
Functional Selectivity and Antidepressant Activity of Serotonin 1A Receptor Ligands

Abstract
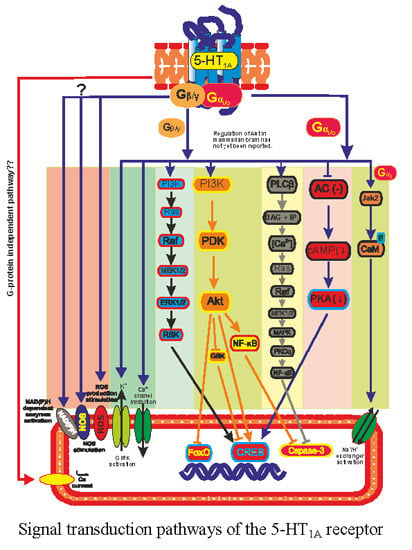
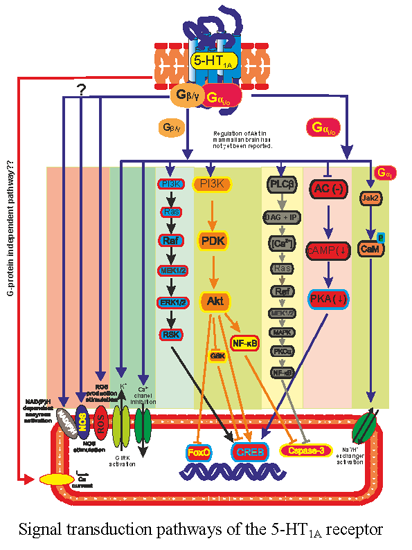
:
1. Introduction
- Low number of neurons: 250,000 5-HT neurons in the human brain (out of a total of 1011).
- Innervation of the whole neuraxis with extensive axon branching (4106 nerve terminals/mm3 in neocortex).
- Slow and regular discharge (pace maker neurons): strong homeostasis.
- Neuronal activity dependent on sleep-wake cycles (REM-off neurons).
- Very sensitive to self-inhibition through activation of 5-HT1A autoreceptors.
- Rich neurochemistry: 14 different postsynaptic receptors.
- Implication in a large number of physiological functions.
- Mutual control with monoaminergic cell groups.
2. 5-HT1A Receptor Distribution
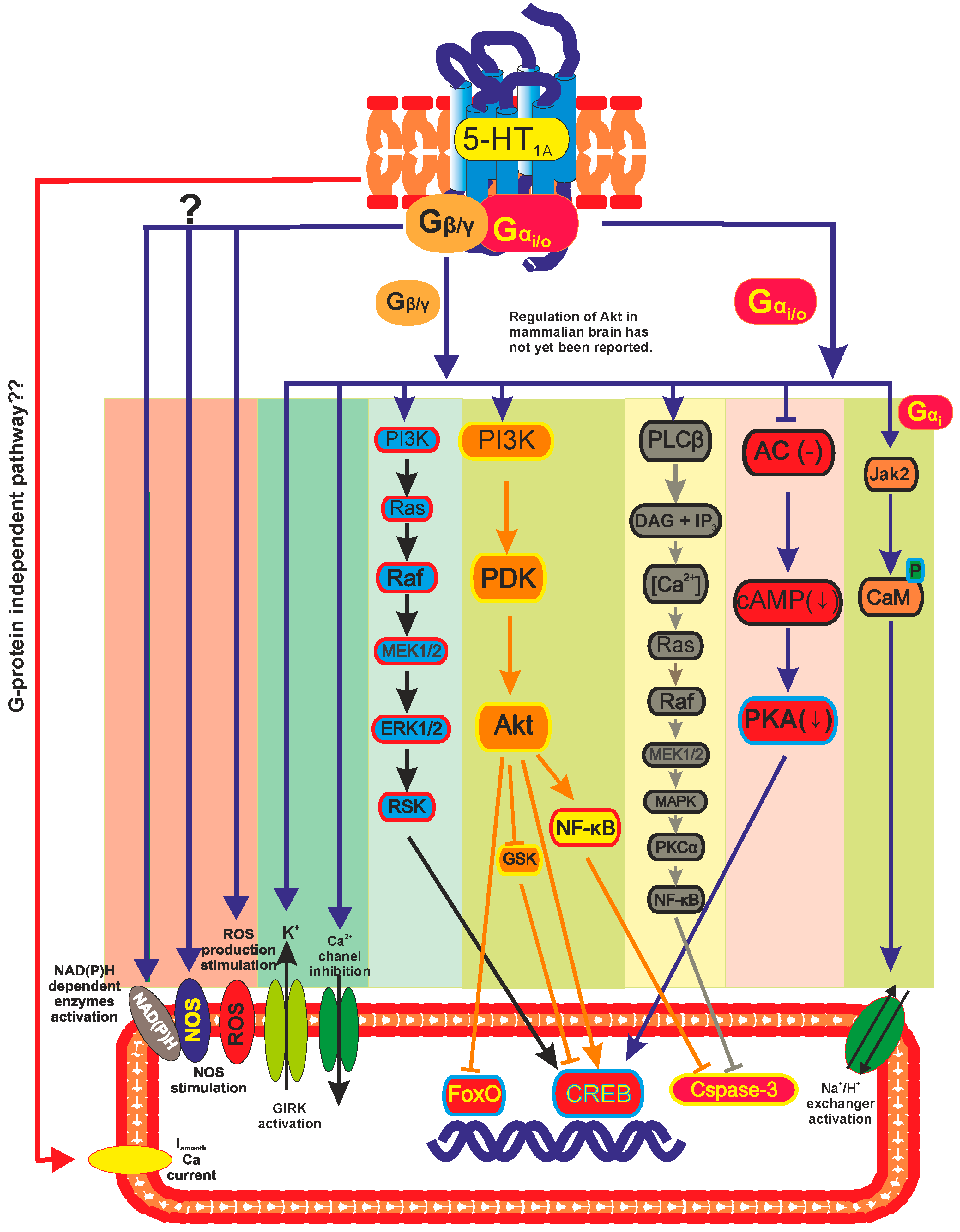
3. 5-HT1A Receptor Regulated Transcription Pathways
3.1. Adenylate Cyclase

3.2. GIRK and Calcium Channel
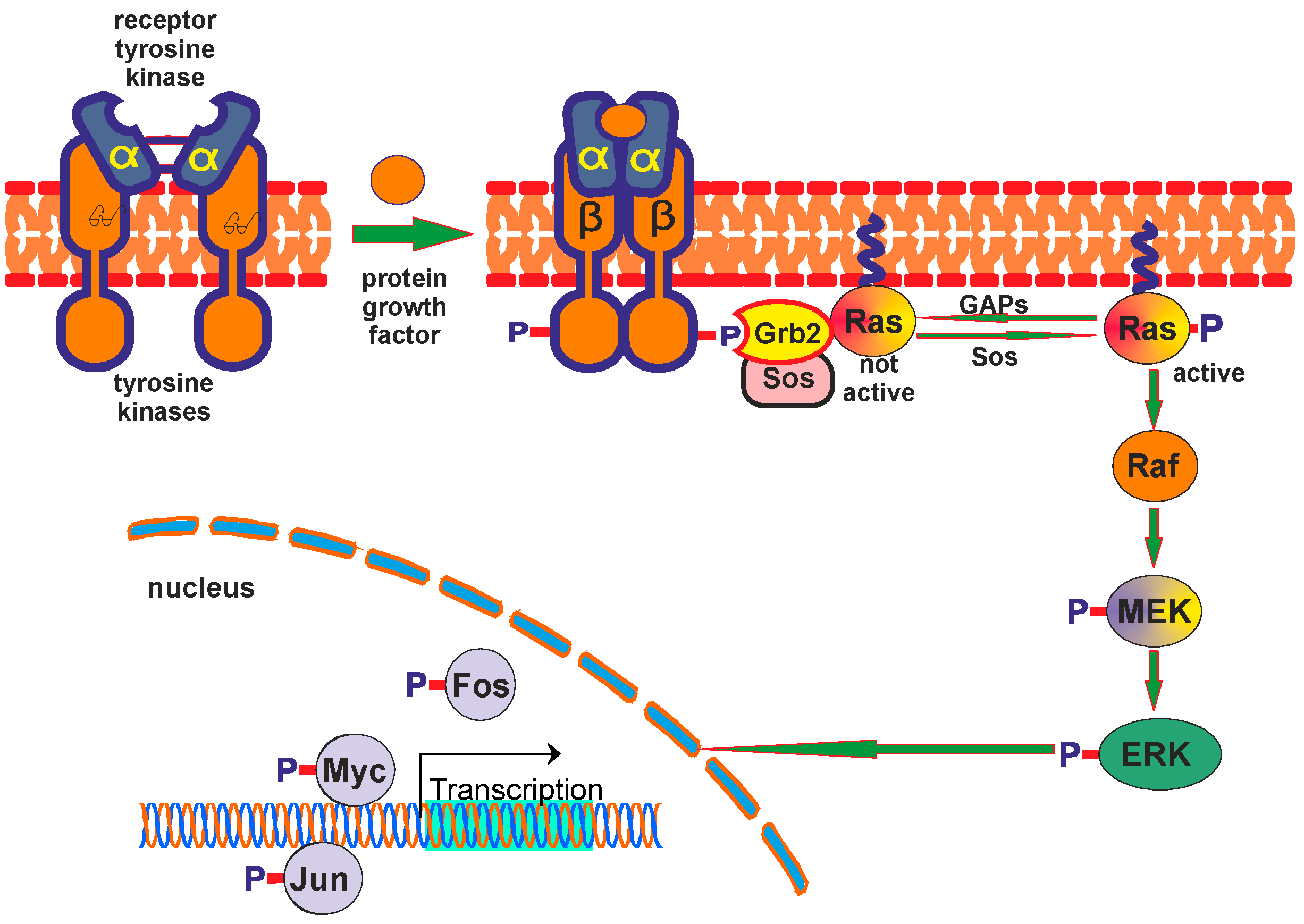
3.3. ERK/MAPK Pathway
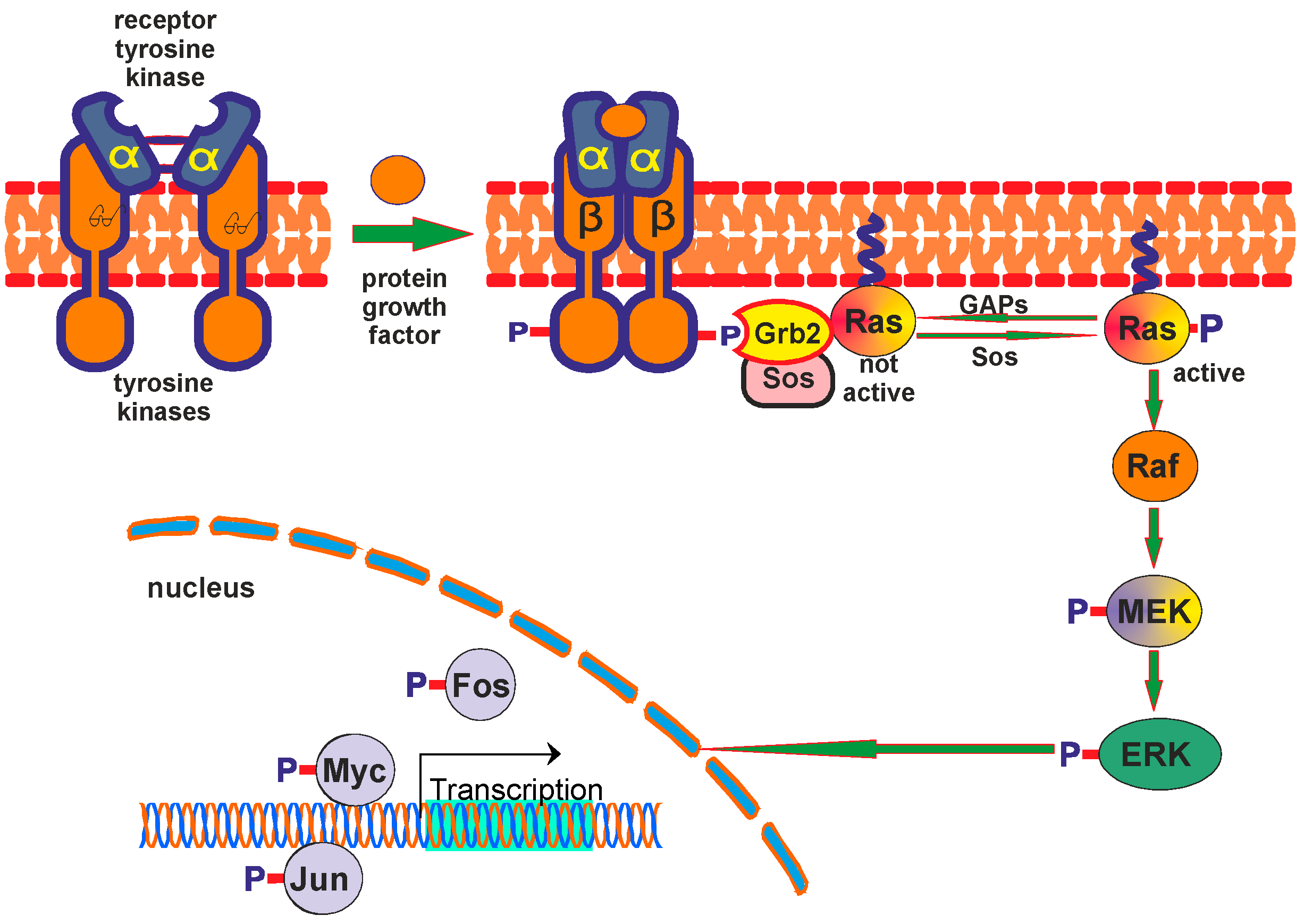
3.4. PI3K and Akt Pathway
3.5. Na+/H+ Exchangers
3.6. Nitric Oxide (NO) Production
3.7. Diversity of 5-HT1A Pathways
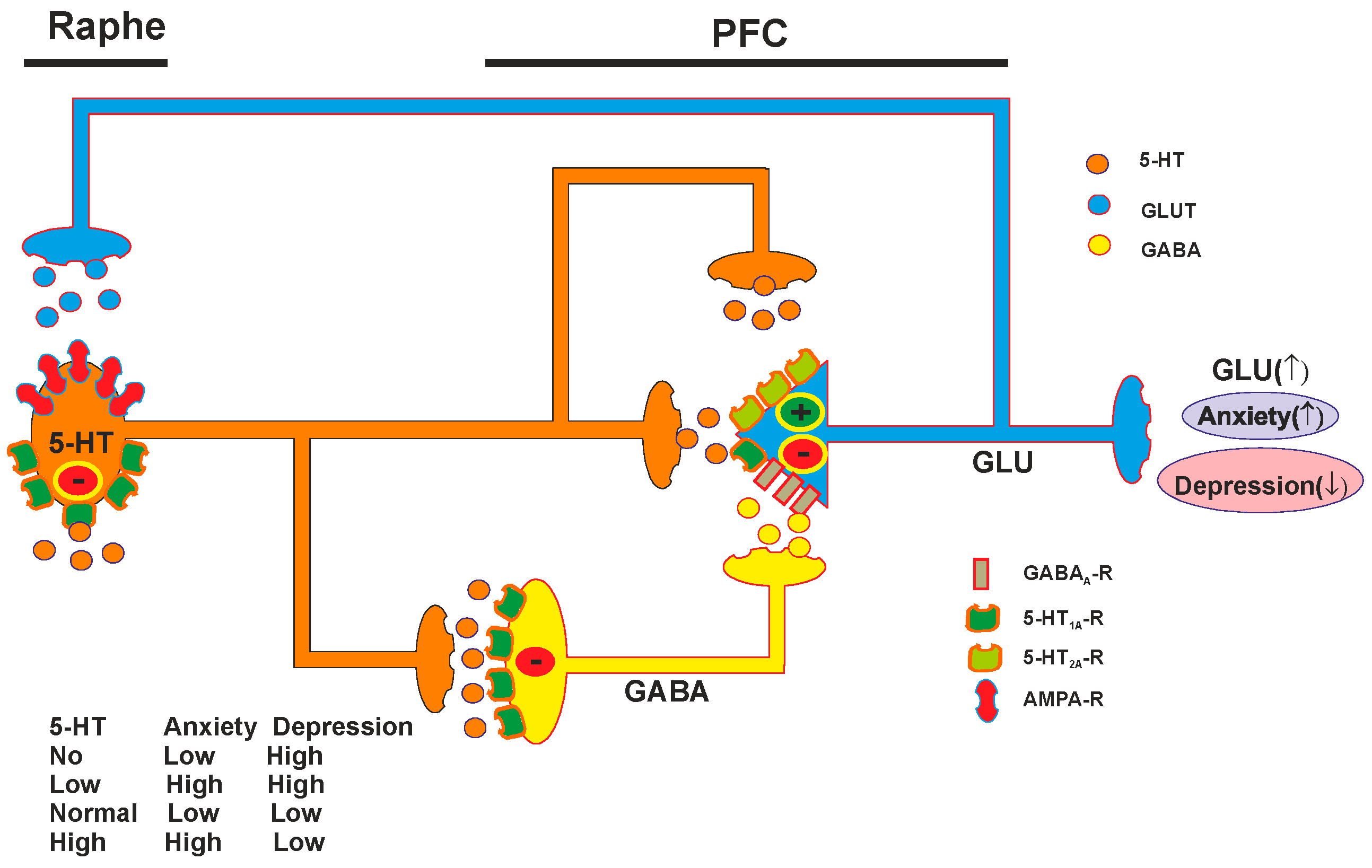
4. 5-HT1A Receptors and Depression
4.1. Serotonin and Depression
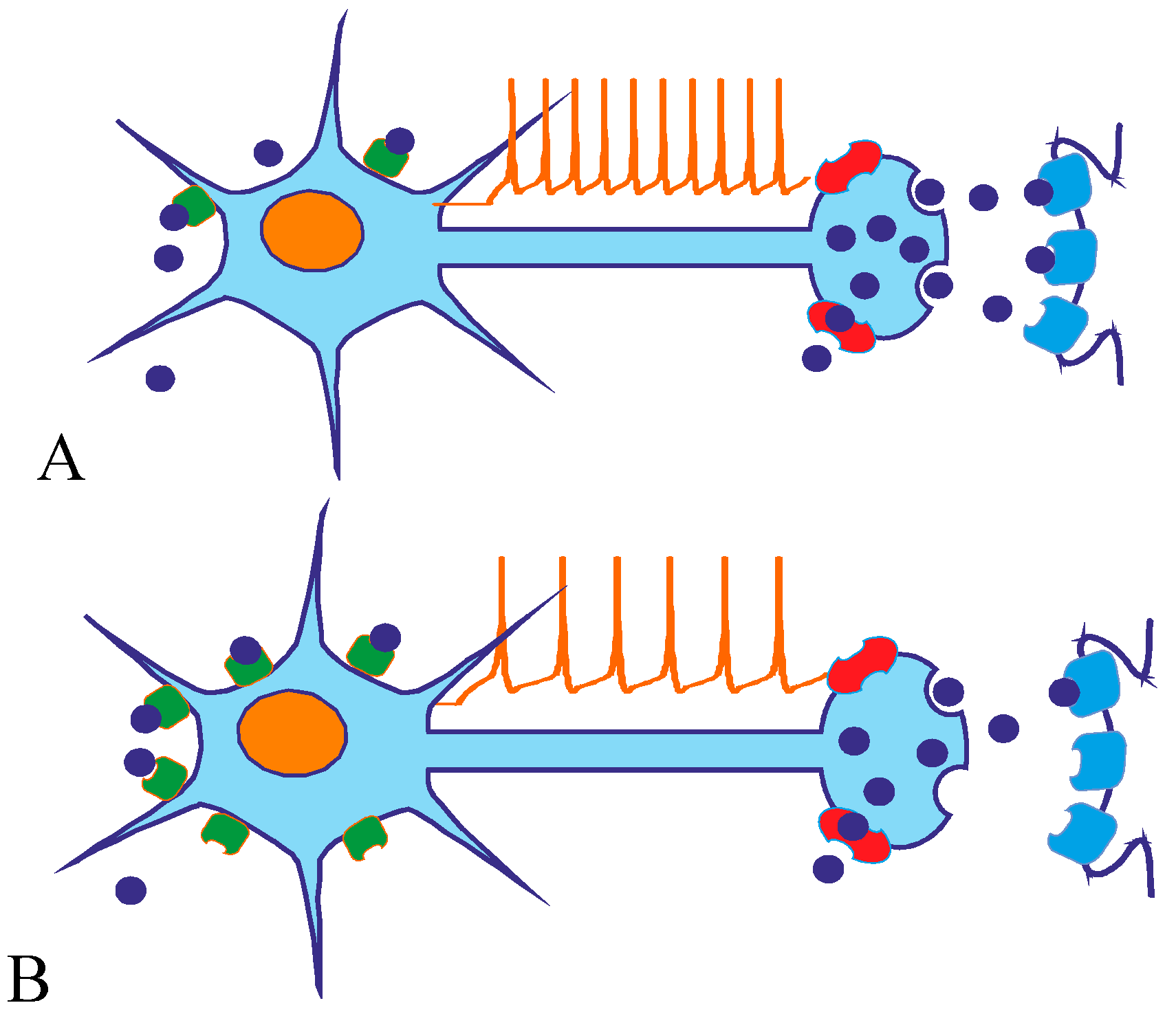
4.2. Role of Pre- and Postsynaptic 5-HT1A Receptors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioural Effect | Auto- and Heteroreceptors | Autoreceptors | Heteroreceptors |
|---|---|---|---|
| Whole life knockout [13,14,104,106,107,108,109,112] | |||
| Anxiety | Elevated | Elevated | No impact |
| Depression | Lower | No impact | Elevated |
| Knock-down between 14 and 30 days (40%) [113] | |||
| Anxiety | - * | Long-term increase | - |
| Depression | - | No effect | - |
| Adulthood [9,13,114] | |||
| Anxiety | - | No effect | - |
| Depression | - | Diminished | - |
| Reduced activity | |||
| Anxiety | - | - | Elevated [115] |
| Depression | - | Diminished [13,116,117] | - |
| Overexpression | |||
| Anxiety | Diminished [105,118] | - | - |
| Depression | - | Elevated [110] | - |
| Increased transcription | |||
| Depression | - | Increased [115] | - |
4.3. Antidepressant Treatment and 5-HT1A Receptors
5. Models of Depression Involved Circuits


6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Raymond, J.R.; Mukhin, Y.V.; Gelasco, A.; Turner, J.; Collinsworth, G.; Gettys, T.W.; Grewal, J.S.; Garnovskaya, M.N. Multiplicity of mechanisms of serotonin receptor signal transduction. Pharmacol. Ther. 2001, 92, 179–212. [Google Scholar] [PubMed]
- Artigas, F. Developments in the field of antidepressants, where do we go now? Eur. Neuropsychopharmacol. 2015, 25, 657–670. [Google Scholar] [PubMed]
- Jacobs, B.L.; Azmitia, E.C. Structure and function of the brain serotonin system. Physiol. Rev. 1992, 72, 165–229. [Google Scholar] [PubMed]
- Artigas, F. Serotonin receptors involved in antidepressant effects. Pharmacol. Ther. 2013, 137, 119–131. [Google Scholar] [PubMed]
- Adell, A.; Celada, P.; Abellan, M.T.; Artigas, F. Origin and functional role of the extracellular serotonin in the midbrain raphe nuclei. Brain Res. Brain Res. Rev. 2002, 39, 154–180. [Google Scholar] [PubMed]
- Beaudoin-Gobert, M.; Sgambato-Faure, V. Serotonergic pharmacology in animal models: From behavioral disorders to dyskinesia. Neuropharmacology 2014, 81, 15–30. [Google Scholar] [PubMed]
- Valdizan, E.M.; Castro, E.; Pazos, A. Agonist-dependent modulation of G-protein coupling and transduction of 5-HT1A receptors in rat dorsal raphe nucleus. Int. J. Neuropsychopharmacol. 2010, 13, 835–843. [Google Scholar] [PubMed]
- Fiorino, F.; Severino, B.; Magli, E.; Ciano, A.; Caliendo, G.; Santagada, V.; Frecentese, F.; Perissutti, E. 5-HT (1A) receptor: An old target as a new attractive tool in drug discovery from central nervous system to cancer. J. Med. Chem. 2014, 57, 4407–4426. [Google Scholar] [PubMed]
- Celada, P.; Bortolozzi, A.; Artigas, F. Serotonin 5-HT1A receptors as targets for agents to treat psychiatric disorders: Rationale and current status of research. CNS Drugs 2013, 27, 703–716. [Google Scholar] [PubMed]
- Albert, P.R.; Le, F.B.; Millar, A.M. Transcriptional dysregulation of 5-HT1A autoreceptors in mental illness. Mol. Brain 2011, 4, 21–24. [Google Scholar] [PubMed]
- Hervas, I.; Queiroz, C.M.; Adell, A.; Artigas, F. Role of uptake inhibition and autoreceptor activation in the control of 5-HT release in the frontal cortex and dorsal hippocampus of the rat. Br. J. Pharmacol. 2000, 130, 160–166. [Google Scholar] [PubMed]
- Bang, S.J.; Jensen, P.; Dymecki, S.M.; Commons, K.G. Projections and interconnections of genetically defined serotonin neurons in mice. Eur. J. Neurosci. 2012, 35, 85–96. [Google Scholar] [PubMed]
- Richardson-Jones, J.W.; Craige, C.P.; Guiard, B.P.; Stephen, A.; Metzger, K.L.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.J.; Beck, S.G.; et al. 5-HT1A autoreceptor levels determine vulnerability to stress and response to antidepressants. Neuron 2010, 65, 40–52. [Google Scholar] [PubMed]
- Garcia-Garcia, A.L.; Newman-Tancredi, A.; Leonardo, E.D. 5-HT (1A) receptors in mood and anxiety: Recent insights into autoreceptor versus heteroreceptor function. Psychopharmacology 2014, 231, 623–636. [Google Scholar] [PubMed]
- Pauwels, P.J. 5-HT 1B/D receptor antagonists. Gen. Pharmacol. 1997, 29, 293–303. [Google Scholar] [PubMed]
- Hoyer, D.; Hannon, J.P.; Martin, G.R. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol. Biochem. Behav. 2002, 71, 533–554. [Google Scholar] [PubMed]
- Malleret, G.; Hen, R.; Guillou, J.L.; Segu, L.; Buhot, M.C. 5-HT1B receptor knock-out mice exhibit increased exploratory activity and enhanced spatial memory performance in the Morris water maze. J. Neurosci. 1999, 19, 6157–6168. [Google Scholar] [PubMed]
