Direct Single-Molecule Observation of Sequential DNA Bending Transitions by the Sox2 HMG Box

 , and
, and
Abstract
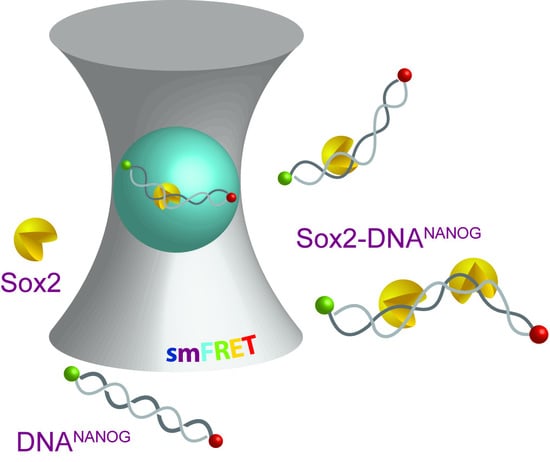
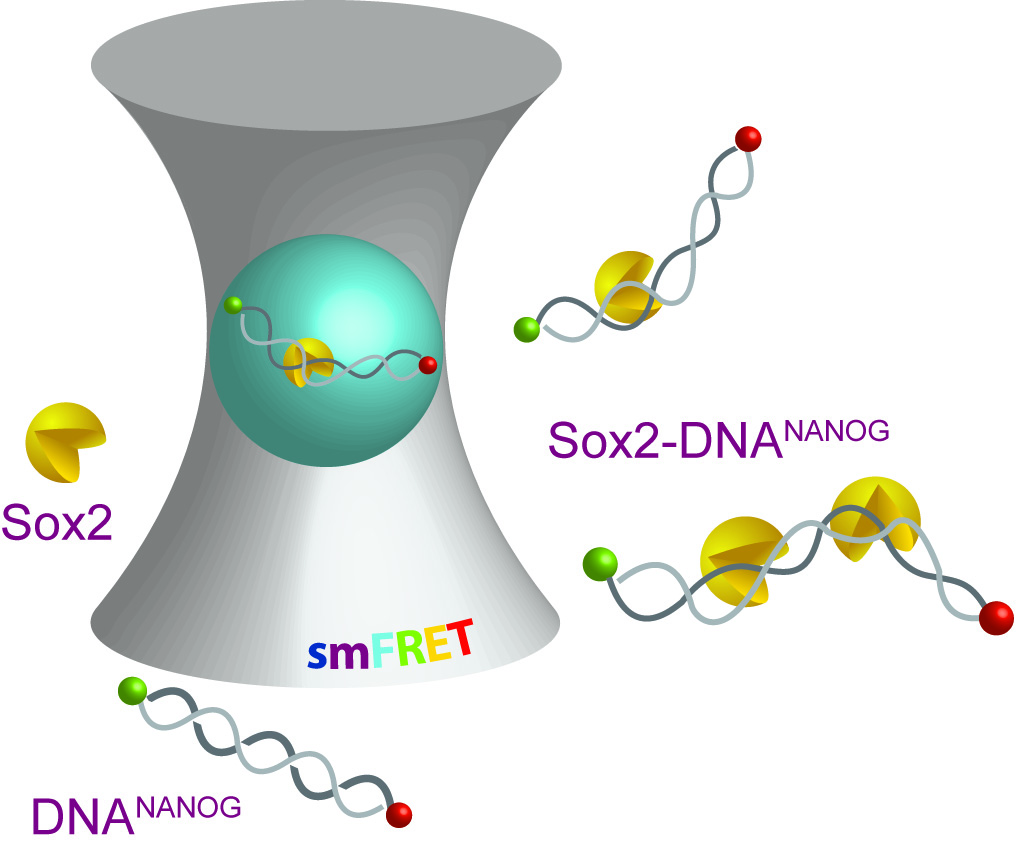
:
{kind=link}
{kind=link}
{kind=link}
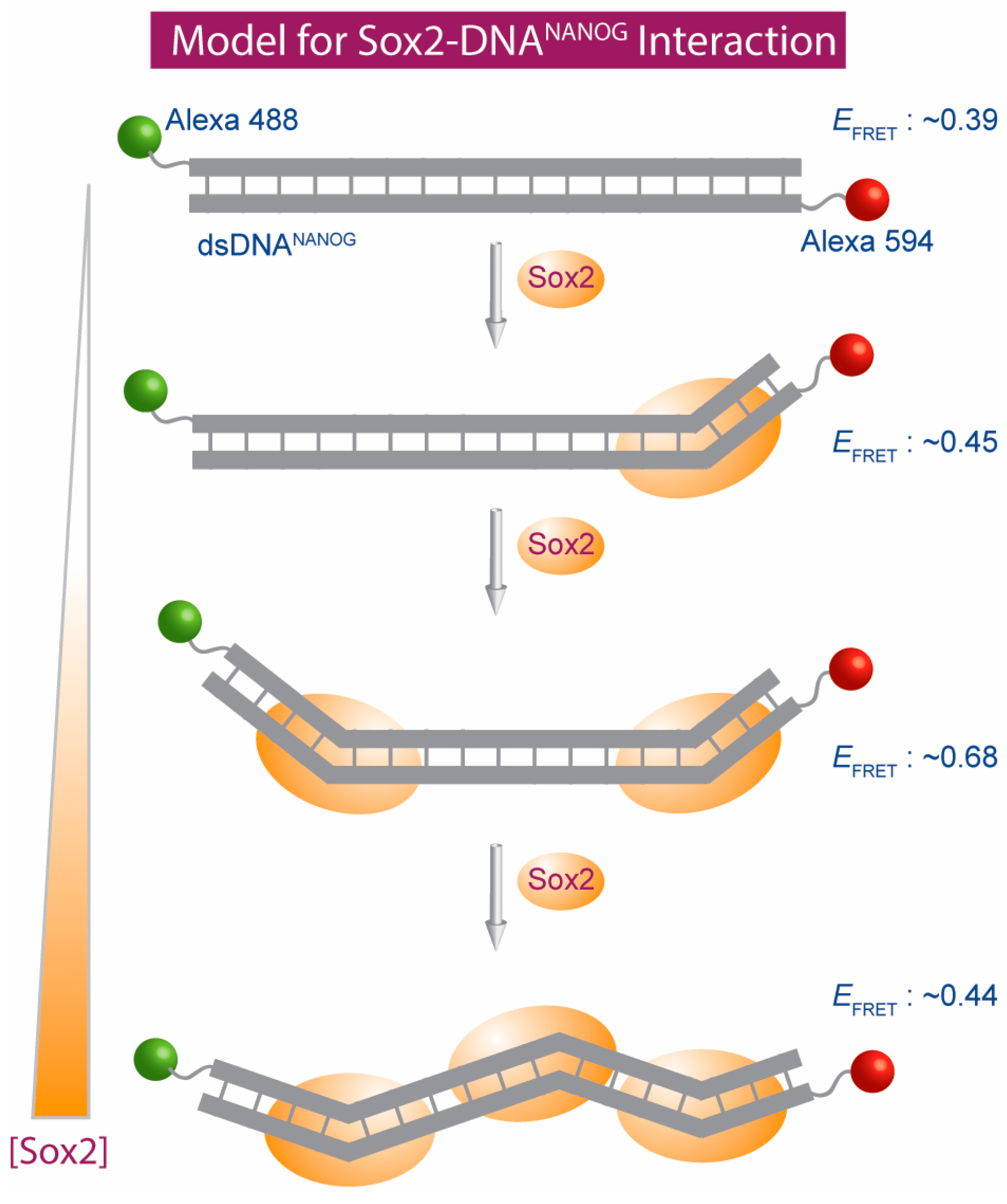{kind=link}
1. Introduction
2. Results
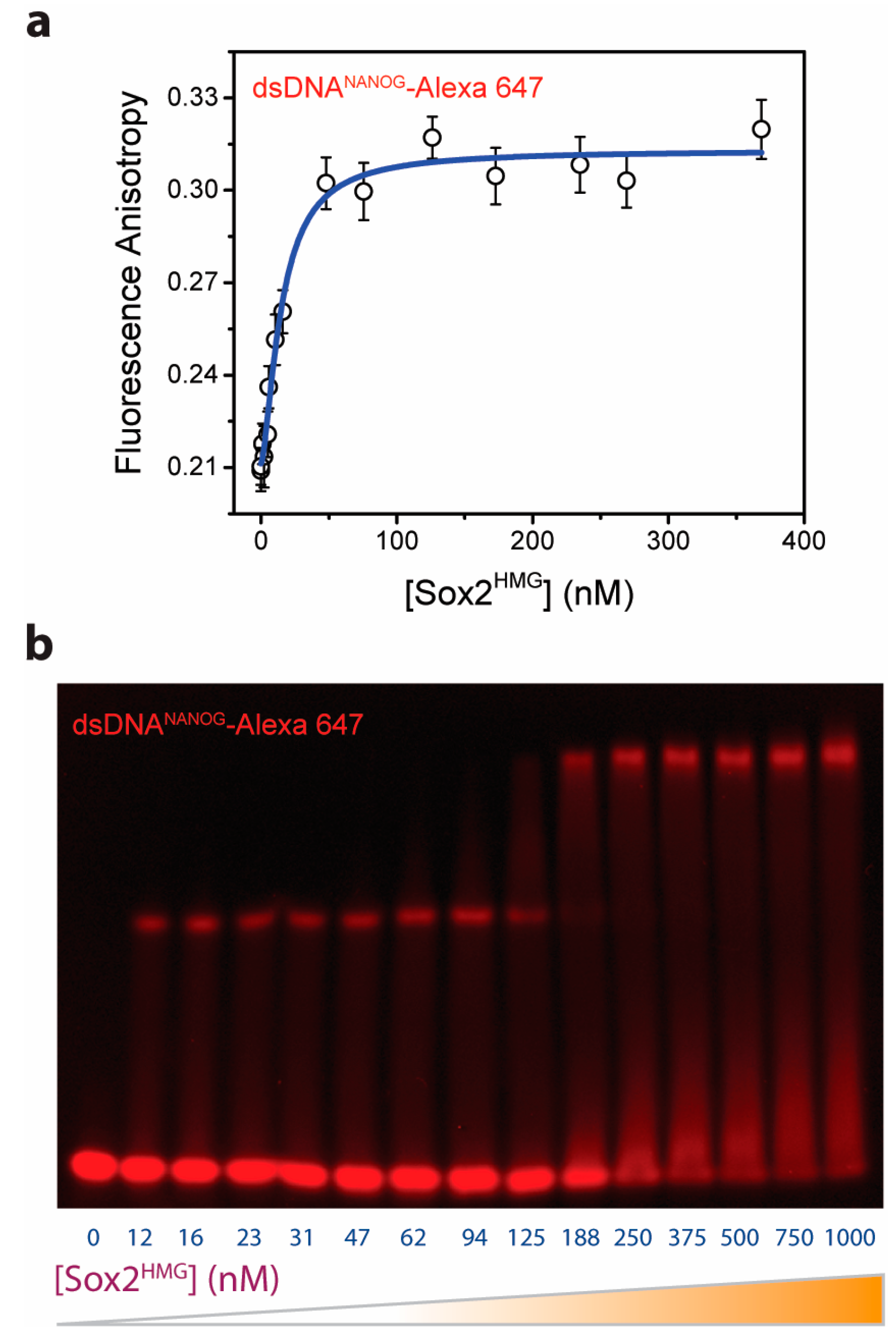
2.1. Multiple Sox2HMG Domains Cooperatively Interact with dsDNANANOG
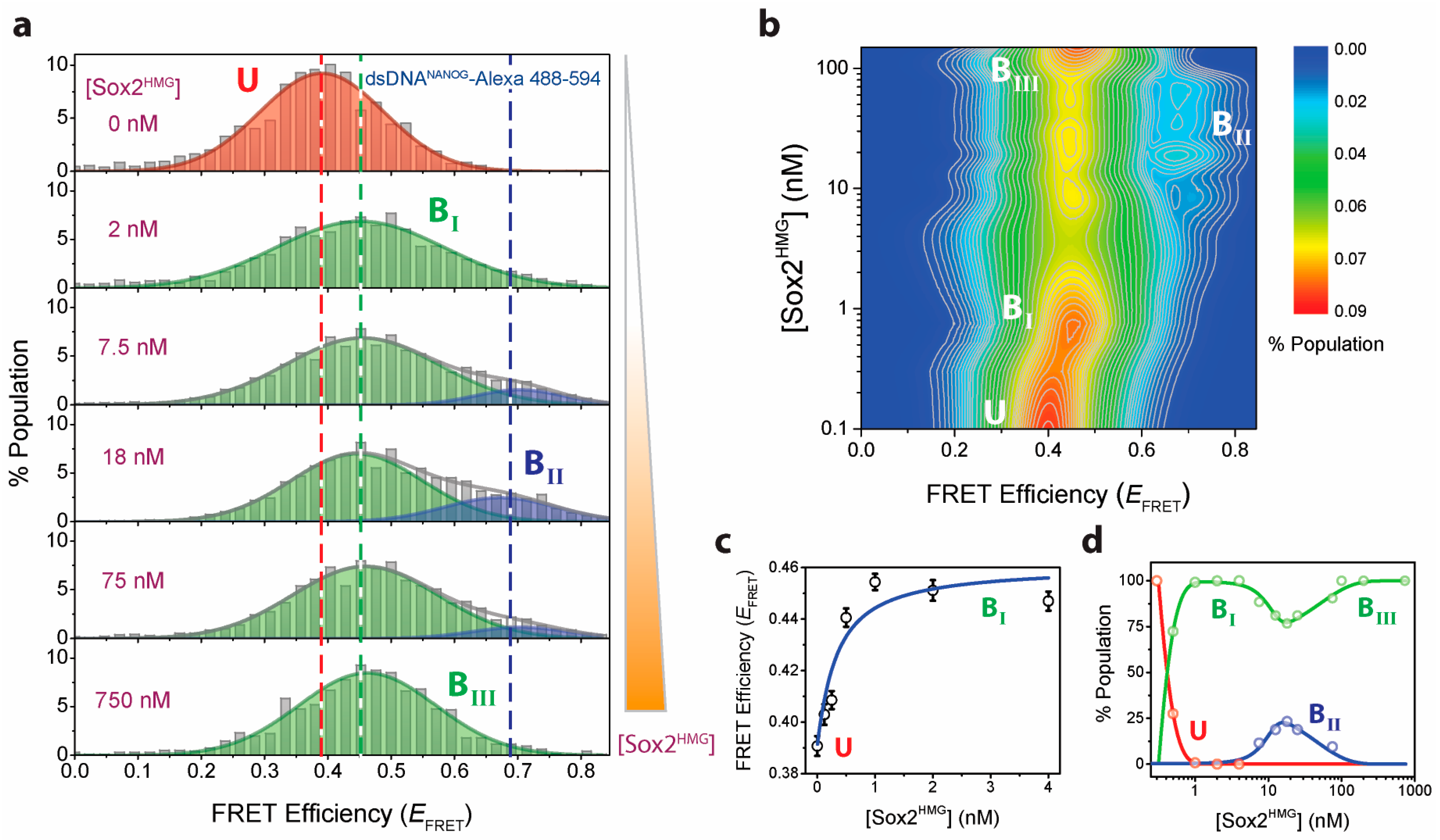
2.2. Sox2HMG Induces Sequential dsDNANANOG Bending Transitions
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| fEMSA | fluorescence electrophoretic mobility shift assay |
| FGF | fibroblast growth factor |
| HMG | high mobility group |
| NLS | nonlinear least squares |
| smFRET | single-molecule Förster resonance energy transfer |
| TF | transcription factor |
References
- Chew, L.J.; Gallo, V. The Yin and Yang of Sox proteins: Activation and repression in development and disease. J. Neurosci. Res. 2009, 87, 3277–3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Cui, W. Sox2, a key factor in the regulation of pluripotency and neural differentiation. World J. Stem Cells 2014, 6, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.R.; Laghari, Z.A.; Novoa, C.A.; Hughes, J.; Webster, J.R.; Goodwin, P.E.; Wheatley, S.P.; Scotting, P.J. Sox2 acts as a transcriptional repressor in neural stem cells. BMC Neurosci. 2014, 15, 95. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, H.; Kamachi, Y. SOX-partner code for cell specification: Regulatory target selection and underlying molecular mechanisms. Int. J. Biochem. Cell Biol. 2010, 42, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, H.; Kamachi, Y. Chapter 8—SOX2–Partner Factor Interactions and Enhancer Regulation. In Sox2; Academic Press: Boston, MA, USA, 2016; pp. 131–144. [Google Scholar]
