The Origin of a Coastal Indigenous Horse Breed in China Revealed by Genome-Wide SNP Data

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
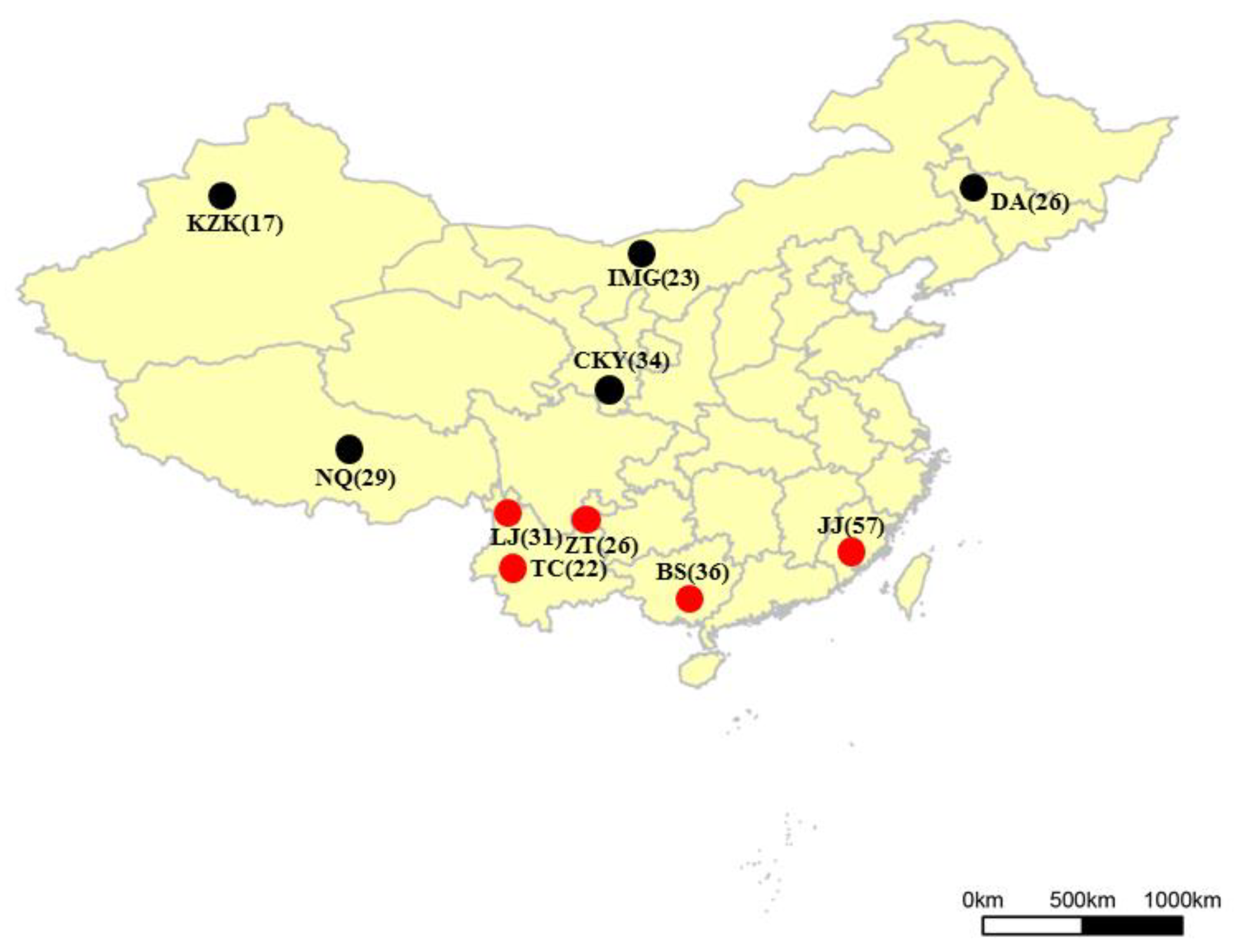
2.1. Sample Collection and DNA Extraction
2.2. Datasets
2.3. Population Divergence
2.4. LD Decay
2.5. Identical by Descent Analyses
2.6. Runs of Homozygosity and Genetic Diversity Analysis
2.7. Formal Test of Ancestor Admixture
2.8. Estimating the Ancestry Proportion of the Jinjiang Horse
3. Results
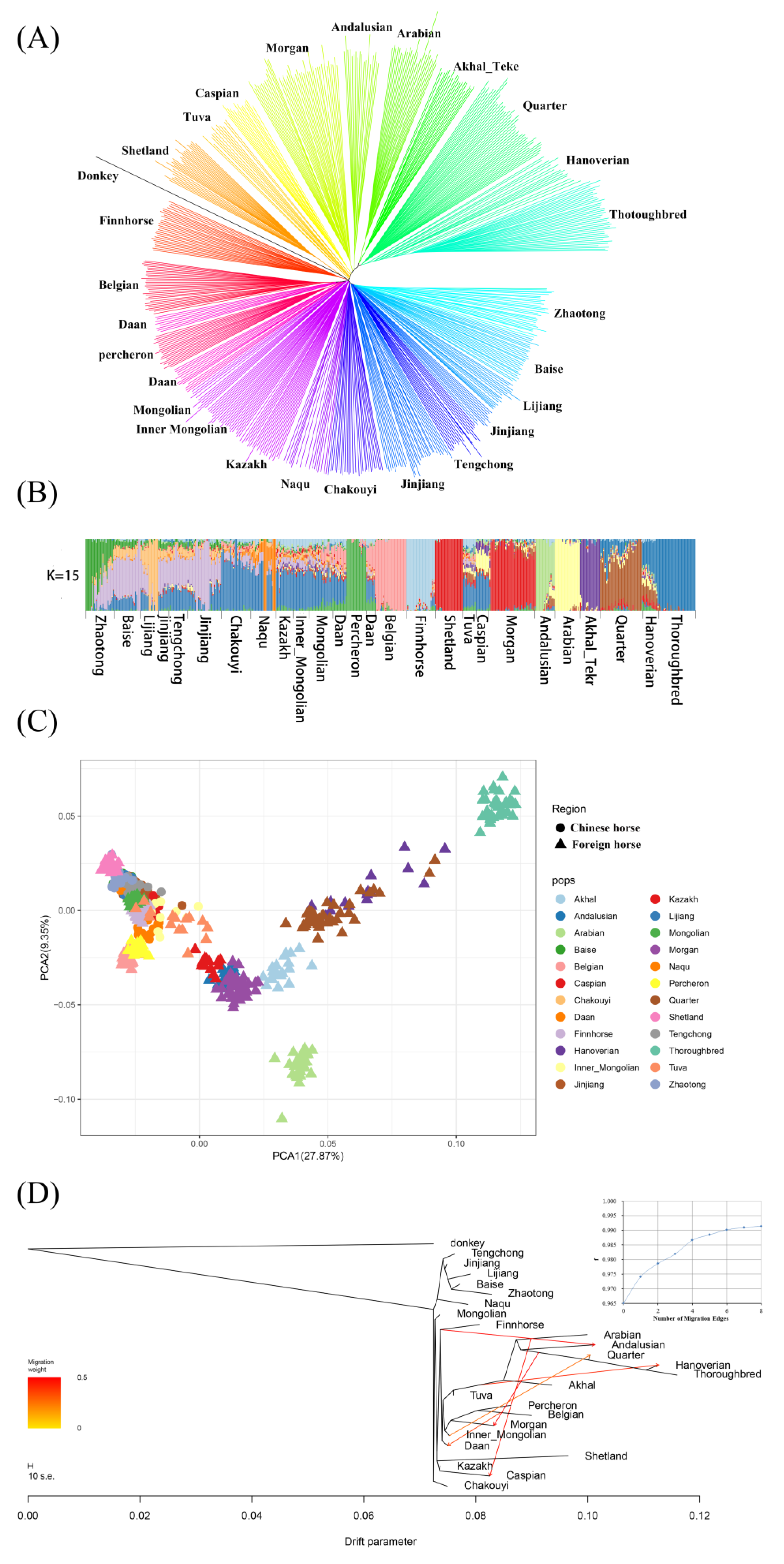
3.1. Genetic Relationships between the Jinjiang Horses and Worldwide Horse Breeds
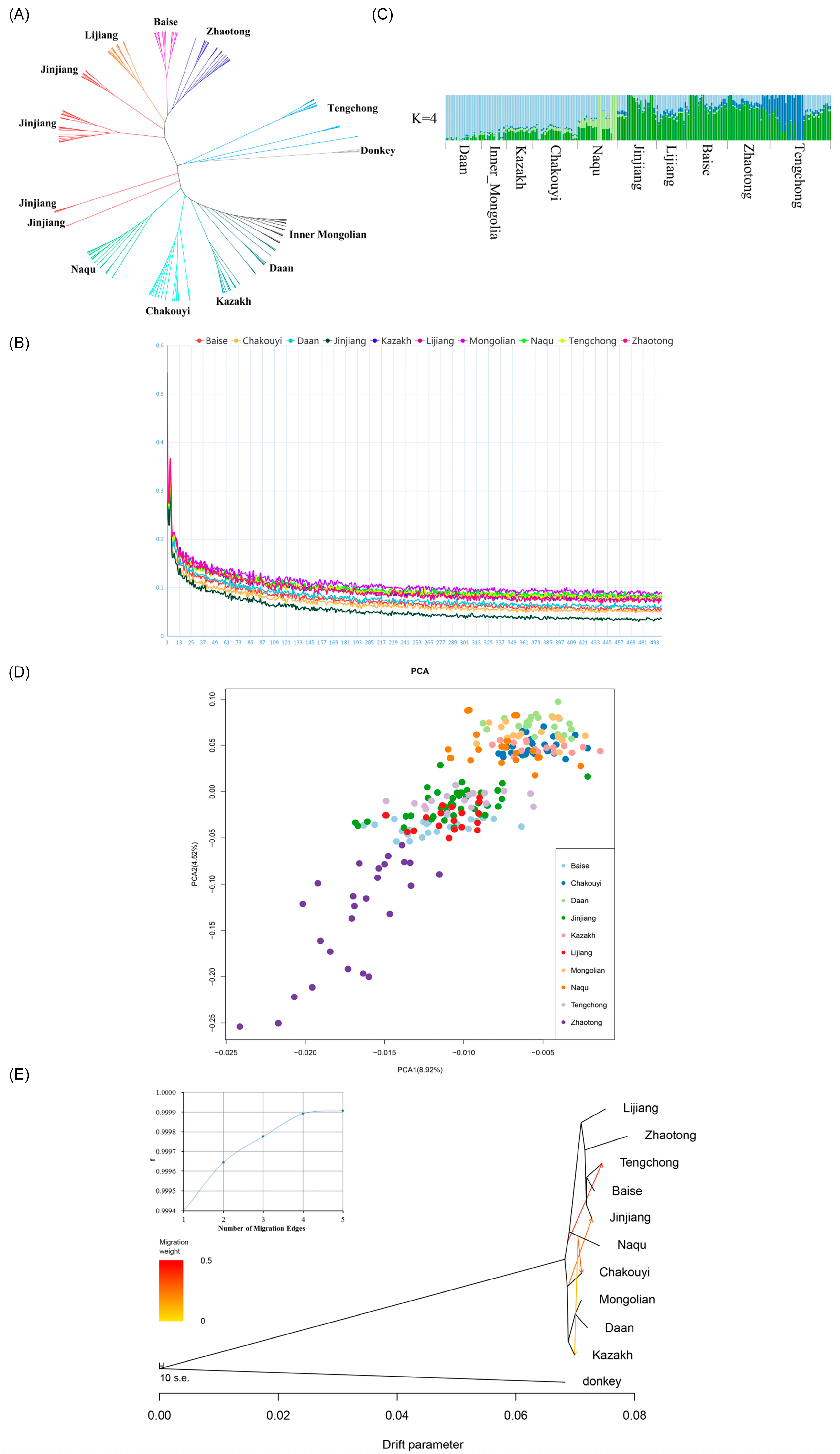
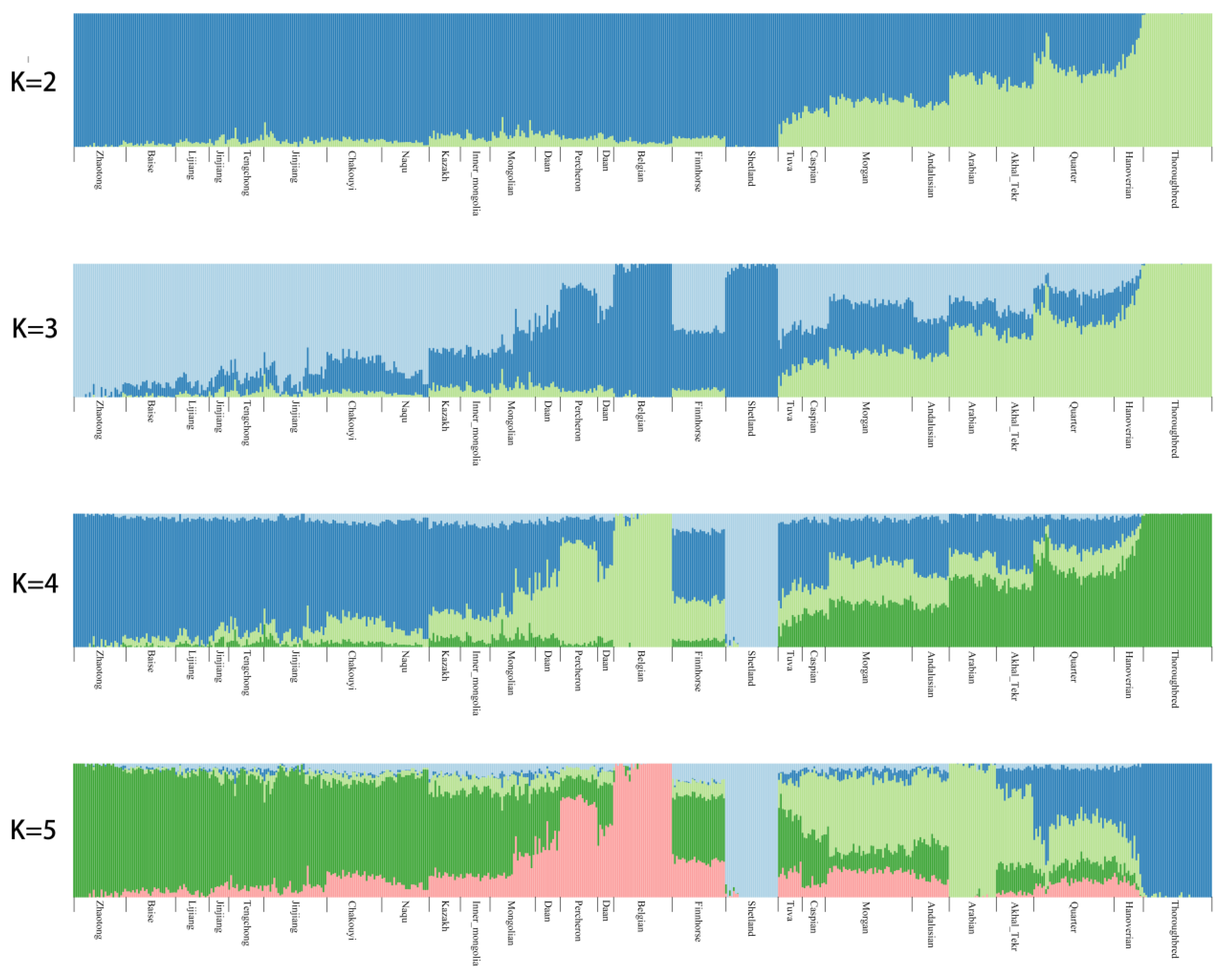
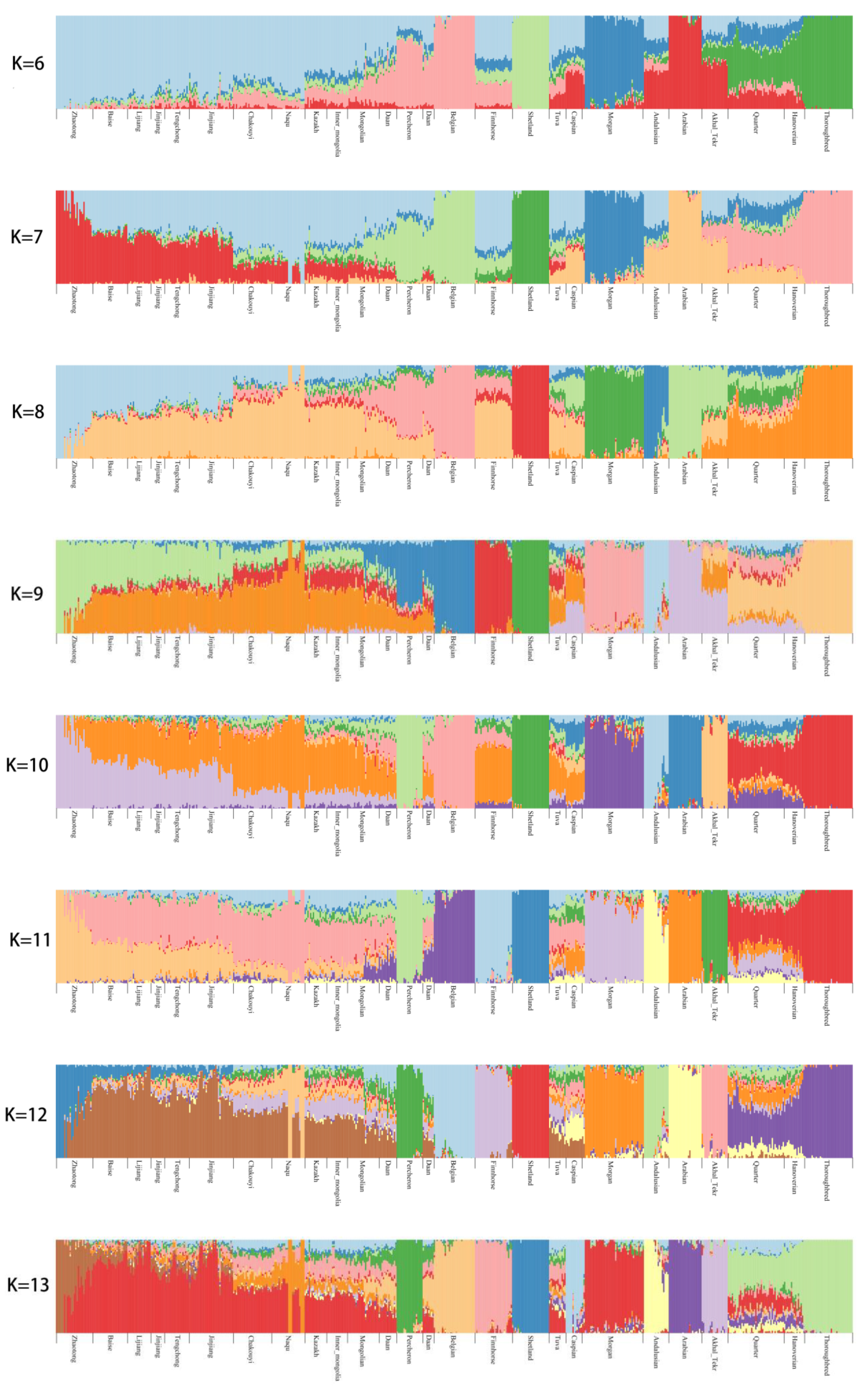
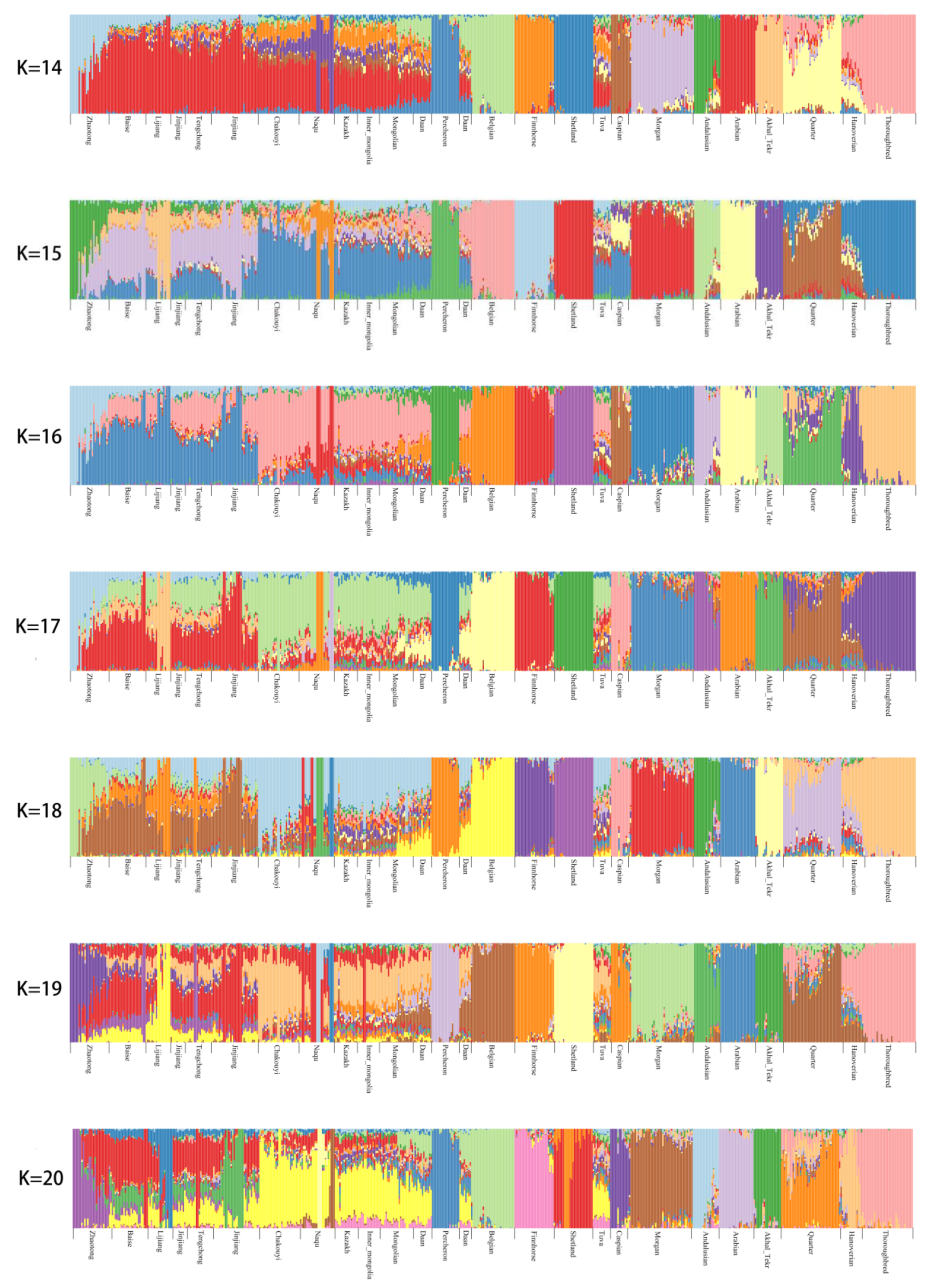
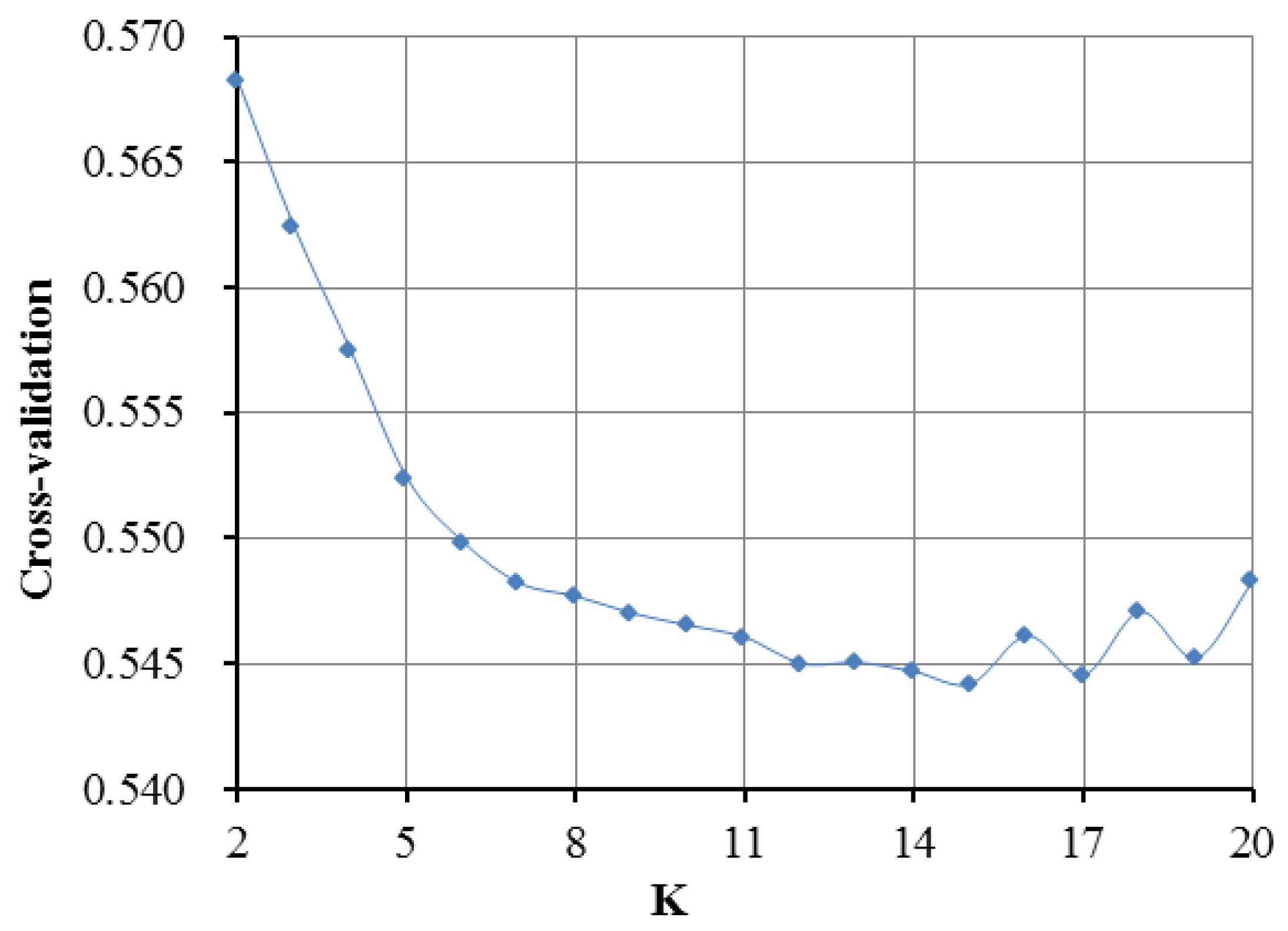
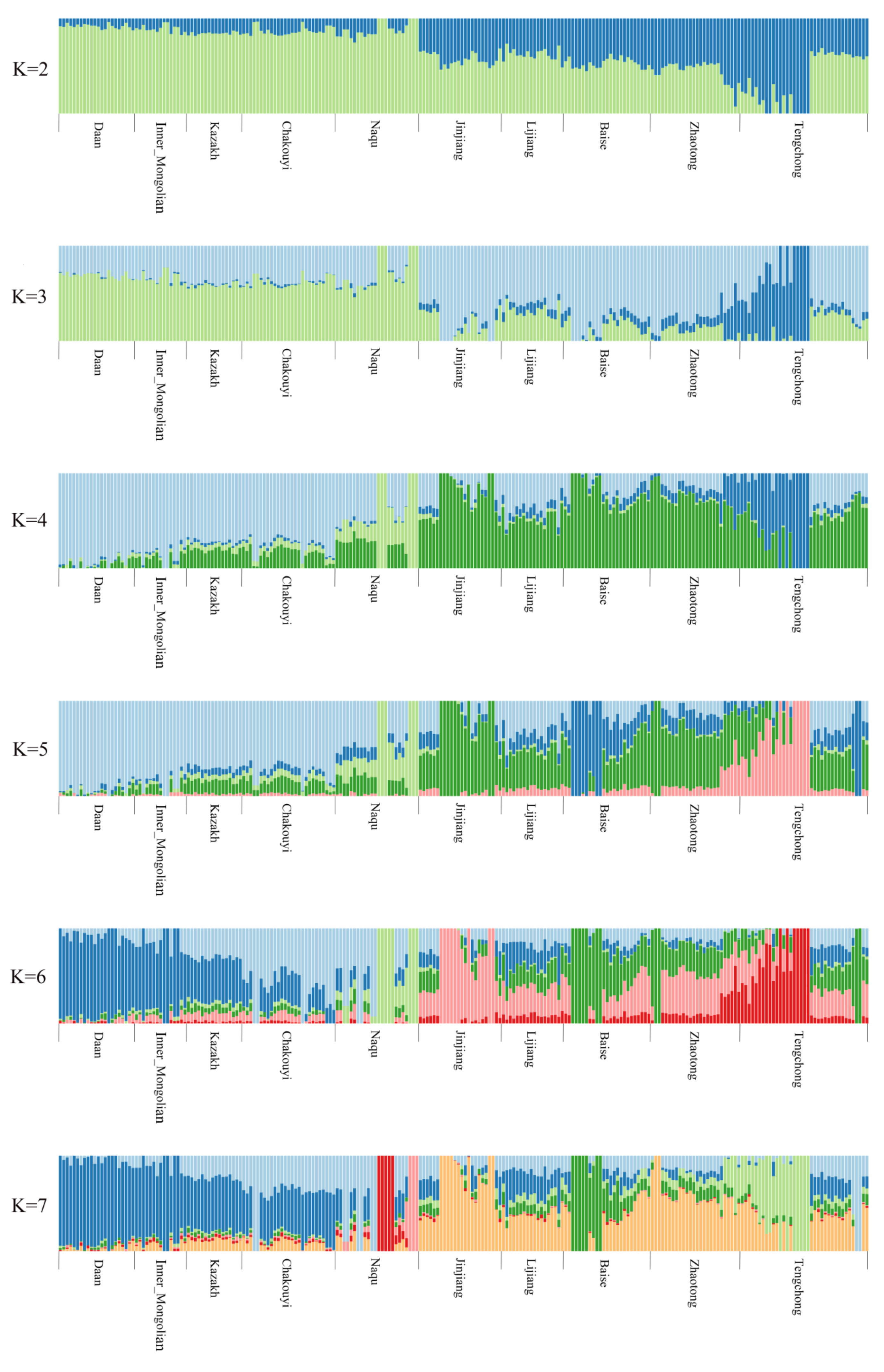
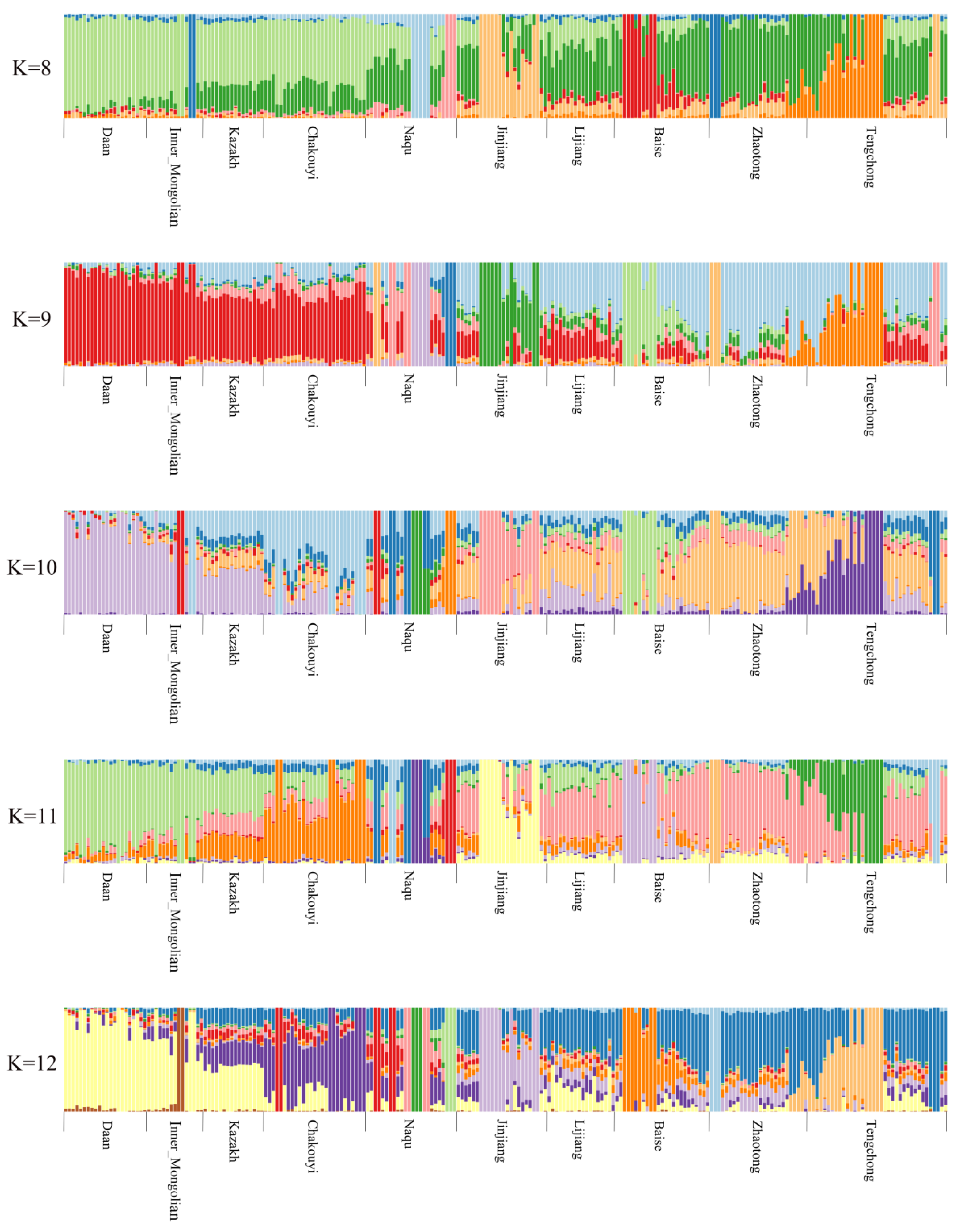
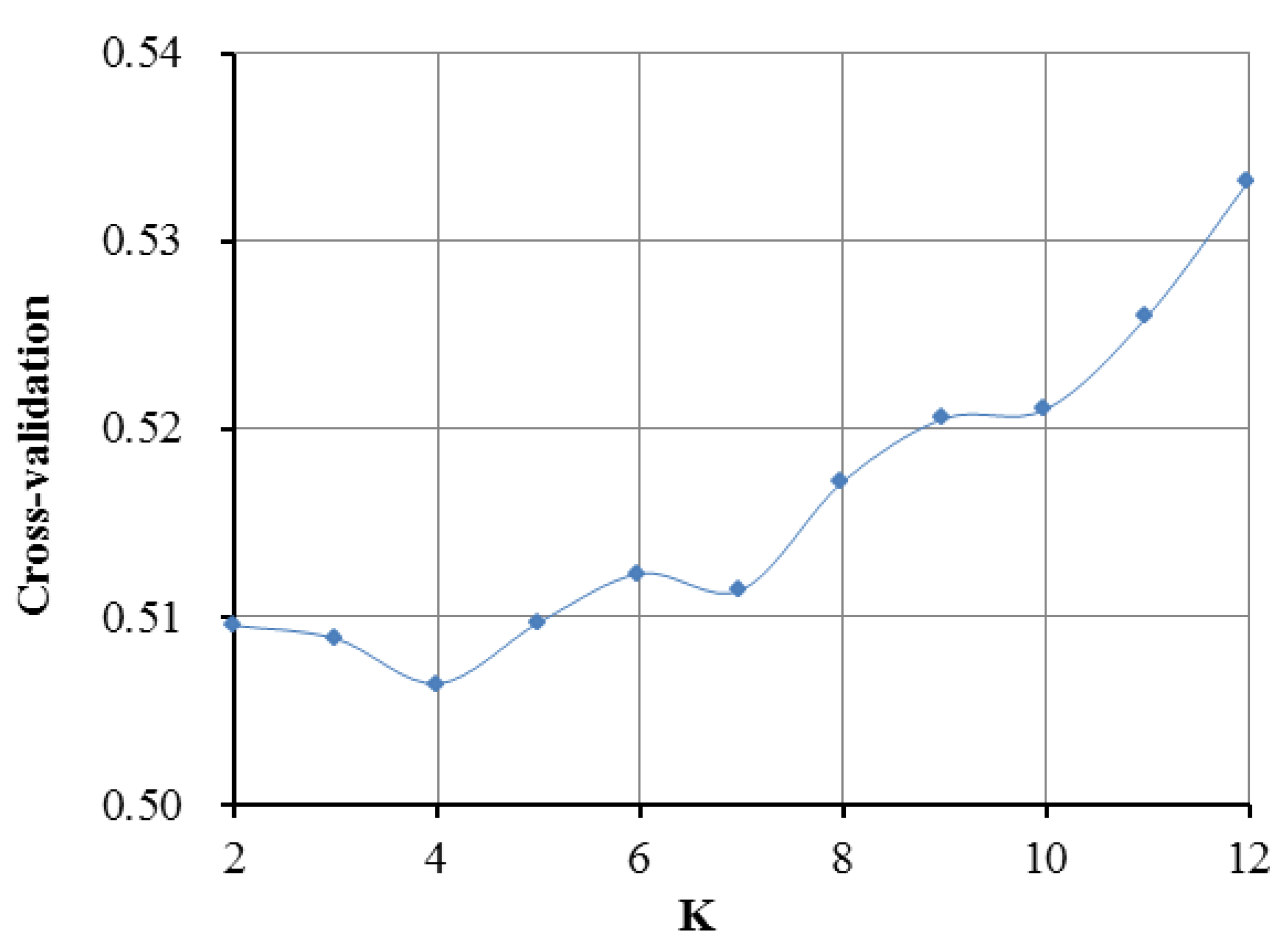
3.2. Population Genetic Relationships between the Jinjiang Horse and Other Chinese Indigenous Horse Breeds
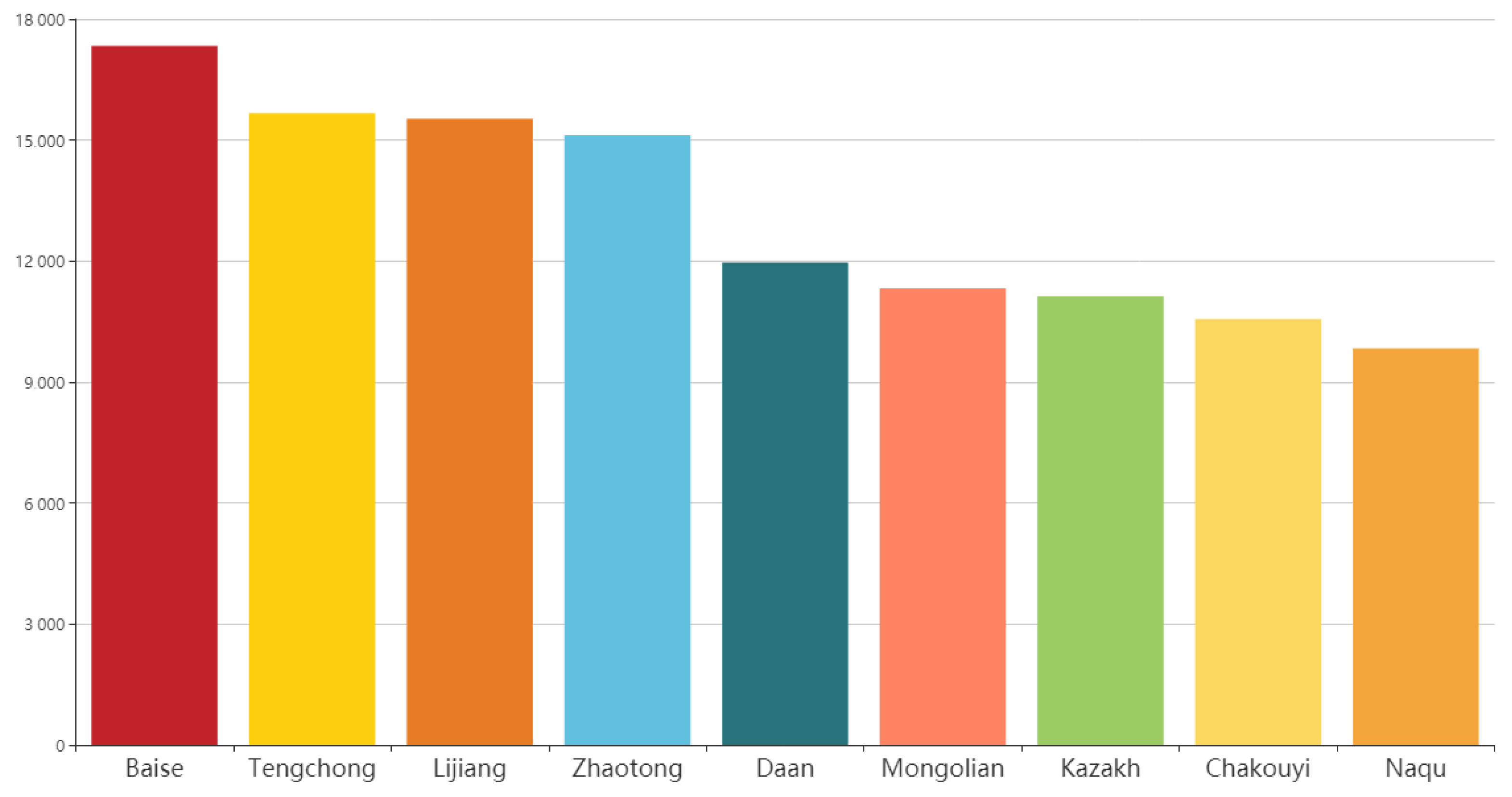
3.3. Runs of Homozygosity and the Genetic Diversity of the Worldwide Horse Breeds
3.4. Identical by Descent Analyses of the Jinjiang Horse
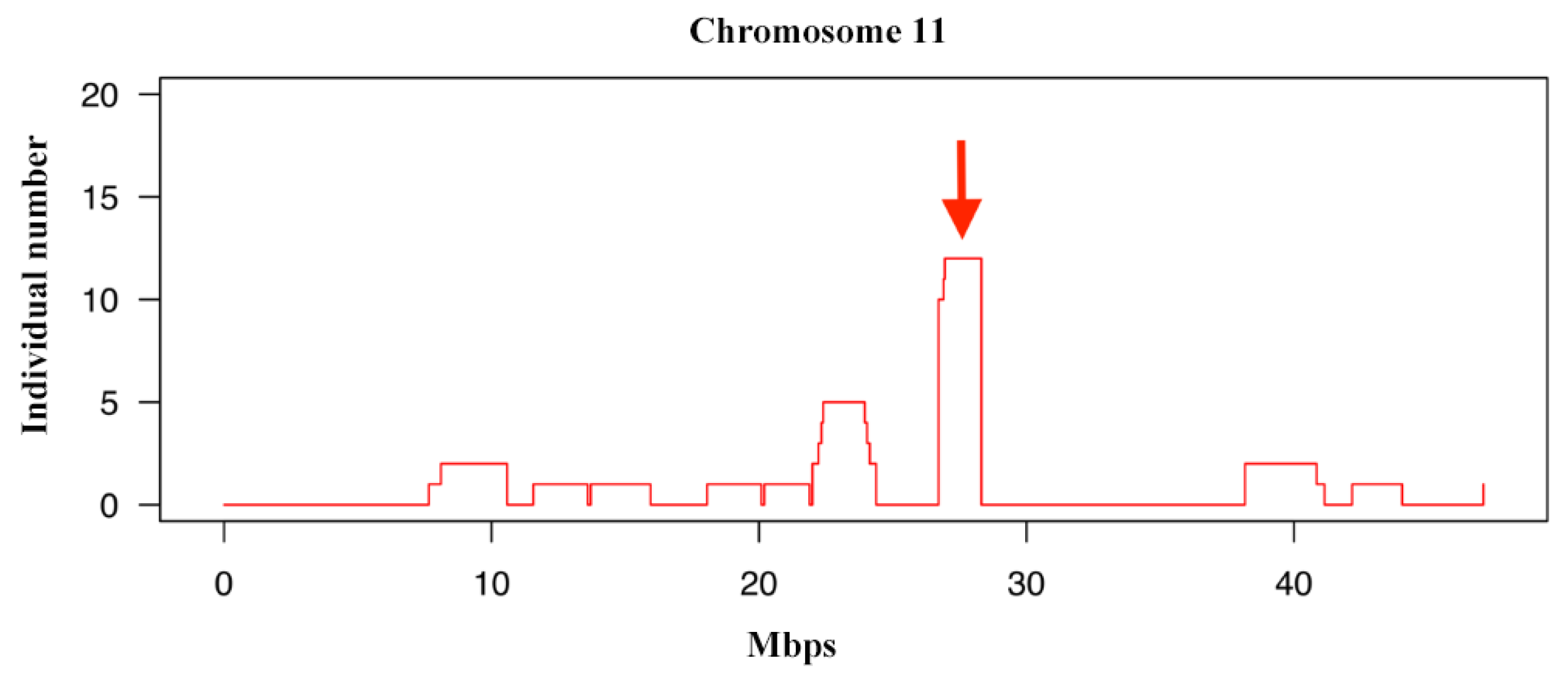
3.5. Identifying Signatures of Selection in the Jinjiang Horse Population with Runs of Homozygosity Analysis
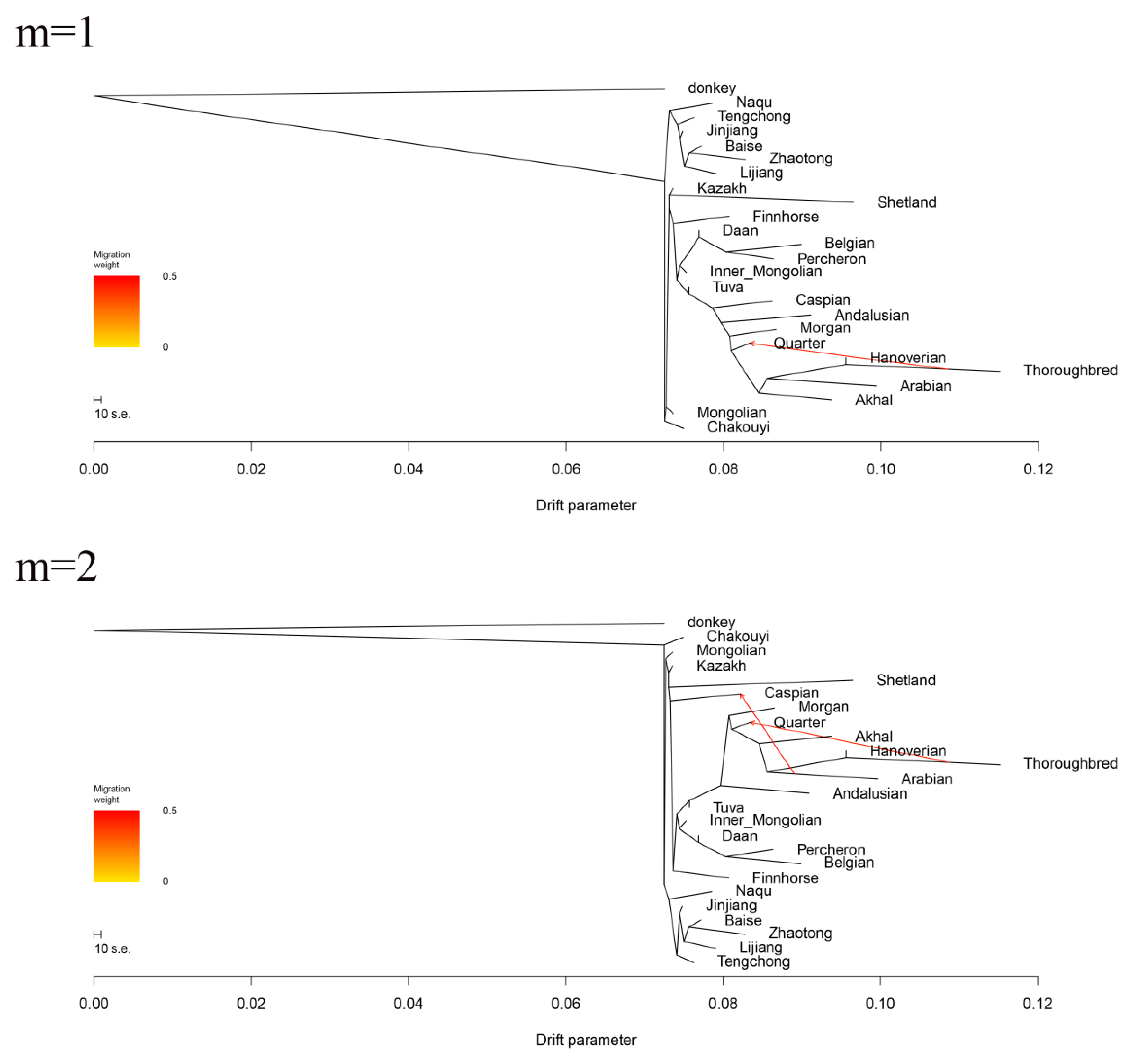
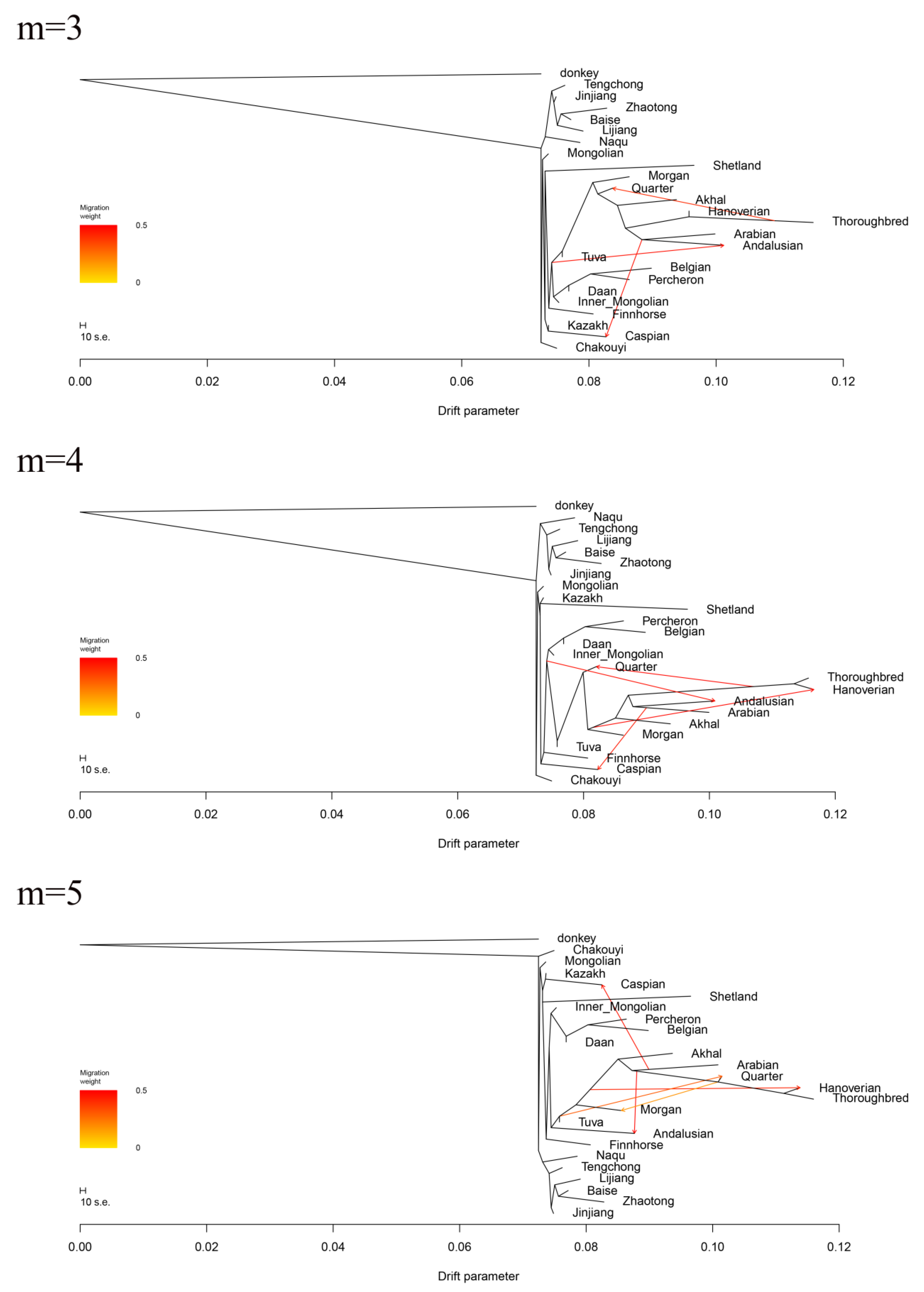
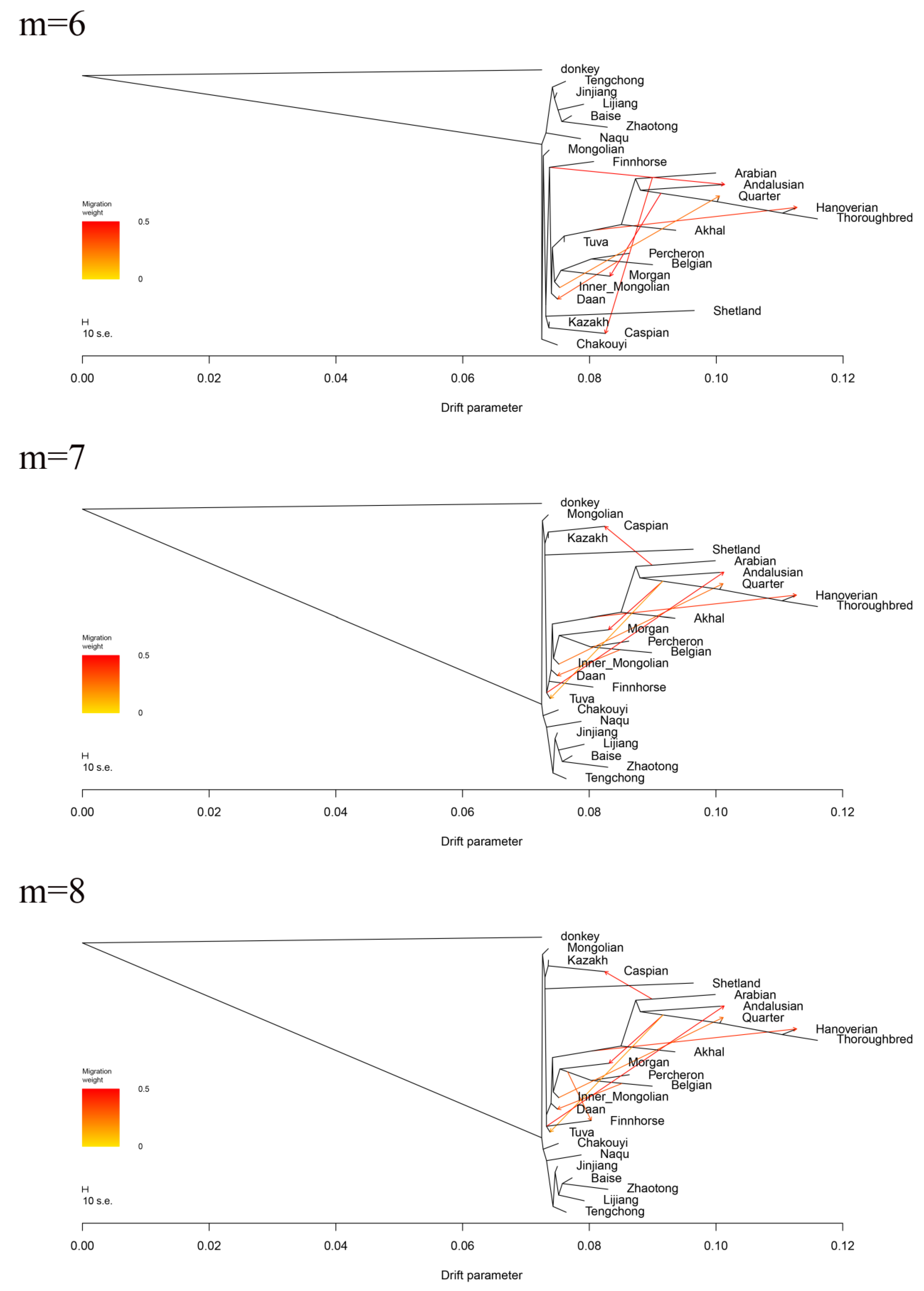
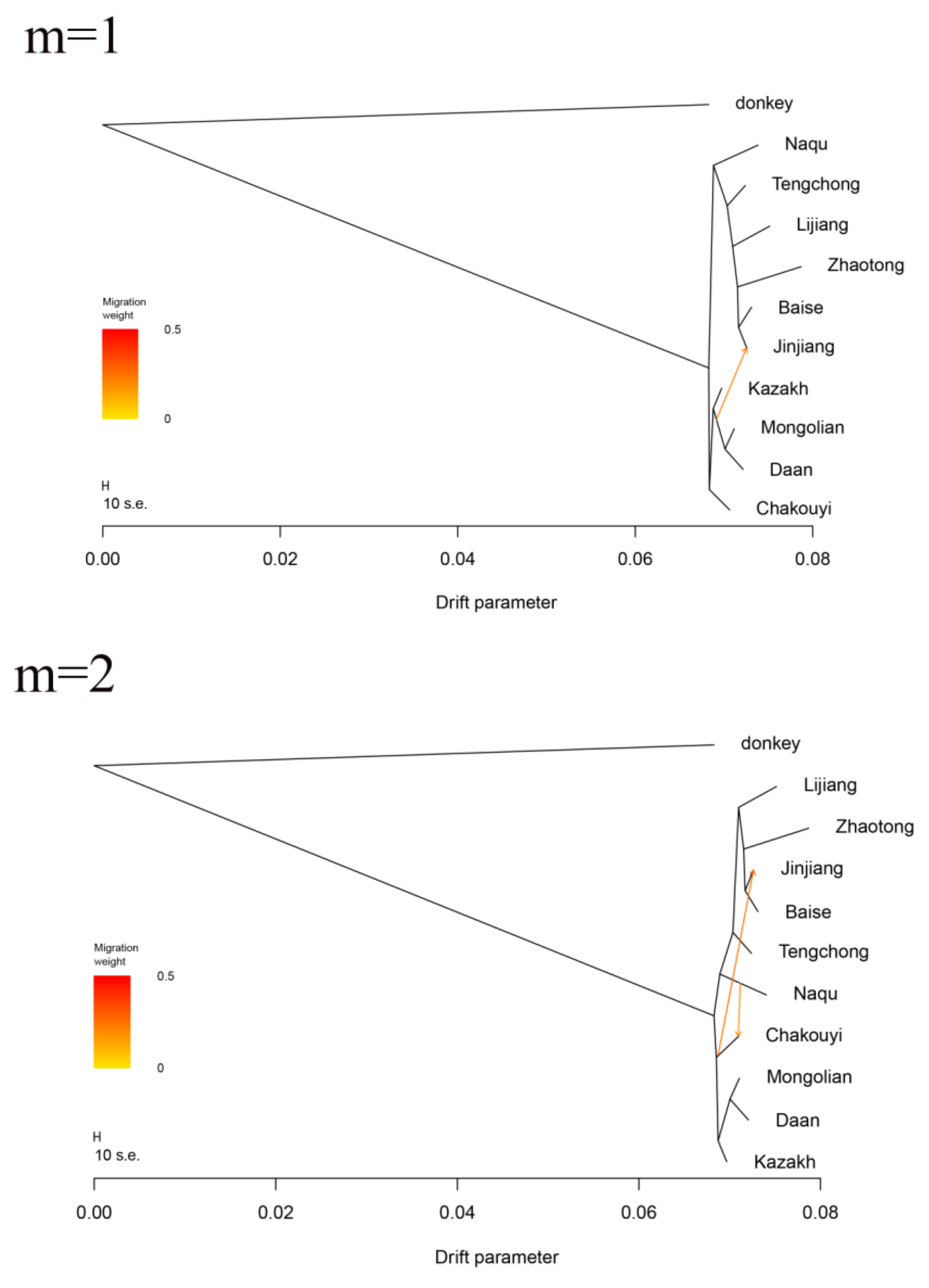
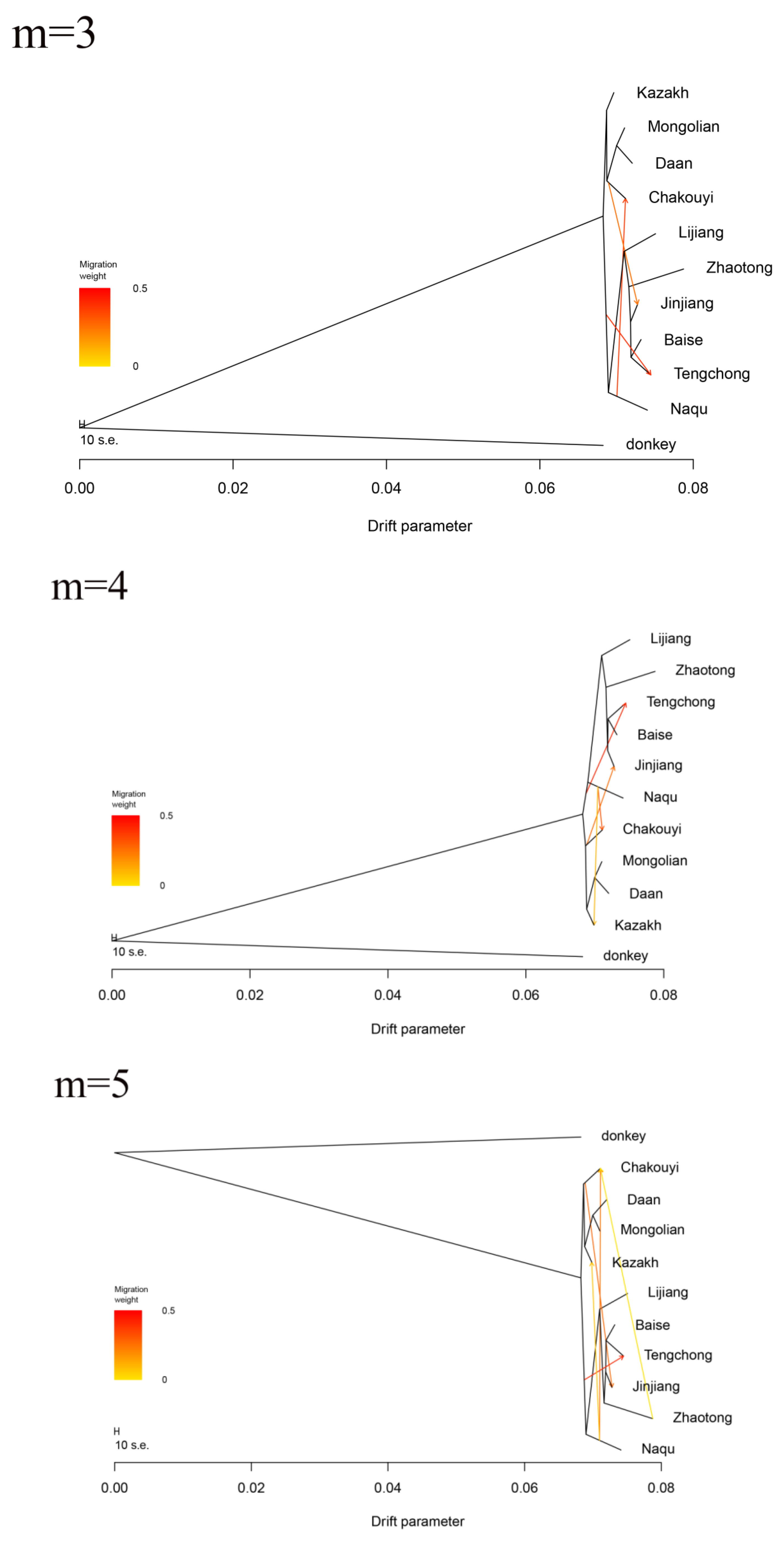
3.6. Estimation of Possible Ancestry with a Formal Test of ADMIXTURE
3.7. Estimating the Proportion of Admixture
4. Discussion
4.1. The Relationship between Jinjiang Horses and Worldwide Horse Populations
4.2. Native Origin of Jinjiang Horses
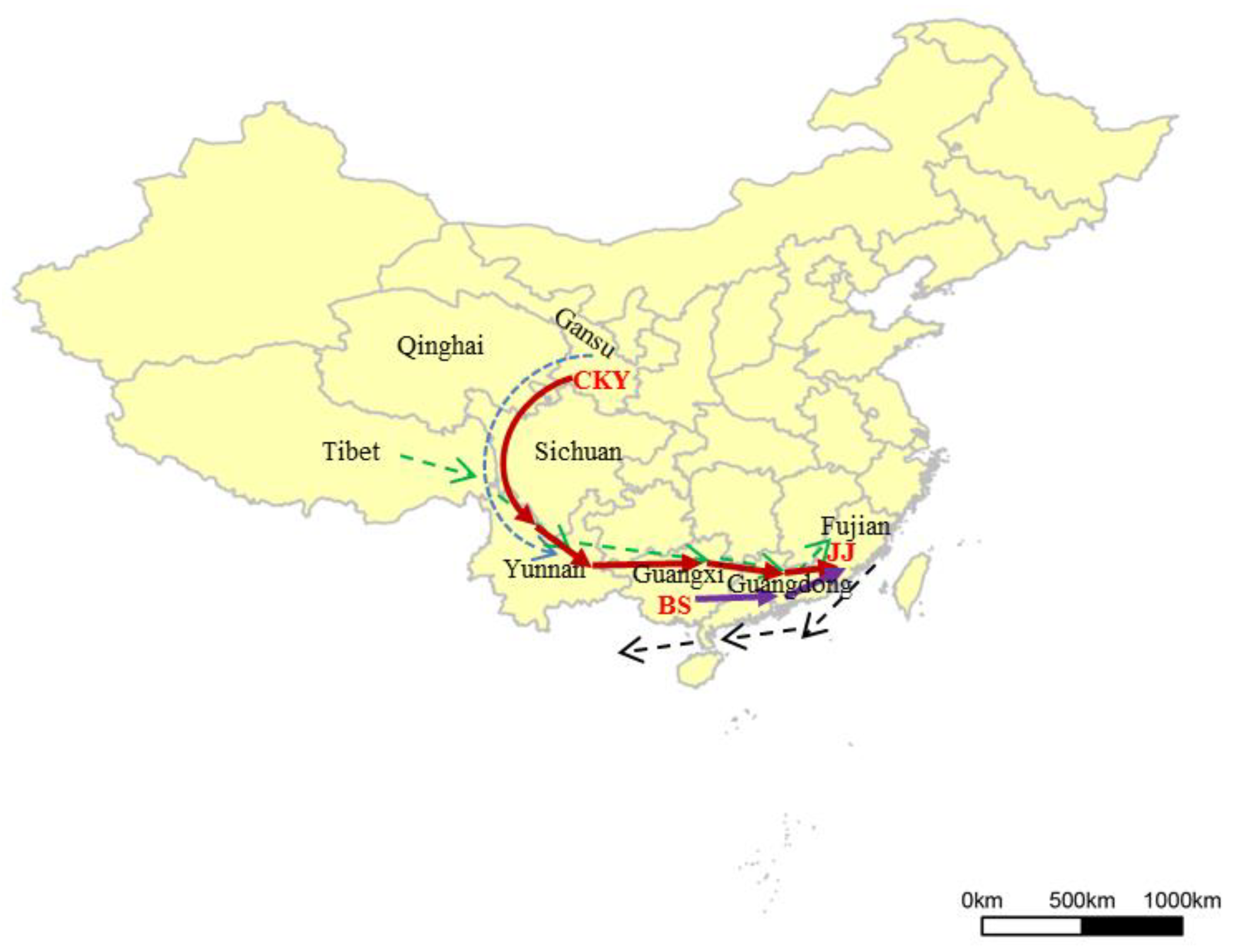
4.3. The Migrating Routes of the Ancestral Horses
4.4. The Conservation of Jinjiang Horses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




Appendix B



Appendix C



Appendix D


References
- Xie, C.X. Variety of Horse Donkey; Shanghai Science and Technology Press: Shanghai, China, 1987. [Google Scholar]
