Circadian Rhythm Abnormalities in Parkinson’s Disease from Humans to Flies and Back


1
Department of Biology, University of Padova, 35131 Padova, Italy
2
Cognitive Neuroscience Center, University of Padova, 35100 Padova, Italy
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(12), 3911; https://doi.org/10.3390/ijms19123911
Submission received: 18 October 2018
/
Revised: 23 November 2018
/
Accepted: 30 November 2018
/
Published: 6 December 2018
(This article belongs to the Special Issue Genetic and Molecular Bases of the Circadian Rhythms)
Abstract
:Clinical and research studies have suggested a link between Parkinson’s disease (PD) and alterations in the circadian clock. Drosophila melanogaster may represent a useful model to study the relationship between the circadian clock and PD. Apart from the conservation of many genes, cellular mechanisms, signaling pathways, and neuronal processes, Drosophila shows an organized central nervous system and well-characterized complex behavioral phenotypes. In fact, Drosophila has been successfully used in the dissection of the circadian system and as a model for neurodegenerative disorders, including PD. Here, we describe the fly circadian and dopaminergic systems and report recent studies which indicate the presence of circadian abnormalities in some fly PD genetic models. We discuss the use of Drosophila to investigate whether, in adults, the disruption of the circadian system might be causative of brain neurodegeneration. We also consider approaches using Drosophila, which might provide new information on the link between PD and the circadian clock. As a corollary, since PD develops its symptomatology over a large part of the organism’s lifespan and given the relatively short lifespan of fruit flies, we suggest that genetic models of PD could be used to perform lifelong screens for drug-modulators of general and/or circadian-related PD traits.
1. Introduction
Parkinson’s disease (PD) is a chronic, progressive neurodegenerative disorder, which is mainly characterized by motor dysfunction (i.e., bradykinesia, postural instability, muscular rigidity, and resting tremor). Motor symptoms arise from the preferential degeneration of dopaminergic neurons in the Substantia nigra pars compacta (SNpc), which represents one hallmark of the disease. The other specific feature of PD is the presence in the surviving neurons of cytoplasmic inclusions, referred to as Lewy bodies, which are mainly formed by ubiquitin and α-synuclein (α-syn) [1].
Besides motor dysfunctions, non-motor symptoms, such as changes in sleep, mood, cognition, and autonomic as well as visual functions, have often been observed during PD progression, and their frequency seems to be related to disease severity [2]. At least some of the non-motor symptoms are thought to be linked to neuronal alterations, as well as to decreased dopamine (DA) levels in brain areas different from the SNpc [3,4,5,6,7].
PD is mainly a sporadic disorder, the pathogenesis of which has not been fully elucidated. However, ~10% of all cases have a genetic origin, and currently at least 23 disease-segregating loci have been identified [8]. The most studied so far are the genes encoding the proteins α-syn, Parkin, PINK1, DJ-1, and LRRK2, and different animal models for these genetic forms have been generated. Although not fully reproducing all the clinical and pathological phenotypes associated with PD, these genetic models have contributed to a better definition of the cellular pathways involved in pathogenesis. These include mitochondrial dysfunction, impaired lysosomal and proteasomal degradation, deficiencies in synaptic transmission, and vesicular recycling [8].
Several lines of evidence have suggested a link between PD pathology and alterations in the circadian clock system [3,4,7], the endogenous ~24 h clock which controls the daily timing of physiological and behavioral processes in organisms. However, since PD is a complex disease, the physiological bases of the circadian alterations associated to this pathology have not been fully clarified yet. In the effort of elucidating such aspects, the possibility of performing basic and translational research studies using multiple model organisms represents a valuable approach.
In this paper, after a brief description of the circadian clock system in mammals, we will discuss a number of studies performed in humans and mammalian models, supporting the association between circadian abnormalities and PD. We will also highlight some aspects emerging from such studies which still remain elusive. Subsequently, we will focus on the insect Drosophila melanogaster as a possible additional model to address at least some of the open questions in investigations concerning the link between PD and the circadian clock.
2. Circadian Abnormalities in PD Pathology
In humans, as in other mammals, the circadian system consists of a hierarchical network of cellular clocks, localized in both the brain and peripheral organs, which are integrated at the level of the organism. The suprachiasmatic nucleus (SCN) of the hypothalamus is the central circadian clock. Light is the main environmental synchronizing stimulus of the circadian clock. The SCN receives light stimuli from the retina through the retinohypothalamic tract, and entrains the endogenous clock to the daily external environmental variations, sending signals to the peripheral clocks, and coordinating their rhythm and phase. This, in turn, determines that clock-controlled phenotypes, such as sleep/wake cycles, body temperature, hormone secretion, and metabolism are synchronized in phase with the environmental daily light-dark (LD) cycle [9].
PD patients frequently show modifications in several physiological aspects, which are fully or partly under the control of the circadian system.
2.1. Circadian Symptoms in PD Patients
A circadian symptom which has been detected in individuals affected by PD consists in alterations at the level of the daily rest/activity rhythm [10,11,12]. In healthy conditions, daily motor activity shows a cycling pattern, with the lowest levels during the night, and a progressive increase during the day. In PD patients, reduced amplitude in the rest/activity rhythm was recorded, with decrements in both diurnal activity and nocturnal rest [10,11,12]. In addition, it has been reported that from 65 to 95% of PD individuals suffer from sleep disturbances [13], which are now recognized as some of the major contributors to the impairment of life quality in patients [2,14,15]. Sleep dysfunctions include sleep fragmentation, excessive daytime sleepiness, and REM sleep behavior disorder (RBD), a pathology characterized by the loss of normal atonia during the REM stage of sleep. Although the causes of sleep impairments in PD are multifactorial, it is however interesting to note that RBD is considered a sleep stage under strong circadian influence [3].
PD may also be associated with several visual problems, such as disturbances in visual acuity, contrast sensitivity, and color vision [16]. These deficits are at least partially due to retinal DA deficiency [16], and some of them are somewhat reversible with levodopa treatment [17,18]. Interestingly, visuo-perceptual deficits have been reported in patients with RBD, with impairments in color vision and visuospatial construction supporting circadian fluctuations in visual perception. In addition, a clinical study found that contrast sensitivity in PD patients showed diurnal fluctuation that is otherwise absent in healthy people [19].
Several studies indicated that many clock-controlled parameters, such as core body temperature, blood pressure, and the daily cycling secretion of melatonin (involved in the regulation of the sleep/wake cycle) appeared to be frequently affected in PD patients [4,20,21,22,23,24,25]. Other analyses indicated altered expression levels of the cardinal human (h) clock gene, hBmal1 in blood samples from PD patients compared to appropriate controls [23,26] and a case-control study associated specific single nucleotide polymorphisms in hBmal1 and hPer1 circadian clock genes to an increased risk of PD in a Chinese population [27].
Finally, it is worth mentioning that different studies reported that PD patients experienced the beneficial effects of bright light therapy (BLT), which ameliorated both non-motor and motor symptoms [3]. BLT has been successfully used to reduce depressive symptoms in several types of mood disorders and is thought to act through modulation of the circadian clock [28].
Taken together, these data support the association between circadian dysfunctions and PD. However, the etiology of the circadian symptoms in PD patients is not well understood. The difficulty is due to several reasons, including the fact that the pathology is extremely heterogenous, with a large and variable spectrum of symptoms [29]; the therapy used to alleviate PD can, per se, cause a variation in circadian parameters, such as the melatonin cycling phase in levodopa-treated patients [25]; some non-motor symptoms, such as sleep disturbances, and their worsening during the progression of the pathology, appear to negatively impact the circadian system of PD patients, leading to a vicious circle [30]. Studies in mammalian models have, however, generated informative data.
2.2. Circadian Dysfunctions in Mammalian PD Models
Both circadian and sleep phenotypes have been analyzed in several mammalian PD models, produced via genetic alterations or toxin-based treatments [3]. Here, we will highlight defects mainly associated with the circadian clock (Table 1).
Variable and age-dependent abnormalities in circadian behavioral phenotypes were detected in a mouse model overexpressing the human α-syn in the whole brain (ASO) [31], as well as in mice showing a progressive degeneration of dopaminergic neurons, due to a specific deletion in the mitochondrial transcription factor A gene in these neurons (MitoPark) [32] (Table 1). In addition, ASO mice displayed a reduced neuronal daytime firing rate in the SCN, although the cycling expression of the mouse (m) circadian factor mPER2 was not affected [31]. Circadian alterations were also detected in PD models induced by stereotaxic injection with toxins such as 6-hydroxydopamine (6-OHDA) [43] or 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [44], both determining dopaminergic neuronal death. 6-OHDA treatments in rodents have been found to deregulate circadian behavioral and physiological outputs, as well as the expression of clock genes in different brain regions [38,39,40,41]. Furthermore, a recent study reported that levodopa administration further modified hormone secretion levels and the expression profiles of some clock genes in specific brain regions in 6-OHDA treated-rats, corroborating the notion that the pro-drug may contribute to circadian alterations seen in PD patients [42]. Contrasting data relating to the effects of MPTP treatment on mouse circadian phenotypes were obtained [33,34,35] (Table 1). However, it is interesting to note that an environmental circadian disruption (induced by a long-term exposure to a 20:4 LD cycle) exacerbated the PD-related motor deficits in this model [45]. In addition, circadian abnormalities were observed in MPTP-treated non-human primates, which showed arrhythmic behavior in constant darkness conditions (DD, when the endogenous clock free runs), indicating that DA depletion caused the loss of circadian locomotor activity, at least in this model [30]. Since MPTP treatment did not seem to influence the SCN circadian functions, the authors suggested that the behavioral defects lay downstream of the master clock [30].
Collectively, these data suggest that the circadian dysfunctions detected in PD can result from a misregulation of the complex interactions between the dopaminergic and circadian systems. Indeed, several lines of evidence indicate DA as one of the modulators of the circadian system, playing a fundamental role in the maintenance of proper rhythmicity [7,46]. DA and dopaminergic transmission seem to have relevant functions at different levels of the circadian system [3,46] as they are, for example, implicated in the modulation of the light input signals from the retina to the SCN [47], but they are also considered as output targets of the timing information from the SCN to other brain regions [3,46]. However, the cellular and molecular processes at the basis of circadian dysfunctions in PD are still unclear. In addition, further studies are required to clarify whether the circadian abnormalities are only a consequence of PD or might also represent a causal/enhancement factor during the progression of the pathology.
Given the above considerations, studies based on the use of relatively simple and well-characterized model organisms, such as D. melanogaster, have the potential of providing relevant information to explore such aspects.
3. Drosophila melanogaster as a Model Organism to Study the Relationship between the Circadian Clock and PD
D. melanogaster is a widely accepted model organism which has been used extensively to study a range of biological and physiological phenomena, as well as several types of human pathology. Despite the evolutionary distance, many genes, cellular mechanisms, signaling pathways, as well as neuronal processes are conserved between insects and mammals. Moreover, Drosophila shows an organized central nervous system and well-characterized complex phenotypes, such as circadian behaviors, locomotion, sleep, memory, and learning [48]. In addition, this insect provides the advantage of a relatively easy implementation of advanced genetic techniques, which are generally more challenging to accomplish in mammals [49]. In the past years, Drosophila has been successfully used in the study of the circadian system and as a model for neurodegenerative disorders, including PD [50,51,52]. In the following sections we will describe the fly circadian and dopaminergic systems and we will report recent studies which indicate the presence of circadian abnormalities in some fly PD genetic models. We will also discuss the use of Drosophila to investigate whether the disruption of the circadian system might be causative of neurodegeneration within adult brains. Finally, we will examine further analyses and approaches which might be useful in the investigation of the link between PD and the circadian clock using Drosophila.
3.1. The Drosophila Circadian Clock
The Drosophila circadian clock controls the timing of several behavioral and physiological processes, such as daily locomotor activity, eclosion, feeding behavior, temperature preferences, memory, and social behaviors [53,54,55,56]. Here, we will focus on the daily locomotor activity, since this represents the circadian behavioral output most studied in Drosophila PD models so far. In laboratory 12:12 LD cycles, D. melanogaster shows locomotor activity with a bimodal profile, characterized by a peak in the morning and a second peak in the evening. In wild-type flies, locomotor activity starts to increase slightly before the lights-on and lights-off transitions, anticipating the LD variation (Figure 1A). In DD conditions, the locomotor activity remains rhythmic, with a periodicity of ~24 h, even though, in the latter condition, the locomotor profiles show slight changes with respect to the LD profiles (Figure 1B).
