An Experimental Hut Evaluation of PBO-Based and Pyrethroid-Only Nets against the Malaria Vector Anopheles funestus Reveals a Loss of Bed Nets Efficacy Associated with GSTe2 Metabolic Resistance

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Hut Design
2.3. Net Treatment/Arm Comparison
2.3.1. Hut Effect
2.3.2. Bioassays and Cone Assays
2.4. Experimental Hut Trial
2.4.1. Mosquito Collection
2.4.2. Bed Nets Performance Assessment
- Deterrence/entry rate: the reduction in hut entry relative to control. Deterrence (%) = 100 × (Du − Dt)/Du, where Du is the total number of mosquitoes found in untreated hut (control) and Dt is the total number of mosquitoes in the treated hut.
- Entry rate (%) = 100 × (Ht/Hn) where Ht is the total number of mosquitoes found in the hut and Hn is the total number of mosquitoes collected in all the 5 huts.
- Exophily (Excito-repellency): the proportion of mosquitoes found exited in the veranda trap Exophily (%) = 100 × (Ev/Et) where Ev is the total number of mosquitoes fund in veranda and Et is the total number of both inside the hut and veranda.
- Blood feeding rate (BFR). This rate was calculated as follows: Blood feeding rate = (N mosquitoes fed) × 100/total N mosquitoes. Where “N mosquitoes fed” was the number of mosquitoes fed, and “total N mosquitoes” was the total number of mosquitoes collected.
- Blood-feeding inhibition (BFI): the reduction in blood-feeding in comparison with the control hut. Blood feeding inhibition is an indicator of personal protection (PP). More precisely, the personal protection effect of each bed net is the reduction of blood feeding percentage induced by the net when compared to control. The protective effect of each bed net can be calculated as follows:
- Personal protection (%) = 100 × (Bu − Bt)/Bu, where Bu is the total number of blood-fed mosquitoes in the huts with untreated nets and Bt is the total number of blood-fed mosquitoes in the huts with treated nets [23].
- Immediate and delay mortality: the proportion of mosquitoes entering the hut that are found dead in the morning (immediate mortality) or after being caught alive and held for 24 h with access to sugar solution (delay mortality) [23]. In this study we focused on the overall mortality calculated as follows: Mortality (%) = 100 × (Mt/MT) where Mt is the total number of mosquitoes found dead in the hut and MT is the total number of mosquitoes collected in the hut.
2.4.3. Ethical Clearance
2.5. Impact of the L119F-GSTe2 Mutation on Insecticide-Treated Nets
2.6. Genotyping
2.7. Data Analysis
2.7.1. Experimental Hut Trial
2.7.2. Test of Association between L119F-Mutation and the Entomological Outcomes
2.7.3. Hut Effect Analysis
3. Results
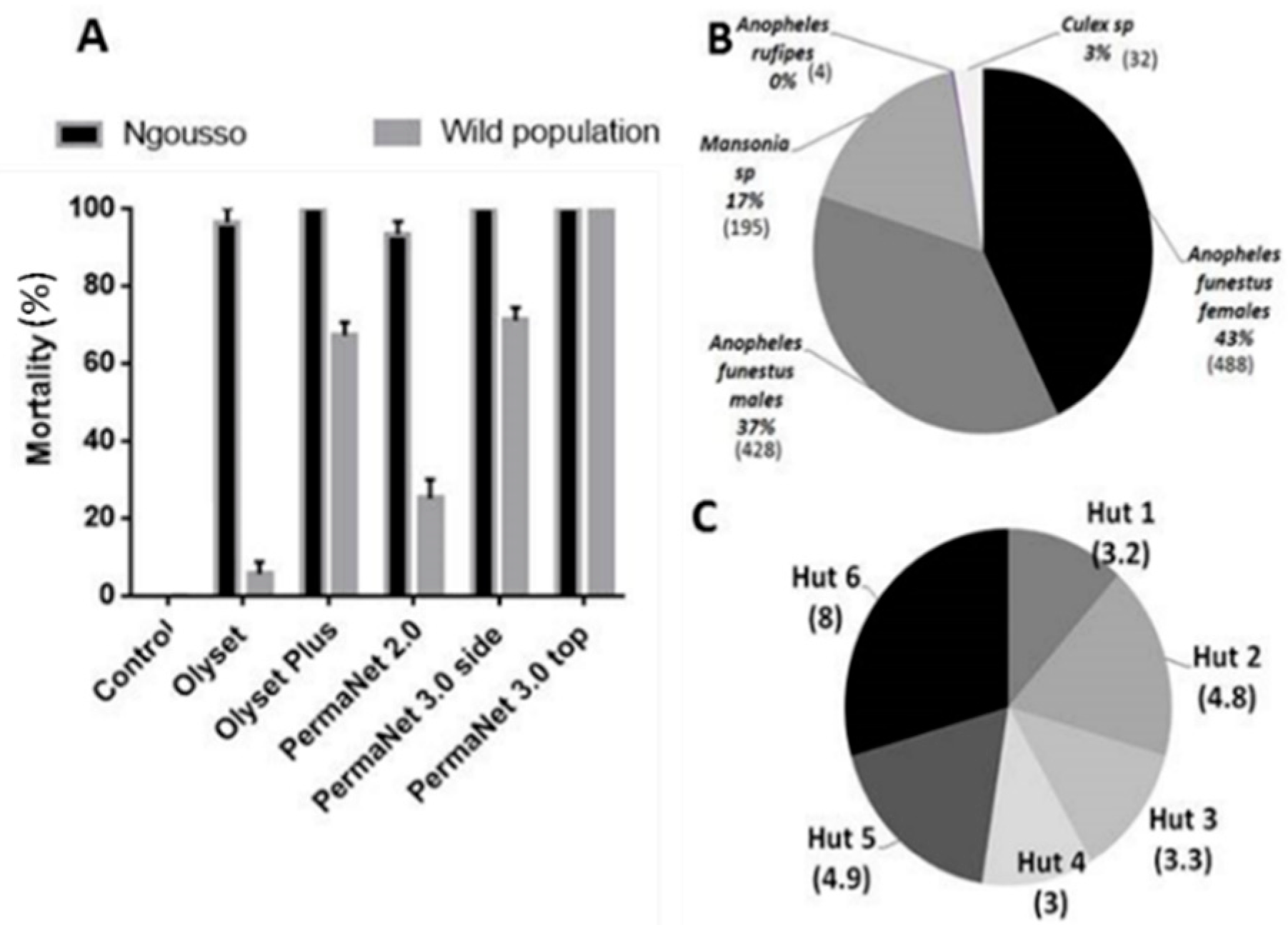