- Huang, J.K. Formation and naming of Jinjiang Horse. China Livest. Poult. Ind. 2010, 6, 43. [Google Scholar]
- Chang, H. Animal Genetic Resources in China, Horses Donkeys Camels; China Agriculture Press: Beijing, China, 2011; pp. 48–52. [Google Scholar]
- Wu, Y.X. Maritime Silk Road and Jinjiang’s Multicultural Culture; The Straits Publishing & Distributing Group: Fuzhou, China, 2016; Volume 180. [Google Scholar]
- Zhuang, X.L. The variety resources and protection measures of Jinjiang horse. Fujian Anim. Sci. Vet. Med. 2006, 28, 69–70. [Google Scholar]
- Vilà, C.; Leonard, J.A.; Götherström, A.; Marklund, S.; Sandberg, K.; Liden, K.; Wayne, R.K.; Ellegren, H. Widespread origins of domestic horse lineages. Science 2001, 291, 474–477. [Google Scholar]
- Jansen, T.; Forster, P.; Levine, M.A.; Oelke, H.; Hurles, M.; Renfrew, C.; Weber, J.; Olek, K. Mitochondrial DNA and the origins of the domestic horse. Proc. Natl. Acad. Sci. USA 2002, 99, 10905–10910. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, M.; Pruvost, M.; Benecke, N.; Hofreiter, M.; Morales, A.; Reissmann, M.; Ludwig, A. Origin and History of Mitochondrial DNA Lineages in Domestic Horses. PLoS ONE 2010, 5, e15311. [Google Scholar] [CrossRef] [PubMed]
- Lippold, S.; Matzke, N.J.; Reissmann, M.; Hofreiter, M. Whole mitochondrial genome sequencing of domestic horses reveals incorporation of extensive wild horse diversity during domestication. BMC Evol. Biol. 2011, 11, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Achilli, A.; Olivieri, A.; Soares, P.; Lancioni, H.; Kashani, B.H.; Perego, U.A.; Nergadze, S.G.; Carossa, V.; Santagostino, M.; Capomaccio, S.; et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 2449–2454. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhu, Q.; Liu, S.; Zhao, C.; Wu, C. The origin of Chinese domestic horses revealed with novel mtDNA variants. Anim. Sci. J. 2017, 88, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wu, Y.; Xiang, H.; Yang, Y.; Wang, M.; Zhao, C.; Wu, C. Some maternal lineages of domestic horses may have origins in East Asia revealed with further evidence of mitochondrial genomes and HVR-1 sequences. Peer J. 2008, 6, 4896. [Google Scholar] [CrossRef] [PubMed]