The circadian locomotor activity is driven by a master clock, located in the brain. This is formed by ~150 neurons, organized into six groups per brain hemisphere and classified as four large ventral lateral neurons (l-LNvs), five small ventral lateral neurons (s-LNvs), six dorsal lateral neurons (LNds), three lateral posterior neurons (LPNs), and dorsal neurons, subdivided into three groups (DNs 1, 2, and 3) [57] (Figure 1C). Initial analyses suggested a primary role of the four s-LNvs in controlling morning activity and of the fifth s-LNv, the LNds, and the DNs in regulating evening activity [58,59]. However, recent data indicate that the whole network of circadian clock neurons contributes to the generation of behavioral rhythmicity in both LD and DD conditions [60,61].
All these neurons express the fundamental clock components required to drive circadian rhythmicity at the molecular level. In a simplified model, the core of the molecular circadian clock is formed by two main interlocking autoregulatory transcriptional/translational feedback loops (TTLs) [62]. The first TTL includes the transcription factors dCLOCK (dCLK) and dCYCLE (dCYC) which form a heterodimer (dCLK/dCYC) and promote the transcription of the dperiod (dper) and dtimeless (dtim) genes (Figure 1D). dPER and dTIM proteins are then regulated by different kinases and phosphatases at a post-translational level to modulate the timing of their rhythmic functions, their nuclear translocation, stability, and their activity as inhibitors of the dCLK/dCYC transcriptional regulators. Among the kinases, dDoubletime (dDBT) targets dPER, while dShaggy is involved in the phosphorylation of both dTIM and dPER. The dCLK/dCYC dimer also represents the positive regulator of a second TTL, which modulates the daily expression of dClk. This TTL is negatively and positively regulated by dVRILLE (dVRI) and dPar Domain Protein 1 (dPDP1), respectively, which probably compete to bind the same DNA elements in the promoter region of dClk [62] (Figure 1D). In mammals, the organization into TTLs of the molecular clockwork is essentially conserved, in spite of the replacement of some elements in both the first (mCryptochromes (mCRYs) instead of dTIM) and second [mRar-related orphan receptor (mROR) and mNuclear receptor subfamily 1, Group D (mREV-ERB) instead of dVRI and dPDP1] TTLs. In addition, mammals show an intrinsic redundancy, as they are characterized by the presence of different paralogs of the same molecular clock element, including three mPer, two mCry, one mCLK, and one mNeuronal PAS domain-containing protein 2 (mNpas2) gene.
In flies, clock neurons show several differences in protein/neuropeptide expression profiles, properties, and activities. This variability has been associated with the roles of the different neurons in driving and fine-tuning the circadian rhythmicity at an organismal level [53,55,63,64]. A first example is linked to the capacity to perceive clock-resetting light signals. As in mammals, light represents the main cue which is responsible for the synchronization of the Drosophila circadian clock with the 24 h LD cycles. At the molecular level, light synchronization is mediated by the blue-light photoreceptor, dCRY [65,66]. Light activates dCRY, which interacts with dTIM, promoting its degradation [67,68] (Figure 1D). However, only a subset of clock neurons expresses dCRY [69,70,71], and the circadian clock network perceives light signals also through other pathways, including eye photoreceptors, ocelli, and the Hofbauer Buchner eyelet [72].
Fundamental differences are also found in the synthesis and activity of neuropeptides, such as the peptide Pigment Dispersing Factor (dPDF) [73], expressed in the l-LNvs and in 4 out of the 5 s-LNvs. dPDF acts as an output neurotransmitter, as a synchronizer of the clock neurons and in the transduction of light signals into the circadian network [57]. Other neuropeptides, including the short neuropeptide F [74] and the ion transport peptide [75], which have a role in controlling different features of the evening locomotor behavior, are expressed with variable patterns within the master clock neurons [53,55,63]. Among clock neurons, a heterogeneity in the sensitivity to several types of neurotransmitters, including glutamate, acetyilcholine, GABA, glycine, and biogenic amines [i.e., DA, serotonin, and octopamine (related to mammalian noradrenaline)], was also demonstrated and associated with different aspects of circadian locomotor behavior [53,63,64].
Recent studies focused on the communications between the central clock and downstream brain regions important in the generation of circadian locomotor behavior. In the current model, timekeeping signals from the master clock reach specific neurons of the Pars Intecerebralis (PI) (Figure 1C), which in turn contact other neurons located in the subesophageal brain region. Detailed information about these neuronal circuits are reported elsewhere [53,64,76,77,78].
Interestingly, it has been suggested that aminergic systems (i.e., serotoninergic, dopaminergic, and octopaminergic) also have a fundamental role as output components acting downstream from the master clock in the control of circadian locomotor rhythmicity, both in LD and DD [79]. DA was demonstrated to be important in circadian light sensitivity and entrainment, as well as in driving a robust circadian rhythmicity in free-running conditions [80]. In addition, a cooperation between at least two aminergic systems seems to be required to guarantee circadian locomotor activity both in LD and DD [79]. In fact, flies carrying a loss-of-function mutation in the vesicular monoamine transporter dVMAT, which functionally inactivates all the aminergic systems, showed an aberrant locomotor activity profile in LD and weak rhythmicity in DD [81]. On the contrary, dVmat− flies overexpressing wild-type dVMAT in at least two out of the three systems (in all the combinations) showed a rescue of the mutant circadian phenotype in both LD and DD. Since dVmat− flies did not show evident defects in the circadian molecular oscillator within the master clock neurons, the cooperative action of the aminergic systems was postulated to act downstream of the brain master clock [79].
Finally, it has been suggested that glia-to-neuron communication also controls rhythmic behavior, since the glia modulates dPDF transport and/or release from LNv projections [82], and at this level, ebony, which encodes an enzyme involved in DA and histamine recycling, seems to play a key role as an output gene [83].
3.2. Drosophila Dopaminergic System
Drosophila has a well-characterized dopaminergic system which encompasses ~130 neurons, subdivided into multiple clusters which are symmetrically distributed in the fly brain [84,85,86]. The major cluster is the protocerebral anterior medial (PAM) group, which is composed of ~100 neurons, while all other groups are constituted by fewer cells (~5–10 each). These smaller clusters include the protocerebral anterior lateral (PAL), protocerebral posterior medial (PPM1–4), the protocerebral posterior lateral (PPL1–5), the Thoracic 1 (T1), and the ventral unpaired median (VUM) group (Figure 1C). While PAM, PAL, and T1 neurons are located in the anterior part of the brain, the PPM and PPL clusters are mapped posteriorly. These different groups are variably involved in the control of locomotion and other complex behaviors, including olfaction, memory, learning, courtship, reward, sleep, and arousal by communicating with organized substructures of the Drosophila brain, such as the Mushroom Bodies (MB) and Central Complex (CC) [84,86,87]. Moreover, small cells expressing tyrosine hydroxylase (TH) (the rate-limiting enzyme involved in DA synthesis) are located in the optic lobe at the level of the medulla neuropile [85] (Figure 1C). In addition, pathways and enzymes involved in DA synthesis, release, and signaling are conserved, although the catabolism of DA follows different mechanisms [87]. As in the case of mammals, in Drosophila, DA shows a rhythmic release in the form of a pronounced bimodal daily pattern with a mild increase during the end of the night in adult fly heads. This pattern seems to be influenced by light during the daytime, while it is controlled by the circadian system during the night [88].
DA in Drosophila is not just a neurotransmitter, but is required as a precursor molecule for cuticle pigmentation; therefore, pale− mutant flies lacking a functional TH present unpigmented cuticles, as well as embryonic lethality. For this reason, in order to investigate the functions of DA at the level of the brain, the main model is based on the generation of individuals that specifically preserve DA synthesis in non-neuronal cells. Interestingly, these flies displayed reduced daily activity and tended to sleep more frequently than controls; they also showed a reduced reactivity to external mechanical stimuli [89]. Remarkably, the administration of known PD drugs, such as levodopa and carbidopa, were found to rescue locomotor hypoactivity, providing evidence that with respect to these drugs, the Drosophila blood-brain barrier has the same permeability properties as that of mammals [90].
Although clock neurons do not express DA [91], and it is suggested that dopaminergic neurons do not possess a functional clock [92], different studies indicate the existence of synaptic contacts between dopaminergic and circadian-clock LNvs neurons. In particular, dopaminergic neurons appear to send input signals to l-LNv dendrites, and to receive input stimuli from s-LNv axons [93,94]. These neuronal communications contribute to regulate diurnal wakefulness in the complex interaction involved in the control of sleep and wakefulness in flies [93,94,95].
3.3. Drosophila PD Models and Circadian Dysfunctions
Drosophila is widely employed for studying the genetic forms of PD. Fly models have been generated to analyze the effects of hα-syn overexpression, as well as knockout (KO) and knockdown (KD) alterations of the Drosophila homologs of mammalian Lrrk2, Parkin, PINK1, and DJ-1. Recent studies have examined the circadian phenotypes in hα-syn overexpressing flies, and in dParkin, dPINK1 single mutants [96,97,98]. In the following paragraphs, we will describe the available data on these models, together with those obtained in our laboratories, analyzing circadian locomotor activity in flies with null mutations in the Dj-1 homologs (Table 2).
3.3.1. α-syn
In humans, α-syn is a small, intrinsically disordered 14 kDa protein which is abundantly expressed in neurons. Mutations and multicopy variants of its encoding gene, hSNCA have been associated with highly penetrant forms of PD and dominant inheritance [109]. At the neuronal level, the protein exists in an equilibrium between a highly disordered cytosolic monomer and a folded, membrane-bound form [110]. Although the physiological role is still under debate, its prevalent localization at presynaptic terminals suggests an involvement in neurotransmitter release and vesicle recycling [111]. Nevertheless, under pathological conditions, the protein conformation is thought to shift from disordered monomers to large multimeric species, forming insoluble cytosolic inclusions. Oligomers are pre-fibrillar species that are considered to be the principal culprits of hα-syn toxicity due to their potential to disrupt membrane integrity, and to alter mitochondrial functionality and protein degradation pathways [109]. Although the Drosophila genome does not harbor an SNCA homolog, different fly models expressing wild-type and mutant variants of the human gene have been generated. These models manifest a variety of aspects of human PD, including neurodegeneration, locomotor dysfunctions, and the formation of sub-cellular protein inclusions [99,112] (Table 2).
Circadian locomotor activity and sleep were analyzed in flies expressing wild-type hα-syn (wt-αS) and two mutant isoforms, A53T-αS and TP-αS, specifically in serotonergic and/or dopaminergic neurons [96]. A53T-αS (carrying a single amino acid substitution) is able to form pre-fibrillar oligomers that aggregate later than wt-αS, while TP-αS (characterized by the modification of three amino acids within the same αS molecule; Table 2) tends to oligomerize but is unable to aggregate. According to this feature, the TP-αS mutant should present a more aggressive phenotype due to its toxicity. In particular, overexpression of the wt-αS or TP-αS isoforms affected the locomotor activity profiles in 12:12 LD, since at light-off, only a mild reduction in locomotion was observed in TP-αS transgenic flies, while this locomotor reduction was clearly evident in control flies. Furthermore, in TP mutants, older individuals (30 days) did not show the typical anticipatory locomotor behavior before lights-off in LD and presented a longer circadian period in DD. This phenomenon might be related to mutant hα-syn oligomerization-induced neuronal malfunction and/or death. Given that the above described genetic manipulations occurred at the level of the aminergic system, the observed circadian behavioral alterations were probably related to effects occurring downstream from the master clock [79].
Besides circadian abnormalities, the expression of wild-type and mutant forms of hα-syn also induced alterations in the sleep profile of transgenic flies. Globally, these abnormalities were related to variations in the amount of sleep and the number and length of sleep episodes during the day or night, with such alterations appearing already in young flies, which is suggestive of neuronal dysfunction rather than neuronal death in the transgenic flies [96].
3.3.2. Parkin and PINK1
hParkin and hPINK1 are mitochondrial-associated proteins which, when mutated, have been reported to cause autosomic recessive forms of PD. The hPARK2 gene encodes hParkin, a cytosolic E3-ubiquitin ligase, with an amino-terminal ubiquitin-like domain and a carboxy-terminal ubiquitin ligase domain. hPARK6 encodes the mitochondrially localized phosphatase and tensin homolog (PTEN)-induced kinase 1 (hPINK1). Although found in different subcellular compartments, the two proteins were shown (for the first time in Drosophila) to play a fundamental role in mitochondrial quality control [101,113,114,115]. Drosophila models of Parkin (in the fruit fly encoded by dpark) and PINK1 present a shortened lifespan, male sterility, swollen mitochondria, and altered locomotor behavior, mainly as a result of muscle damage. The two proteins are thought to belong to the same pathway, with dPINK1 acting upstream of dParkin [116].
Experiments to evaluate the endogenous circadian rhythmicity of dPink1 and dpark loss-of-function mutant flies indicate that dPink1 mutants show the strongest circadian locomotor activity defects in DD, when compared to dpark null mutants. Furthermore, both mutations were associated with alterations in the electrophysiological properties of the l-LNvs, although they influenced the electrophysiological features of these clock neurons in different ways [97] (Table 2).