- Malagie, I.; David, D.J.; Jolliet, P.; Hen, R.; Bourin, M.; Gardier, A.M. Improved efficacy of fluoxetine in increasing hippocampal 5-hydroxytryptamine outflow in 5-HT(1B) receptor knock-out mice. Eur. J. Pharmacol. 2002, 443, 99–104. [Google Scholar] [PubMed]
- Tatarczynska, E.; Klodzinska, A.; Stachowicz, K.; Chojnacka-Wojcik, E. Effects of a selective 5-HT1B receptor agonist and antagonists in animal models of anxiety and depression. Behav. Pharmacol. 2004, 15, 523–534. [Google Scholar] [PubMed]
- Chenu, F.; David, D.J.; Leroux-Nicollet, I.; Le, M.E.; Gardier, A.M.; Bourin, M. Serotonin1B heteroreceptor activation induces an antidepressant-like effect in mice with an alteration of the serotonergic system. J. Psychiatry Neurosci. 2008, 33, 541–550. [Google Scholar] [PubMed]
- Raymond, J.R.; Turner, J.H.; Gelasco, A.K.; Ayiku, H.B.; Coaxum, S.D.; Arthur, J.M.; Garnovskaya, M.N. 5-HT Receptor Signal Transduction Pathways. In The Serotonin Receptors. From Molecular Pharmacology to Human Therapeutics; Roth, B.R., Ed.; Humana Press: Toyota, NJ, USA, 2006. [Google Scholar]
- Hsiung, S.C.; Tamir, H.; Franke, T.F.; Liu, K.P. Roles of extracellular signal-regulated kinase and Akt signaling in coordinating nuclear transcription factor-kappaB-dependent cell survival after serotonin 1A receptor activation. J. Neurochem. 2005, 95, 1653–1666. [Google Scholar] [PubMed]
- Adayev, T.; Ray, I.; Sondhi, R.; Sobocki, T.; Banerjee, P. The G protein-coupled 5-HT1A receptor causes suppression of caspase-3 through MAPK and protein kinase Calpha. Biochim. Biophys. Acta 2003, 1640, 85–96. [Google Scholar] [PubMed]
- Polter, A.M.; Li, X. 5-HT1A receptor-regulated signal transduction pathways in brain. Cell Signal. 2010, 22, 1406–1412. [Google Scholar] [PubMed]
- Clarke, W.P.; Yocca, F.D.; Maayani, S. Lack of 5-hydroxytryptamine1A-mediated inhibition of adenylyl cyclase in dorsal raphe of male and female rats. J. Pharmacol. Exp. Ther. 1996, 277, 1259–1266. [Google Scholar] [PubMed]
- Mannoury la, C.C.; El, M.S.; Hanoun, N.; Hamon, M.; Lanfumey, L. Regional differences in the coupling of 5-hydroxytryptamine-1A receptors to G proteins in the rat brain. Mol. Pharmacol. 2006, 70, 1013–1021. [Google Scholar]
- Marazziti, D.; Palego, L.; Giromella, A.; Mazzoni, M.R.; Borsini, F.; Mayer, N.; Naccarato, A.G.; Lucacchini, A.; Cassano, G.B. Region-dependent effects of flibanserin and buspirone on adenylyl cyclase activity in the human brain. Int. J. Neuropsychopharmacol. 2002, 5, 131–140. [Google Scholar] [PubMed]
- Martel, J.C.; Ormiere, A.M.; Leduc, N.; Assie, M.B.; Cussac, D.; Newman-Tancredi, A. Native rat hippocampal 5-HT1A receptors show constitutive activity. Mol. Pharmacol. 2007, 71, 638–643. [Google Scholar] [CrossRef]
- Moyano, S.; Del, R.J.; Frechilla, D. Role of hippocampal CaMKII in serotonin 5-HT (1A) receptor-mediated learning deficit in rats. Neuropsychopharmacology 2004, 29, 2216–2224. [Google Scholar] [CrossRef] [PubMed]
- Luscher, C.; Jan, L.Y.; Stoffel, M.; Malenka, R.C.; Nicoll, R.A. G protein-coupled inwardly rectifying K+ channels (GIRKs) mediate postsynaptic but not presynaptic transmitter actions in hippocampal neurons. Neuron 1997, 19, 687–695. [Google Scholar] [CrossRef]
- Oleskevich, S. G alpha o1 decapeptide modulates the hippocampal 5-HT1A potassium current. J. Neurophysiol. 1995, 74, 2189–2193. [Google Scholar] [PubMed]
- Colino, A.; Halliwell, J.V. Differential modulation of three separate K-conductances in hippocampal CA1 neurons by serotonin. Nature 1987, 328, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.; Nicoll, R.A. Pharmacologically distinct actions of serotonin on single pyramidal neurones of the rat hippocampus recorded in vitro. J. Physiol. 1987, 394, 99–124. [Google Scholar] [CrossRef] [PubMed]
- Clarke, W.P.; De, V.M.; Beck, S.G.; Maayani, S.; Goldfarb, J. Serotonin decreases population spike amplitude in hippocampal cells through a pertussis toxin substrate. Brain Res. 1987, 410, 357–361. [Google Scholar] [CrossRef]
- Penington, N.J.; Kelly, J.S.; Fox, A.P. Whole-cell recordings of inwardly rectifying K+ currents activated by 5-HT1A receptors on dorsal raphe neurones of the adult rat. J. Physiol. 1993, 469, 387–405. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, D.A.; Li, Y.W.; Talley, E.M. Effects of serotonin on caudal raphe neurons: Activation of an inwardly rectifying potassium conductance. J. Neurophysiol. 1997, 77, 1349–1361. [Google Scholar] [PubMed]
- Ehrengruber, M.U.; Doupnik, C.A.; Xu, Y.; Garvey, J.; Jasek, M.C.; Lester, H.A.; Davidson, N. Activation of heteromeric G protein-gated inward rectifier K+ channels overexpressed by adenovirus gene transfer inhibits the excitability of hippocampal neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 7070–7075. [Google Scholar] [CrossRef] [PubMed]
- Loucif, A.J.; Bonnavion, P.; Macri, B.; Golmard, J.L.; Boni, C.; Melfort, M.; Leonard, G.; Lesch, K.P.; Adrien, J.; Jacquin, T.D. Gender-dependent regulation of G-protein-gated inwardly rectifying potassium current in dorsal raphe neurons in knock-out mice devoid of the 5-hydroxytryptamine transporter. J. Neurobiol. 2006, 66, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Doupnik, C.A.; Davidson, N.; Lester, H.A.; Kofuji, P. RGS proteins reconstitute the rapid gating kinetics of gbetagamma-activated inwardly rectifying K+ channels. Proc. Natl. Acad. Sci. USA 1997, 94, 10461–10466. [Google Scholar] [CrossRef] [PubMed]
- Doupnik, C.A.; Dessauer, C.W.; Slepak, V.Z.; Gilman, A.G.; Davidson, N.; Lester, H.A. Time resolved kinetics of direct G beta 1 gamma 2 interactions with the carboxyl terminus of Kir3.4 inward rectifier K+ channel subunits. Neuropharmacology 1996, 35, 923–931. [Google Scholar] [CrossRef]
- Kovoor, A.; Lester, H.A. Gi Irks GIRKs. Neuron 2002, 33, 6–8. [Google Scholar] [CrossRef]