3.1. Cone Assays Using the An. gambiae Susceptible Lab Strain Ngousso
3.2. Cone Assays with An. Funestus from the Field (Mibellon)
3.3. Hut Effect
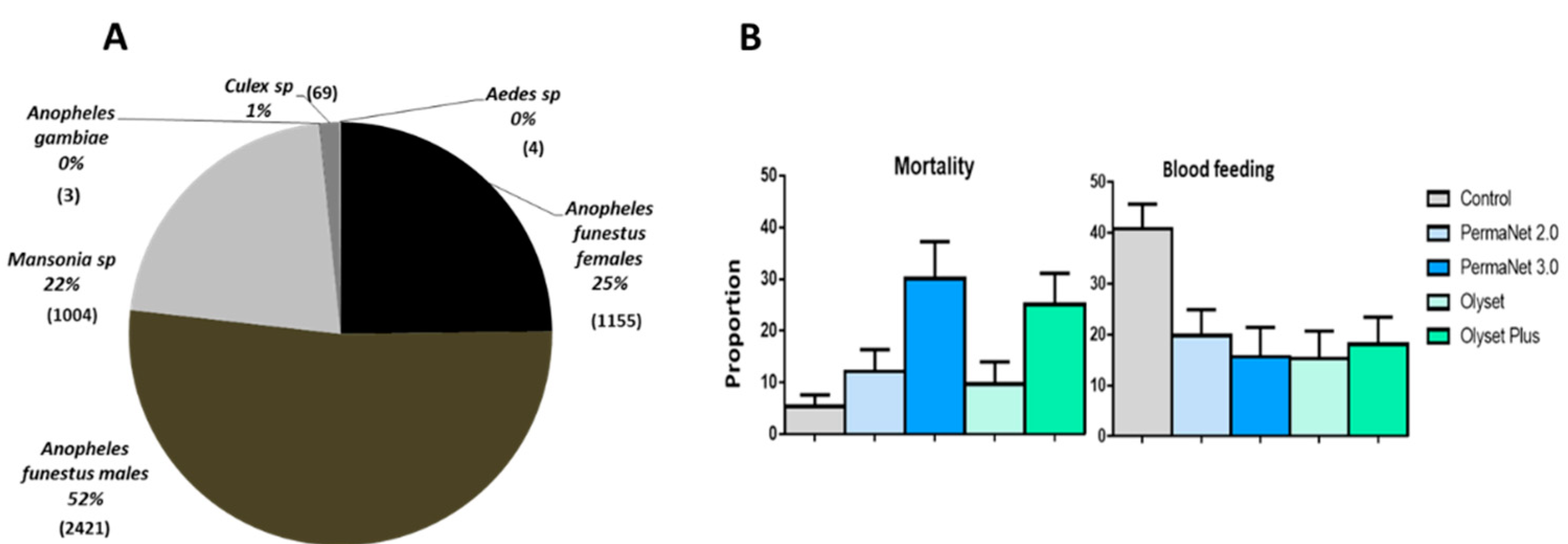
3.4. Mosquito Abundance
3.5. Performance of the Nets against An. Funestus s.s. Population
3.5.1. Deterrent Effect/Entry Rate
3.5.2. Induced Exophily Rate
3.5.3. Mortality (Overall Mortality)
3.5.4. Blood Feeding Inhibition (BFI)
3.5.5. Personal Protection (PP)
3.6. Performance of the Nets against Mansonia spp. Population
3.6.1. Deterrent Effect
3.6.2. Induced Exophily Rate
3.6.3. Mortality
3.6.4. Blood Feeding Inhibition (BFI)
3.6.5. Personal Protection (PP)
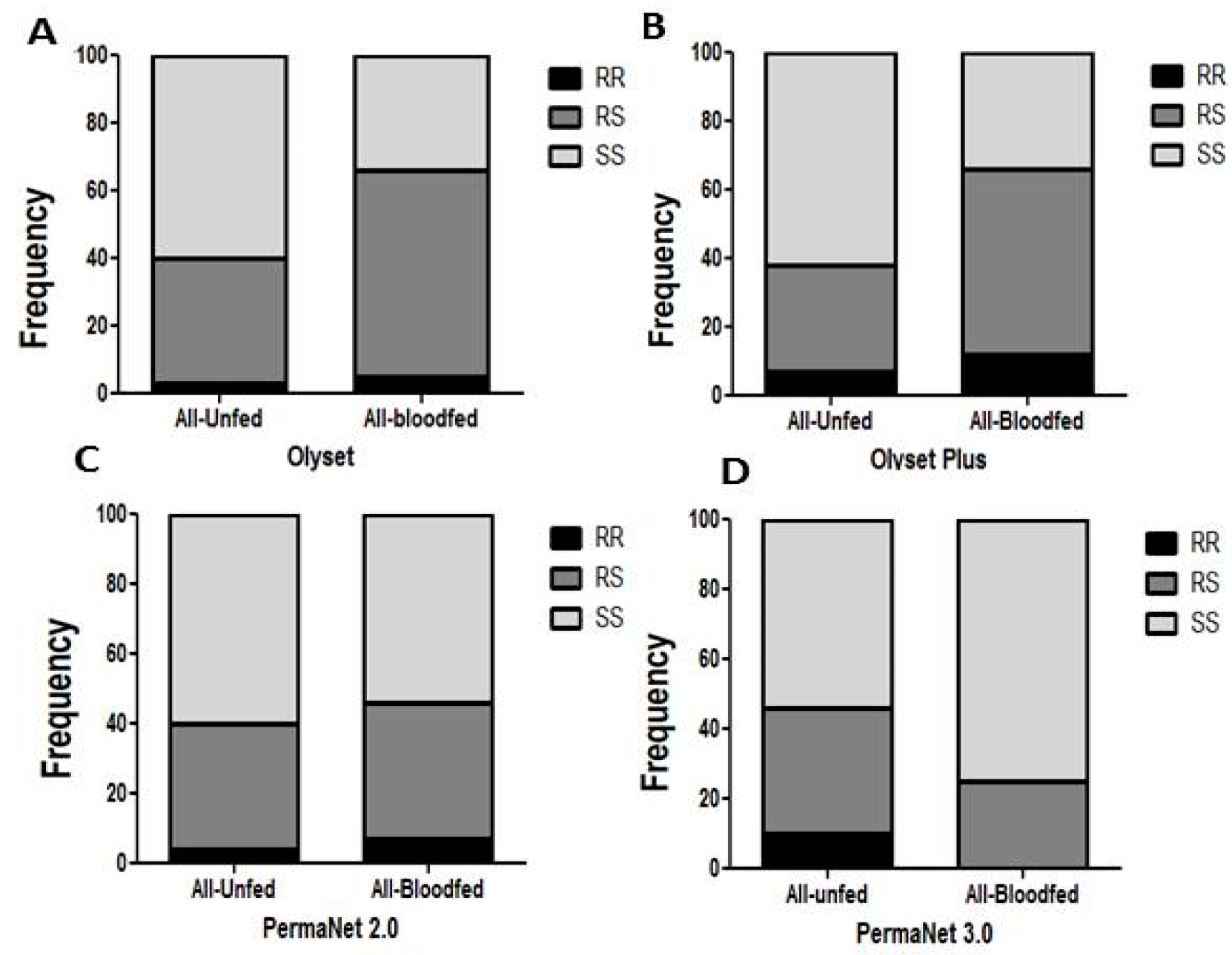
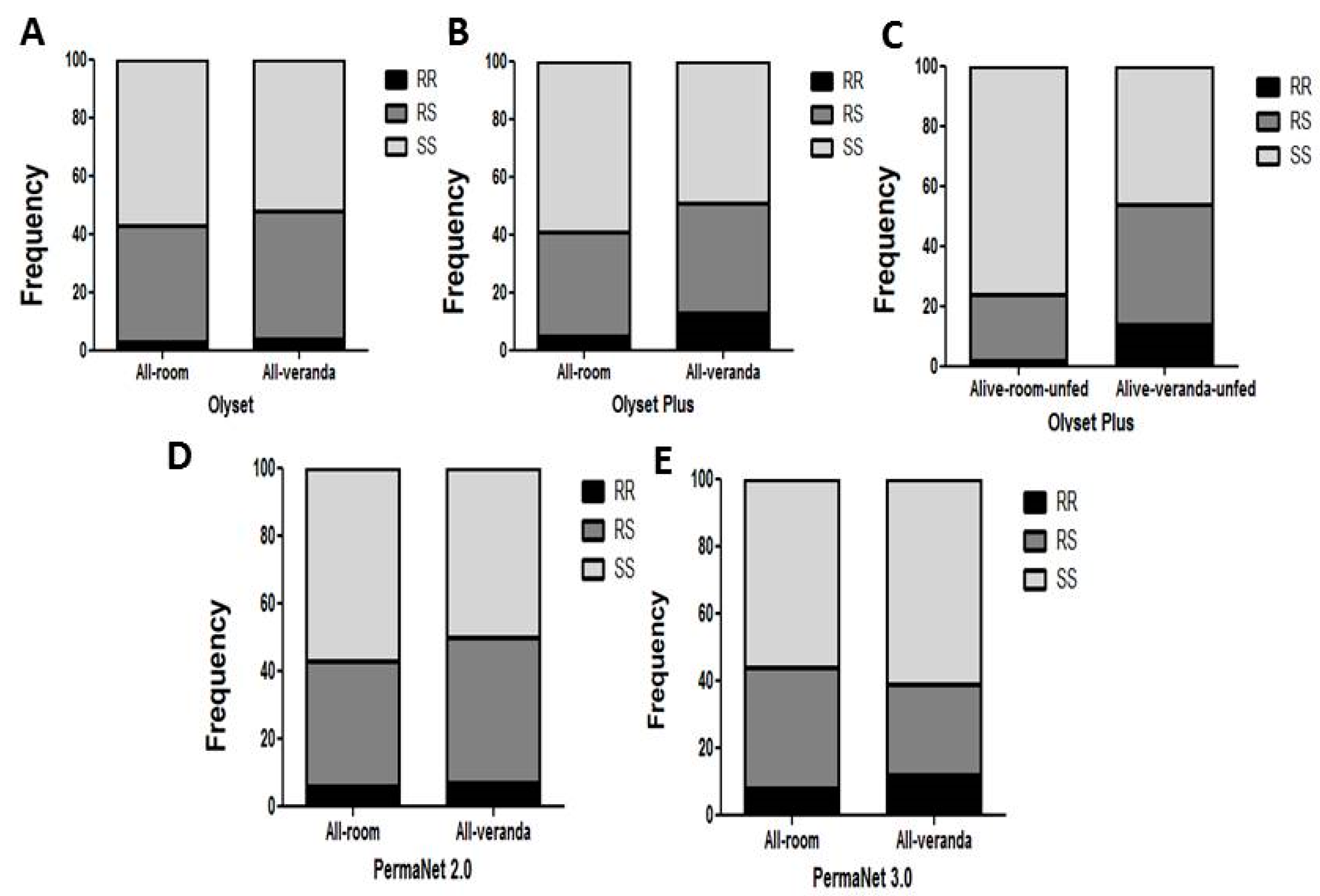
3.7. Comparative Analysis of the Impact of L119F-GSTe2 Mutation on the Efficacy of Conventional and PBO-Based Nets
Impact on Mortality
3.8. Impact on Blood Feeding
3.9. Impact on Exophily
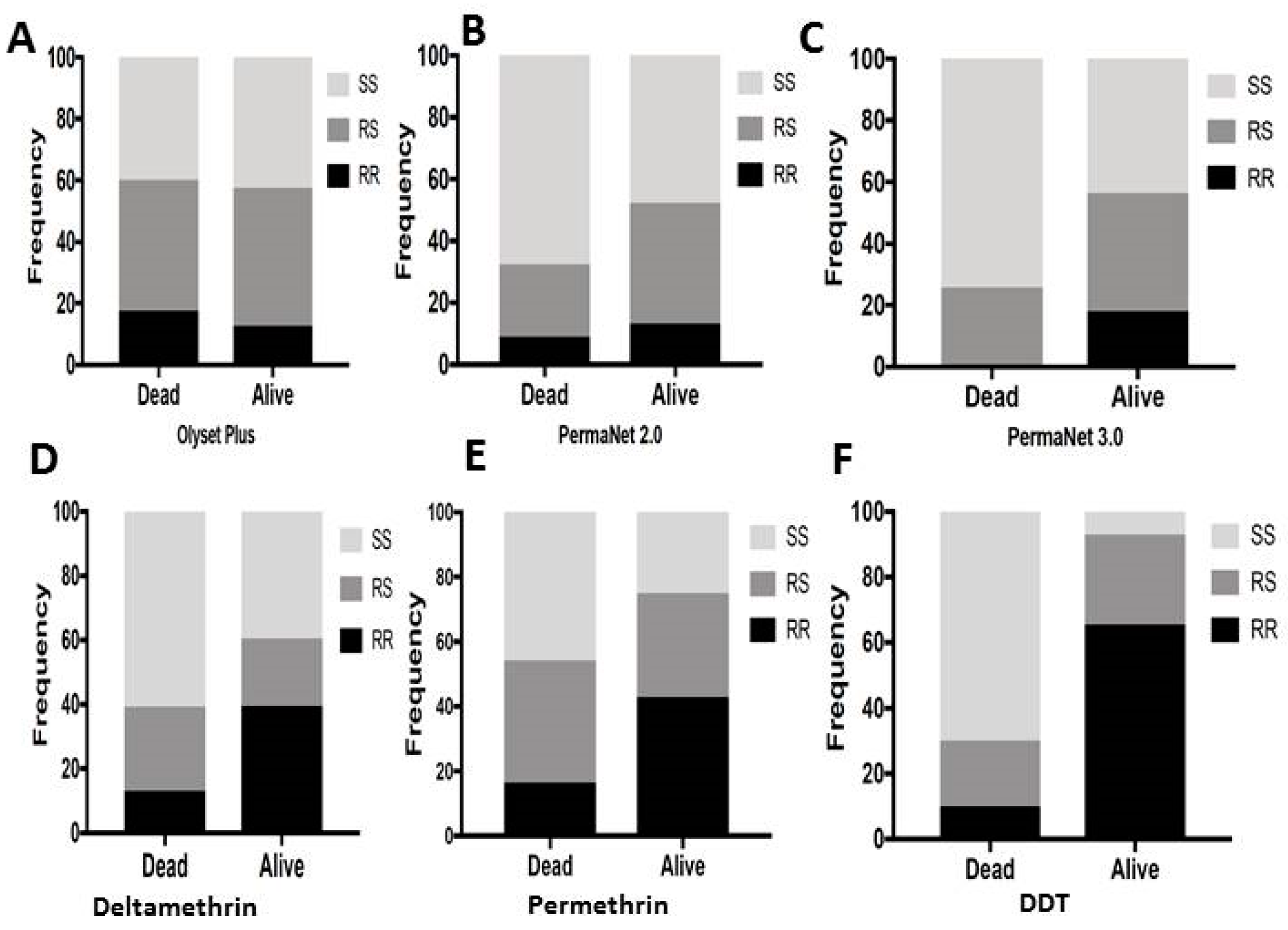
3.10. Correlation between L119F-GSTe2 and Mortality from Cone Assays
3.10.1. Olyset Plus
3.10.2. PermaNet 2.0
3.10.3. PermaNet 3.0
3.11. Validation of the Association between L119F-GSTe2 and Resistance to Pyrethroids and DDT in Mibellon
4. Discussion
4.1. PBO plus pyrethroid-Based Nets are More Effective than Pyrethroid-Only Nets against Pyrethroid Resistant An. funestus
4.1.1. Mortality
4.1.2. Blood Feeding
4.1.3. Exophily
4.2. GSTe2 Mediated Metabolic Resistance Is Reducing Efficacy of LLINs
4.2.1. Experimental Hut Study
4.2.2. Impact from Cone Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.E.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinschmidt, I.; Bradley, J.; Knox, T.B.; Mnzava, A.P.; Kafy, H.T.; Mbogo, C.; Ismail, B.A.; Bigoga, J.D.; Adechoubou, A.; Raghavendra, K.; et al. Implications of insecticide resistance for malaria vector control with long-lasting insecticidal nets: A WHO-coordinated, prospective, international, observational cohort study. Lancet Infect. Dis. 2018, 18, 640–649. [Google Scholar] [CrossRef] [Green Version]
- Protopopoff, N.; Mosha, J.F.; Lukole, E.; Charlwood, J.D.; Wright, A.; Mwalimu, C.D.; Manjurano, A.; Mosha, F.W.; Kisinza, W.; Kleinschmidt, I.; et al. Effectiveness of a long-lasting piperonyl butoxide-treated insecticidal net and indoor residual spray interventions, separately and together, against malaria transmitted by pyrethroid-resistant mosquitoes: A cluster, randomised controlled, two-by-two factorial design trial. Lancet 2018, 391, 1577–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbel, V.; Chabi, J.; Dabire, R.K.; Etang, J.; Nwane, P.; Pigeon, O.; Akogbeto, M.; Hougard, J.M. Field efficacy of a new mosaic long-lasting mosquito net (PermaNet® 3.0) against pyrethroid-resistant malaria vectors: A multi centre study in Western and Central Africa. Malar. J. 2010, 9, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- N’Guessan, R.; Asidi, A.; Boko, P.; Odjo, A.; Akogbeto, M.; Pigeon, O.; Rowland, M. An experimental hut evaluation of PermaNet® 3.0, a deltamethrin-piperonyl butoxide combination net, against pyrethroid-resistant Anopheles gambiae and Culex quinquefasciatus mosquitoes in southern Benin. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 758–765. [Google Scholar] [CrossRef]
- Pennetier, C.; Bouraima, A.; Chandre, F.; Piameu, M.; Etang, J.; Rossignol, M.; Sidick, I.; Zogo, B.; Lacroix, M.N.; Yadav, R.; et al. Efficacy of Olyset® Plus, a new long-lasting insecticidal net incorporating permethrin and piperonyl-butoxide against multi-resistant malaria vectors. PLoS ONE 2013, 8, e75134. [Google Scholar] [CrossRef]
- World Health Organization (WHO). World Malaria Report 2017; WHO: Geneva, Switzerland, 2017; Licencce: CC BY-NC-SA 3.0 IGO. [Google Scholar]
- Riveron, J.M.; Yunta, C.; Ibrahim, S.S.; Djouaka, R.; Irving, H.; Menze, B.D.; Ismail, H.M.; Hemingway, J.; Ranson, H.; Albert, A.; et al. A single mutation in the GSTe2 gene allows tracking of metabolically-based insecticide resistance in a major malaria vector. Genome Biol. 2014, 15, R27. [Google Scholar] [CrossRef] [Green Version]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 369–389. [Google Scholar] [CrossRef]
- Mitchell, S.N.; Rigden, D.J.; Dowd, A.J.; Lu, F.; Wilding, C.S.; Weetman, D.; Dadzie, S.; Jenkins, A.M.; Regna, K.; Boko, P.; et al. Metabolic and target-site mechanisms combine to confer strong DDT resistance in Anopheles gambiae. PLoS ONE 2014, 9, e92662. [Google Scholar] [CrossRef]
- Lumjuan, N.; Rajatileka, S.; Changsom, D.; Wicheer, J.; Leelapat, P.; Prapanthadara, L.A.; Somboon, P.; Lycett, G.; Ranson, H. The role of the Aedes aegypti Epsilon glutathione transferases in conferring resistance to DDT and pyrethroid insecticides. Insect. Biochem. Mol. Biol. 2011, 41, 203–209. [Google Scholar] [CrossRef]
- Tchouakui, M.; Chiang, M.C.; Ndo, C.; Kuicheu, C.K.; Amvongo-Adjia, N.; Wondji, M.J.; Tchoupo, M.; Kusimo, M.O.; Riveron, J.M.; Wondji, C.S. A marker of glutathione S-transferase-mediated resistance to insecticides is associated with higher Plasmodium infection in the African malaria vector Anopheles funestus. Sci. Rep. 2019, 9, 5772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riveron, J.M.; Watsenga, F.; Irving, H.; Irish, S.R.; Wondji, C.S. High Plasmodium Infection Rate and Reduced Bed Net Efficacy in Multiple Insecticide-Resistant Malaria Vectors in Kinshasa, Democratic Republic of Congo. J. Infect. Dis. 2017, 217, 320–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weedall, G.D.; Mugenzi, L.M.; Menze, B.D.; Tchouakui, M.; Ibrahim, S.S.; Amvongo-Adjia, N.; Irving, H.; Wondji, M.J.; Tchoupo, M.; Djouaka, R. A