Kang, J.; Wang, Z. Polygonatum odoratum Polysaccharides Modulate Gut Microbiota and Mitigate Experimentally Induced Obesity in Rats. Int. J. Mol. Sci. 2018, 19, 3587. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, C.; Lv, W.; Chao, L.; Li, Z.; Shi, D.; Guo, S. Structural Modulation of Gut Microbiota during Alleviation of Suckling Piglets Diarrhoea with Herbal Formula. Evid. Based Complement. Altern. Med. 2017, 2017, 8358151. [Google Scholar] [CrossRef]
- Yan, X.; Yan, J.; Huang, K.; Pan, T.; Xu, Z.; Lu, H. Protective effect of baicalin on the small intestine in rats with food allergy. Life Sci. 2017, 191, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Pei, T.; Huang, C.; Chen, X.; Bai, Y.; Xue, J.; Wu, Z.; Mu, J.; Li, Y.; Wang, Y. A novel systems pharmacology platform to dissect action mechanisms of traditional Chinese medicines for bovine viral diarrhea disease. Eur. J. Pharm. Sci. 2016, 94, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.S.; Liang, X.; Wei, X.H.; Chen, F.L.; Tang, Q.F.; Tan, X.M. Comparative metabolism of the eight main bioactive ingredients of gegen qinlian decoction by the intestinal flora of diarrhoeal and healthy piglets. Biomed. Chromatogr. 2018. [Google Scholar] [CrossRef]
- Ziese, A.L.; Suchodolski, J.S.; Hartmann, K.; Busch, K.; Anderson, A.; Sarwar, F.; Sindern, N.; Unterer, S. Effect of probiotic treatment on the clinical course, intestinal microbiome, and toxigenic Clostridium perfringens in dogs with acute hemorrhagic diarrhea. PLoS ONE 2018, 13, e0204691. [Google Scholar] [CrossRef]
- Paganini, D.; Zimmermann, M.B. The effects of iron fortification and supplementation on the gut microbiome and diarrhea in infants and children: A review. Am. J. Clin. Nutr. 2017, 106, 1688s–1693s. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Han, X.; Zhan, J.; You, Y.; Huang, W. Vanillin Alleviates High Fat Diet-Induced Obesity and Improves the Gut Microbiota Composition. Front. Microbiol. 2018, 9, 2733. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Tang, R.; Yang, S.; Lu, Y.; Luo, J.; Liu, Z. Rutin and Its Combination with Inulin Attenuate Gut Dysbiosis, the Inflammatory Status and Endoplasmic Reticulum Stress in Paneth Cells of Obese Mice Induced by High-Fat Diet. Front. Microbiol. 2018, 9, 2651. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Chung, H.J.; Lee, I.A.; D’Souza, R.; Kim, H.J.; Hong, S.T. Elucidation of the anti-hyperammonemic mechanism of Lactobacillus amylovorus JBD401 by comparative genomic analysis. BMC Genom. 2018, 19, 292. [Google Scholar] [CrossRef]
- Sunmola, A.A.; Ogbole, O.O.; Faleye, T.O.C.; Adetoye, A.; Adeniji, J.A.; Ayeni, F.A. Antiviral potentials of Lactobacillus plantarum, Lactobacillus amylovorus, and Enterococcus hirae against selected Enterovirus. Folia Microbiol. 2019, 64, 257–264. [Google Scholar] [CrossRef]
- Hynonen, U.; Kant, R.; Lahteinen, T.; Pietila, T.E.; Beganovic, J.; Smidt, H.; Uroic, K.; Avall-Jaaskelainen, S.; Palva, A. Functional characterization of probiotic surface layer protein-carrying Lactobacillus amylovorus strains. BMC Microbiol. 2014, 14, 199. [Google Scholar] [CrossRef]
- Sayan, H.; Assavacheep, P.; Angkanaporn, K.; Assavacheep, A. Effect of Lactobacillus salivarius on growth performance, diarrhea incidence, fecal bacterial population and intestinal morphology of suckling pigs challenged with F4+ enterotoxigenic Escherichia coli. Asian Austr. J. Anim. Sci. 2018, 31, 1308–1314. [Google Scholar] [CrossRef]
- Wasfi, R.; Abd El-Rahman, O.A.; Zafer, M.M.; Ashour, H.M. Probiotic Lactobacillus sp. inhibit growth, biofilm formation and gene expression of caries-inducing Streptococcus mutans. J. Cell. Mol. Med. 2018, 22, 1972–1983. [Google Scholar] [CrossRef]