- Thomson, M.; Liu, S.J.; Zou, L.N.; Smith, Z.; Meissner, A.; Ramanathan, S. Pluripotency factors in embryonic stem cells regulate differentiation into germ layers. Cell 2011, 145, 875–889. [Google Scholar] [CrossRef] [PubMed]
- Soufi, A.; Donahue, G.; Zaret, K.S. Facilitators and impediments of the pluripotency reprogramming factors’ initial engagement with the genome. Cell 2012, 151, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Z.; Li, L.; Chen, B.C.; Revyakin, A.; Hajj, B.; Legant, W.; Dahan, M.; Lionnet, T.; Betzig, E.; Tjian, R. Single-molecule dynamics of enhanceosome assembly in embryonic stem cells. Cell 2014, 156, 1274–1285. [Google Scholar] [CrossRef]
- Soufi, A.; Garcia, M.F.; Jaroszewicz, A.; Osman, N.; Pellegrini, M.; Zaret, K.S. Pioneer transcription factors target partial DNA motifs on nucleosomes to initiate reprogramming. Cell 2015, 161, 555–568. [Google Scholar] [CrossRef]
- White, M.D.; Angiolini, J.F.; Alvarez, Y.D.; Kaur, G.; Zhao, Z.W.; Mocskos, E.; Bruno, L.; Bissiere, S.; Levi, V.; Plachta, N. Long-Lived Binding of Sox2 to DNA Predicts Cell Fate in the Four-Cell Mouse Embryo. Cell 2016, 165, 75–87. [Google Scholar] [CrossRef]
- Hou, L.; Srivastava, Y.; Jauch, R. Molecular basis for the genome engagement by Sox proteins. Semin. Cell Dev. Biol. 2017, 63, 2–12. [Google Scholar] [CrossRef]
- Soullier, S.; Jay, P.; Poulat, F.; Vanacker, J.M.; Berta, P.; Laudet, V. Diversification pattern of the HMG and SOX family members during evolution. J. Mol. Evol. 1999, 48, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Malarkey, C.S.; Churchill, M.E. The high mobility group box: The ultimate utility player of a cell. Trends Biochem. Sci. 2012, 37, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.A. Floppy SOX: Mutual induced fit in hmg (high-mobility group) box-DNA recognition. Mol. Endocrinol. 2001, 15, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Schlierf, B.; Ludwig, A.; Klenovsek, K.; Wegner, M. Cooperative binding of Sox10 to DNA: Requirements and consequences. Nucleic Acids Res. 2002, 30, 5509–5516. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.K.; Li, N.X.; Chee, S.; Prabhakar, S.; Kolatkar, P.R.; Jauch, R. Deciphering the Sox-Oct partner code by quantitative cooperativity measurements. Nucleic Acids Res. 2012, 40, 4933–4941. [Google Scholar] [CrossRef] [PubMed]
- Clore, G.M. Chapter 3—Dynamics of SOX2 Interactions with DNA A2—Kondoh, Hisato. In Sox2; Lovell-Badge, R., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 25–41. [Google Scholar]
- Scaffidi, P.; Bianchi, M.E. Spatially precise DNA bending is an essential activity of the sox2 transcription factor. J. Biol. Chem. 2001, 276, 47296–47302. [Google Scholar] [CrossRef] [PubMed]
- Dragan, A.I.; Read, C.M.; Makeyeva, E.N.; Milgotina, E.I.; Churchill, M.E.; Crane-Robinson, C.; Privalov, P.L. DNA binding and bending by HMG boxes: Energetic determinants of specificity. J. Mol. Biol. 2004, 343, 371–393. [Google Scholar] [CrossRef]
- Slattery, M.; Riley, T.; Liu, P.; Abe, N.; Gomez-Alcala, P.; Dror, I.; Zhou, T.; Rohs, R.; Honig, B.; Bussemaker, H.J.; Mann, R.S. Cofactor binding evokes latent differences in DNA binding specificity between Hox proteins. Cell 2011, 147, 1270–1282. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, G.; Cui, K.; Liu, Y.; Xu, S.; Pei, D. A dominant-negative form of mouse SOX2 induces trophectoderm differentiation and progressive polyploidy in mouse embryonic stem cells. J. Biol. Chem. 2007, 282, 19481–19492. [Google Scholar] [CrossRef]
- Cox, J.L.; Mallanna, S.K.; Luo, X.; Rizzino, A. Sox2 uses multiple domains to associate with proteins present in Sox2-protein complexes. PLoS ONE 2010, 5, e15486. [Google Scholar] [CrossRef]