- Tsetsenis, T.; Ma, X.H.; Lo, I.L.; Beck, S.G.; Gross, C. Suppression of conditioning to ambiguous cues by pharmacogenetic inhibition of the dentate gyrus. Nat. Neurosci. 2007, 10, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Penington, N.J. Differential effects of protein kinase C activation on 5-HT1A receptor coupling to Ca2+ and K+ currents in rat serotonergic neurones. J. Physiol. 1996, 496, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Penington, N.J.; Fox, A.P. Toxin-insensitive Ca current in dorsal raphe neurons. J. Neurosci. 1995, 15, 5719–5726. [Google Scholar] [PubMed]
- Bayliss, D.A.; Li, Y.W.; Talley, E.M. Effects of serotonin on caudal raphe neurons: Inhibition of N- and P/Q-type calcium channels and the afterhyperpolarization. J. Neurophysiol. 1997, 77, 1362–1374. [Google Scholar] [PubMed]
- Chen, Y.; Penington, N.J. QEHA27, a peptide that binds to G-protein beta gamma-subunits, reduces the inhibitory effect of 5-HT on the Ca2+ current of rat dorsal raphe neurons. Neurosci. Lett. 1997, 224, 87–90. [Google Scholar] [CrossRef]
- Blumer, K.J.; Johnson, G.L. Diversity in function and regulation of MAP kinase pathways. Trends Biochem. Sci. 1994, 19, 236–240. [Google Scholar] [CrossRef]
- Garnovskaya, M.N.; van Biesen, T.; Hawe, B.; Ramos, S.C.; Lefkowitz, R.J.; Raymond, J.R. Ras-dependent activation of fibroblast mitogen-activated protein kinase by 5-HT1A receptor via a G protein beta gamma-subunit-initiated pathway. Biochemistry 1996, 35, 13716–13722. [Google Scholar] [CrossRef] [PubMed]
- Cowen, D.S.; Sowers, R.S.; Manning, D.R. Activation of a mitogen-activated protein kinase (ERK2) by the 5-hydroxytryptamine1A receptor is sensitive not only to inhibitors of phosphatidylinositol 3-kinase, but to an inhibitor of phosphatidylcholine hydrolysis. J. Biol. Chem. 1996, 271, 22297–22300. [Google Scholar] [PubMed]
- Millan, M.J.; Newman-Tancredi, A.; Duqueyroix, D.; Cussac, D. Agonist properties of pindolol at h5-HT1A receptors coupled to mitogen-activated protein kinase. Eur. J. Pharmacol. 2001, 424, 13–17. [Google Scholar] [CrossRef]
- Garnovskaya, M.N.; Mukhin, Y.; Raymond, J.R. Rapid activation of sodium-proton exchange and extracellular signal-regulated protein kinase in fibroblasts by G protein-coupled 5-HT1A receptor involves distinct signalling cascades. Biochem. J. 1998, 330, 489–495. [Google Scholar] [PubMed]
- la Rocca, G.J.; Mukhin, Y.V.; Garnovskaya, M.N.; Daaka, Y.; Clark, G.J.; Luttrell, L.M.; Lefkowitz, R.J.; Raymond, J.R. Serotonin 5-HT1A receptor-mediated Erk activation requires calcium/calmodulin-dependent receptor endocytosis. J. Biol. Chem. 1999, 274, 4749–4753. [Google Scholar] [CrossRef]
- Sullivan, N.R.; Crane, J.W.; Damjanoska, K.J.; Carrasco, G.A.; D'Souza, D.N.; Garcia, F.; van de Kar, L.D. Tandospirone activates neuroendocrine and ERK (MAP kinase) signaling pathways specifically through 5-HT1A receptor mechanisms in vivo. Naunyn Schmiedebergs Arch. Pharmacol. 2005, 371, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Buritova, J.; Berrichon, G.; Cathala, C.; Colpaert, F.; Cussac, D. Region-specific changes in 5-HT1A agonist-induced Extracellular signal-Regulated Kinases 1/2 phosphorylation in rat brain: A quantitative ELISA study. Neuropharmacology 2009, 56, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Newman-Tancredi, A.; Martel, J.C.; Assie, M.B.; Buritova, J.; Lauressergues, E.; Cosi, C.; Heusler, P.; Bruins, S.L.; Colpaert, F.C.; Vacher, B.; et al. Signal transduction and functional selectivity of F15599, a preferential post-synaptic 5-HT1A receptor agonist. Br. J. Pharmacol. 2009, 156, 338–353. [Google Scholar] [CrossRef] [PubMed]
- Adayev, T.; El-Sherif, Y.; Barua, M.; Penington, N.J.; Banerjee, P. Agonist stimulation of the serotonin1A receptor causes suppression of anoxia-induced apoptosis via mitogen-activated protein kinase in neuronal HN2-5 cells. J. Neurochem. 1999, 72, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Cowen, D.S.; Johnson-Farley, N.N.; Travkina, T. 5-HT1A receptors couple to activation of Akt, but not extracellular-regulated kinase (ERK), in cultured hippocampal neurons. J. Neurochem. 2005, 93, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Druse, M.; Tajuddin, N.F.; Gillespie, R.A.; Le, P. Signaling pathways involved with serotonin1A agonist-mediated neuroprotection against ethanol-induced apoptosis of fetal rhombencephalic neurons. Brain Res. Dev. Brain Res. 2005, 159, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, N.; Albert, P.R. Coupling of 5-HT1A autoreceptors to inhibition of mitogen-activated protein kinase activation via G beta gamma subunit signaling. Eur. J. Neurosci. 2005, 21, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.W.; Shimizu, K.; Carrasco, G.A.; Garcia, F.; Jia, C.; Sullivan, N.R.; D’Souza, D.N.; Zhang, Y.; Van de Kar, L.D.; Muma, N.A.; et al. 5-HT1A receptors mediate (+)8-OH-DPAT-stimulation of extracellular signal-regulated kinase (MAP kinase) in vivo in rat hypothalamus: Time dependence and regional differences. Brain Res. 2007, 1183, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shen, C.; Meller, E. 5-HT1A receptor-mediated regulation of mitogen-activated protein kinase phosphorylation in rat brain. Eur. J. Pharmacol. 2002, 452, 155–162. [Google Scholar] [CrossRef]
- Sturgill, T.W.; Ray, L.B.; Erikson, E.; Maller, J.L. Insulin-stimulated MAP-2 kinase phosphorylates and activates ribosomal protein S6 kinase II. Nature 1988, 334, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Cowen, D.S.; Molinoff, P.B.; Manning, D.R. 5-hydroxytryptamine1A receptor-mediated increases in receptor expression and activation of nuclear factor-kappaB in transfected Chinese hamster ovary cells. Mol. Pharmacol. 1997, 52, 221–226. [Google Scholar] [PubMed]