- Tozaki, T.; Takezaki, N.; Hasegawa, T.; Ishida, N.; Kurosawa, M.; Tomita, M.; Saitou, N.; Mukiyama, H. Microsatellite variation in Japanese and Asian horses and their phylogenetic relationship using a European horse outgroup. J. Hered. 2003, 94, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Curik, I.; Zechner, P.; Sölkner, J.; Achmann, R.; Bodo, I.; Dovc, P.; Kavar, T.; Marti, E.; Brem, G. Inbreeding, microsatellite heterozygosity, and morphological traits in Lipizzan horses. J. Hered. 2003, 94, 125–132. [Google Scholar] [CrossRef]
- Solis, A.; Jugo, B.M.; Mériaux, J.C.; Iriondo, M.; Mazón, L.I.; Aguirre, A.I.; Vicario, A.; Estomba, A. Genetic diversity within and among four South European native horse breeds based on microsatellite DNA analysis: Implications for conservation. J. Hered. 2005, 96, 670–678. [Google Scholar] [CrossRef]
- Ling, Y.H.; Ma, Y.H.; Guan, W.J.; Chen, Y.J.; Wang, Y.P.; Han, J.L.; Mang, L.; Zhao, Q.J.; He, X.H.; Pu, Y.B.; et al. Evaluation of the genetic diversity and population structure of Chinese indigenous horse breeds using 27 microsatellite markers. Anim. Genet. 2010, 42, 56–65. [Google Scholar] [CrossRef]
- Lindgren, G.; Backström, N.; Swinburne, J.; Hellborg, L.; Einarsson, A.; Sandberg, K.; Cothran, G.; Vilà, C.; Binns, M.; Ellegren, H. Limited number of patrilines in horse domestication. Nat. Genet. 2004, 36, 335–336. [Google Scholar] [CrossRef]
- Lippold, S.; Knapp, M.; Kuznetsova, T.; Leonard, J.A.; Benecke, N.; Ludwig, A.; Rasmussen, M.; Cooper, A.; Weinstock, J.; Willerslev, E.; et al. Discovery of lost diversity of paternal horse lineages using ancient DNA. Nat. Commun. 2011, 2, 450–456. [Google Scholar] [CrossRef]
- Wallner, B.; Palmieri, N.; Vogl, C.; Rigler, D.; Bozlak, E.; Druml, T.; Jagannathan, V.; Leeb, T.; Fries, R.; Tetens, J.; et al. Y Chromosome Uncovers the Recent Oriental Origin of Modern Stallions. Curr. Biol. 2017, 27, 2029–2035. [Google Scholar] [CrossRef]
- Orozco-Terwengel, P.; Barbato, M.; Nicolazzi, E.; Biscarini, F.; Milanesi, M.; Davies, W.; Williams, D.; Stella, A.; Ajmone-Marsan, P.; Bruford, M.W. Revisiting demographic processes in cattle with genome-wide population genetic analysis. Front. Genet. 2015, 6, 191–205. [Google Scholar] [CrossRef]