- Xia, P.; Wang, S.; Ye, B.; Du, Y.; Huang, G.; Zhu, P.; Fan, Z. Sox2 functions as a sequence-specific DNA sensor in neutrophils to initiate innate immunity against microbial infection. Nat. Immunol. 2015, 16, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.N. The Hill equation revisited: Uses and misuses. FASEB J. 1997, 11, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Hellman, L.M.; Fried, M.G. Electrophoretic mobility shift assay (EMSA) for detecting protein-nucleic acid interactions. Nat. Protoc. 2007, 2, 1849–1861. [Google Scholar] [CrossRef] [PubMed]
- Ferreon, A.C.; Deniz, A.A. Protein folding at single-molecule resolution. Biochim. Biophys. Acta 2011, 1814, 1021–1029. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.D.; Crick, F.H.C. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid. Nature 1953, 171, 737. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, P.S.; Choi, K.J.; Leonard, P.G.; Sizovs, A.; Moosa, M.M.; MacKenzie, K.R.; Ferreon, J.C.; Ferreon, A.C. The N-Terminal Domain of ALS-Linked TDP-43 Assembles without Misfolding. Angew. Chem. Int. Ed. Engl. 2017, 56, 12590–12593. [Google Scholar] [CrossRef] [PubMed]
- Deniz, A.A.; Dahan, M.; Grunwell, J.R.; Ha, T.; Faulhaber, A.E.; Chemla, D.S.; Weiss, S.; Schultz, P.G. Single-pair fluorescence resonance energy transfer on freely diffusing molecules: Observation of Forster distance dependence and subpopulations. Proc. Natl. Acad. Sci. USA 1999, 96, 3670–3675. [Google Scholar] [CrossRef] [Green Version]
- Dey, S.K.; Pettersson, J.R.; Topacio, A.Z.; Das, S.R.; Peteanu, L.A. Eliminating Spurious Zero-Efficiency FRET States in Diffusion-Based Single-Molecule Confocal Microscopy. J. Phys. Chem. Lett. 2018, 9, 2259–2265. [Google Scholar] [CrossRef]
- Johnson, I.D.; Spence, M.T.Z. The Molecular Probes Handbook: A Guide to Fluorescent Probes and Labeling Technologies; Molecular Probes: Eugene, OR, USA, 2010. [Google Scholar]