- Blendy, J.A. The role of CREB in depression and antidepressant treatment. Biol. Psychiatry 2006, 59, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Einat, H.; Yuan, P.; Gould, T.D.; Li, J.; Du, J.; Zhang, L.; Manji, H.K.; Chen, G. The role of the extracellular signal-regulated kinase signaling pathway in mood modulation. J. Neurosci. 2003, 23, 7311–7316. [Google Scholar] [PubMed]
- Ailing, F.; Fan, L.; Li, S.; Manji, S. Role of extracellular signal-regulated kinase signal transduction pathway in anxiety. J. Psychiatr. Res. 2008, 43, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Mailliet, F.; Spedding, M.; Rocher, C.; Zhang, X.; Delagrange, P.; McEwen, B.; Jay, T.M.; Svenningsson, P. Antidepressants reverse the attenuation of the neurotrophic MEK/MAPK cascade in frontal cortex by elevated platform stress; reversal of effects on LTP is associated with GluA1 phosphorylation. Neuropharmacology 2009, 56, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Creson, T.K.; Hao, Y.; Engel, S.; Shen, Y.; Hamidi, A.; Zhuo, M.; Manji, H.K.; Chen, G. The anterior cingulate ERK pathway contributes to regulation of behavioral excitement and hedonic activity. Bipolar Disord. 2009, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Duman, C.H.; Schlesinger, L.; Kodama, M.; Russell, D.S.; Duman, R.S. A role for MAP kinase signaling in behavioral models of depression and antidepressant treatment. Biol. Psychiatry 2007, 61, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhao, Y.; Zhu, X.; Cai, Z.; Wang, S.; Yao, S.; Qi, Z.; Xie, P. Fluoxetine upregulates phosphorylated-AKT and phosphorylated-ERK1/2 proteins in neural stem cells: Evidence for a crosstalk between AKT and ERK1/2 pathways. J. Mol. Neurosci. 2013, 49, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Sale, E.M.; Sale, G.J. Protein kinase B: Signalling roles and therapeutic targeting. Cell Mol. Life Sci. 2008, 65, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Cheon, S.H.; Yoo, S.J.; Kwon, J.; Park, J.H.; Kim, C.G.; Rhee, K.; You, S.; Lee, J.Y.; Roh, S.I.; et al. Contribution of the PI3K/Akt/PKB signal pathway to maintenance of self-renewal in human embryonic stem cells. FEBS Lett. 2005, 579, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Saunders, C.; Siuta, M.; Robertson, S.D.; Davis, A.R.; Sauer, J.; Matthies, H.J.; Gresch, P.J.; Airey, D.C.; Lindsley, C.W.; Schetz, J.A.; et al. Neuronal ablation of p-Akt at Ser473 leads to altered 5-HT1A/2A receptor function. Neurochem. Int. 2014, 73, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fanburg, B.L. Serotonin-induced growth of pulmonary artery smooth muscle requires activation of phosphatidylinositol 3-kinase/serine-threonine protein kinase B/mammalian target of rapamycin/p70 ribosomal S6 kinase 1. Am. J. Respir. Cell Mol. Biol. 2006, 34, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Cross, D.A.; Alessi, D.R.; Cohen, P.; Andjelkovich, M.; Hemmings, B.A. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 1995, 378, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Goode, N.; Hughes, K.; Woodgett, J.R.; Parker, P.J. Differential regulation of glycogen synthase kinase-3 beta by protein kinase C isotypes. J. Biol. Chem. 1992, 267, 16878–16882. [Google Scholar] [PubMed]
- Li, M.; Wang, X.; Meintzer, M.K.; Laessig, T.; Birnbaum, M.J.; Heidenreich, K.A. Cyclic AMP promotes neuronal survival by phosphorylation of glycogen synthase kinase 3beta. Mol. Cell. Biol. 2000, 20, 9356–9363. [Google Scholar] [CrossRef] [PubMed]
- Klein, P.S.; Melton, D.A. A molecular mechanism for the effect of lithium on development. Proc. Natl. Acad. Sci. USA 1996, 93, 8455–8459. [Google Scholar] [CrossRef] [PubMed]
- De Sarno, P.; Li, X.; Jope, R.S. Regulation of Akt and glycogen synthase kinase-3 beta phosphorylation by sodium valproate and lithium. Neuropharmacology 2002, 43, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
- Kaidanovich-Beilin, O.; Milman, A.; Weizman, A.; Pick, C.G.; Eldar-Finkelman, H. Rapid antidepressive-like activity of specific glycogen synthase kinase-3 inhibitor and its effect on beta-catenin in mouse hippocampus. Biol. Psychiatry 2004, 55, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Gould, T.D.; Einat, H.; Bhat, R.; Manji, H.K. AR-A014418, a selective GSK-3 inhibitor, produces antidepressant-like effects in the forced swim test. Int. J. Neuropsychopharmacol. 2004, 7, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Sotnikova, T.D.; Yao, W.D.; Kockeritz, L.; Woodgett, J.R.; Gainetdinov, R.R.; Caron, M.G. Lithium antagonizes dopamine-dependent behaviors mediated by an AKT/glycogen synthase kinase 3 signaling cascade. Proc. Natl. Acad. Sci. USA 2004, 101, 5099–5104. [Google Scholar] [CrossRef] [PubMed]
- Prickaerts, J.; Moechars, D.; Cryns, K.; Lenaerts, I.; van Craenendonck, H.; Goris, I.; Daneels, G.; Bouwknecht, J.A.; Steckler, T. Transgenic mice overexpressing glycogen synthase kinase 3beta: A putative model of hyperactivity and mania. J. Neurosci. 2006, 26, 9022–9029. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Moussaif, M.; Kuan, C.J.; Gargus, J.J.; Sze, J.Y. Serotonin targets the DAF-16/FOXO signaling pathway to modulate stress responses. Cell Metab. 2006, 4, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Polter, A.; Yang, S.; Zmijewska, A.A.; van Groen, T.; Paik, J.H.; Depinho, R.A.; Peng, S.L.; Jope, R.S.; Li, X. Forkhead box, class O transcription factors in brain: Regulation and behavioral manifestation. Biol. Psychiatry 2009, 65, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Liu, L.; Zhang, R.; Li, X. Lithium reduces FoxO3a transcriptional activity by decreasing its intracellular content. Biol. Psychiatry 2007, 62, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.H.; Garnovskaya, M.N.; Coaxum, S.D.; Vlasova, T.M.; Yakutovich, M.; Lefler, D.M.; Raymond, J.R. Ca2+-calmodulin and janus kinase 2 are required for activation of sodium-proton exchange by the Gi-coupled 5-hydroxytryptamine 1a receptor. J. Pharmacol. Exp. Ther. 2007, 320, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Carmena, M.J.; Camacho, A.; Solano, R.M.; Montalvo, L.; Garcia-Lopez, E.; Arias, A.; Prieto, J.C. 5-hydroxytryptamine1A receptor-mediated effects on adenylate cyclase and nitric oxide synthase activities in rat ventral prostate. Cell Signal. 1998, 10, 583–587. [Google Scholar] [CrossRef]