In addition, a recent report showed that dPink1 and dpark mutant flies were characterized in LD conditions by the absence of circadian locomotor anticipatory activity in the morning (and a reduced anticipation in the evening), and more fragmented sleep compared to wild-type controls [98]. A specific downregulation of these genes mapped the circadian abnormalities at the level of the LNvs and sleep defects in the brain insulin-producing cells (IPCs), known for their role in sleep maintenance [117]. Interestingly, the morning, anticipation abnormalities were specifically due to an excessive number of endoplasmic-reticulum (ER)-mitochondrial contacts, which determined anomalous lipid trafficking and a depletion in the phosphatidylserine ER content at the level of the mutant LNvs. This alteration, in turn, caused a disruption in the physiological production of the dPDF-containing vesicles, and in the failure of proper neuropeptide localization in the LNv terminals [98]. It is noteworthy that an abnormal distribution of the circadian neuropeptide, hVasoactive Intestinal Peptide (analogous to the dPDF) was also detected in hypothalamic neurons differentiated from pluripotent stem cells of PD patients carrying mutations in hPARK6 and hPARK2 genes, indicating the evolutionary conservation of Pink1 and Parkin functions [98].
Cellular, circadian, and sleep defects were rescued by feeding dPink1 and dpark mutant flies with phosphatidylserine, suggesting that these abnormalities were not linked to neurodegeneration, but rather to a neuronal disfunction. Moreover, the same feeding rescue results suggest that the circadian and sleep alterations were mainly associated with neuronal networks different from the dopaminergic system (i.e., LNvs and IPCs for circadian and sleep defects, respectively) [98].
3.3.3. DJ-1
In humans, mutations in DJ-1 have been associated with early-onset forms of PD, with recessive inheritance. hDJ-1 is a small dimeric protein, with ubiquitous expression and a conserved amino acid sequence from Prokaryotes to Eukaryotes. The protein is mainly cytosolic, but a small fraction has also been detected within mitochondria [118,119] and in the nucleus [120]. Although the physiological role of hDJ-1 is still controversial, many activities have been ascribed to the protein, including oncogenesis, antioxidative responses, male fertility, and transcription [121]. Nonetheless, the most corroborated function is its involvement in anti-oxidant responses, activating pro-survival pathways while inhibiting cell death [121]. Differently from mammals, Drosophila’s genome encodes two DJ-1 paralogs, ddj-1α and ddj-1β. While ddj-1α is principally transcribed in the male testes [107] and partly in the brain [122], ddj-1β, similarly to the human protein, is ubiquitously expressed [107]. Double knock-out (DKO) flies are viable and fertile and are characterized by a normal lifespan, but show sensitivity to oxidative insults determined by chronic exposure to 20 mM Paraquat (PQ). In particular, dDj-1β seems to be primarily involved in oxidative stress protection, since dj-1β− single mutants display sensitivity to oxidative stressors similarly to ddj-1α−; ddj-1β− DKO flies [107]. In our laboratories, we investigated the consequences of ddj-1s loss on locomotor activity profiles in 12:12 LD, and circadian rhythmicity in DD. As PD is an age-related pathology, young (3–5-day-old) and old (30-day-old) flies were analyzed. Both young and old individuals exhibited a normal locomotor activity profile in LD, with both anticipatory morning and evening activity (Figure 2A–D). No particular abnormalities were detected in the circadian periodicity of neither young nor old ddj-1α− and ddj-1β− DKO flies in DD conditions (Figure 2E). Circadian locomotor behavior of ddj-1α− and ddj-1β− DKO flies was not even affected under a mild oxidative-stress treatment, consisting of chronic exposure to a low concentration of PQ (1 mM). Similar treatments are known to cause arrhythmicity in foxo− mutant flies, a genetic situation which is known to produce an oxidative stress-sensitized background [123]. In our case, 1 mM PQ chronic exposure was able to determine a slight, but significant, decrease in vitality in ddj-1α− and ddj-1β− DKO flies, indicating the effectiveness of the oxidative-stress treatment (Figure 2F). However, neither the percentage of rhythmic flies nor the periodicity were significantly modified under this oxidative-stress condition (Figure 2E). Since DJ-1 is known to have a role in the protection against oxidative stress, our data suggest that this factor does not influence the physiological mechanisms controlling rhythmic behavior in flies.
4. Drosophila as a Model to Evaluate the Effect of Circadian Disruptions on Neurodegenerative Processes
An interesting aspect which requires further investigation is whether circadian clock disruptions can promote/enhance neurodegenerative processes, in this way representing an additional risk factor for the development of PD pathology. Indeed, it is currently accepted that an impairment in normal rhythmicity favors the activation of gene expression and pro-inflammatory responses, which may play a role also in neurodegeneration. A proposed pathway that associates the master clock to neurodegeneration is through the modulation of oxidative stress responses. However other mechanisms such as neuroinflammation or an impaired degradation of pathogenic protein aggregates might also be involved [125].
Studies in mice showed that the KO of mBmal1 or the DKO of mClk and its paralog mNpas2, promoted premature aging, increased oxidative stress, age-dependent neuropathology as well as synaptic degeneration, and indicated these factors as possible players in the anti-oxidant response, through a downstream activation of antioxidant response transcription factors [126,127,128]. In addition, as mentioned above, it was demonstrated that an environmental disruption of the circadian clock intensified PD-related motor deficits in MPTP-treated mice. These abnormalities were associated with a loss of dopaminergic neurons and a robust neuroinflammatory reaction in the Substantia Nigra [45]. Similarly, in 6-OHDA treated mice, mutations of the circadian clock element mREV-ERBα (the mBmal1 transcriptional repressor) promoted 6-OHDA-dependent locomotor deficits, dopaminergic neurodegeneration, and neuroinflammation in the vertebral midbrain [129].
Compelling evidence indicates that D. melanogaster represents a valuable model to investigate these aspects. In flies the oxidative stress response is under circadian control, as wild-type individuals exposed to an oxidative insult during the day time were more sensitive than those treated at night. Moreover, flies with a disrupted circadian clock, as a result of either maintaining wild-type flies under constant light (LL) conditions, or by using clock mutants such as per0, did not show daily fluctuations in this response [130]. In fact, aged per0 mutant flies displayed higher impaired locomotor ability, increased susceptibility to oxidative damage, and expanded vacuolization in the central brain. Furthermore, the absence of functional dPER exacerbated damage in neurodegeneration-prone mutants, suggesting that in flies this clock gene acts as a neuroprotective factor during aging [131,132]. Moreover, in flies a long term arrythmicity is able to determine an acceleration in the manifestation of phenotypes typical of the aging process, since wild-type flies in LL conditions as well as amorphic (per0, tim0 or cyc0) or hypomorphic (ClkAR) clock mutants showed a significantly reduced lifespan and more pronounced locomotor impairments compared to their age-related controls [131,133].
In Drosophila, a connection between clock molecular elements, circadian neuronal cells and neurodegeneration was recently demonstrated by two different groups [133,134].
In particular, within the circadian neuronal network, Means and colleagues identified a molecular pathway involving the circadian kinase dDBT, the tetratricopeptide repeat-containing protein dSpaghetti (dSpag), and the dDronc caspase, implicated in the cleavage of the tubulin-associated unit (dTau) protein [134]. Tau is an evolutionarily conserved protein, which, in the truncated form, shows enhanced aggregation, contributing to the neuronal death typical of human neurodegenerative diseases, including Alzheimer’s disease [135] and, albeit to a lesser extent, PD [136]. Following an extensive series of experiments, and based on the use of mutant and transgenic flies, the authors obtained compelling indications to the effect that under normal conditions, dSpag controls the stability of dDBT, which in turn inhibits the activation of dDronc. On the contrary, following perturbations which determine the degradation of dDBT (for example via a dspag KD), dDronc is activated, leading to an enhanced assembly of truncated dTau (and likely other still unknown targets), which in turn contributes to neuronal death [134].
The same authors also demonstrated that a dspag KD, driven specifically in clock neurons, determined the activation of dDronc in regions outside the circadian network (i.e., optic lobe). dDronc activation was evident when flies were exposed to light and modulated by dPDF signaling, indicating a light-mediated communication between circadian neurons and other brain regions, leading to a modulation of the activity of molecular factors involved in apoptosis [134]. Similar abnormal activation of dDronc was also detected in young flies expressing a mutated form of dDBT (DBTK/R) [137] in the circadian network, in dClk null mutant individuals (Clk Jrk) and in very old wild-type flies, which are characterized by alterations in dDBT metabolism. These data suggest the dSpag-dDBT-dDronc pathway as being one of the possible molecular mechanisms by means of which a disrupted clock might affect the state of health and vitality of neuronal cells.
Evidence for a connection between the circadian clock and neurodegeneration more specifically involving the dopaminergic system was derived from the work by Vaccaro and coauthors [133]. In this study, the authors demonstrated that, in flies, the core clock element dClk showed additional clock-independent activity within the s-LNvs, which was required to avoid locomotor impairments typical of ClkAR hypomorphic mutants. Experiments performed by specifically knocking down or expressing dClk within s-LNvs in a ClkAR mutant background indicated that this function was, however, linked to dPDF-mediated communication and protected against the neurodegeneration of the PPL1 dopaminergic neurons, conceivably through the control of apoptotic phenomena, possibly acting via paracrine signaling between the two systems [133]. However, the communication might also be synaptic, since synaptic connections between axons of the sLNvs and dopaminergic neurons were recently described, although the specific dopaminergic cluster/s involved in this connection has not yet been clearly identified [94]. These data revealed a functional communication between the circadian clock network and the dopaminergic systems in Drosophila, and implied a direct involvement of a circadian clock element in the maintenance of healthy dopaminergic neurons during aging in flies. Interestingly, an age-dependent neuroinflammatory phenotype, with astrogliosis and synaptic terminal degeneration, was detected in the brain cortex of mice following the simultaneous deletion of mClk and its paralog mNpas2 [127]. However, in this case, the presence of specific degeneration in dopaminergic neurons was not investigated.
5. Conclusions and Further Perspectives
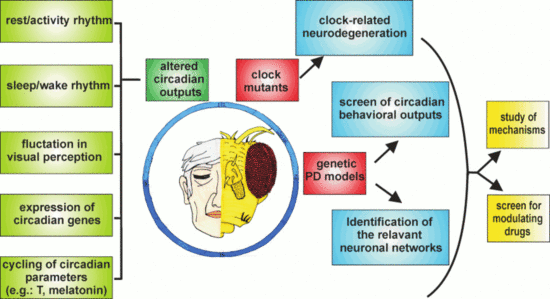
Although the number of studies concerning the relationship between PD and the circadian clock performed so far using Drosophila is still limited, the results obtained are, in our opinion, promising, particularly in view of considering this organism as an additional model to explore these aspects (Figure 3).
The use of circadian locomotor activity as a first-level behavioral output has indicated that some of the genes implicated in the early-onset forms of PD (e.g., hSNCA, dPink1, and dpark) were able to affect the circadian rhythmicity in Drosophila, but others, such as the ddj-1s, were not. Given the multifactorial etiology of PD, globally, these data suggest that the genetic factors involved in the pathology might not control the same phenotypes, and an initial screen in Drosophila could provide a useful indication to guide subsequent analyses in more complex organisms.
In addition, the relative ease with which downregulation or overexpression of candidate genes in specific neurons can be obtained in flies allows to address the question of which, among the different types of neurons, are mainly involved in affecting the circadian output, as seen in the case of hα-syn in aminergic neurons [96] and dPink1 or dpark in LNv neurons [98]. This strategy could contribute to elucidate the causal factors of the circadian abnormalities in PD, as demonstrated by Valadas and colleagues who associated the dPink1− and dpark− circadian defects mainly to alterations in the neuropeptide vesicle formation at the level of the circadian neurons [98]. The same approach might therefore be extended to other PD-related genes evaluating whether they directly affect the molecular and/or the electrophysiological properties of the circadian neurons, or whether they act through the dopaminergic system or other circuits, which in turn modify the activity of the clock neuronal network.
Moreover, besides locomotor activity, other phenotypes show a circadian pattern in Drosophila. Among these is the optomotor response [138,139], the study of which might constitute a useful phenotypic output for the symptoms related to visual acuity and contrast sensitivity detected in some PD patients and fly PD models [16,140,141]. The study of these circadian phenotypes in Drosophila PD models (including the manipulation of the expression of PD-related gene products in specific sets of cells and/or neurons) might contribute to the identification of the cellular network/s, the malfunction of which might determine such vision-related defects in flies. Despite the differences in the interactions between the visual and the circadian systems in insects and mammals, this kind of information might be relevant to the clarification of the mechanisms underlying the presence of circadian alterations also in individuals affected by idiopathic forms of PD. Generally, dopaminergic neurons are considered to be the primary target in the explanation of the motor symptoms observed in PD patients. However, non-motor symptoms, including some circadian abnormalities, might originate from the “spreading” of alterations to other (among which are possibly also non-dopaminergic) neurons [142,143].