cytochrome P450 allele confers pyrethroid resistance on a major African malaria vector, reducing insecticide-treated bednet efficacy. Sci. Transl. Med. 2019, 11, eaat7386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riveron, J.M.; Irving, H.; Ndula, M.; Barnes, K.G.; Ibrahim, S.S.; Paine, M.J.; Wondji, C.S. Directionally selected cytochrome P450 alleles are driving the spread of pyrethroid resistance in the major malaria vector Anopheles funestus. Proc. Natl. Acad. Sci. USA 2013, 110, 252–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irving, H.; Wondji, C.S. Investigating knockdown resistance (kdr) mechanism against pyrethroids/DDT in the malaria vector Anopheles funestus across Africa. BMC Genet. 2017, 18, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menze, B.D.; Riveron, J.M.; Ibrahim, S.S.; Irving, H.; Antonio-Nkondjio, C.; Awono-Ambene, P.H.; Wondji, C.S. Multiple Insecticide Resistance in the Malaria Vector Anopheles funestus from Northern Cameroon Is Mediated by Metabolic Resistance Alongside Potential Target Site Insensitivity Mutations. PLoS ONE 2016, 11, e0163261. [Google Scholar] [CrossRef]
- Menze, B.D.; Wondji, M.J.; Tchapga, W.; Tchoupo, M.; Riveron, J.M.; Wondji, C.S. Bionomics and insecticides resistance profiling of malaria vectors at a selected site for experimental hut trials in central Cameroon. Malar. J. 2018, 17, 317. [Google Scholar] [CrossRef]
- N’Guessan, R.; Corbel, V.; Akogbéto, M.; Rowland, M. Reduced efficacy of insecticide-treated nets and indoor residual spraying for malaria control in pyrethroid resistance area, Benin. Emerg. Infect. Dis. 2007, 13, 199. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global Plan for Insecticide Resistance Management in Malaria Vectors; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- Ngufor, C.; Chouaibou, M.; Tchicaya, E.; Loukou, B.; Kesse, N.; N’Guessan, R.; Johnson, P.; Koudou, B.; Rowland, M. Combining organophosphate-treated wall linings and long-lasting insecticidal nets fails to provide additional control over long-lasting insecticidal nets alone against multiple insecticide-resistant Anopheles gambiae in Cote d’Ivoire: An experimental hut trial. Malar. J. 2014, 13, 396. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- World Health Organization (WHO). Guidelines for Laboratory and Field-Testing of Long-Lasting Insecticidal Nets; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Gillies, M.; Coetzee, M. A Supplement to the Anophelinae of Africa South of the Sahara (Afrotropical Region). Publ. S. Afr. Inst. Med. Res. 1987, 55, 1–143. [Google Scholar]