- Shinka, T.; Onodera, D.; Tanaka, T.; Shoji, N.; Miyazaki, T.; Moriuchi, T.; Fukumoto, T. Serotonin synthesis and metabolism-related molecules in a human prostate cancer cell line. Oncol. Lett. 2011, 2, 211–215. [Google Scholar] [PubMed]
- Strosznajder, J.; Chalimoniuk, M.; Samochocki, M. Activation of serotonergic 5-HT1A receptor reduces Ca2+- and glutamatergic receptor-evoked arachidonic acid and No/cGMP release in adult hippocampus. Neurochem. Int. 1996, 28, 439–444. [Google Scholar] [CrossRef]
- Maura, G.; Marcoli, M.; Pepicelli, O.; Rosu, C.; Viola, C.; Raiteri, M. Serotonin inhibition of the NMDA receptor/nitric oxide/cyclic GMP pathway in human neocortex slices: Involvement of 5-HT(2C) and 5-HT(1A) receptors. Br. J. Pharmacol. 2000, 130, 1853–1858. [Google Scholar] [CrossRef] [PubMed]
- Volke, V.; Wegener, G.; Bourin, M.; Vasar, E. Antidepressant- and anxiolytic-like effects of selective neuronal NOS inhibitor 1-(2-trifluoromethylphenyl)-imidazole in mice. Behav. Brain Res. 2003, 140, 141–147. [Google Scholar] [CrossRef]
- Yildiz, F.; Erden, B.F.; Ulak, G.; Utkan, T.; Gacar, N. Antidepressant-like effect of 7-nitroindazole in the forced swimming test in rats. Psychopharmacology 2000, 149, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Ulak, G.; Mutlu, O.; Tanyeri, P.; Komsuoglu, F.I.; Akar, F.Y.; Erden, B.F. Involvement of serotonin receptor subtypes in the antidepressant-like effect of TRIM in the rat forced swimming test. Pharmacol. Biochem. Behav. 2010, 95, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Heusler, P.; Pauwels, P.J.; Wurch, T.; Newman-Tancredi, A.; Tytgat, J.; Colpaert, F.C.; Cussac, D. Differential ion current activation by human 5-HT(1A) receptors in Xenopus oocytes: Evidence for agonist-directed trafficking of receptor signalling. Neuropharmacology 2005, 49, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Shopsin, B.; Friedman, E.; Gershon, S. Parachlorophenylalanine reversal of tranylcypromine effects in depressed patients. Arch. Gen. Psychiatry 1976, 33, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Young, S.N.; Smith, S.E.; Pihl, R.O.; Ervin, F.R. Tryptophan depletion causes a rapid lowering of mood in normal males. Psychopharmacology 1985, 87, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Stark, P.; Hardison, C.D. A review of multicenter controlled studies of fluoxetine vs. imipramine and placebo in outpatients with major depressive disorder. J. Clin. Psychiatry 1985, 46, 53–58. [Google Scholar] [PubMed]
- Franklin, M.; Bermudez, I.; Murck, H.; Singewald, N.; Gaburro, S. Sub-chronic dietary tryptophan depletion--an animal model of depression with improved face and good construct validity. J. Psychiatr. Res. 2012, 46, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Chau, D.T.; Rada, P.V.; Kim, K.; Kosloff, R.A.; Hoebel, B.G. Fluoxetine alleviates behavioral depression while decreasing acetylcholine release in the nucleus accumbens shell. Neuropsychopharmacology 2011, 36, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Mouillet-Richard, S.; Schneider, B.; Launay, J.M.; Kellermann, O. miR-16 targets the serotonin transporter: A new facet for adaptive responses to antidepressants. Science 2010, 329, 1537–1541. [Google Scholar] [CrossRef] [PubMed]
- Temel, Y.; Boothman, L.J.; Blokland, A.; Magill, P.J.; Steinbusch, H.W.; Visser-Vandewalle, V.; Sharp, T. Inhibition of 5-HT neuron activity and induction of depressive-like behavior by high-frequency stimulation of the subthalamic nucleus. Proc. Natl. Acad. Sci. USA 2007, 104, 17087–17092. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.; Santarelli, L.; Brunner, D.; Zhuang, X.; Hen, R. Altered fear circuits in 5-HT (1A) receptor KO mice. Biol. Psychiatry 2000, 48, 1157–1163. [Google Scholar] [CrossRef]
- Kusserow, H.; Davies, B.; Hortnagl, H.; Voigt, I.; Stroh, T.; Bert, B.; Deng, D.R.; Fink, H.; Veh, R.W.; Theuring, F. Reduced anxiety-related behaviour in transgenic mice overexpressing serotonin 1A receptors. Brain Res. Mol. Brain Res. 2004, 129, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Heisler, L.K.; Chu, H.M.; Brennan, T.J.; Danao, J.A.; Bajwa, P.; Parsons, L.H.; Tecott, L.H. Elevated anxiety and antidepressant-like responses in serotonin 5-HT1A receptor mutant mice. Proc. Natl. Acad. Sci. USA 1998, 95, 15049–15054. [Google Scholar] [CrossRef] [PubMed]
- Klemenhagen, K.C.; Gordon, J.A.; David, D.J.; Hen, R.; Gross, C.T. Increased fear response to contextual cues in mice lacking the 5-HT1A receptor. Neuropsychopharmacology 2006, 31, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Parks, C.L.; Robinson, P.S.; Sibille, E.; Shenk, T.; Toth, M. Increased anxiety of mice lacking the serotonin1A receptor. Proc. Natl. Acad. Sci. USA 1998, 95, 10734–10739. [Google Scholar] [CrossRef] [PubMed]
- Ramboz, S.; Oosting, R.; Amara, D.A.; Kung, H.F.; Blier, P.; Mendelsohn, M.; Mann, J.J.; Brunner, D.; Hen, R. Serotonin receptor 1A knockout: An animal model of anxiety-related disorder. Proc. Natl. Acad. Sci. USA 1998, 95, 14476–14481. [Google Scholar] [CrossRef] [PubMed]
- Gunther, L.; Rothe, J.; Rex, A.; Voigt, J.P.; Millan, M.J.; Fink, H.; Bert, B. 5-HT(1A)-receptor over-expressing mice: Genotype and sex dependent responses to antidepressants in the forced swim-test. Neuropharmacology 2011, 61, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Maletic, V.; Robinson, M.; Oakes, T.; Iyengar, S.; Ball, S.G.; Russell, J. Neurobiology of depression: An integrated view of key findings. Int. J. Clin. Pract. 2007, 61, 2030–2040. [Google Scholar] [CrossRef] [PubMed]