- Melegh, B.I.; Banfai, Z.; Hadzsiev, K.; Miseta, A.; Melegh, B. Refining the South Asian Origin of the Romani people. BMC Genet. 2017, 18, 82–94. [Google Scholar] [CrossRef]
- Rahmatalla, S.A.; Arends, D.; Reissmann, M.; Ahmed, A.S.; Wimmers, K.; Reyer, H.; Brockmann, G.A. Whole genome population genetics analysis of Sudanese goats identifies regions harboring genes associated with major traits. BMC Genet. 2017, 18, 92–101. [Google Scholar] [CrossRef]
- Colli, L.; Milanesi, M.; Vajana, E.; Iamartino, D.; Bomba, L.; Puglisi, F.; Del Corvo, M.; Nicolazzi, E.L.; Ahmed, S.S.E.; Herrera, J.R.V.; et al. New Insights on Water Buffalo Genomic Diversity and Post-Domestication Migration Routes From Medium Density SNP Chip Data. Front. Genet. 2018, 9, 53–69. [Google Scholar] [CrossRef]
- Mccue, M.E.; Bannasch, D.L.; Petersen, J.L.; Gurr, J.; Bailey, E.; Binns, M.M.; Distl, O.; Guérin, G.; Hasegawa, T.; Hill, E.W.; et al. A High Density SNP Array for the Domestic Horse and Extant Perissodactyla: Utility for Association Mapping, Genetic Diversity, and Phylogeny Studies. PLoS Genet. 2012, 8, e1002451. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.L.; Mickelson, J.R.; Cothran, E.G.; Andersson, L.S.; Axelsson, J.; Bailey, E.; Bannasch, D.; Binns, M.M.; Borges, A.S.; Brama, P.; et al. Genetic Diversity in the Modern Horse Illustrated from Genome-Wide SNP Data. PLoS ONE 2013, 8, e54997. [Google Scholar] [CrossRef] [PubMed]
- Kader, A.; Li, Y.; Dong, K.; Irwin, D.M.; Zhao, Q.; He, X.; Liu, J.; Pu, Y.; Gorkhali, N.A.; Liu, X.; et al. Population Variation Reveals Independent Selection toward Small Body Size in Chinese Debao Pony. Genome Biol. Evol. 2015, 8, 42–50. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinf. 2008, 9, 299–306. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef]
- Falush, D.; Wirth, T.; Linz, B.; Pritchard, J.K.; Stephens, M.; Kidd, M.; Blaser, M.J.; Graham, D.Y.; Vacher, S.; Perez-Perez, G.I.; et al. Traces of human migrations in Helicobacter pylori populations. Science 2003, 299, 1582–1585. [Google Scholar] [CrossRef]