Data obtained from neurodegeneration studies in flies support the notion that Drosophila might be an additional model to evaluate the impact of a misregulated or disrupted circadian clock in the neurodegenerative processes. Given the differences in the structure and organization of the circadian and dopaminergic systems between flies and mammals, it is likely that not all the information might be directly transposable from one model to the other. For example, it is important to mention that, while in flies, dper seems to have a role as a neuroprotective factor in aging brains [131,132], in mice, the DKO of the paralogs mPer1 and mPer2 did not cause any neuroinflammatory phenotype [127]. In such DKO mice, we can speculate that the presence of a third Per gene (mPer3) might (partially) substitute for the lack of mPer1 and mPer2 in the putative neuroprotective function. However, no data exploring this aspect are yet available. It is worth underlining that when data obtained from both models are consistent, the use of Drosophila allows to more readily perform experiments which both take longer and are usually more complex in mammals. For example, tissue-specific rescue experiments in mutant backgrounds, which can be conducted with relative ease in Drosophila, allow to rapidly explore multiple experimental conditions in order to understand the cellular and molecular mechanisms involved in the control of a specific phenotype, including neurodegeneration.
Furthermore, because PD, like most neurodegenerative diseases, progresses over a period of decades, pharmacological interventions that even slightly modulate one or more of the pathways involved in the pathogenesis of the disease could delay the progression or improve function, leading to an enhancement in the quality of life for patients suffering from such terribly debilitating disorders. In this respect, the various available Drosophila melanogaster genetic models of PD could provide an advantage as first-stage screens for modulatory drugs (as seen in [98]). In particular, the relatively short lifespan of fruit flies facilitates the search for modulators of PD, which develops its symptomatology over periods of time often spanning a large part of the organism’s lifespan. In this respect, in Drosophila, it is feasible to conduct lifelong studies during which it would be possible to explore the pharmacological modulation, for instance, of perturbations in the circadian rhythms resulting from the manipulation of genetic determinants of PD, such as hα-syn, and the homologs of the human Pink1, Parkin, or LRRK2.
An aspect that has been considered partly in PD models of Drosophila so far is sleep, which has been examined in dPink1 and dpark mutants [98] and in flies overexpressing hα-syn, either by characterizing sleep profiles under basal conditions [96] or by evaluating the effects of sleep deprivation on memory [144]. In Drosophila and mammals, sleep is controlled by both circadian and homeostatic systems and, as mentioned above, is often affected in PD patients. Under this perspective, it is interesting to remind that, using the appropriate sampling frequency, the same raw data collected for the analysis of daily locomotor behavior in flies could be processed to determine the structure of sleep [145,146]. This would therefore allow to evaluate the presence of sleep abnormalities in the different genetic PD-related forms, and possibly to understand the neuronal circuit/s mainly affected in Drosophila, as well as to evaluate the rescue of altered sleep patterns as an end-point in primary screens for pharmacological interventions.
Author Contributions
Conceptualization of the manuscript, F.D.L., M.B., M.A.Z. and F.S.; data analysis, F.D.L. and F.S. All authors wrote, read and approved the manuscript.
Funding
This work was founded by grant from the Department of Biology, University of Padova (Italy) to MB PRID-SEED 2018 and to FS PRID-SEED 2017.
Acknowledgments
We would like to thank Milo Tolando for the support in the graphical abstract.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shults, C.W. Lewy bodies. Proc. Natl. Acad. Sci. USA 2006, 103, 1661–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barone, P.; Antonini, A.; Colosimo, C.; Marconi, R.; Morgante, L.; Avarello, T.P.; Bottacchi, E.; Cannas, A.; Ceravolo, G.; Ceravolo, R.; et al. The PRIAMO study: A multicenter assessment of nonmotor symptoms and their impact on quality of life in Parkinson’s disease. Mov. Disord. 2009, 24, 1641–1649. [Google Scholar] [CrossRef]
- Videnovic, A.; Golombek, D. Circadian Dysregulation in Parkinson’s Disease. Neurobiol. Sleep Circadian Rhythm. 2017, 2, 53–58. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Wang, F.; Hu, L.-F.; Liu, C.-F. A New Perspective for Parkinson’s Disease: Circadian Rhythm. Neurosci. Bull. 2017, 33, 62–72. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K. Neuropathological Staging of Brain Pathology in Sporadic Parkinson’s disease: Separating the Wheat from the Chaff. J. Parkinson’s Dis. 2017, 7, S71–S85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeve, B.F. REM sleep behavior disorder: Updated review of the core features, the REM sleep behavior disorder-neurodegenerative disease association, evolving concepts, controversies, and future directions. Ann. N. Y. Acad. Sci. 2010, 1184, 15–54. [Google Scholar] [CrossRef] [PubMed]
- Korshunov, K.S.; Blakemore, L.J.; Trombley, P.Q. Dopamine: A Modulator of Circadian Rhythms in the Central Nervous System. Front. Cell. Neurosci. 2017, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Karimi-Moghadam, A.; Charsouei, S.; Bell, B.; Jabalameli, M.R. Parkinson Disease from Mendelian Forms to Genetic Susceptibility: New Molecular Insights into the Neurodegeneration Process. Cell. Mol. Neurobiol. 2018, 38, 1153–1178. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, U. Timing to perfection: The biology of central and peripheral circadian clocks. Neuron 2012, 74, 246–260. [Google Scholar] [CrossRef]
- Piccini, P.; Del Dotto, P.; Pardini, C.; D’Antonio, P.; Rossi, G.; Bonuccelli, U. Diurnal worsening in Parkinson patients treated with levodopa. Riv. Neurol. 1991, 61, 219–224. [Google Scholar]
- Van Hilten, J.J.; Hoogland, G.; van der Velde, E.A.; Middelkoop, H.A.; Kerkhof, G.A.; Roos, R.A. Diurnal effects of motor activity and fatigue in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 1993, 56, 874–877. [Google Scholar] [CrossRef] [PubMed]
- Placidi, F.; Izzi, F.; Romigi, A.; Stanzione, P.; Marciani, M.G.; Brusa, L.; Sperli, F.; Galati, S.; Pasqualetti, P.; Pierantozzi, M. Sleep-wake cycle and effects of cabergoline monotherapy in de novo Parkinson’s disease patients. An ambulatory polysomnographic study. J. Neurol. 2008, 255, 1032–1037. [Google Scholar] [CrossRef] [PubMed]
- Prudon, B.; Duncan, G.W.; Khoo, T.K.; Yarnall, A.J.; Anderson, K.N. Primary sleep disorder prevalence in patients with newly diagnosed Parkinson’s disease. Mov. Disord. 2014, 29, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, K.R.; Prieto-Jurcynska, C.; Naidu, Y.; Mitra, T.; Frades-Payo, B.; Tluk, S.; Ruessmann, A.; Odin, P.; Macphee, G.; Stocchi, F.; et al. The nondeclaration of nonmotor symptoms of Parkinson’s disease to health care professionals: An international study using the nonmotor symptoms questionnaire. Mov. Disord. 2010, 25, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Avidan, A.; Hays, R.D.; Diaz, N.; Bordelon, Y.; Thompson, A.W.; Vassar, S.D.; Vickrey, B.G. Associations of sleep disturbance symptoms with health-related quality of life in Parkinson’s disease. J. Neuropsychiatry Clin. Neurosci. 2013, 25, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Weil, R.S.; Schrag, A.E.; Warren, J.D.; Crutch, S.J.; Lees, A.J.; Morris, H.R. Visual dysfunction in Parkinson’s disease. Brain 2016, 139, 2827–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulens, C.; Meerwaldt, J.D.; Van der Wildt, G.J.; Van Deursen, J.B. Effect of levodopa treatment on contrast sensitivity in Parkinson’s disease. Ann. Neurol. 1987, 22, 365–369. [Google Scholar] [CrossRef]
- Büttner, T.; Kuhn, W.; Patzold, T.; Przuntek, H. L-Dopa improves colour vision in Parkinson’s disease. J. Neural Transm. Park Dis. Dement. Sect. 1994, 7, 13–19. [Google Scholar] [CrossRef]
- Struck, L.K.; Rodnitzky, R.L.; Dobson, J.K. Circadian fluctuations of contrast sensitivity in Parkinson’s disease. Neurology 1990, 40, 467–470. [Google Scholar] [CrossRef]
- Zhong, G.; Bolitho, S.; Grunstein, R.; Naismith, S.L.; Lewis, S.J.G. The relationship between thermoregulation and REM sleep behaviour disorder in Parkinson’s disease. PLoS ONE 2013, 8, e72661. [Google Scholar] [CrossRef]
- Ejaz, A.A.; Sekhon, I.S.; Munjal, S. Characteristic findings on 24-h ambulatory blood pressure monitoring in a series of patients with Parkinson’s disease. Eur. J. Intern. Med. 2006, 17, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, T.; Kitano, Y.; Kuno, S. Blood pressure fluctuation and hypertension in patients with Parkinson’s disease. Brain Behav. 2013, 3, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Breen, D.P.; Vuono, R.; Nawarathna, U.; Fisher, K.; Shneerson, J.M.; Reddy, A.B.; Barker, R.A. Sleep and circadian rhythm regulation in early Parkinson disease. JAMA Neurol. 2014, 71, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Videnovic, A.; Noble, C.; Reid, K.J.; Peng, J.; Turek, F.W.; Marconi, A.; Rademaker, A.W.; Simuni, T.; Zadikoff, C.; Zee, P.C. Circadian melatonin rhythm and excessive daytime sleepiness in Parkinson disease. JAMA Neurol. 2014, 71, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Bolitho, S.J.; Naismith, S.L.; Rajaratnam, S.M.W.; Grunstein, R.R.; Hodges, J.R.; Terpening, Z.; Rogers, N.; Lewis, S.J.G. Disturbances in melatonin secretion and circadian sleep-wake regulation in Parkinson disease. Sleep Med. 2014, 15, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Liu, S.; Sothern, R.B.; Xu, S.; Chan, P. Expression of clock genes Per1 and Bmal1 in total leukocytes in health and Parkinson’s disease. Eur. J. Neurol. 2010, 17, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Wang, B.; Zhang, Y.-B.; Ding, H.; Zhang, Y.; Yu, J.; Gu, M.; Chan, P.; Cai, Y. Association of ARNTL and PER1 genes with Parkinson’s disease: A case-control study of Han Chinese. Sci. Rep. 2015, 5, 15891. [Google Scholar] [CrossRef] [PubMed]
- Schnell, A.; Albrecht, U.; Sandrelli, F. Rhythm and mood: Relationships between the circadian clock and mood-related behavior. Behav. Neurosci. 2014, 128, 326–343. [Google Scholar] [CrossRef] [PubMed]
- Thenganatt, M.A.; Jankovic, J. Parkinson disease subtypes. JAMA Neurol. 2014, 71, 499–504. [Google Scholar] [CrossRef]
- Fifel, K. Alterations of the circadian system in Parkinson’s disease patients. Mov. Disord. 2017, 32, 682–692. [Google Scholar] [CrossRef]
- Kudo, T.; Loh, D.H.; Truong, D.; Wu, Y.; Colwell, C.S. Circadian dysfunction in a mouse model of Parkinson’s disease. Exp. Neurol. 2011, 232, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Fifel, K.; Cooper, H.M. Loss of dopamine disrupts circadian rhythms in a mouse model of Parkinson’s disease. Neurobiol. Dis. 2014, 71, 359–369. [Google Scholar] [CrossRef]