- Richardson-Jones, J.W.; Craige, C.P.; Nguyen, T.H.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.J.; Guiard, B.P.; Beck, S.G.; Hen, R.; et al. Serotonin-1A autoreceptors are necessary and sufficient for the normal formation of circuits underlying innate anxiety. J. Neurosci. 2011, 31, 6008–6018. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, Z.R.; Piel, D.A.; Santos, T.L.; Richardson-Jones, J.; Leonardo, E.D.; Beck, S.G.; Champagne, F.A.; Hen, R. Developmental effects of serotonin 1A autoreceptors on anxiety and social behavior. Neuropsychopharmacology 2014, 39, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Bortolozzi, A.; Castane, A.; Semakova, J.; Santana, N.; Alvarado, G.; Cortes, R.; Ferres-Coy, A.; Fernandez, G.; Carmona, M.C.; Toth, M.; et al. New antidepressant strategy based on acute siRNA silencing of 5-HT1A autoreceptors. Mol. Psychiatry 2012, 17, 567. [Google Scholar] [CrossRef] [PubMed]
- Albert, P.R.; Francois, B.L. Modifying 5-HT1A receptor gene expression as a new target for antidepressant therapy. Front. Neurosci. 2010, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Ferres-Coy, A.; Santana, N.; Castane, A.; Cortes, R.; Carmona, M.C.; Toth, M.; Montefeltro, A.; Artigas, F.; Bortolozzi, A. Acute 5-HT(1)A autoreceptor knockdown increases antidepressant responses and serotonin release in stressful conditions. Psychopharmacology 2013, 225, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Bortolozzi, A.; Castane, A.; Semakova, J.; Santana, N.; Alvarado, G.; Cortes, R.; Ferres-Coy, A.; Fernandez, G.; Carmona, M.C.; Toth, M.; et al. Selective siRNA-mediated suppression of 5-HT1A autoreceptors evokes strong anti-depressant-like effects. Mol. Psychiatry 2012, 17, 612–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordukalo-Niksic, T.; Mokrovic, G.; Stefulj, J.; Zivin, M.; Jernej, B.; Cicin-Sain, L. 5HT-1A receptors and anxiety-like behaviours: Studies in rats with constitutionally upregulated/downregulated serotonin transporter. Behav. Brain Res. 2010, 213, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Assie, M.B.; Lomenech, H.; Ravailhe, V.; Faucillon, V.; Newman-Tancredi, A. Rapid desensitization of somatodendritic 5-HT1A receptors by chronic administration of the high-efficacy 5-HT1A agonist, F13714: A microdialysis study in the rat. Br. J. Pharmacol. 2006, 149, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Depoortere, R.; Auclair, A.L.; Bardin, L.; Colpaert, F.C.; Vacher, B.; Newman-Tancredi, A. F15599, a preferential post-synaptic 5-HT1A receptor agonist: Activity in models of cognition in comparison with reference 5-HT1A receptor agonists. Eur. Neuropsychopharmacol. 2010, 20, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Llado-Pelfort, L.; Assie, M.B.; Newman-Tancredi, A.; Artigas, F.; Celada, P. Preferential in vivo action of F15599, a novel 5-HT(1A) receptor agonist, at postsynaptic 5-HT(1A) receptors. Br. J. Pharmacol. 2010, 160, 1929–1940. [Google Scholar] [CrossRef] [PubMed]
- Assie, M.B.; Bardin, L.; Auclair, A.L.; Carilla-Durand, E.; Depoortere, R.; Koek, W.; Kleven, M.S.; Colpaert, F.; Vacher, B.; Newman-Tancredi, A. F15599, a highly selective post-synaptic 5-HT(1A) receptor agonist: In vivo profile in behavioural models of antidepressant and serotonergic activity. Int. J. Neuropsychopharmacol. 2010, 13, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Savitz, J.; Lucki, I.; Drevets, W.C. 5-HT (1A) receptor function in major depressive disorder. Prog. Neurobiol. 2009, 88, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Lemonde, S.; Turecki, G.; Bakish, D.; Du, L.; Hrdina, P.D.; Bown, C.D.; Sequeira, A.; Kushwaha, N.; Morris, S.J.; Basak, A.; et al. Impaired repression at a 5-hydroxytryptamine 1A receptor gene polymorphism associated with major depression and suicide. J. Neurosci. 2003, 23, 8788–8799. [Google Scholar] [PubMed]
- Stockmeier, C.A.; Shapiro, L.A.; Dilley, G.E.; Kolli, T.N.; Friedman, L.; Rajkowska, G. Increase in serotonin-1A autoreceptors in the midbrain of suicide victims with major depression-postmortem evidence for decreased serotonin activity. J. Neurosci. 1998, 18, 7394–7401. [Google Scholar] [PubMed]
- Neff, C.D.; Abkevich, V.; Packer, J.C.; Chen, Y.; Potter, J.; Riley, R.; Davenport, C.; DeGrado, W.J.; Jammulapati, S.; Bhathena, A.; et al. Evidence for HTR1A and LHPP as interacting genetic risk factors in major depression. Mol. Psychiatry 2009, 14, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Sargent, P.A.; Kjaer, K.H.; Bench, C.J.; Rabiner, E.A.; Messa, C.; Meyer, J.; Gunn, R.N.; Grasby, P.M.; Cowen, P.J. Brain serotonin1A receptor binding measured by positron emission tomography with [11C]WAY-100635: Effects of depression and antidepressant treatment. Arch. Gen. Psychiatry 2000, 57, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Bhagwagar, Z.; Rabiner, E.A.; Sargent, P.A.; Grasby, P.M.; Cowen, P.J. Persistent reduction in brain serotonin1A receptor binding in recovered depressed men measured by positron emission tomography with [11C]WAY-100635. Mol. Psychiatry 2004, 9, 386–392. [Google Scholar] [CrossRef] [PubMed]
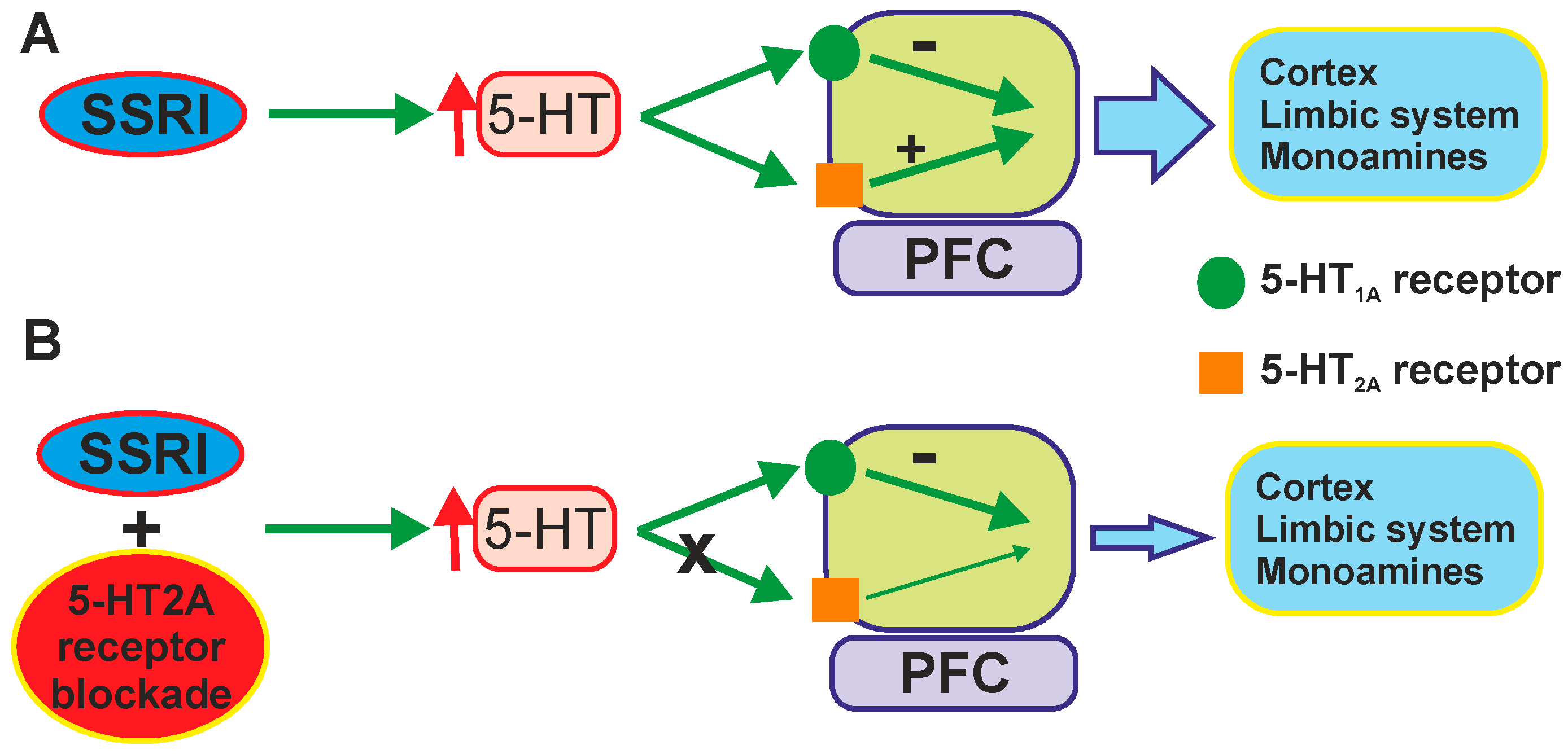
- Artigas, F.; Romero, L.; de, M.C.; Blier, P. Acceleration of the effect of selected antidepressant drugs in major depression by 5-HT1A antagonists. Trends Neurosci. 1996, 19, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Nord, M.; Finnema, S.J.; Halldin, C.; Farde, L. Effect of a single dose of escitalopram on serotonin concentration in the non-human and human primate brain. Int. J. Neuropsychopharmacol. 2013, 16, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Artigas, F.; Perez, V.; Alvarez, E. Pindolol induces a rapid improvement of depressed patients treated with serotonin reuptake inhibitors. Arch. Gen. Psychiatry 1994, 51, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Portella, M.J.; de Diego-Adelino, J.; Ballesteros, J.; Puigdemont, D.; Oller, S.; Santos, B.; Alvarez, E.; Artigas, F.; Perez, V. Can we really accelerate and enhance the selective serotonin reuptake inhibitor antidepressant effect? A randomized clinical trial and a meta-analysis of pindolol in nonresistant depression. J. Clin. Psychiatry 2011, 72, 962–969. [Google Scholar] [CrossRef] [PubMed]