- Pickrell, J.K.; Pritchard, J.K. Inference of population splits and mixtures from genome-wide allele frequency data. PLoS Genet. 2012, 8, e1002967. [Google Scholar] [CrossRef]
- Han, L.; Abney, M. Identity by descent estimation with dense genome-wide genotype data. Genet. Epidemiol. 2011, 35, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Patterson, N.; Moorjani, P.; Luo, Y.; Mallick, S.; Rohland, N.; Zhan, Y.; Genschoreck, T.; Webster, T.; Reich, D. Ancient admixture in human history. Genetics 2012, 192, 1065–1093. [Google Scholar] [CrossRef]
- Loh, P.R.; Lipson, M.; Patterson, N.; Moorjani, P.; Pickrell, J.K.; Reich, D.; Berger, B. Inferring admixture histories of human populations using linkage disequilibrium. Genetics 2013, 193, 1233–1254. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.M.P.; Utsunomiya, Y.T.; Meszaros, G.; Bickhart, D.M.; Liu, G.E.; Van Tassell, C.P.; Sonstegard, T.S.; Da Silva, M.V.; Garcia, J.F.; Sölkner, J. Assessing signatures of selection through variation in linkage disequilibrium between taurine and indicine cattle. Genet. Sel. Evol. 2014, 46, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dong, Q.; Miao, Y.; Fu, L.; Lin, X.; Wang, E. Clinical significance and biological roles of SPAG9 overexpression in non-small cell lung cancer. Lung Cancer 2013, 81, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Shen, B.; Zhang, J.; Zhang, Q. Sperm-associated antigen 9 overexpression correlates with poor prognosis and insensitive to Taxol treatment in breast cancer. Biomarkers 2015, 21, 1–6. [Google Scholar] [CrossRef]
- Ren, B.; Luo, S.; Xu, F.; Zou, G.; Xu, G.; He, J.; Huang, Y.; Zhu, H.; Li, Y. The expression of DAMP proteins HSP70 and cancer-testis antigen SPAG9 in peripheral blood of patients with HCC and lung cancer. Cell Stress Chaperones 2016, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cropp, C.S.; Lidereau, R.; Leone, A.; Liscia, D.; Cappa, A.P.; Campbell, G.; Barker, E.; Le Doussal, V.; Steeg, P.S.; Callahan, R. NME1 Protein Expression and Loss of Heterozygosity Mutations in Primary Human Breast Tumors. JNCI J. Natl. Cancer Inst. 1994, 86, 1167–1169. [Google Scholar] [CrossRef]