- Fifel, K.; Dkhissi-Benyahya, O.; Cooper, H.M. Lack of long-term changes in circadian, locomotor, and cognitive functions in acute and chronic MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) mouse models of Parkinson’s disease. Chronobiol. Int. 2013, 30, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Yamaguchi, E.; Takahashi, M.; Hashimura, K.; Shibata, T.; Nakamura, W.; Nakamura, T.J. Effects of age-related dopaminergic neuron loss in the substantia nigra on the circadian rhythms of locomotor activity in mice. Neurosci. Res. 2012, 74, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, A.; Matsunaga, N.; Okazaki, H.; Kakimoto, K.; Kimura, Y.; Azuma, H.; Ikeda, E.; Shiba, T.; Yamato, M.; Yamada, K.-I.; et al. A disruption mechanism of the molecular clock in a MPTP mouse model of Parkinson’s disease. Neuromol. Med. 2013, 15, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Hineno, T.; Mizobuchi, M.; Hiratani, K.; Inami, Y.; Kakimoto, Y. Disappearance of circadian rhythms in Parkinson’s disease model induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in dogs. Brain Res. 1992, 580, 92–99. [Google Scholar] [CrossRef]
- Fifel, K.; Vezoli, J.; Dzahini, K.; Claustrat, B.; Leviel, V.; Kennedy, H.; Procyk, E.; Dkhissi-Benyahya, O.; Gronfier, C.; Cooper, H.M. Alteration of daily and circadian rhythms following dopamine depletion in MPTP treated non-human primates. PLoS ONE 2014, 9, e86240. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, D.; Liu, W.; Li, S.; Chen, J.; Shen, Y.; Wang, F.; Hu, L.-F.; Liu, C.-F. Disruption of the Circadian Clock Alters Antioxidative Defense via the SIRT1-BMAL1 Pathway in 6-OHDA-Induced Models of Parkinson’s Disease. Oxid. Med. Cell. Longev. 2018, 2018, 4854732. [Google Scholar] [CrossRef]
- Gravotta, L.; Gavrila, A.M.; Hood, S.; Amir, S. Global depletion of dopamine using intracerebroventricular 6-hydroxydopamine injection disrupts normal circadian wheel-running patterns and PERIOD2 expression in the rat forebrain. J. Mol. Neurosci. 2011, 45, 162–171. [Google Scholar] [CrossRef]
- Isobe, Y.; Nishino, H. Circadian rhythm of drinking and running-wheel activity in rats with 6-hydroxydopamine lesions of the ventral tegmental area. Brain Res. 2001, 899, 187–192. [Google Scholar] [CrossRef]
- Ben, V.; Bruguerolle, B. Effects of bilateral striatal 6-OHDA lesions on circadian rhythms in the rat: A radiotelemetric study. Life Sci. 2000, 67, 1549–1558. [Google Scholar] [CrossRef]
- Li, S.-Y.; Wang, Y.-L.; Liu, W.-W.; Lyu, D.-J.; Wang, F.; Mao, C.-J.; Yang, Y.-P.; Hu, L.-F.; Liu, C.-F. Long-term Levodopa Treatment Accelerates the Circadian Rhythm Dysfunction in a 6-hydroxydopamine Rat Model of Parkinson’s Disease. Chin. Med. J. 2017, 130, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Glinka, Y.; Gassen, M.; Youdim, M.B. Mechanism of 6-hydroxydopamine neurotoxicity. J. Neural Transm. Suppl. 1997, 50, 55–66. [Google Scholar] [PubMed]
- Burns, R.S.; Chiueh, C.C.; Markey, S.P.; Ebert, M.H.; Jacobowitz, D.M.; Kopin, I.J. A primate model of parkinsonism: Selective destruction of dopaminergic neurons in the pars compacta of the substantia nigra by N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Proc. Natl. Acad. Sci. USA 1983, 80, 4546–4550. [Google Scholar] [CrossRef] [PubMed]
- Lauretti, E.; Di Meco, A.; Merali, S.; Praticò, D. Circadian rhythm dysfunction: A novel environmental risk factor for Parkinson’s disease. Mol. Psychiatry 2017, 22, 280–286. [Google Scholar] [CrossRef]
- Mendoza, J.; Challet, E. Circadian insights into dopamine mechanisms. Neuroscience 2014, 282, 230–242. [Google Scholar] [CrossRef]
- Witkovsky, P. Dopamine and retinal function. Doc. Ophthalmol. 2004, 108, 17–40. [Google Scholar] [CrossRef]
- Bellen, H.J.; Tong, C.; Tsuda, H. 100 years of Drosophila research and its impact on vertebrate neuroscience: A history lesson for the future. Nat. Rev. Neurosci. 2010, 11, 514–522. [Google Scholar] [CrossRef]
- Hales, K.G.; Korey, C.A.; Larracuente, A.M.; Roberts, D.M. Genetics on the Fly: A Primer on the Drosophila Model System. Genetics 2015, 201, 815–842. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Soriano, V.; Paricio, N. Drosophila models of Parkinson’s disease: Discovering relevant pathways and novel therapeutic strategies. Parkinson’s Dis. 2011, 2011, 520640. [Google Scholar] [CrossRef]
- Xiong, Y.; Yu, J. Modeling Parkinson’s Disease in Drosophila: What Have We Learned for Dominant Traits? Front. Neurol. 2018, 9, 228. [Google Scholar] [CrossRef]
- Nagoshi, E. Drosophila Models of Sporadic Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 3343. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.L.; Frenkel, L.; Ceriani, M.F. The Underlying Genetics of Drosophila Circadian Behaviors. Physiology 2018, 33, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Zordan, M.A.; Sandrelli, F. Circadian Clock Dysfunction and Psychiatric Disease: Could Fruit Flies have a Say? Front. Neurol. 2015, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Dubowy, C.; Sehgal, A. Circadian Rhythms and Sleep in Drosophila melanogaster. Genetics 2017, 205, 1373–1397. [Google Scholar] [CrossRef] [Green Version]
- Allada, R.; Chung, B.Y. Circadian Organization of Behavior and Physiology in Drosophila. Annu. Rev. Physiol. 2010, 72, 605–624. [Google Scholar] [CrossRef] [PubMed]
- Peschel, N.; Helfrich-Förster, C. Setting the clock--by nature: Circadian rhythm in the fruitfly Drosophila melanogaster. FEBS Lett. 2011, 585, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Grima, B.; Chélot, E.; Xia, R.; Rouyer, F. Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain. Nature 2004, 431, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Stoleru, D.; Peng, Y.; Agosto, J.; Rosbash, M. Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature 2004, 431, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Shafer, O.T. The Drosophila circadian clock is a variably coupled network of multiple peptidergic units. Science 2014, 343, 1516–1520. [Google Scholar] [CrossRef]
- Dissel, S.; Hansen, C.N.; Özkaya, Ö.; Hemsley, M.; Kyriacou, C.P.; Rosato, E. The logic of circadian organization in Drosophila. Curr. Biol. 2014, 24, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Ozkaya, O.; Rosato, E. The circadian clock of the fly: A neurogenetics journey through time. Adv. Genet. 2012, 77, 79–123. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Wu, B.; Price, J.L.; Zhao, Z. Circadian Rhythm Neuropeptides in Drosophila: Signals for Normal Circadian Function and Circadian Neurodegenerative Disease. Int. J. Mol. Sci. 2017, 18, 886. [Google Scholar] [CrossRef] [PubMed]
- King, A.N.; Sehgal, A. Molecular and circuit mechanisms mediating circadian clock output in the Drosophila brain. Eur. J. Neurosci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Emery, P.; So, W.V.; Kaneko, M.; Hall, J.C.; Rosbash, M. CRY, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity. Cell 1998, 95, 669–679. [Google Scholar] [CrossRef]
- Stanewsky, R.; Kaneko, M.; Emery, P.; Beretta, B.; Wager-Smith, K.; Kay, S.A.; Rosbash, M.; Hall, J.C. The cryb mutation identifies cryptochrome as a circadian photoreceptor in Drosophila. Cell 1998, 95, 681–692. [Google Scholar] [CrossRef]
- Lin, F.J.; Song, W.; Meyer-Bernstein, E.; Naidoo, N.; Sehgal, A. Photic signaling by cryptochrome in the Drosophila circadian system. Mol. Cell. Biol. 2001, 21, 7287–7294. [Google Scholar] [CrossRef] [PubMed]
- Koh, K.; Zheng, X.; Sehgal, A. JETLAG resets the Drosophila circadian clock by promoting light-induced degradation of TIMELESS. Science 2006, 312, 1809–1812. [Google Scholar] [CrossRef]
- Yoshii, T.; Todo, T.; Wülbeck, C.; Stanewsky, R.; Helfrich-Förster, C. Cryptochrome is present in the compound eyes and a subset of Drosophila’s clock neurons. J. Comp. Neurol. 2008, 508, 952–966. [Google Scholar] [CrossRef]
- Yoshii, T.; Hermann, C.; Helfrich-Förster, C. Cryptochrome-positive and -negative clock neurons in Drosophila entrain differentially to light and temperature. J. Biol. Rhythm. 2010, 25, 387–398. [Google Scholar] [CrossRef]
- Benito, J.; Houl, J.H.; Roman, G.W.; Hardin, P.E. The blue-light photoreceptor CRYPTOCHROME is expressed in a subset of circadian oscillator neurons in the Drosophila CNS. J. Biol. Rhythm. 2008, 23, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Helfrich-Förster, C.; Winter, C.; Hofbauer, A.; Hall, J.C.; Stanewsky, R. The circadian clock of fruit flies is blind after elimination of all known photoreceptors. Neuron 2001, 30, 249–261. [Google Scholar] [CrossRef]
- Renn, S.C.; Park, J.H.; Rosbash, M.; Hall, J.C.; Taghert, P.H. A pdf neuropeptide gene mutation and ablation of PDF neurons each cause severe abnormalities of behavioral circadian rhythms in Drosophila. Cell 1999, 99, 791–802. [Google Scholar] [CrossRef]
- He, C.; Cong, X.; Zhang, R.; Wu, D.; An, C.; Zhao, Z. Regulation of circadian locomotor rhythm by neuropeptide Y-like system in Drosophila melanogaster. Insect Mol. Biol. 2013, 22, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Johard, H.A.D.; Yoishii, T.; Dircksen, H.; Cusumano, P.; Rouyer, F.; Helfrich-Förster, C.; Nässel, D.R. Peptidergic clock neurons in Drosophila: Ion transport peptide and short neuropeptide F in subsets of dorsal and ventral lateral neurons. J. Comp. Neurol. 2009, 516, 59–73. [Google Scholar] [CrossRef] [PubMed]
- King, A.N.; Barber, A.F.; Smith, A.E.; Dreyer, A.P.; Sitaraman, D.; Nitabach, M.N.; Cavanaugh, D.J.; Sehgal, A. A Peptidergic Circuit Links the Circadian Clock to Locomotor Activity. Curr. Biol. 2017, 27, 1915.e5–1927.e5. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, D.J.; Geratowski, J.D.; Wooltorton, J.R.A.; Spaethling, J.M.; Hector, C.E.; Zheng, X.; Johnson, E.C.; Eberwine, J.H.; Sehgal, A. Identification of a circadian output circuit for rest:activity rhythms in Drosophila. Cell 2014, 157, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Cavey, M.; Collins, B.; Bertet, C.; Blau, J. Circadian rhythms in neuronal activity propagate through output circuits. Nat. Neurosci. 2016, 19, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.; Ng, F.; Lebestky, T.; Grygoruk, A.; Djapri, C.; Lawal, H.O.; Zaveri, H.A.; Mehanzel, F.; Najibi, R.; Seidman, G.; et al. Dispensable, redundant, complementary, and cooperative roles of dopamine, octopamine, and serotonin in Drosophila melanogaster. Genetics 2013, 193, 159–176. [Google Scholar] [CrossRef]
- Hirsh, J.; Riemensperger, T.; Coulom, H.; Iché, M.; Coupar, J.; Birman, S. Roles of dopamine in circadian rhythmicity and extreme light sensitivity of circadian entrainment. Curr. Biol. 2010, 20, 209–214. [Google Scholar] [CrossRef]
- Hanna, M.E.; Bednářová, A.; Rakshit, K.; Chaudhuri, A.; O’Donnell, J.M.; Krishnan, N. Perturbations in dopamine synthesis lead to discrete physiological effects and impact oxidative stress response in Drosophila. J. Insect Physiol. 2015, 73, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, F.S.; Tangredi, M.M.; Jackson, F.R. Glial cells physiologically modulate clock neurons and circadian behavior in a calcium-dependent manner. Curr. Biol. 2011, 21, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.; Jackson, F.R. Drosophila ebony activity is required in glia for the circadian regulation of locomotor activity. Neuron 2007, 55, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Davis, R.L. Eight different types of dopaminergic neurons innervate the Drosophila mushroom body neuropil: Anatomical and physiological heterogeneity. Front. Neural Circuits 2009, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Friggi-Grelin, F.; Coulom, H.; Meller, M.; Gomez, D.; Hirsh, J.; Birman, S. Targeted gene expression in Drosophila dopaminergic cells using regulatory sequences from tyrosine hydroxylase. J. Neurobiol. 2003, 54, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Kasture, A.S.; Hummel, T.; Sucic, S.; Freissmuth, M. Big Lessons from Tiny Flies: Drosophila melanogaster as a Model to Explore Dysfunction of Dopaminergic and Serotonergic Neurotransmitter Systems. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Seto, E.S. Dopamine dynamics and signaling in Drosophila: An overview of genes, drugs and behavioral paradigms. Exp. Anim. 2014, 63, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Gomez, A.; Turiegano, E.; León, Y.; Molina, I.; Torroja, L.; Canal, I. Ih current is necessary to maintain normal dopamine fluctuations and sleep consolidation in Drosophila. PLoS ONE 2012, 7, e36477. [Google Scholar] [CrossRef]