- Koekemoer, L.L.; Kamau, L.; Hunt, R.H.; Coetzee, M. A cocktail polymerase chain reaction assay to identify members of the Anopheles funestus (Diptera: Culicidae) group. Am. J. Trop. Med. Hyg. 2002, 66, 804–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J. Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics 1984, 107, 611–634. [Google Scholar] [PubMed]
- Tchouakui, M.; Riveron, J.M.; Djonabaye, D.; Tchapga, W.; Irving, H.; Takam, P.S.; Njiokou, F.; Wondji, C.S. Fitness Costs of the Glutathione S-Transferase Epsilon 2 (L119F-GSTe2) Mediated Metabolic Resistance to Insecticides in the Major African Malaria Vector Anopheles Funestus. Genes 2018, 9, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchouakui, M.; Fossog, B.T.; Ngannang, B.V.; Djonabaye, D.; Tchapga, W.; Njiokou, F.; Wondji, C. Investigation of the influence of a glutathione S-transferase metabolic resistance to pyrethroids/DDT on mating competitiveness in males Anopheles funestus, African malaria vector. Wellcome Open Res. 2019, 4, 13. [Google Scholar]
- Badolo, A.; Guelbéogo, W.M.; Tiono, A.B.; Traoré, A.; Sirima, S.B. Laboratory evaluation of Fendona 6SC® treated bednets and Interceptor® long-lasting nets against Anopheles gambiae s.l. in Burkina Faso. Parasitol. Res. 2014, 113, 1069–1075. [Google Scholar] [CrossRef]
- Chouaibou, M.; Simard, F.; Chandre, F.; Etang, J.; Darriet, F.; Hougard, J.M. Efficacy of bifenthrin-impregnated bednets against Anopheles funestus and pyrethroid-resistant Anopheles gambiae in North Cameroon. Malar. J. 2006, 5, 77. [Google Scholar] [CrossRef] [Green Version]
- Toe, K.H.; Muller, P.; Badolo, A.; Traore, A.; Sagnon, N.; Dabire, R.K.; Ranson, H. Do bednets including piperonyl butoxide offer additional protection against populations of Anopheles gambiae s.l. that are highly resistant to pyrethroids? An experimental hut evaluation in Burkina Fasov. Med. Vet. Entomol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Koudou, B.G.; Koffi, A.A.; Malone, D.; Hemingway, J. Efficacy of PermaNet® 2.0 and PermaNet® 3.0 against insecticide-resistant Anopheles gambiae in experimental huts in Cote d’Ivoire. Malar. J. 2011, 10, 172. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.P.; Sabesan, S.; Panicker, K. The resting and house frequenting behavior of Mansonia annulifera, Ma. uniformis and Ma. indiana, the vectors of Malayan filariasis in Kerala State, India. Southeast Asian J. Trop. Med. Public Health 1992, 23, 324–327. [Google Scholar]
- Mugenzi, L.M.; Menze, B.D.; Tchouakui, M.; Wondji, M.J.; Irving, H.; Tchoupo, M.; Hearn, J.; Weedall, G.D.; Riveron, J.M.; Wondji, C.S. Cis-regulatory CYP6P9b P450 variants associated with loss of insecticide-treated bed net efficacy against Anopheles funestus. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Riveron, J.M.; Ibrahim, S.S.; Chanda, E.; Mzilahowa, T.; Cuamba, N.; Irving, H.; Barnes, K.G.; Ndula, M.; Wondji, C.S. The highly polymorphic CYP6M7 cytochrome P450 gene partners with the directionally selected CYP6P9a and CYP6P9b genes to expand the pyrethroid resistance front in the malaria vector Anopheles funestus in Africa. BMC Genom. 2014, 15, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riveron, J.M.; Osae, M.; Egyir-Yawson, A.; Irving, H.; Ibrahim, S.S.; Wondji, C.S. Multiple insecticide resistance in the major malaria vector Anopheles funestus in southern Ghana: Implications for malaria control. Parasites Vectors 2016, 9, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berticat, C.; Duron, O.; Heyse, D.; Raymond, M. Insecticide resistance genes confer a predation cost on mosquitoes, Culex pipiens. Genet. Res. 2004, 83, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Arm | Description | Manufacturer |
|---|---|---|
| Untreated | 100% polyester with no insecticide | Local market |
| Olyset | 8.6 × 10−4 kg/m2 (2%) of permethrin incorporated into polyethylene | Sumitomo Chemical |
| Olyset Plus | 8.6 × 10−4 kg/m2 (2%) of permethrin and 4.3 × 10−4 kg/m2 (1%) of Piperonyl butoxide (PBO) incorporated into polyethylene | Sumitomo Chemical |
| PermaNet 2.0 | 100% polyester coated with 1.8 g/kg of deltamethrin | Vestergaard Frandsen |
| PermaNet 3.0 | Combination of 2.8 g/kg of deltamethrin coated on polyester with strengthened border (side panels) and deltamethrin (4.0 g/kg) and PBO (25 g/kg) | Vestergaard Frandsen |
| Treatments | |||||
|---|---|---|---|---|---|
| Control | PermaNet 2.0 | PermaNet 3.0 | Olyset | Olyset Plus | |
| Females caught | 390 | 237 | 153 | 176 | 199 |
| Exophily% | 6.9 | 17.7 *** | 17.0 *** | 29.0 *** | 22.1 *** |
| 95% Confidence limits | 4.40–9.44) | (12.85–22.58) | (11.04–22.94) | (22.27–35.68) | (16.34–27.88) |
| Blood fed (%) | 40.8 | 19.8 *** | 15.7 *** | 15.3 *** | 17.2 *** |
| 95% Confidence limits | (35.89–45.65) | (14.75–24.91) | 9.92–21.45) | 10.02–20.67) | (12.74–23.44) |
| Blood feed inh. (%) | 0.0 | 51.36 | 61.52 | 62.37 | 55.63 |
| Personal protection (%) | 0.0 | 70.44 | 84.90 | 83.01 | 77.35 |
| Overall mortality (%) | 5.4 | 12.2 ** | 30.1 *** | 9.7 * | 25.1 *** |
| 95% Confidence limits | (31.14–7.62) | (8.06–16.41) | (22.80–37.33) | (5.29–14.02) | (19.10–31.15) |
| Immediate mortality | 2.6 | 5.4 | 27.6 *** | 6.1 * | 18.3 *** |
| entry rate | 33.7 | 20.5 * | 13.2 *** | 15.2 *** | 13.6 *** |
| Deterrence (%) | 0.0 | 39.2 | 60.8 | 54.9 | 49.0 |
| Control | PermaNet 2.0 | PermaNet 3.0 | Olyset | Olyset Plus | |
|---|---|---|---|---|---|
| Total | 342 | 142 | 216 | 141 | 163 |
| Exophily% | 42.4 | 37.3 * | 31.9 ** | 28.4 ** | 31.3 ** |
| 95% Confidence limits | (37.16–47.64) | (29.37–45.28) | (25.73–38.16) | (20.93–35.81) | (24.17–38.41) |
| Blood feeding (%) | 30.1 | 16.2 ** | 9.3 *** | 15.6 *** | 20.2 ** |