- Qu, S.; Long, J.; Cai, Q.; Shu, X.O.; Cai, H.; Gao, Y.T.; Zheng, W. Genetic Polymorphisms of Metastasis Suppressor Gene NME1 and Breast Cancer Survival. Clin. Cancer Res. An Off. J. Am. Assoc. Cancer Res. 2008, 14, 4787–4793. [Google Scholar] [CrossRef]
- Postel, E.H.; Zou, X.; Notterman, D.A.; La Perle, K.M. Double knockout Nme1/Nme2 mouse model suggests a critical role for NDP kinases in erythroid development. Mol. Cell. Biochem. 2009, 329, 45–50. [Google Scholar] [CrossRef]
- Yao, Y.; Li, C.; Zhou, X.; Zhang, Y.; Lu, Y.; Chen, J.; Zheng, X.; Tao, D.; Liu, Y.; Ma, Y. PIWIL2 induces c-Myc expression by interacting with NME2 and regulates c-Myc-mediated tumor cell proliferation. Oncotarget 2014, 5, 8466–8477. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.B.; Ma, J.Y.; Zhang, Q.H.; Lin, F.; Wang, Z.W.; Huang, L.; Schatten, H.; Sun, Q.Y. MBTD1 is associated with Pr-Set7 to stabilize H4K20me1 in mouse oocyte meiotic maturation. Cell Cycle 2013, 12, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Park, E.H.; Kim, M.D. Genome-Wide Screening of Saccharomyces cerevisiae Genes Regulated by Vanillin. J. Microbiol. Biotechnol. 2014, 25, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T.; Scozzafava, A. Carbonic anhydrase inhibitors and their therapeutic potential. Expert Opin. Ther. Pat. 2000, 10, 575–600. [Google Scholar] [CrossRef]
- Huang, G.M. General History of Fujian Economic Geography; Fujian Science & Technology Publishing House: Fuzhou, China, 2005; pp. 202–203. [Google Scholar]
- Xiong, Z.Y. Study on the Origin of Jinjiang Horse with DNA Genetic Polymorphisms. Master’s Thesis, China Agricultural University, Beijing, China, 2017; p. 49. [Google Scholar]
- Cui, Y.X. Horse Science; Agricultural Press: Beijing, China, 1990; pp. 108–109. [Google Scholar]
- Seyedabadi, H.; Amirinia, C.; Banabazi, M.H.; Emrani, H. Parentage verification of Iranian Caspian horse using microsatellites markers. Iran. J. Biotechnol. 2006, 4, 260–264. [Google Scholar]
- Qin, Y.Z. On the scope and geographical features of “Northwest Ethnic corridor”. J. Minzu Univ. China (Philos. Soc. Sci.) 2011, 3, 67–72. [Google Scholar]
- Liang, Y.B.; Xu, D.X.; Su, J.L. Investigation of Baise Horse Breeds. China Herbiv. Sci. 2010, 30, 76–78. [Google Scholar]