- Whale, R.; Terao, T.; Cowen, P.; Freemantle, N.; Geddes, J. Pindolol augmentation of serotonin reuptake inhibitors for the treatment of depressive disorder: A systematic review. J. Psychopharmacol. 2010, 24, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Bel, N.; Artigas, F. Chronic treatment with fluvoxamine increases extracellular serotonin in frontal cortex but not in raphe nuclei. Synapse 1993, 15, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Blier, P.; Pineyro, G.; el Mansari, M.M.; Bergeron, R.; de, M.C. Role of somatodendritic 5-HT autoreceptors in modulating 5-HT neurotransmission. Ann. N. Y. Acad. Sci. 1998, 861, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Blier, P.; Ward, N.M. Is there a role for 5-HT1A agonists in the treatment of depression? Biol. Psychiatry 2003, 53, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Blier, P.; de, M.C. Modification of 5-HT neuron properties by sustained administration of the 5-HT1A agonist gepirone: Electrophysiological studies in the rat brain. Synapse 1987, 1, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Wieland, S.; Lucki, I. Antidepressant-like activity of 5-HT1A agonists measured with the forced swim test. Psychopharmacology 1990, 101, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Haddjeri, N.; Blier, P.; de, M.C. Long-term antidepressant treatments result in a tonic activation of forebrain 5-HT1A receptors. J. Neurosci. 1998, 18, 10150–10156. [Google Scholar] [PubMed]
- Scorza, M.C.; Llado-Pelfort, L.; Oller, S.; Cortes, R.; Puigdemont, D.; Portella, M.J.; Perez-Egea, R.; Alvarez, E.; Celada, P.; Perez, V.; et al. Preclinical and clinical characterization of the selective 5-HT(1A) receptor antagonist DU-125530 for antidepressant treatment. Br. J. Pharmacol. 2012, 167, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Stahl, S.M.; Kaiser, L.; Roeschen, J.; Keppel Hesselink, J.M.; Orazem, J. Effectiveness of ipsapirone, a 5-HT-1A partial agonist, in major depressive disorder: Support for the role of 5-HT-1A receptors in the mechanism of action of serotonergic antidepressants. Int. J. Neuropsychopharmacol. 1998, 1, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Sprouse, J.S.; Aghajanian, G.K. Responses of hippocampal pyramidal cells to putative serotonin 5-HT1A and 5-HT1B agonists: A comparative study with dorsal raphe neurons. Neuropharmacology 1988, 27, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, L.; Verdurand, M.; Vacher, B.; Blanc, E.; Le Bars, D.; Newman-Tancredi, A.; Zimmer, L. [18F]F15599, a novel 5-HT1A receptor agonist, as a radioligand for PET neuroimaging. Eur. J. Nucl. Med. Mol. Imaging 2010, 37, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Artigas, F. 5-HT and antidepressants: New views from microdialysis studies. Trends Pharmacol. Sci. 1993, 14, 262. [Google Scholar] [CrossRef]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef]
- Duman, R.S.; Monteggia, L.M. A neurotrophic model for stress-related mood disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Page, M.E.; Cryan, J.F.; Sullivan, A.; Dalvi, A.; Saucy, B.; Manning, D.R.; Lucki, I. Behavioral and neurochemical effects of 5-(4-[4-(5-Cyano-3-indolyl)-butyl)-butyl]-1-piperazinyl)-benzofuran-2-carboxamide (EMD 68843): A combined selective inhibitor of serotonin reuptake and 5-hydroxytryptamine(1A) receptor partial agonist. J. Pharmacol. Exp. Ther. 2002, 302, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Mork, A.; Pehrson, A.; Brennum, L.T.; Nielsen, S.M.; Zhong, H.; Lassen, A.B.; Miller, S.; Westrich, L.; Boyle, N.J.; Sanchez, C.; et al. Pharmacological effects of Lu AA21004: A novel multimodal compound for the treatment of major depressive disorder. J. Pharmacol. Exp. Ther. 2012, 340, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Mayorga, A.J.; Dalvi, A.; Page, M.E.; Zimov-Levinson, S.; Hen, R.; Lucki, I. Antidepressant-like behavioral effects in 5-hydroxytryptamine(1A) and 5-hydroxytryptamine(1B) receptor mutant mice. J. Pharmacol. Exp. Ther. 2001, 298, 1101–1107. [Google Scholar] [PubMed]
- Duman, R.S. Depression: A case of neuronal life and death? Biol. Psychiatry 2004, 56, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Lucas, G.; Rymar, V.V.; Du, J.; Mnie-Filali, O.; Bisgaard, C.; Manta, S.; Lambas-Senas, L.; Wiborg, O.; Haddjeri, N.; Pineyro, G.; et al. Serotonin(4) (5-HT(4)) receptor agonists are putative antidepressants with a rapid onset of action. Neuron 2007, 55, 712–725. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, D.V.; Gage, F.H. Neurogenesis and neuroadaptation. Neuromol. Med. 2004, 5, 1–9. [Google Scholar] [CrossRef]
- Santarelli, L.; Saxe, M.; Gross, C.; Surget, A.; Battaglia, F.; Dulawa, S.; Weisstaub, N.; Lee, J.; Duman, R.; Arancio, O.; et al. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 2003, 301, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.L.; van Praag, H.; Gage, F.H. Adult brain neurogenesis and psychiatry: A novel theory of depression. Mol. Psychiatry. 2000, 5, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.; Fricker, A.D.; Devi, L.A.; Gomes, I. Mechanisms of action of antidepressants: From neurotransmitter systems to signaling pathways. Cell Signal. 2005, 17, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Blier, P.; de, M.C. Current advances and trends in the treatment of depression. Trends Pharmacol. Sci. 1994, 15, 220–226. [Google Scholar] [CrossRef]