- Pljevaljcic, G.; Millar, D.P.; Deniz, A.A. Freely diffusing single hairpin ribozymes provide insights into the role of secondary structure and partially folded states in RNA folding. Biophys. J. 2004, 87, 457–467. [Google Scholar] [CrossRef]
- Morgan, M.A.; Okamoto, K.; Kahn, J.D.; English, D.S. Single-molecule spectroscopic determination of lac repressor-DNA loop conformation. Biophys. J. 2005, 89, 2588–2596. [Google Scholar] [CrossRef]
- Schuler, B. Single-molecule FRET of protein structure and dynamics—A primer. J. Nanobiotechnol. 2013, 11 (Suppl. 1), S2. [Google Scholar] [CrossRef]
- Tyagi, S.; VanDelinder, V.; Banterle, N.; Fuertes, G.; Milles, S.; Agez, M.; Lemke, E.A. Continuous throughput and long-term observation of single-molecule FRET without immobilization. Nat. Methods 2014, 11, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Ferreon, A.C.; Gambin, Y.; Lemke, E.A.; Deniz, A.A. Interplay of α-synuclein binding and conformational switching probed by single-molecule fluorescence. Proc. Natl. Acad. Sci. USA 2009, 106, 5645–5650. [Google Scholar] [CrossRef] [PubMed]
- Ferreon, A.C.; Moran, C.R.; Ferreon, J.C.; Deniz, A.A. Alteration of the α-synuclein folding landscape by a mutation related to Parkinson’s disease. Angew. Chem. Int. Ed. Engl. 2010, 49, 3469–3472. [Google Scholar] [CrossRef]
- Gambin, Y.; VanDelinder, V.; Ferreon, A.C.; Lemke, E.A.; Groisman, A.; Deniz, A.A. Visualizing a one-way protein encounter complex by ultrafast single-molecule mixing. Nat. Methods 2011, 8, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Moosa, M.M.; Ferreon, A.C.; Deniz, A.A. Forced folding of a disordered protein accesses an alternative folding landscape. Chemphyschem 2015, 16, 90–94. [Google Scholar] [CrossRef]
- Su, W.; Jackson, S.; Tjian, R.; Echols, H. DNA looping between sites for transcriptional activation: Self-association of DNA-bound Sp1. Genes Dev. 1991, 5, 820–826. [Google Scholar] [CrossRef]