- Zhu, W.H. Try to talk about the Song Dynasty Guangnan west road to the development of outside trade. Jiangxi Acad. Soc. Sci. 2010, 5, 160–164. [Google Scholar]
- Shi, S. Ethnic flows from Upper reaches of the Yellow River to Tibetan-Yi corridor at the Neolithic Culture. J. Southwest Univ. Natl. (Nat. Sci. Ed.) 2008, 29, 1–7. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | N | NROH | SROH | LROH | FROH | HO | HE | f |
|---|---|---|---|---|---|---|---|---|
| Baise | 9 | 2.3 | 32,000.9 | 9850.0 | 0.0143 | 0.2818 | 0.3031 | 0.1024 |
| Chakouyi | 18 | 1.8 | 13,500.2 | 7574.4 | 0.0060 | 0.2824 | 0.3032 | 0.0397 |
| Daan | 17 | 2.9 | 29,691.3 | 9136.1 | 0.0132 | 0.2872 | 0.3064 | 0.0702 |
| Jinjiang | 19 | 5.8 | 74,697.5 | 9550.1 | 0.0333 | 0.2942 | 0.3102 | 0.0970 |
| Kazakh | 7 | 3.1 | 63,988.7 | 14,983.1 | 0.0285 | 0.2828 | 0.3041 | 0.0768 |
| Lijiang | 8 | 6.0 | 104,407.2 | 15,405.5 | 0.0465 | 0.2853 | 0.3067 | 0.1024 |
| Inner Mongolian | 10 | 4.4 | 69,589.7 | 9846.7 | 0.0310 | 0.2891 | 0.3078 | 0.0500 |
| Naqu | 14 | 1.9 | 14,013.9 | 6859.1 | 0.0062 | 0.2935 | 0.3092 | 0.0658 |
| Tengchong | 12 | 10.5 | 196,409.9 | 15,743.5 | 0.0876 | 0.2798 | 0.3105 | 0.0923 |
| Zhaotong | 21 | 7.9 | 112,960.4 | 12,476.9 | 0.0504 | 0.2802 | 0.3089 | 0.1446 |
| Akhal_Teke | 20 | 10.7 | 106,929.8 | 9919.2 | 0.0477 | 0.2748 | 0.3082 | 0.0499 |
| Andalusian | 18 | 16.9 | 229,341.4 | 12,841.3 | 0.1022 | 0.2810 | 0.3102 | 0.1182 |
| Arabian | 24 | 14.0 | 156,306.3 | 10,144.7 | 0.0697 | 0.2829 | 0.3115 | 0.0857 |
| Belgian | 30 | 11.7 | 116,769.2 | 9753.1 | 0.0521 | 0.2928 | 0.3116 | 0.1566 |
| Caspian | 11 | 5.2 | 47,479.8 | 8895.2 | 0.0212 | 0.2864 | 0.3081 | 0.0210 |
| Finnhorse | 27 | 5.3 | 59,737.2 | 11,224.1 | 0.0266 | 0.2879 | 0.3064 | 0.1001 |
| Hanoverian | 15 | 9.5 | 82,050.0 | 8402.7 | 0.0366 | 0.2872 | 0.3078 | −0.0433 |
| Mongolian | 3 | 3.3 | 29,864.8 | 8312.9 | 0.0133 | 0.2891 | 0.3078 | 0.0793 |
| Morgan | 43 | 13.1 | 167,297.9 | 11,061.9 | 0.0746 | 0.2955 | 0.3088 | 0.0780 |
| Percheron | 20 | 7.1 | 63,734.8 | 8336.1 | 0.0284 | 0.2871 | 0.3082 | 0.1212 |
| Quarter | 40 | 9.3 | 88,200.5 | 9245.7 | 0.0393 | 0.2911 | 0.3090 | −0.0260 |
