Novel Characteristics of Mitochondrial Electron Transport Chain from Eimeria tenella

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sequence Analysis
2.2. Preparations of Eimeria tenella Oocysts and Sporozoitess
2.3. Transmission Electron Microscopy (TEM) of Eimeria tenella
2.4. Preparation of Eimeria tenella Mitochondria-rich Fraction
2.5. Enzyme Assays
2.6. Analysis of the Quinone from Eimeria tenella
2.7. Enzyme Inhibition Assay of Eimeria tenella Succinate Dehydrogenase
2.8. High Resolution Clear Native Electrophoresis (HrCNE) and Succinate Dehydrogenase Activity Staining
2.9. Two Dimentional Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (2D-SDS-PAGE) and Nano- Liquid Chromatography Tandem-Mass Spectrometry (LC MS/MS) Analysis of Eimeria tenella Succinate Dehydrogenase Stained Band
3. Results
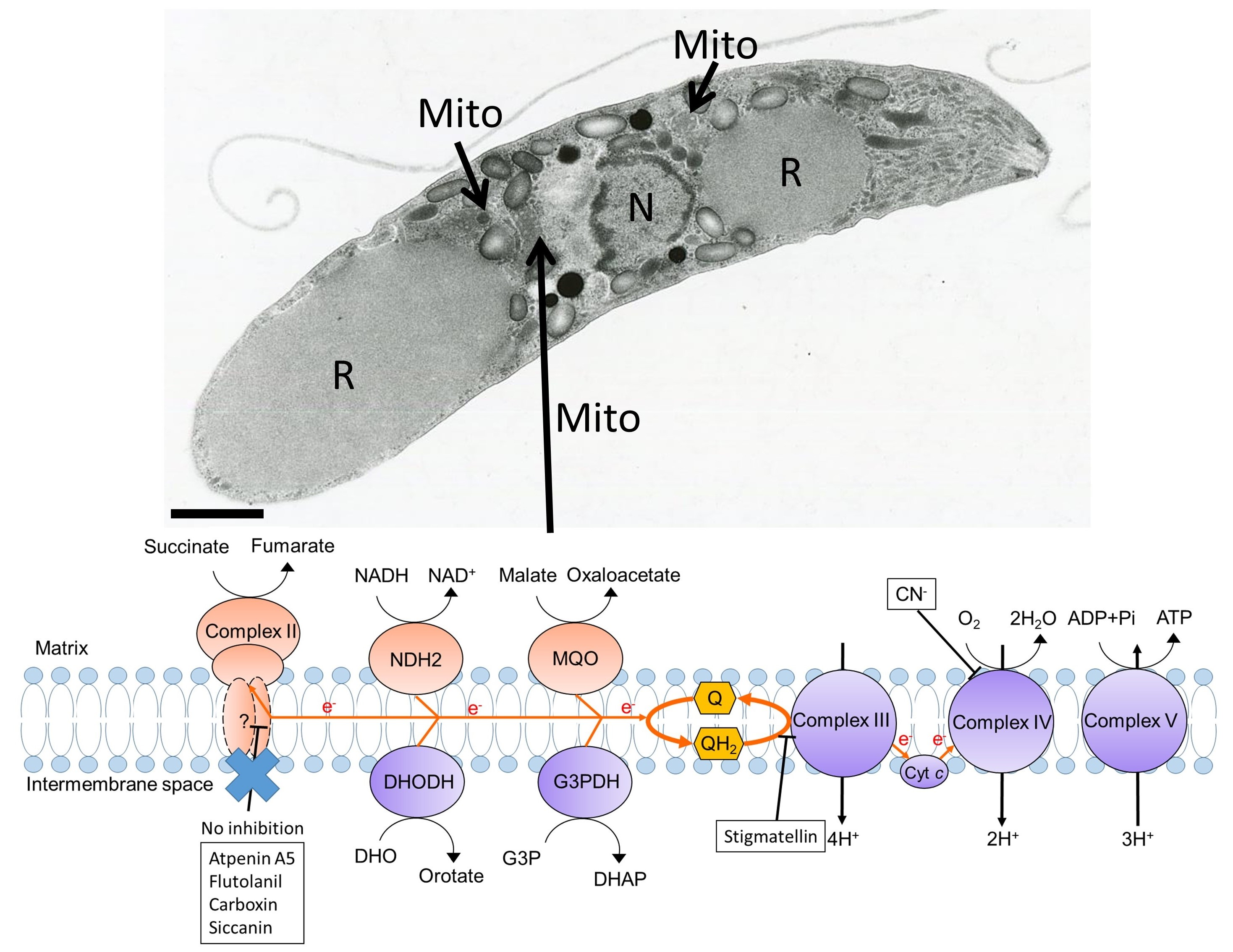
3.1. Electron Transport Chain Dehydrogenases from Eimeria tenella
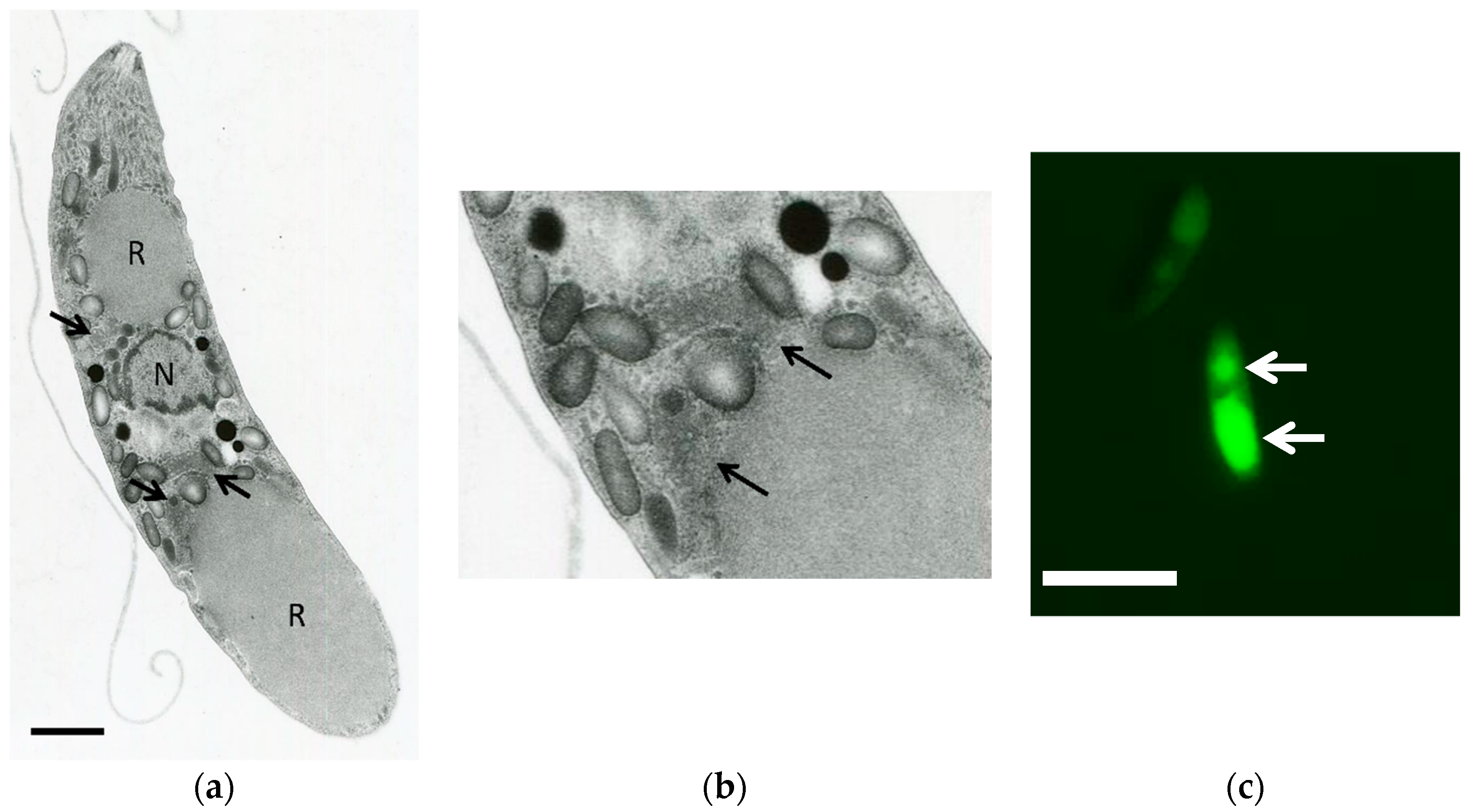
3.2. Transmission Electron Microscopy Analysis of Eimeria tenella
3.3. Preparation of Mitochondria-Rich Fraction
3.4. Characterization of Eimeria tenella Mitochondrial Electron Transport Chain.
3.5. Inhibition Studies of Eimeria tenella Mitochondrial Complex II
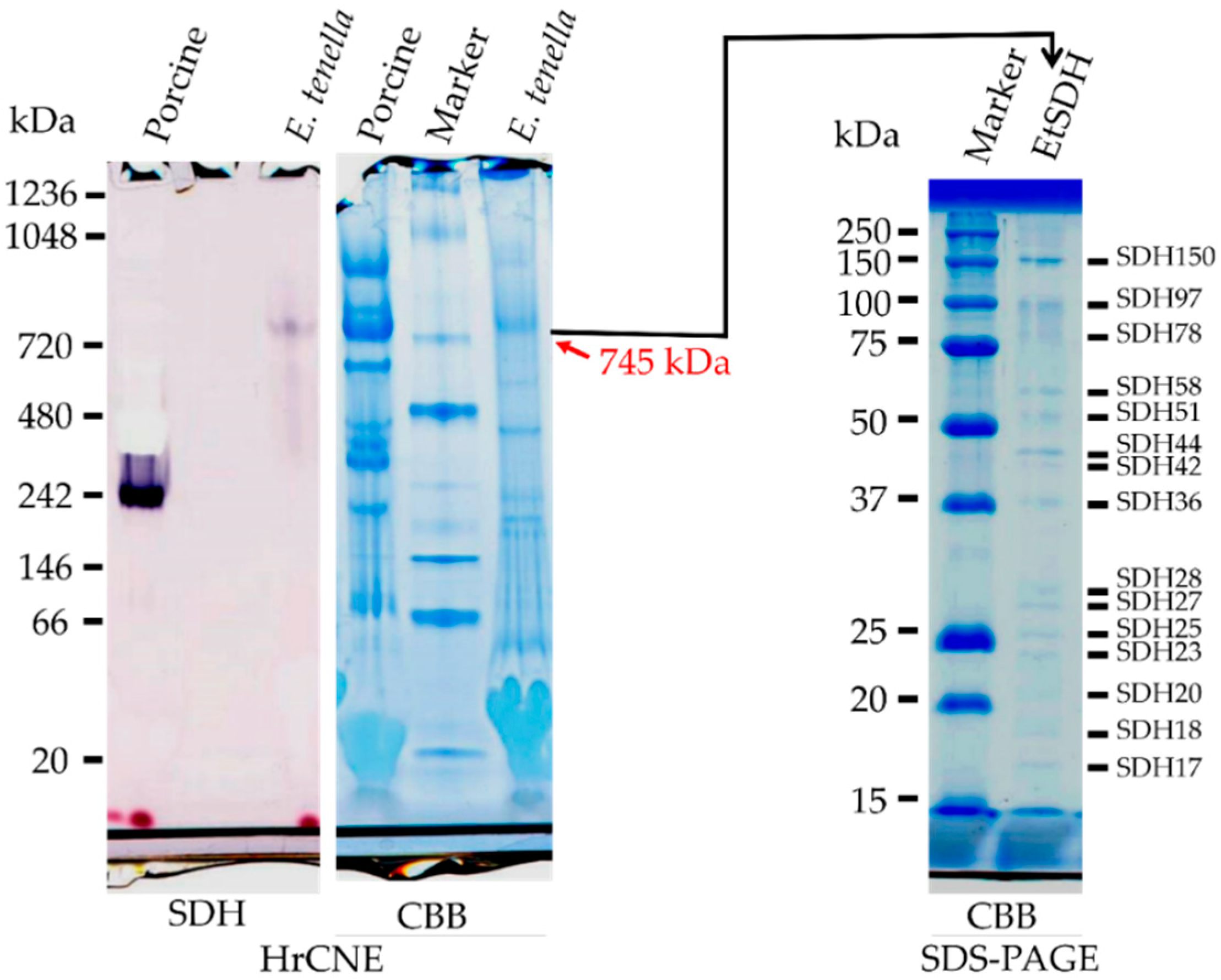
3.6. HrCNE and Nano-LC MS/MS Analyses of Eimeria tenella Mitochondrial Complex II
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Naciri, P.M.; Brossier, F. Avian coccidiosis: Importance and research prospects. Bull. Acad. Vét France 2009, 162, 4. [Google Scholar] [CrossRef]