- Riemensperger, T.; Isabel, G.; Coulom, H.; Neuser, K.; Seugnet, L.; Kume, K.; Iché-Torres, M.; Cassar, M.; Strauss, R.; Preat, T.; et al. Behavioral consequences of dopamine deficiency in the Drosophila central nervous system. Proc. Natl. Acad. Sci. USA 2011, 108, 834–839. [Google Scholar] [CrossRef]
- Cichewicz, K.; Garren, E.J.; Adiele, C.; Aso, Y.; Wang, Z.; Wu, M.; Birman, S.; Rubin, G.M.; Hirsh, J. A new brain dopamine-deficient Drosophila and its pharmacological and genetic rescue. Genes Brain Behav. 2017, 16, 394–403. [Google Scholar] [CrossRef]
- Hamasaka, Y.; Nässel, D.R. Mapping of serotonin, dopamine, and histamine in relation to different clock neurons in the brain of Drosophila. J. Comp. Neurol. 2006, 494, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Abruzzi, K.C.; Zadina, A.; Luo, W.; Wiyanto, E.; Rahman, R.; Guo, F.; Shafer, O.; Rosbash, M. RNA-seq analysis of Drosophila clock and non-clock neurons reveals neuron-specific cycling and novel candidate neuropeptides. PLoS Genet. 2017, 13, e1006613. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Haynes, P.; Pírez, N.; Harrington, K.I.; Guo, F.; Pollack, J.; Hong, P.; Griffith, L.C.; Rosbash, M. Imaging analysis of clock neurons reveals light buffers the wake-promoting effect of dopamine. Nat. Neurosci. 2011, 14, 889–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potdar, S.; Sheeba, V. Wakefulness Is Promoted during Day Time by PDFR Signalling to Dopaminergic Neurons in Drosophila melanogaster. eNeuro 2018, 5. [Google Scholar] [CrossRef]
- Helfrich-Förster, C. Sleep in Insects. Annu. Rev. Entomol. 2018, 63, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Gajula Balija, M.B.; Griesinger, C.; Herzig, A.; Zweckstetter, M.; Jäckle, H. Pre-fibrillar α-synuclein mutants cause Parkinson’s disease-like non-motor symptoms in Drosophila. PLoS ONE 2011, 6, e24701. [Google Scholar] [CrossRef] [PubMed]
- Julienne, H.; Buhl, E.; Leslie, D.S.; Hodge, J.J.L. Drosophila PINK1 and parkin loss-of-function mutants display a range of non-motor Parkinson’s disease phenotypes. Neurobiol. Dis. 2017, 104, 15–23. [Google Scholar] [CrossRef]
- Valadas, J.S.; Esposito, G.; Vandekerkhove, D.; Miskiewicz, K.; Deaulmerie, L.; Raitano, S.; Seibler, P.; Klein, C.; Verstreken, P. ER Lipid Defects in Neuropeptidergic Neurons Impair Sleep Patterns in Parkinson’s Disease. Neuron 2018, 98, 1155–1169. [Google Scholar] [CrossRef]
- Feany, M.B.; Bender, W.W. A Drosophila model of Parkinson’s disease. Nature 2000, 404, 394–398. [Google Scholar] [CrossRef]
- Karpinar, D.P.; Balija, M.B.G.; Kügler, S.; Opazo, F.; Rezaei-Ghaleh, N.; Wender, N.; Kim, H.-Y.; Taschenberger, G.; Falkenburger, B.H.; Heise, H.; et al. Pre-fibrillar alpha-synuclein variants with impaired beta-structure increase neurotoxicity in Parkinson’s disease models. EMBO J. 2009, 28, 3256–3268. [Google Scholar] [CrossRef]
- Park, J.; Lee, S.B.; Lee, S.; Kim, Y.; Song, S.; Kim, S.; Bae, E.; Kim, J.; Shong, M.; Kim, J.-M.; et al. Mitochondrial dysfunction in Drosophila PINK1 mutants is complemented by parkin. Nature 2006, 441, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, A.J.; Theodore, D.A.; Greene, J.C.; Benes, H.; Wes, P.D.; Pallanck, L.J. Increased glutathione S-transferase activity rescues dopaminergic neuron loss in a Drosophila model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2005, 102, 8024–8029. [Google Scholar] [CrossRef]
- Yang, Y.; Gehrke, S.; Haque, M.E.; Imai, Y.; Kosek, J.; Yang, L.; Beal, M.F.; Nishimura, I.; Wakamatsu, K.; Ito, S.; et al. Inactivation of Drosophila DJ-1 leads to impairments of oxidative stress response and phosphatidylinositol 3-kinase/Akt signaling. Proc. Natl. Acad. Sci. USA 2005, 102, 13670–13675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzies, F.M.; Yenisetti, S.C.; Min, K.-T. Roles of Drosophila DJ-1 in survival of dopaminergic neurons and oxidative stress. Curr. Biol. 2005, 15, 1578–1582. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, S.Y.; Cha, G.-H.; Lee, S.B.; Kim, S.; Chung, J. Drosophila DJ-1 mutants show oxidative stress-sensitive locomotive dysfunction. Gene 2005, 361, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Song, S.; Hong, Y.K.; Choi, G.; Suh, Y.S.; Han, S.Y.; Lee, M.; Park, S.H.; Lee, J.H.; Lee, S.; et al. Drosophila DJ-1 Decreases Neural Sensitivity to Stress by Negatively Regulating Daxx-Like Protein through dFOXO. PLoS Genet. 2013, 9, e1003412. [Google Scholar] [CrossRef]
- Meulener, M.; Whitworth, A.J.; Armstrong-Gold, C.E.; Rizzu, P.; Heutink, P.; Wes, P.D.; Pallanck, L.J.; Bonini, N.M. Drosophila DJ-1 mutants are selectively sensitive to environmental toxins associated with Parkinson’s disease. Curr. Biol. 2005, 15, 1572–1577. [Google Scholar] [CrossRef]
- Hao, L.-Y.; Giasson, B.I.; Bonini, N.M. DJ-1 is critical for mitochondrial function and rescues PINK1 loss of function. Proc. Natl. Acad. Sci. USA 2010, 107, 9747–9752. [Google Scholar] [CrossRef] [Green Version]
- Bengoa-Vergniory, N.; Roberts, R.F.; Wade-Martins, R.; Alegre-Abarrategui, J. Alpha-synuclein oligomers: A new hope. Acta Neuropathol. 2017, 134, 819–838. [Google Scholar] [CrossRef]
- Burré, J.; Vivona, S.; Diao, J.; Sharma, M.; Brunger, A.T.; Südhof, T.C. Properties of native brain α-synuclein. Nature 2013, 498, E4. [Google Scholar] [CrossRef]
- Burré, J. The Synaptic Function of α-Synuclein. J. Parkinsons Dis 2015, 5, 699–713. [Google Scholar] [CrossRef] [PubMed]
- Auluck, P.K.; Chan, H.Y.E.; Trojanowski, J.Q.; Lee, V.M.Y.; Bonini, N.M. Chaperone suppression of alpha-synuclein toxicity in a Drosophila model for Parkinson’s disease. Science 2002, 295, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.E.; Dodson, M.W.; Jiang, C.; Cao, J.H.; Huh, J.R.; Seol, J.H.; Yoo, S.J.; Hay, B.A.; Guo, M. Drosophila pink1 is required for mitochondrial function and interacts genetically with parkin. Nature 2006, 441, 1162–1166. [Google Scholar] [CrossRef] [PubMed]
- Cackovic, J.; Gutierrez-Luke, S.; Call, G.B.; Juba, A.; O’Brien, S.; Jun, C.H.; Buhlman, L.M. Vulnerable Parkin Loss-of-Function Drosophila Dopaminergic Neurons Have Advanced Mitochondrial Aging, Mitochondrial Network Loss and Transiently Reduced Autophagosome Recruitment. Front. Cell. Neurosci. 2018, 12, 39. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, T.; Vilain, S.; Vints, K.; Gounko, N.; Verstreken, P.; Vandenberghe, W. Deficiency of parkin and PINK1 impairs age-dependent mitophagy in Drosophila. Elife 2018, 7. [Google Scholar] [CrossRef]
- Pickrell, A.M.; Youle, R.J. The roles of PINK1, parkin, and mitochondrial fidelity in Parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Crocker, A.; Shahidullah, M.; Levitan, I.B.; Sehgal, A. Identification of a neural circuit that underlies the effects of octopamine on sleep:wake behavior. Neuron 2010, 65, 670–681. [Google Scholar] [CrossRef]
- Canet-Avilés, R.M.; Wilson, M.A.; Miller, D.W.; Ahmad, R.; McLendon, C.; Bandyopadhyay, S.; Baptista, M.J.; Ringe, D.; Petsko, G.A.; Cookson, M.R. The Parkinson’s disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. USA 2004, 101, 9103–9108. [Google Scholar] [CrossRef]
- Junn, E.; Jang, W.H.; Zhao, X.; Jeong, B.S.; Mouradian, M.M. Mitochondrial localization of DJ-1 leads to enhanced neuroprotection. J. Neurosci. Res. 2009, 87, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Park, Y.-J.; Hwang, I.-Y.; Youdim, M.B.H.; Park, K.-S.; Oh, Y.J. Nuclear translocation of DJ-1 during oxidative stress-induced neuronal cell death. Free Radic. Biol. Med. 2012, 53, 936–950. [Google Scholar] [CrossRef]
- Biosa, A.; Sandrelli, F.; Beltramini, M.; Greggio, E.; Bubacco, L.; Bisaglia, M. Recent findings on the physiological function of DJ-1: Beyond Parkinson’s disease. Neurobiol. Dis. 2017, 108, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Aradska, J.; Bulat, T.; Sialana, F.J.; Birner-Gruenberger, R.; Erich, B.; Lubec, G. Gel-free mass spectrometry analysis of Drosophila melanogaster heads. Proteomics 2015, 15, 3356–3360. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yang, Z.; Yue, Z.; Alvarez, J.D.; Sehgal, A. FOXO and insulin signaling regulate sensitivity of the circadian clock to oxidative stress. Proc. Natl. Acad. Sci. USA 2007, 104, 15899–15904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zordan, M.A.; Benna, C.; Mazzotta, G. Monitoring and analyzing Drosophila circadian locomotor activity. Methods Mol. Biol. 2007, 362, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Musiek, E.S.; Holtzman, D.M. Mechanisms linking circadian clocks, sleep, and neurodegeneration. Science 2016, 354, 1004–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondratov, R.V.; Vykhovanets, O.; Kondratova, A.A.; Antoch, M.P. Antioxidant N-acetyl-L-cysteine ameliorates symptoms of premature aging associated with the deficiency of the circadian protein BMAL1. Aging 2009, 1, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Musiek, E.S.; Lim, M.M.; Yang, G.; Bauer, A.Q.; Qi, L.; Lee, Y.; Roh, J.H.; Ortiz-Gonzalez, X.; Dearborn, J.T.; Culver, J.P.; et al. Circadian clock proteins regulate neuronal redox homeostasis and neurodegeneration. J. Clin. Investig. 2013, 123, 5389–5400. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Moulik, M.; Fang, Z.; Saha, P.; Zou, F.; Xu, Y.; Nelson, D.L.; Ma, K.; Moore, D.D.; Yechoor, V.K. Bmal1 and β-cell clock are required for adaptation to circadian disruption, and their loss of function leads to oxidative stress-induced β-cell failure in mice. Mol. Cell. Biol. 2013, 33, 2327–2338. [Google Scholar] [CrossRef]
- Kim, J.; Jang, S.; Choi, M.; Chung, S.; Choe, Y.; Choe, H.K.; Son, G.H.; Rhee, K.; Kim, K. Abrogation of the Circadian Nuclear Receptor REV-ERBα Exacerbates 6-Hydroxydopamine-Induced Dopaminergic Neurodegeneration. Mol. Cells 2018, 41, 742–752. [Google Scholar] [CrossRef]
- Krishnan, N.; Davis, A.J.; Giebultowicz, J.M. Circadian regulation of response to oxidative stress in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2008, 374, 299–303. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, N.; Kretzschmar, D.; Rakshit, K.; Chow, E.; Giebultowicz, J.M. The circadian clock gene period extends healthspan in aging Drosophila melanogaster. Aging 2009, 1, 937–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, N.; Rakshit, K.; Chow, E.S.; Wentzell, J.S.; Kretzschmar, D.; Giebultowicz, J.M. Loss of circadian clock accelerates aging in neurodegeneration-prone mutants. Neurobiol. Dis. 2012, 45, 1129–1135. [Google Scholar] [CrossRef] [Green Version]