- Neumaier, J.F.; Root, D.C.; Hamblin, M.W. Chronic fluoxetine reduces serotonin transporter mRNA and 5-HT1B mRNA in a sequential manner in the rat dorsal raphe nucleus. Neuropsychopharmacology 1996, 15, 515–522. [Google Scholar] [CrossRef]
- Gobert, A.; Rivet, J.M.; Cistarelli, L.; Millan, M.J. Potentiation of the fluoxetine-induced increase in dialysate levels of serotonin (5-HT) in the frontal cortex of freely moving rats by combined blockade of 5-HT1A and 5-HT1B receptors with WAY 100,635 and GR 127,935. J. Neurochem. 1997, 68, 1159–1163. [Google Scholar] [CrossRef] [PubMed]
- Gardier, A.M.; Trillat, A.C.; Malagie, I.; David, D.; Hascoet, M.; Colombel, M.C.; Jolliet, P.; Jacquot, C.; Hen, R.; Bourin, M. 5-HT1B serotonin receptors and antidepressant effects of selective serotonin reuptake inhibitors. C. R. Acad. Sci. III 2001, 324, 433–441. [Google Scholar] [PubMed]
- Artigas, F.; Celada, P.; Laruelle, M.; Adell, A. How does pindolol improve antidepressant action? Trends Pharmacol. Sci. 2001, 22, 224–228. [Google Scholar] [PubMed]
- Albert, P.R.; Vahid-Ansari, F.; Luckhart, C. Serotonin-prefrontal cortical circuitry in anxiety and depression phenotypes: Pivotal role of pre- and post-synaptic 5-HT1A receptor expression. Front. Behav. Neurosci. 2014, 8, 199. [Google Scholar] [PubMed]
- De Almeida, J.; Mengod, G. Serotonin 1A receptors in human and monkey prefrontal cortex are mainly expressed in pyramidal neurons and in a GABAergic interneuron subpopulation: Implications for schizophrenia and its treatment. J. Neurochem. 2008, 107, 488–496. [Google Scholar] [PubMed]
- Santana, N.; Bortolozzi, A.; Serrats, J.; Mengod, G.; Artigas, F. Expression of serotonin1A and serotonin2A receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb. Cortex 2004, 14, 1100–1109. [Google Scholar] [PubMed]
- Amargos-Bosch, M.; Bortolozzi, A.; Puig, M.V.; Serrats, J.; Adell, A.; Celada, P.; Toth, M.; Mengod, G.; Artigas, F. Co-expression and in vivo interaction of serotonin1A and serotonin2A receptors in pyramidal neurons of prefrontal cortex. Cereb. Cortex 2004, 14, 281–299. [Google Scholar] [PubMed]
- Bevilacqua, L.; Doly, S.; Kaprio, J.; Yuan, Q.; Tikkanen, R.; Paunio, T.; Zhou, Z.; Wedenoja, J.; Maroteaux, L.; Diaz, S.; et al. A population-specific HTR2B stop codon predisposes to severe impulsivity. Nature 2010, 468, 1061–1066. [Google Scholar] [PubMed]
- Launay, J.M.; Schneider, B.; Loric, S.; Da, P.M.; Kellermann, O. Serotonin transport and serotonin transporter-mediated antidepressant recognition are controlled by 5-HT2B receptor signaling in serotonergic neuronal cells. FASEB J. 2006, 20, 1843–1854. [Google Scholar] [PubMed]
- Doly, S.; Bertran-Gonzalez, J.; Callebert, J.; Bruneau, A.; Banas, S.M.; Belmer, A.; Boutourlinsky, K.; Herve, D.; Launay, J.M.; Maroteaux, L. Role of serotonin via 5-HT2B receptors in the reinforcing effects of MDMA in mice. PLoS ONE 2009, 4, e7952. [Google Scholar] [PubMed]
- Doly, S.; Valjent, E.; Setola, V.; Callebert, J.; Herve, D.; Launay, J.M.; Maroteaux, L. Serotonin 5-HT2B receptors are required for 3,4-methylenedioxymethamphetamine-induced hyperlocomotion and 5-HT release in vivo and in vitro. J. Neurosci. 2008, 28, 2933–2940. [Google Scholar] [PubMed]
- Banas, S.M.; Doly, S.; Boutourlinsky, K.; Diaz, S.L.; Belmer, A.; Callebert, J.; Collet, C.; Launay, J.M.; Maroteaux, L. Deconstructing antiobesity compound action: Requirement of serotonin 5-HT2B receptors for dexfenfluramine anorectic effects. Neuropsychopharmacology 2011, 36, 423–433. [Google Scholar] [PubMed]
- Diaz, S.L.; Maroteaux, L. Implication of 5-HT (2B) receptors in the serotonin syndrome. Neuropharmacology 2011, 61, 4954502. [Google Scholar]
- Diaz, S.L.; Doly, S.; Narboux-Neme, N.; Fernandez, S.; Mazot, P.; Banas, S.M.; Boutourlinsky, K.; Moutkine, I.; Belmer, A.; Roumier, A.; et al. 5-HT(2B) receptors are required for serotonin-selective antidepressant actions. Mol. Psychiatry 2012, 17, 154–163. [Google Scholar] [PubMed]
- Rosenzweig-Lipson, S.; Sabb, A.; Stack, G.; Mitchell, P.; Lucki, I.; Malberg, J.E.; Grauer, S.; Brennan, J.; Cryan, J.F.; Sukoff Rizzo, S.J.; et al. Antidepressant-like effects of the novel, selective, 5-HT2C receptor agonist WAY-163909 in rodents. Psychopharmacology (Berl.) 2007, 192, 159–170. [Google Scholar] [PubMed]
- Cryan, J.F.; Lucki, I. Antidepressant-like behavioral effects mediated by 5-Hydroxytryptamine (2C) receptors. J. Pharmacol. Exp. Ther. 2000, 295, 1120–1126. [Google Scholar] [PubMed]
- Hervas, I.; Vilaro, M.T.; Romero, L.; Scorza, M.C.; Mengod, G.; Artigas, F. Desensitization of 5-HT(1A) autoreceptors by a low chronic fluoxetine dose effect of the concurrent administration of WAY-100635. Neuropsychopharmacology 2001, 24, 11–20. [Google Scholar] [PubMed]
- Riad, M.; Zimmer, L.; Rbah, L.; Watkins, K.C.; Hamon, M.; Descarries, L. Acute treatment with the antidepressant fluoxetine internalizes 5-HT1A autoreceptors and reduces the in vivo binding of the PET radioligand [18F]MPPF in the nucleus raphe dorsalis of rat. J. Neurosci. 2004, 24, 5420–5426. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chilmonczyk, Z.; Bojarski, A.J.; Pilc, A.; Sylte, I. Functional Selectivity and Antidepressant Activity of Serotonin 1A Receptor Ligands. Int. J. Mol. Sci. 2015, 16, 18474-18506. https://doi.org/10.3390/ijms160818474
Chilmonczyk Z, Bojarski AJ, Pilc A, Sylte I. Functional Selectivity and Antidepressant Activity of Serotonin 1A Receptor Ligands. International Journal of Molecular Sciences. 2015; 16(8):18474-18506. https://doi.org/10.3390/ijms160818474
Chicago/Turabian StyleChilmonczyk, Zdzisław, Andrzej Jacek Bojarski, Andrzej Pilc, and Ingebrigt Sylte. 2015. "Functional Selectivity and Antidepressant Activity of Serotonin 1A Receptor Ligands" International Journal of Molecular Sciences 16, no. 8: 18474-18506. https://doi.org/10.3390/ijms160818474