| 95% Confidence limits | (25.25–34.98) | (10.14–22.26) | (5.39–13.12) | (9.61–21.59) | (14.08–26.41) |
| Blood feeding inh. (%) | 0.0 | 46.22 | 69.26 | 48.19 | 32.78 |
| Personal protection (%) | 0.0 | 77.66 | 80.53 | 78.64 | 67.96 |
| Overall mortality (%) | 37.4 | 51.4 ** | 65.5 *** | 40.4 ** | 68.1 *** |
| Immediate mortality | 36.8 | 46.5 | 62.0 *** | 39.0 *** | 65.0 *** |
| Entry rate | 34.0 | 14.1 *** | 21.5 ** | 14.0 *** | 16.2 *** |
| Deterrence (%) | 0.0 | 58.5 | 36.8 | 58.8 | 52.3 |
| Mortality | ||||
|---|---|---|---|---|
| Genotype | OR | PV | CI | |
| Olyset Plus | RR vs. SS | 0.61 | ˃0.05 | 0.22–1.66 |
| RR vs. RS | 0.59 | ˃0.05 | 0.21–1.68 | |
| RS vs. SS | 1.02 | 1 | 0.56–1.8 | |
| R vs. S | 0.85 | ˃0.05 | 0.45–1.59 | |
| PermaNet 3.0 | RR vs. SS | 3.47 | ˃0.05 | 1–11.9 |
| RR vs. RS | 3.56 | ˃0.05 | 1–12.7 | |
| RS vs. SS | 0.97 | 1 | 0.53–1.76 | |
| R vs. S | 1.42 | ˃0.05 | 0.74–2.7 | |
| PermaNet 2.0 | RR vs. SS | |||
| RR vs. RS | ||||
| RS vs. SS | 1.1 | 1 | 0.6 | |
| R vs. S | 1.18 | ˃0.05 | 0.6–2.2 | |
| Genotype | OR | PV | CI | |
|---|---|---|---|---|
| Olyset Plus | RR vs. SS | 3.3 | <0.05 | 1.19–9.2 |
| RR vs. RS | 1.07 | ˃0.05 | 0.38–3.02 | |
| RS vs. SS | 3.08 | <0.01 | 1.67–5.66 | |
| R vs. S | 2.23 | <0.05 | 1.2–4.1 | |
| Mosquitoes in Room | ||||
| RR vs. SS | 12.3 | <0.001 | 2.5–60.4 | |
| RR vs. RS | 1.46 | ˃0.05 | 0.29–7.2 | |
| RS vs. SS | 8.42 | <0.001 | 4.37–16.2 | |
| R vs. S | 4.56 | <0.001 | 2.26–9.2 | |
| Olyset | RR vs. SS | 2.6 | ˃0.05 | 0.6–11.7 |
| RR vs. RS | 0.89 | ˃0.05 | 0.2–3.9 | |
| RS vs. SS | 2.97 | <0.001 | 1.65–5.35 | |
| R vs. S | 2 | <0.05 | 1.06–3.7 | |
| Blood feeding in Room | ||||
| RS vs. SS | 3 | <0.001 | 1.67–5.4 | |
| R vs. S | 1.71 | ˃0.05 | 0.89–3.3 | |
| PermaNet 3.0 | RR vs. SS | inf | ||
| RR vs. RS | inf | |||
| RS vs. SS | 0.5 | <0.05 | 0.26–0.92 | |
| R vs. S | 0.35 | <0.05 | 0.17–0.73 | |
| Blood feeding in Room | ||||
| RS vs. SS | 0.41 | ˃0.05 | 0.21–0.77 | |
| R vs. S | 0.29 | ˃0.05 | 0.14–0.63 | |
| PermaNet 2.0 | RR vs. SS | 0.4 | ˃0.05 | 0.1–1.56 |
| RR vs. RS | 0.48 | ˃0.05 | 0.12–1.9 | |
| RS vs. SS | 0.8 | ˃0.05 | 0.47–1.5 | |
| R vs. S | 1.31 | ˃0.05 | 0.6–2.5 | |
| Exophily | ||||
|---|---|---|---|---|
| Genotype | OR | PV | CI | |
| Olyset Plus | RR vs. SS | 3.1 | ˃0.05 | 1.1–9 |
| RR vs. RS | 2.48 | ˃0.05 | 0.83–7.42 | |
| RS vs. SS | 1.25 | ˃0.05 | 0.67–2.26 | |
| R vs. S | 1.1 | ˃0.05 | 0.6–2.2 | |
| Unfed Alive | ||||
| RR vs. SS | 11.76 | <0.01 | 2.59–53.4 | |
| RR vs. RS | 3.9 | 1 | 0.8–18.59 | |
| RS vs. SS | 2.99 | <0.01 | 1.58–5.6 | |
| R vs. S | 3.4 | <0.001 | 1.67–6.9 | |
| RS vs. SS | 1.18 | ˃0.05 | 0.6–2.1 | |
| R vs. S | 1.17 | ˃0.05 | 0.62–2.2 | |
| PermaNet 3.0 | RR vs. SS | 1.22 | 1 | 0.46–3.2 |
| RR vs. RS | 1.68 | ˃0.05 | 0.6–4.7 | |