- Vaccaro, A.; Issa, A.-R.; Seugnet, L.; Birman, S.; Klarsfeld, A. Drosophila Clock Is Required in Brain Pacemaker Neurons to Prevent Premature Locomotor Aging Independently of Its Circadian Function. PLoS Genet. 2017, 13, e1006507. [Google Scholar] [CrossRef]
- Means, J.C.; Venkatesan, A.; Gerdes, B.; Fan, J.-Y.; Bjes, E.S.; Price, J.L. Drosophila Spaghetti and Doubletime Link the Circadian Clock and Light to Caspases, Apoptosis and Tauopathy. PLOS Genet. 2015, 11, e1005171. [Google Scholar] [CrossRef] [PubMed]
- Chong, F.P.; Ng, K.Y.; Koh, R.Y.; Chye, S.M. Tau Proteins and Tauopathies in Alzheimer’s Disease. Cell. Mol. Neurobiol. 2018, 38, 965–980. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Ayton, S.; Finkelstein, D.I.; Adlard, P.A.; Masters, C.L.; Bush, A.I. Tau protein: Relevance to Parkinson’s disease. Int. J. Biochem. Cell Biol. 2010, 42, 1775–1778. [Google Scholar] [CrossRef] [PubMed]
- Muskus, M.J.; Preuss, F.; Fan, J.-Y.; Bjes, E.S.; Price, J.L. Drosophila DBT Lacking Protein Kinase Activity Produces Long-Period and Arrhythmic Circadian Behavioral and Molecular Rhythms. Mol. Cell. Biol. 2007, 27, 8049–8064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, M.; Schultze, M.; Schuster, C.M.; Strauss, R. Circadian Plasticity in Photoreceptor Cells Controls Visual Coding Efficiency in Drosophila melanogaster. PLoS ONE 2010, 5, e9217. [Google Scholar] [CrossRef] [PubMed]
- Mazzotta, G.; Rossi, A.; Leonardi, E.; Mason, M.; Bertolucci, C.; Caccin, L.; Spolaore, B.; Martin, A.J.M.; Schlichting, M.; Grebler, R.; et al. Fly cryptochrome and the visual system. Proc. Natl. Acad. Sci. USA 2013, 110, 6163–6168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, R.J.H.; Furmston, R.; Williams, C.A.C.; Elliott, C.J.H. Neurophysiology of Drosophila Models of Parkinson’s Disease. Parkinson’s Dis. 2015, 2015, 1–11. [Google Scholar] [CrossRef]
- Afsari, F.; Christensen, K.V.; Smith, G.P.; Hentzer, M.; Nippe, O.M.; Elliott, C.J.H.; Wade, A.R. Abnormal visual gain control in a Parkinson’s disease model. Hum. Mol. Genet. 2014, 23, 4465–4478. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, H.; Brandt, M.D.; Klingelhoefer, L. The nonmotor features of Parkinson’s disease: Pathophysiology and management advances. Curr. Opin. Neurol. 2016, 29, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J.; Obeso, J.A.; Halliday, G.M. Selective neuronal vulnerability in Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 101–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seugnet, L.; Suzuki, Y.; Vine, L.; Gottschalk, L.; Shaw, P.J. D1 Receptor Activation in the Mushroom Bodies Rescues Sleep-Loss-Induced Learning Impairments in Drosophila. Curr. Biol. 2008, 18, 1110–1117. [Google Scholar] [CrossRef] [PubMed]
- Faville, R.; Kottler, B.; Goodhill, G.J.; Shaw, P.J.; van Swinderen, B. How deeply does your mutant sleep? Probing arousal to better understand sleep defects in Drosophila. Sci. Rep. 2015, 5, 8454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilestro, G.F. Video tracking and analysis of sleep in Drosophila melanogaster. Nat. Protoc. 2012, 7, 995–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Locomotor activity and organization of the circadian clock and dopaminergic system in the brain of D. melanogaster (A,B) Locomotor activity (mean ± SEM) of ~50 wild-type flies in (A) 12:12 light-dark (LD) and (B) constant darkness (DD) conditions, as a function of time. In (A) red arrows indicate the morning and evening anticipatory activities; ZT: Zeitgeber Time. In (B) CT: Circadian Time; (C) Schematic representation of the adult fly brain, in which the relative positions of the circadian neurons (right), the dopaminergic neurons (left), the mushroom bodies (MB), the central complex (CC), the Pars Intercerebralis (PI) are reported. Right hemisphere: lLNv: large ventral lateral neurons; sLNv: small LNvs; 5th sLNv: the fifth PDF-negative sLNv; LPN: lateral posterior neuron; LNd: dorsal LNs; DN1: dorsal neurons group 1; DN2: DN group 2; DN3: DN group 3; ey: relative position of the compound eye. Left hemisphere: PAM: Protocerebral Anterior Medial group; PAL: Protocerebral Anterior Lateral group; PPM1-4: Protocerebral Posterior Medial groups 1–4; PPL1–5: Protocerebral Posterior Lateral groups 1–5; VUM: Ventral Unpaired Median group; open circles in the left optic lobe indicate the TH positive cells of the medulla. (D) The two major TTLs of the circadian molecular clock in Drosophila. In the first Drosophila TTL, dCLK and dCYC form a dimer which binds the Enhancer boxes (E-boxes) in the promoter of the dper and dtim clock genes. dPER and dTIM proteins interact to form a complex, enter into the nucleus, and inhibit dCLK-dCYC activity. A second TTL modulates dClk expression: the dCLK-dCYC dimer induces the transcription of dvri and dPdp1 δ/ε genes. dVRI and dPDP1 δ/ε compete for the same element (D-box) in the dClk promoter, controlling dClk transcription. Post-translational modifications mediated by the kinases dDBT and dSGG modulate clock protein activities, regulating protein–protein interactions, nuclear translocation and degradation. In some clock neurons, light activates the internal photoreceptor dCRY which associates with dTIM and mediates its degradation. CLOCK: Circadian Locomotor Output Cycles Kaput; CRY: Cryptochrome; CYC: Cycle; PER: Period; TIM: Timeless; VRI: Vrille; PDP1: Par Domain Protein 1; DBT: Doubletime; SGG: Shaggy. Black arrows indicate the flow of the process; black sinusoidal lines and red arrows indicate transcriptional activity; blue arrows indicate the protein translation process (A,B) Graphs derived from data from our laboratory; (C,D) Modified from [54].
Figure 1.
Locomotor activity and organization of the circadian clock and dopaminergic system in the brain of D. melanogaster (A,B) Locomotor activity (mean ± SEM) of ~50 wild-type flies in (A) 12:12 light-dark (LD) and (B) constant darkness (DD) conditions, as a function of time. In (A) red arrows indicate the morning and evening anticipatory activities; ZT: Zeitgeber Time. In (B) CT: Circadian Time; (C) Schematic representation of the adult fly brain, in which the relative positions of the circadian neurons (right), the dopaminergic neurons (left), the mushroom bodies (MB), the central complex (CC), the Pars Intercerebralis (PI) are reported. Right hemisphere: lLNv: large ventral lateral neurons; sLNv: small LNvs; 5th sLNv: the fifth PDF-negative sLNv; LPN: lateral posterior neuron; LNd: dorsal LNs; DN1: dorsal neurons group 1; DN2: DN group 2; DN3: DN group 3; ey: relative position of the compound eye. Left hemisphere: PAM: Protocerebral Anterior Medial group; PAL: Protocerebral Anterior Lateral group; PPM1-4: Protocerebral Posterior Medial groups 1–4; PPL1–5: Protocerebral Posterior Lateral groups 1–5; VUM: Ventral Unpaired Median group; open circles in the left optic lobe indicate the TH positive cells of the medulla. (D) The two major TTLs of the circadian molecular clock in Drosophila. In the first Drosophila TTL, dCLK and dCYC form a dimer which binds the Enhancer boxes (E-boxes) in the promoter of the dper and dtim clock genes. dPER and dTIM proteins interact to form a complex, enter into the nucleus, and inhibit dCLK-dCYC activity. A second TTL modulates dClk expression: the dCLK-dCYC dimer induces the transcription of dvri and dPdp1 δ/ε genes. dVRI and dPDP1 δ/ε compete for the same element (D-box) in the dClk promoter, controlling dClk transcription. Post-translational modifications mediated by the kinases dDBT and dSGG modulate clock protein activities, regulating protein–protein interactions, nuclear translocation and degradation. In some clock neurons, light activates the internal photoreceptor dCRY which associates with dTIM and mediates its degradation. CLOCK: Circadian Locomotor Output Cycles Kaput; CRY: Cryptochrome; CYC: Cycle; PER: Period; TIM: Timeless; VRI: Vrille; PDP1: Par Domain Protein 1; DBT: Doubletime; SGG: Shaggy. Black arrows indicate the flow of the process; black sinusoidal lines and red arrows indicate transcriptional activity; blue arrows indicate the protein translation process (A,B) Graphs derived from data from our laboratory; (C,D) Modified from [54].

Figure 2.
Locomotor activity in 12:12 LD and circadian periodicity in DD conditions of young (3–5-day-old) and old (30-day-old) ddj-1α−; ddj-1β− double knock-out (DKO) males. ddj-1α−; ddj-1β− DKO and w1118 control flies were reared at 23 °C in 12:12 LD conditions (light intensity ~800 lux). 3–5 or ~30-day-old adult males were transferred into vials (~30 individuals each) containing normal or 1 mM Paraquat (PQ)-supplemented medium for 4 days. Subsequently, individuals were singly placed in glass tubes, containing normal or oxidative stress-inducing food. Locomotor activity was recorded as 1 min bins for 10 days in 12:12 LD conditions, or for 3 days in 12:12 LD followed by 7 days in DD regimes, using the Trikinetics Activity Monitor (DAM2). Circadian periodicity was analyzed as described in [124]. At least 3 replicates for each experiment were performed. (A–D) Locomotor activity profiles (mean ± SEM) of 50 young w1118 (A), 61 young ddj-1α−; ddj-1β− DKO (B), 38 old w1118 (C), 79 old ddj-1α−; ddj-1β− DKO (D) males for 3 days in 12:12 LD conditions. (E) Free-running periodicity of young and old ddj-1α−; ddj-1β− DKO and w1118 control flies in normal or mild oxidative stress conditions (1 mM PQ). N: number of analyzed flies; %R: percentage of rhythmic flies; tau: periodicity in DD. (F) Survival analysis to evaluate the effectiveness of the chronic exposure to 1 mM PQ on vitality in young flies. Only PQ-treated ddj-1α−; ddj-1β− DKO males showed a significant decrease in vitality (***: p < 0.0001 for all paired comparisons in Mantel-Cox test; for each genotype and treatment at least 90 individuals were monitored in 3 independent replicates).
Figure 2.
Locomotor activity in 12:12 LD and circadian periodicity in DD conditions of young (3–5-day-old) and old (30-day-old) ddj-1α−; ddj-1β− double knock-out (DKO) males. ddj-1α−; ddj-1β− DKO and w1118 control flies were reared at 23 °C in 12:12 LD conditions (light intensity ~800 lux). 3–5 or ~30-day-old adult males were transferred into vials (~30 individuals each) containing normal or 1 mM Paraquat (PQ)-supplemented medium for 4 days. Subsequently, individuals were singly placed in glass tubes, containing normal or oxidative stress-inducing food. Locomotor activity was recorded as 1 min bins for 10 days in 12:12 LD conditions, or for 3 days in 12:12 LD followed by 7 days in DD regimes, using the Trikinetics Activity Monitor (DAM2). Circadian periodicity was analyzed as described in [124]. At least 3 replicates for each experiment were performed. (A–D) Locomotor activity profiles (mean ± SEM) of 50 young w1118 (A), 61 young ddj-1α−; ddj-1β− DKO (B), 38 old w1118 (C), 79 old ddj-1α−; ddj-1β− DKO (D) males for 3 days in 12:12 LD conditions. (E) Free-running periodicity of young and old ddj-1α−; ddj-1β− DKO and w1118 control flies in normal or mild oxidative stress conditions (1 mM PQ). N: number of analyzed flies; %R: percentage of rhythmic flies; tau: periodicity in DD. (F) Survival analysis to evaluate the effectiveness of the chronic exposure to 1 mM PQ on vitality in young flies. Only PQ-treated ddj-1α−; ddj-1β− DKO males showed a significant decrease in vitality (***: p < 0.0001 for all paired comparisons in Mantel-Cox test; for each genotype and treatment at least 90 individuals were monitored in 3 independent replicates).

Figure 3.