- Lim, F.L.; Hayes, A.; West, A.G.; Pic-Taylor, A.; Darieva, Z.; Morgan, B.A.; Oliver, S.G.; Sharrocks, A.D. Mcm1p-induced DNA bending regulates the formation of ternary transcription factor complexes. Mol. Cell Biol. 2003, 23, 450–461. [Google Scholar] [CrossRef]
- Petrascheck, M.; Escher, D.; Mahmoudi, T.; Verrijzer, C.P.; Schaffner, W.; Barberis, A. DNA looping induced by a transcriptional enhancer in vivo. Nucleic Acids Res. 2005, 33, 3743–3750. [Google Scholar] [CrossRef] [Green Version]
- Whittington, J.E.; Delgadillo, R.F.; Attebury, T.J.; Parkhurst, L.K.; Daugherty, M.A.; Parkhurst, L.J. TATA-binding protein recognition and bending of a consensus promoter are protein species dependent. Biochemistry 2008, 47, 7264–7273. [Google Scholar] [CrossRef]
- Gietl, A.; Grohmann, D. Modern biophysical approaches probe transcription-factor-induced DNA bending and looping. Biochem. Soc. Trans. 2013, 41, 368–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.C.; Cai, M., Jr.; Clore, G.M. Molecular basis for synergistic transcriptional activation by Oct1 and Sox2 revealed from the solution structure of the 42-kDa Oct1.Sox2.Hoxb1-DNA ternary transcription factor complex. J. Biol. Chem. 2004, 279, 1449–1457. [Google Scholar] [CrossRef]
- Palasingam, P.; Jauch, R.; Ng, C.K.; Kolatkar, P.R. The structure of Sox17 bound to DNA reveals a conserved bending topology but selective protein interaction platforms. J. Mol. Biol. 2009, 388, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Rodda, D.J.; Chew, J.L.; Lim, L.H.; Loh, Y.H.; Wang, B.; Ng, H.H.; Robson, P. Transcriptional regulation of nanog by OCT4 and SOX2. J. Biol. Chem. 2005, 280, 24731–24737. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Hirano, K.; Nagata, S.; Tada, T. Sox2 expression effects on direct reprogramming efficiency as determined by alternative somatic cell fate. Stem Cell Res. 2011, 6, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, N. Chapter 4—Posttranscriptional Modulation of Sox2 Activity by miRNAs A2—Kondoh, Hisato. In Sox2; Lovell-Badge, R., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 43–71. [Google Scholar]