| RS vs. SS | 0.59 | ˃0.05 | 0.31–1.12 | |
| R vs. S | 0.94 | 1 | 0.5–1.8 | |
| PermaNet 2.0 | RS vs. SS | 1.35 | <0.01 | 0.75–2.43 |
| R vs. S | 1.22 | ˃0.05 | 0.65–2.3 | |
| Unfed Alive | ||||
| RS vs. SS | 3.37 | <0.001 | 1.84–6.17 | |
| R vs. S | 1.8 | ˃0.05 | 0.99–3.38 | |
| Cone Assays | ||||
| Olyset Plus 40 alive vs. 40 dead | OR | PV | CI | |
| RR vs. SS | 1.1 | ˃0.05 | 0.4–2.7 | |
| RS vs. SS | 0.9 | 1.0 | 0.5–1.8 | |
| RR vs. RS | 1.1 | ˃0.05 | 0.4–2.7 | |
| R vs. S | 1.04 | 1.0 | 0.5–1.8 | |
| PermaNet 2.0 46 alive vs. 34 dead | RR vs. SS | 2.09 | <0.01 | 1.1–4.2 |
| RS vs. SS | 2.3 | <0.001 | 1.2–4.4 | |
| RR vs. RS | 0.8 | ˃0.05 | 0.3–2.4 | |
| R vs. S | 1.8 | <0.01 | 0.9–3.5 | |
| PermaNet 3.0 39 alive vs. 31 dead | RR vs. SS | 30.1 | <0.001 | 0.8–5.3 |
| RS vs. SS | 2.4 | <0.01 | 1.3–4.5 | |
| RR vs. RS | 12.3 | <0.001 | 1.5–98.6 | |
| R vs. S | 3.8 | <0.001 | 1.8–7.7 | |
| WHO Bioassays | ||||
| Deltamethrin 38 alive vs. 38 dead | OR | PV | CI | |
| RR vs. SS | 4.6 | <0.001 | 2.5–8.4 | |
| RS vs. SS | 1.22 | ˃0.05 | 0.69–2.15 | |
| RR vs. RS | 3.75 | <0.001 | 2.1–6.8 | |
| R vs. S | 2.80 | <0.01 | 1.54–5.1 | |
| Permethrin 28 alive vs. 37 dead | RR vs. SS | 4.8 | <0.001 | 2.66–8.8 |
| RS vs. SS | 1.6 | ˃0.05 | 0.86–2.8 | |
| RR vs. RS | 3.1 | <0.001 | 1.7–5.5 | |
| R vs. S | 2.99 | <0.001 | 1.7–5.3 | |
| DDT 29 alive vs. 30 dead | RR vs. SS | 66.7 | <0.001 | 27.1–164 |
| RS vs. SS | 14.0 | <0.001 | 6.3–31.2 | |
| RR vs. RS | 4.8 | <0.001 | 2.4–9.5 | |
| R v S | 15.3 | <0.001 | 7.7–30.5 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menze, B.D.; Kouamo, M.F.; Wondji, M.J.; Tchapga, W.; Tchoupo, M.; Kusimo, M.O.; Mouhamadou, C.S.; Riveron, J.M.; Wondji, C.S. An Experimental Hut Evaluation of PBO-Based and Pyrethroid-Only Nets against the Malaria Vector Anopheles funestus Reveals a Loss of Bed Nets Efficacy Associated with GSTe2 Metabolic Resistance. Genes 2020, 11, 143. https://doi.org/10.3390/genes11020143
Menze BD, Kouamo MF, Wondji MJ, Tchapga W, Tchoupo M, Kusimo MO, Mouhamadou CS, Riveron JM, Wondji CS. An Experimental Hut Evaluation of PBO-Based and Pyrethroid-Only Nets against the Malaria Vector Anopheles funestus Reveals a Loss of Bed Nets Efficacy Associated with GSTe2 Metabolic Resistance. Genes. 2020; 11(2):143. https://doi.org/10.3390/genes11020143
Chicago/Turabian StyleMenze, Benjamin D., Mersimine F. Kouamo, Murielle J. Wondji, Williams Tchapga, Micareme Tchoupo, Michael O. Kusimo, Chouaibou S. Mouhamadou, Jacob M. Riveron, and Charles S. Wondji. 2020. "An Experimental Hut Evaluation of PBO-Based and Pyrethroid-Only Nets against the Malaria Vector Anopheles funestus Reveals a Loss of Bed Nets Efficacy Associated with GSTe2 Metabolic Resistance" Genes 11, no. 2: 143. https://doi.org/10.3390/genes11020143