- Scanlan, P.D. Bacteria-Bacteriophage Coevolution in the Human Gut: Implications for Microbial Diversity and Functionality. Trends Microbiol. 2017, 25, 614–623. [Google Scholar] [CrossRef]
- Ogilvie, L.A.; Caplin, J.; Dedi, C.; Diston, D.; Cheek, E.; Bowler, L.; Taylor, H.; Ebdon, J.; Jones, B.V. Comparative (meta)genomic analysis and ecological profiling of human gut-specific bacteriophage phiB124-14. PLoS ONE 2012, 7, e35053. [Google Scholar] [CrossRef] [PubMed]
- Bakhshinejad, B.; Ghiasvand, S. Bacteriophages in the human gut: Our fellow travelers throughout life and potential biomarkers of heath or disease. Virus Res. 2017, 240, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Manrique, P.; Dills, M.; Young, M.J. The Human Gut Phage Community and Its Implications for Health and Disease. Viruses 2017. [Google Scholar] [CrossRef]
- Manrique, P.; Bolduc, B.; Walk, S.T.; Van der Oost, J.; De Vos, W.M.; Young, M.J. Healthy human gut phageome. Proc. Natl. Acad. Sci. USA 2016, 113, 10400–10405. [Google Scholar] [CrossRef] [Green Version]
- Locher, K.P. Mechanistic diversity in ATP-binding cassette (ABC) transporters. Nat. Struct. Mol. Biol. 2016, 23, 487–493. [Google Scholar] [CrossRef]
- Qi, P.F.; Zhang, Y.Z.; Liu, C.H.; Zhu, J.; Chen, Q.; Guo, Z.R.; Wang, Y.; Xu, B.J.; Zheng, T.; Jiang, Y.F.; et al. Fusarium graminearum ATP-Binding Cassette Transporter Gene FgABCC9 Is Required for Its Transportation of Salicylic Acid, Fungicide Resistance, Mycelial Growth and Pathogenicity towards Wheat. Int. J. Mol. Sci. 2018, 19, 2351. [Google Scholar] [CrossRef]
- Brayboy, L.M.; Knapik, L.O.; Long, S.; Westrick, M.; Wessel, G.M. Ovarian hormones modulate multidrug resistance transporters in the ovary. Contracept. Reprod. Med. 2018, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Domenichini, A.; Adamska, A.; Falasca, M. ABC transporters as cancer drivers: Potential functions in cancer development. Biochim. Biophys. acta 2019, 1863, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Petryszyn, P.W.; Wiela-Hojenska, A. The importance of the polymorphisms of the ABCB1 gene in disease susceptibility, behavior and response to treatment in inflammatory bowel disease: A literature review. Adv. Clin. Exp. Med. 2018, 27, 1459–1463. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.P.; He, Q.Q.; Ouyang, H.M.; Peng, H.S.; Wang, Q.; Li, J.; Lv, X.F.; Zheng, Y.N.; Li, S.C.; Liu, H.L.; Yin, A.H. Human Gut Microbiota Associated with Obesity in Chinese Children and Adolescents. BioMed Res. Int. 2017, 2017, 7585989. [Google Scholar] [CrossRef] [PubMed]
- Saier, M.H. The Bacterial Phosphotransferase System: New Frontiers 50 Years after Its Discovery. J. Mol. Microbiol. Biotechnol. 2015, 25, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Perez, W.A.; Yepes-Perez, A.F.; Martinez-Pabon, M.C. Molecular docking and in silico studies of the physicochemical properties of potential inhibitors for the phosphotransferase system of Streptococcus mutans. Arch. Oral Biol. 2019, 98, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Ding, M.; Yang, C.H.; Fan, H.; Chai, Y.; Li, Y. The phosphotransferase system gene ptsH plays an important role in MnSOD production, biofilm formation, swarming motility, and root colonization in Bacillus cereus 905. Res. Microbiol. 2018. [Google Scholar] [CrossRef]
- Wang, L.; Huang, L.; Su, Y.; Qin, Y.; Kong, W.; Ma, Y.; Xu, X.; Lin, M.; Zheng, J.; Yan, Q. Involvement of the flagellar assembly pathway in Vibrio alginolyticus adhesion under environmental stresses. Front. Cell. Infect. Microbiol. 2015, 5, 59. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, M. Strategies to Block Bacterial Pathogenesis by Interference with Motility and Chemotaxis. Curr. Top. Microbiol. Immunol. 2016, 398, 185–205. [Google Scholar]