- Bertolini, J.; Mercurio, S.; Favaro, R.; Mariani, J.; Ottolenghi, S.; Nicolis, S.K. Chapter 11—Sox2-Dependent Regulation of Neural Stem Cells and CNS Development A2—Kondoh, Hisato. In Sox2; Lovell-Badge, R., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 187–216. [Google Scholar]
- Van Heyningen, V. Chapter 13—Congenital Abnormalities and SOX2 Mutations A2—Kondoh, Hisato. In Sox2; Lovell-Badge, R., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 235–242. [Google Scholar]
- Rizzoti, K.; Lovell-Badge, R. Chapter 14—Role of SOX2 in the Hypothalamo–Pituitary Axis. In Sox2; Academic Press: Boston, MA, USA, 2016; pp. 243–262. [Google Scholar]
- Iwafuchi-Doi, M.; Zaret, K.S. Cell fate control by pioneer transcription factors. Development 2016, 143, 1833–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuebben, E.L.; Rizzino, A. The dark side of SOX2: Cancer—A comprehensive overview. Oncotarget 2017, 8, 44917–44943. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Hochedlinger, K. The sox family of transcription factors: Versatile regulators of stem and progenitor cell fate. Cell Stem Cell 2013, 12, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Lin, B.; Zhao, M.; Yang, X.; Chen, M.; Gao, A.; Liu, F.; Que, J.; Lan, X. The multiple roles for Sox2 in stem cell maintenance and tumorigenesis. Cell Signal. 2013, 25, 1264–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamachi, Y.; Kondoh, H. Sox proteins: Regulators of cell fate specification and differentiation. Development 2013, 140, 4129–4144. [Google Scholar] [CrossRef]
- Hagey, D.W.; Muhr, J. Sox2 acts in a dose-dependent fashion to regulate proliferation of cortical progenitors. Cell Rep. 2014, 9, 1908–1920. [Google Scholar] [CrossRef] [PubMed]