Evaluation of the relationship between PD and circadian clock alterations using D. melanogaster. Clock gene mutants (green) can be used to determine the possible presence of clock-related neurodegeneration, while circadian abnormalities might be evaluated in genetic PD models (red), at the behavioral, physiological, and molecular levels. Similar analyses could be performed to screen for modulating drugs (magenta), in order to ameliorate circadian alterations.
Figure 3.
Evaluation of the relationship between PD and circadian clock alterations using D. melanogaster. Clock gene mutants (green) can be used to determine the possible presence of clock-related neurodegeneration, while circadian abnormalities might be evaluated in genetic PD models (red), at the behavioral, physiological, and molecular levels. Similar analyses could be performed to screen for modulating drugs (magenta), in order to ameliorate circadian alterations.


{kind=link}
{kind=link}
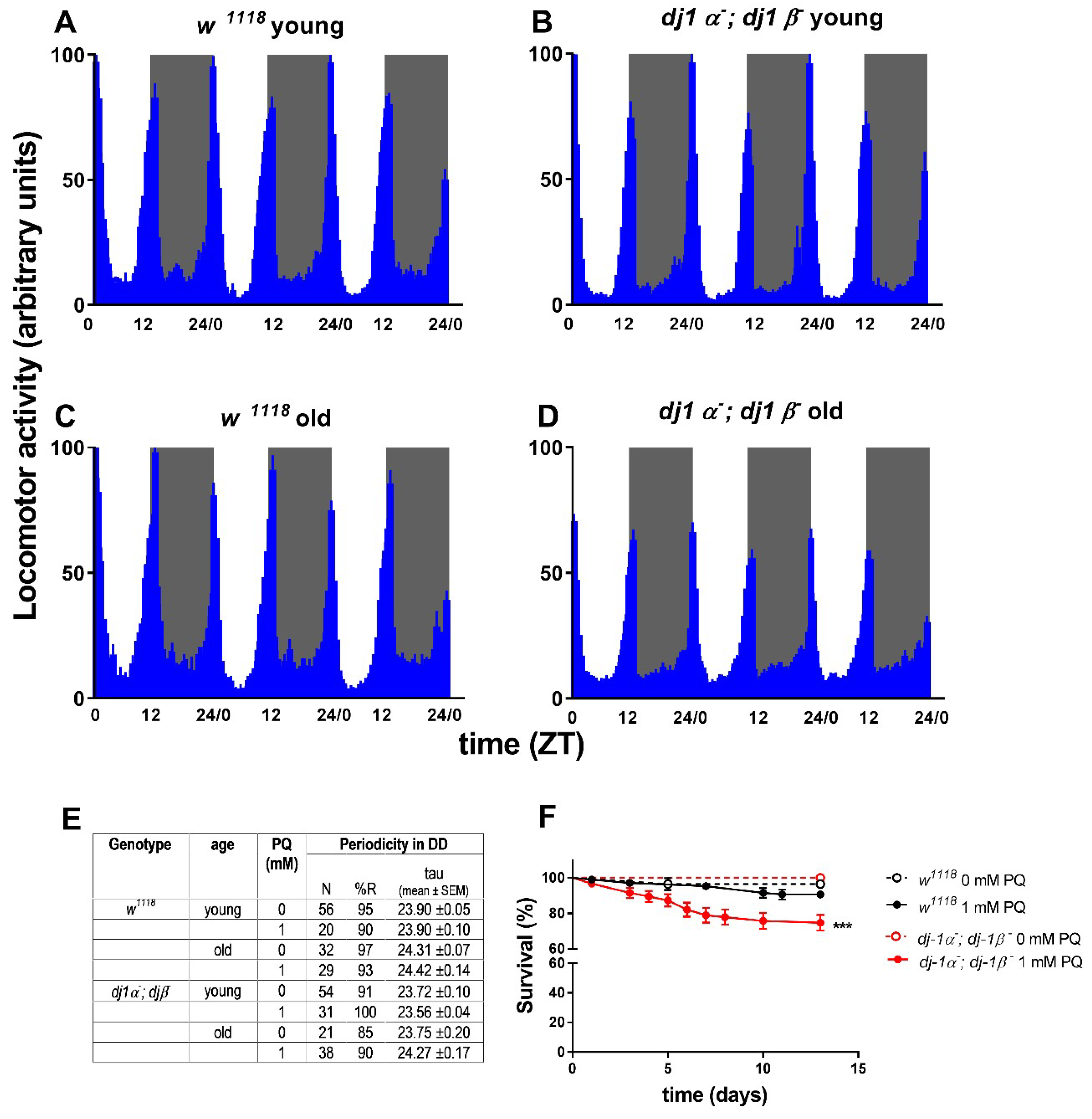{kind=link}
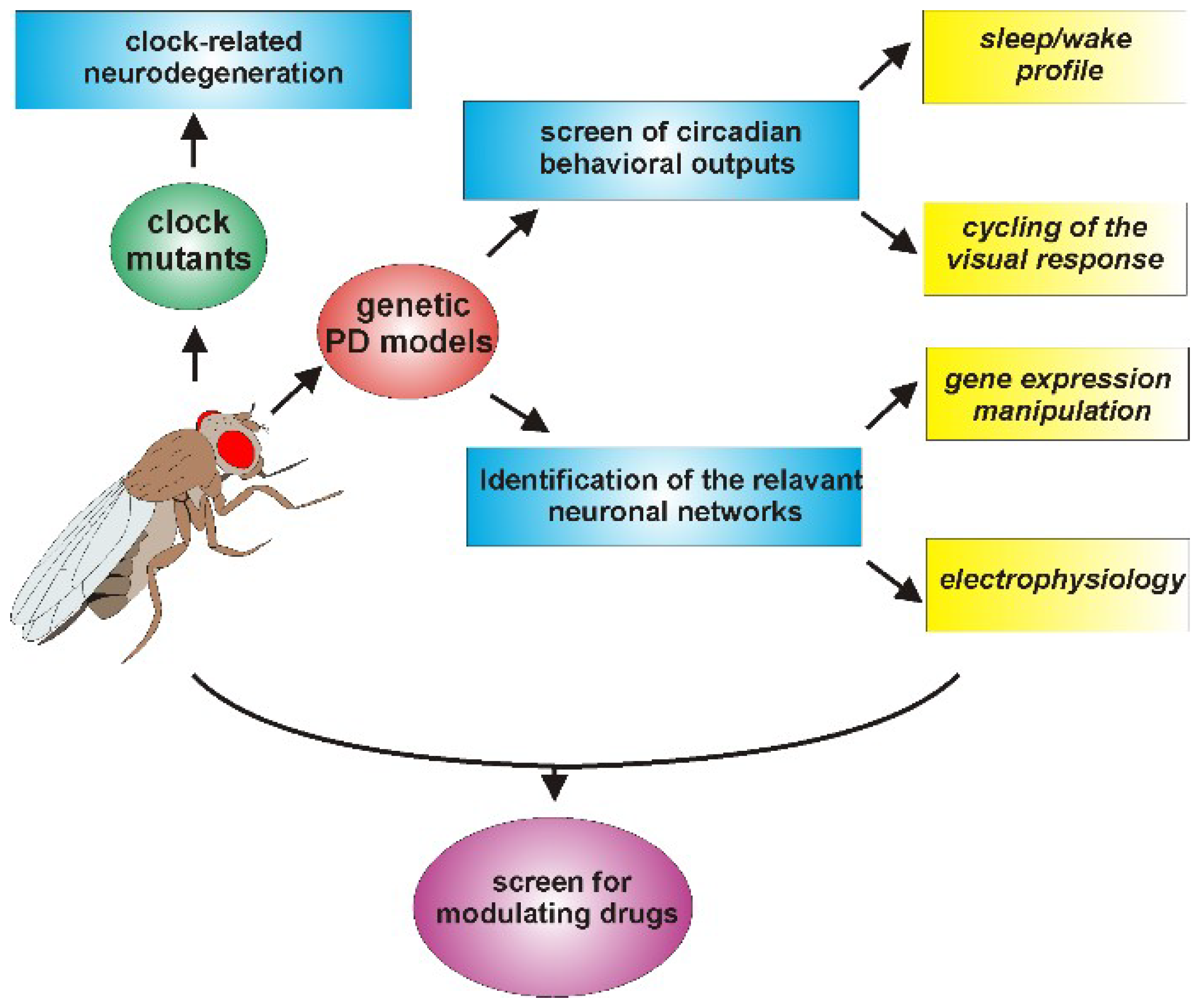{kind=link}
Table 1.
Circadian dysfunctions in mammalian Parkinson’s disease (PD) models.
| Mammalian Models | Intervention | Animal Model | Circadian Phenotype |
|---|---|---|---|
| ASO | Overexpression of wt α-syn in all brain regions | Mouse | Fragmented circadian rhythms and a reduced firing rate of SCN neurons during the day [31] |
| MitoPark | Deletion for mitochondrial transcription factor A in dopaminergic neurons | Mouse | Age-dependent rhythm decline and disturbed circadian activity rhythms under constant high light conditions [32] |
| MPTP-injection | Intraperitoneally and subcutaneously [33] Intraperitoneally [34] | Mouse | No significant changes in circadian parameters [33,34] |
| Not specified [35] | Mouse | Reduced amplitude in locomotor rhythm; altered clock gene expression in the SCN [35] | |
| Intravenously [36] | Dog | Circadian urine volume and vasopressin release alteration [36] | |
| Intramuscular [37] | Non-human Primates | Loss of circadian locomotor activity in the absence of light/dark cues [37] | |
| 6-OHDA-injection | Bilateral in the striatum [38] | Rat | Altered clock gene expression in the striatum [38] |
| Intracerebroventricular and unilateral infusion in medial forebrain bundle [39] | Rat | Disorganized wheel-running pattern in constant darkness and blunted PER2 expression rise in the dorsal striatum [39] | |
| Bilateral in the ventral tegmental area [40] | Rat | Reduced locomotor activity period in LD and longer activity rhythm periodicity under constant dim light [40] | |
| Bilateral in the striatum [41] | Rat | Decreased amplitude of the heart rate rhythm [41] | |
| Bilateral in the striatum [42] | Rat | Altered clock gene expression profile in SNC and in the striatum [42] |
Table 2.
Drosophila genetic PD models and associated circadian abnormalities.
| Fly Genetic PD Model | |||||
|---|---|---|---|---|---|
| Gene | Genetic Manipulation | PD Phenotype | Circadian Phenotype | ||
| Dopaminergic Cells Death | Protein Inclusions | Locomotor Deficit | |||
| hSNCA | wt-αS overexpression | Yes [99] | No [99] | Yes [99] | Altered locomotor activity profiles [96] |
| A53T overexpression | Yes [99] | Yes [99] | Yes [99] | n.d | |
| TP-αS (A30P, A56P, A76P) overexpression | Yes [96] | Yes [100] | Yes [100] | Circadian locomotor periodicity shift with aging [96] | |
| dPink1 | Loss of function | Yes [101] | n.d. | Yes [101] | Arrhythmic, hyperexcitability of l-LNvs neurons, day/night difference in RMP less pronounced [97] |
| Absence of circadian locomotor anticipatory activity in the morning in LD [98] | |||||
| dpark | Loss of function | Yes [102] | n.d. | Yes [102] | Weakly rhythmic, No difference in day/night SFR ratio [97] |
| Absence of circadian locomotor anticipatory activity in the morning in LD [98] | |||||
| ddj-1α | Loss of function | Yes [103] | n.d. | n.d. | n.d. |
| ddj-1β | Loss of function | No [104] | No [105] | Yes [105,106] | n.d. |
| ddj-1α; ddj-1β | Loss of function | No [107] | n.d. | Yes [108] | No evident abnormalities observed (this work) |
n.d: not determined; RMP: Resting Membrane Potential; SFR: Spontaneous Firing Rate.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
De Lazzari, F.; Bisaglia, M.; Zordan, M.A.; Sandrelli, F. Circadian Rhythm Abnormalities in Parkinson’s Disease from Humans to Flies and Back. Int. J. Mol. Sci. 2018, 19, 3911. https://doi.org/10.3390/ijms19123911
AMA Style
De Lazzari F, Bisaglia M, Zordan MA, Sandrelli F. Circadian Rhythm Abnormalities in Parkinson’s Disease from Humans to Flies and Back. International Journal of Molecular Sciences. 2018; 19(12):3911. https://doi.org/10.3390/ijms19123911
Chicago/Turabian StyleDe Lazzari, Federica, Marco Bisaglia, Mauro Agostino Zordan, and Federica Sandrelli. 2018. "Circadian Rhythm Abnormalities in Parkinson’s Disease from Humans to Flies and Back" International Journal of Molecular Sciences 19, no. 12: 3911. https://doi.org/10.3390/ijms19123911
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.