- Matilla, M.A.; Krell, T. The effect of bacterial chemotaxis on host infection and pathogenicity. FEMS Microbiol. Rev. 2018, 42. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, X.; Dou, Y.; Wang, S.; Tian, M.; Qi, J.; Li, T.; Ding, C.; Wu, Y.; Yu, S. Riemerella anatipestifer M949_1360 Gene Functions on the Lipopolysaccharide Biosynthesis and Bacterial Virulence. PLoS ONE 2016, 11, e0160708. [Google Scholar] [CrossRef]
- Karasartova, D.; Gureser, A.S.; Ruh, E.; Turegun-Atasoy, B.; Calgin, M.K.; Tasci, L.; Taylan-Ozkan, A. An alternative DNA extraction method for detection of Blastocystis spp. in human fecal samples. Exp. Parasitol. 2018, 186, 36–41. [Google Scholar] [CrossRef]
- Forouzan, E.; Shariati, P.; Mousavi Maleki, M.S.; Karkhane, A.A.; Yakhchali, B. Practical evaluation of 11 de novo assemblers in metagenome assembly. J. Microbiol. Methods. 2018, 151, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Ismail, W.M.; Ye, Y.; Tang, H. Gene finding in metatranscriptomic sequences. BMC Bioinform. 2014, 15, S8. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jiang, G.F.; Dong, S.Y. A head transcriptome provides insights into odorant binding proteins of the bamboo grasshopper. Genes Genom. 2018, 40, 991–1000. [Google Scholar] [CrossRef]
- Wang, Q.; Li, F.; Liang, B.; Liang, Y.; Chen, S.; Mo, X.; Ju, Y.; Zhao, H.; Jia, H.; Spector, T.D.; et al. A metagenome-wide association study of gut microbiota in asthma in UK adults. BMC Microbiol. 2018, 18, 114. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Forslund, K.; Cook, H.; Heller, D.; Walter, M.C.; Rattei, T.; Mende, D.R.; Sunagawa, S.; Kuhn, M.; et al. eggNOG 4.5: A hierarchical orthology framework with improved functional annotations for eukaryotic, prokaryotic and viral sequences. Nucleic Acids Res. 2016, 44, D286–D293. [Google Scholar] [CrossRef]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. MetaPhlAn2 for enhanced metagenomic taxonomic profiling. Nat. Methods 2015, 12, 902–903. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, F.; Crippa, G.; Garbelli, C.; Griesshaber, E. Microstructural data of six recent brachiopod species: SEM, EBSD, morphometric and statistical analyses. Data Brief 2018, 18, 300–318. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, 60. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Raw Reads | Clean Reads | Clean Ratio (%) | Mapped Ratio (%) |
|---|---|---|---|---|
| Treated with BAA | 89,537,684 ± 5,405,796.2 | 87,247,590.3 ± 5,144,406.2 | 94.9 ± 0.2 | 99.1 ± 0.1 |
| Untreated | 103,588,655.3 ± 13,288,842.4 | 100,319,056.3 ± 12,734,141.2 | 92.6 ± 0.2 | 99.0 ± 0.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.; Zhuang, F.; Guo, L.; Qiu, Y.; Xiong, J.; Ye, C.; Liu, Y.; Wu, Z.; Hou, Y.; Hu, C.-A.A. Effect of Baicalin-Aluminum Complexes on Fecal Microbiome in Piglets. Int. J. Mol. Sci. 2019, 20, 2390. https://doi.org/10.3390/ijms20102390
Fu S, Zhuang F, Guo L, Qiu Y, Xiong J, Ye C, Liu Y, Wu Z, Hou Y, Hu C-AA. Effect of Baicalin-Aluminum Complexes on Fecal Microbiome in Piglets. International Journal of Molecular Sciences. 2019; 20(10):2390. https://doi.org/10.3390/ijms20102390
Chicago/Turabian StyleFu, Shulin, Feng Zhuang, Ling Guo, Yinsheng Qiu, Jianglin Xiong, Chun Ye, Yu Liu, Zhongyuan Wu, Yongqing Hou, and Chien-An Andy Hu. 2019. "Effect of Baicalin-Aluminum Complexes on Fecal Microbiome in Piglets" International Journal of Molecular Sciences 20, no. 10: 2390. https://doi.org/10.3390/ijms20102390