- Peirano, R.I.; Wegner, M. The glial transcription factor Sox10 binds to DNA both as monomer and dimer with different functional consequences. Nucleic Acids Res. 2000, 28, 3047–3055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, Y.; Clore, G.M. Impact of protein/protein interactions on global intermolecular translocation rates of the transcription factors Sox2 and Oct1 between DNA cognate sites analyzed by z-exchange NMR spectroscopy. J. Biol. Chem. 2012, 287, 26962–26970. [Google Scholar] [CrossRef] [PubMed]
- Morimura, H.; Tanaka, S.I.; Ishitobi, H.; Mikami, T.; Kamachi, Y.; Kondoh, H.; Inouye, Y. Nano-analysis of DNA conformation changes induced by transcription factor complex binding using plasmonic nanodimers. ACS Nano 2013, 7, 10733–10740. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; De, D.; Hidaka, K.; Kim, K.K.; Endo, M.; Sugiyama, H. Single molecule visualization and characterization of Sox2-Pax6 complex formation on a regulatory DNA element using a DNA origami frame. Nano Lett. 2014, 14, 2286–2292. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moosa, M.M.; Tsoi, P.S.; Choi, K.-J.; Ferreon, A.C.M.; Ferreon, J.C. Direct Single-Molecule Observation of Sequential DNA Bending Transitions by the Sox2 HMG Box. Int. J. Mol. Sci. 2018, 19, 3865. https://doi.org/10.3390/ijms19123865
Moosa MM, Tsoi PS, Choi K-J, Ferreon ACM, Ferreon JC. Direct Single-Molecule Observation of Sequential DNA Bending Transitions by the Sox2 HMG Box. International Journal of Molecular Sciences. 2018; 19(12):3865. https://doi.org/10.3390/ijms19123865
Chicago/Turabian StyleMoosa, Mahdi Muhammad, Phoebe S. Tsoi, Kyoung-Jae Choi, Allan Chris M. Ferreon, and Josephine C. Ferreon. 2018. "Direct Single-Molecule Observation of Sequential DNA Bending Transitions by the Sox2 HMG Box" International Journal of Molecular Sciences 19, no. 12: 3865. https://doi.org/10.3390/ijms19123865