| Shetland | 27 | 17.5 | 185,601.4 | 10,109.3 | 0.0827 | 0.2844 | 0.3054 | 0.2434 |
| Thoroughbred | 36 | 23.6 | 222,096.8 | 9489.8 | 0.0990 | 0.2875 | 0.3059 | −0.0019 |
| Tuva | 11 | 4.0 | 58,334.8 | 10,996.3 | 0.0260 | 0.2851 | 0.3088 | 0.0675 |
| Gene Stable ID | Chr. | Begin | End | Know Genes |
|---|---|---|---|---|
| ENSECAG00000015256 | 11 | 26,497,130 | 26,746,723 | SPAG9 |
| ENSECAG00000028963 | 11 | 26,767,419 | 26,776,603 | NME1 |
| ENSECAG00000021658 | 11 | 26,779,860 | 26,782,544 | NME2 |
| ENSECAG00000022700 | 11 | 26,787,953 | 26,824,364 | MBTD1 |
| ENSECAG00000008260 | 11 | 26,852,773 | 26,888,439 | UTP18 |
| ENSECAG00000013061 | 11 | 27,143,031 | 27,609,842 | CA10 |
| A | O | X | C | A | O | B | C | F4 Ratio | Std.err | Z (null = 0) |
|---|---|---|---|---|---|---|---|---|---|---|
| NQ | Donkey | JJ | BS | NQ | Donkey | CKY | BS | 39.2431 | 12.654 | 3.1019d |
| NQ | Donkey | JJ | BS | NQ | Donkey | DA | BS | −0.0990 | 0.2196 | −0.4519d |
| NQ | Donkey | JJ | BS | NQ | Donkey | KZK | BS | −0.0674 | 0.5070 | −0.1339d |
| NQ | Donkey | JJ | BS | NQ | Donkey | LJ | BS | −0.6912 | 1.2101 | −0.5719d |
| NQ | Donkey | JJ | BS | NQ | Donkey | IMG | BS | −0.1099 | 0.3299 | −0.3339d |
| NQ | Donkey | JJ | BS | NQ | Donkey | TC | BS | −5.3502 | 3.0578 | −1.7509d |
| NQ | Donkey | JJ | BS | NQ | Donkey | ZT | BS | 0.3187 | 0.4074 | 0.7829d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, H.; Wang, S.; Zeng, G.; Guo, J.; Guo, M.; Dong, X.; Hua, G.; Liu, Y.; Wang, M.; Ling, Y.; et al. The Origin of a Coastal Indigenous Horse Breed in China Revealed by Genome-Wide SNP Data. Genes 2019, 10, 241. https://doi.org/10.3390/genes10030241
Ma H, Wang S, Zeng G, Guo J, Guo M, Dong X, Hua G, Liu Y, Wang M, Ling Y, et al. The Origin of a Coastal Indigenous Horse Breed in China Revealed by Genome-Wide SNP Data. Genes. 2019; 10(3):241. https://doi.org/10.3390/genes10030241
Chicago/Turabian StyleMa, Hongying, Sheng Wang, Guorong Zeng, Jintu Guo, Minghao Guo, Xianggui Dong, Guoying Hua, Yu Liu, Min Wang, Yao Ling, and et al. 2019. "The Origin of a Coastal Indigenous Horse Breed in China Revealed by Genome-Wide SNP Data" Genes 10, no. 3: 241. https://doi.org/10.3390/genes10030241