- Li, G.Q.; Kanu, S.; Xiao, S.M.; Xiang, F.Y. Responses of chickens vaccinated with a live attenuated multi-valent ionophore-tolerant Eimeria vaccine. Vet. Parasitol. 2005, 129, 179–186. [Google Scholar] [CrossRef] [PubMed]
- McDonald, V.; Shirley, M.W. The endogenous development of virulent strains and attenuated precocious lines of Eimeria tenella and E. necatrix. J. Parasitol. 1987, 73, 993–997. [Google Scholar] [CrossRef]
- Kheysin, Y.M. Life Cycles of Coccidia of Domestic Animals; Willian Heinemann Medical Books Limited: London, UK, 1972. [Google Scholar]
- Hermans, D.; Van Deun, K.; Martel, A.; Van Immerseel, F.; Messens, W.; Heyndrickx, M.; Haesebrouck, F.; Pasmans, F. Colonization factors of Campylobacter jejuni in the chicken gut. Vet. Res. 2011, 42. [Google Scholar] [CrossRef] [PubMed]
- Harada, S.; Inaoka, D.K.; Ohmori, J.; Kita, K. Diversity of parasite complex II. Biochim. Biophys. Acta 2013, 1827, 658–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inaoka, D.K.; Shiba, T.; Sato, D.; Balogun, E.O.; Sasaki, T.; Nagahama, M.; Oda, M.; Matsuoka, S.; Ohmori, J.; Honma, T.; et al. Structural insights into the molecular design of flutolanil derivatives targeted for fumarate respiration of parasite mitochondria. Int. J. Mol. Sci. 2015, 16, 15287–15308. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Miyadera, H.; Ui, H.; Shiomi, K.; Yamaguchi, Y.; Masuma, R.; Nagamitsu, T.; Takano, D.; Sunazuka, T.; Harder, A.; et al. An anthelmintic compound, nafuredin, shows selective inhibition of complex I in helminth mitochondria. Proc. Natl. Acad. Sci. USA 2001, 98, 60–62. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, J.; Sakamoto, K.; Shinjyo, N.; Kido, Y.; Yamamoto, N.; Yagi, K.; Miyoshi, H.; Nonaka, N.; Katakura, K.; Kita, K.; et al. Anaerobic NADH-fumarate reductase system is predominant in the respiratory chain of Echinococcus multilocularis, providing a novel target for the chemotherapy of alveolar echinococcosis. Antimicrob. Agents Chemother. 2008, 52, 164–170. [Google Scholar] [CrossRef]
- Iwata, F.; Shinjyo, N.; Amino, H.; Sakamoto, K.; Islam, M.K.; Tsuji, N.; Kita, K. Change of subunit composition of mitochondrial complex II (succinate-ubiquinone reductase/quinol-fumarate reductase) in Ascaris suum during the migration in the experimental host. Parasitol. Int. 2008, 57, 54–61. [Google Scholar] [CrossRef]
- Sakai, C.; Tomitsuka, E.; Esumi, H.; Harada, S.; Kita, K. Mitochondrial fumarate reductase as a target of chemotherapy: From parasites to cancer cells. Biochim. Biophys. Acta 2012, 1820, 643–651. [Google Scholar] [CrossRef]
- Bulusu, V.; Jayaraman, V.; Balaram, H. Metabolic fate of fumarate, a side product of the purine salvage pathway in the intraerythrocytic stages of Plasmodium falciparum. J. Biol. Chem. 2011, 286, 9236–9245. [Google Scholar] [CrossRef]
- Hartuti, E.D.; Inaoka, D.K.; Komatsuya, K.; Miyazaki, Y.; Miller, R.J.; Xinying, W.; Sadikin, M.; Prabandari, E.E.; Waluyo, D.; Kuroda, M.; et al. Biochemical studies of membrane bound Plasmodium falciparum mitochondrial L-malate:quinone oxidoreductase, a potential drug target. Biochim. Biophys. Acta 2018, 1859, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Niikura, M.; Komatsuya, K.; Inoue, S.I.; Matsuda, R.; Asahi, H.; Inaoka, D.K.; Kita, K.; Kobayashi, F. Suppression of experimental cerebral malaria by disruption of malate:quinone oxidoreductase. Malar. J. 2017, 16, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonhosolo, R.; Gabriel, H.B.; Matsumura, M.Y.; Cabral, F.J.; Yamamoto, M.M.; D’Alexandri, F.L.; Sussmann, R.A.C.; Belmonte, R.; Peres, V.J.; Crick, D.C.; et al. Intraerythrocytic stages of Plasmodium falciparum biosynthesize menaquinone. FEBS Lett. 2010, 584, 4761–4768. [Google Scholar] [CrossRef] [PubMed]
- Painter, H.J.; Morrisey, J.M.; Mather, M.W.; Vaidya, A.B. Specific role of mitochondrial electron transport in blood-stage Plasmodium falciparum. Nature 2007, 446, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Scholtyseck, E. The Coccidia—Eimeria, Isospora, Toxoplasma and Related Genera; Hammond, D.M., Long, P.L., Eds.; University Park Press: Baltimore, MD, USA; London, UK, 1973; pp. 81–144. [Google Scholar]
- Reid, A.J.; Blake, D.P.; Ansari, H.R.; Billington, K.; Browne, H.P.; Bryant, J.; Dunn, M.; Hung, S.S.; Kawahara, F.; Miranda-Saavedra, D.; et al. Genomic analysis of the causative agents of coccidiosis in domestic chickens. Genome Res. 2014, 24, 1676–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubayashi, M.; Hatta, T.; Miyoshi, T.; Anisuzzaman; Sasai, K.; Shimura, K.; Isobe, T.; Kita, K.; Tsuji, N. High-throughput RNA sequencing profiles and transcriptional evidence of aerobic respiratory enzymes in sporulating oocysts and sporozoites of Eimeria tenella. Infect. Genet. Evol. 2013, 18, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Sheather, A.L. The detection of intestinal protozoa and mange parasites by a floatation technique. J. Comp. Pathol. Ther. 1923, 36, 10. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Shimada, H.; Kodjabachian, D.; Ishida, M. Specific and rapid analysis of ubiquinones using Craven’s reaction and HPLC with postcolumn derivatization. J. Lipid Res. 2007, 48, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- Miyadera, H.; Shiomi, K.; Ui, H.; Yamaguchi, Y.; Masuma, R.; Tomoda, H.; Miyoshi, H.; Osanai, A.; Kita, K.; Omura, S. Atpenins, potent and specific inhibitors of mitochondrial complex II (succinate-ubiquinone oxidoreductase). Proc. Natl. Acad. Sci. USA 2003, 100, 473–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogi, T.; Kawakami, T.; Arai, H.; Igarashi, Y.; Matsushita, K.; Mori, M.; Shiomi, K.; Omura, S.; Harada, S.; Kita, K. Siccanin rediscovered as a species-selective succinate dehydrogenase inhibitor. J. Biochem. 2009, 146, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Nihashi, N.; Inaoka, D.K.; Tsuge, C.; Balogun, E.O.; Osada, Y.; Goto, Y.; Matsumoto, Y.; Nara, T.; Mogi, T.; Harada, S.; et al. Siccanin is a novel selective inhibitor of trypanosomatid complex II (succinate-ubiquinone reductase) and a potent broad-spectrum anti-trypanosomatid drug candidate. In Kala Azar in South Asia: Current Status and Sustainable Challenges; Noiri, E., Jha, T.K., Eds.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Osanai, A.; Harada, S.; Sakamoto, K.; Shimizu, H.; Inaoka, D.K.; Kita, K. Crystallization of mitochondrial rhodoquinol-fumarate reductase from the parasitic nematode Ascaris suum with the specific inhibitor flutolanil. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2009, 65, 941–944. [Google Scholar] [CrossRef] [PubMed]
- Von Jagow, G.; Ohnishi, T. The chromone inhibitor stigmatellin—Binding to the ubiquinol oxidation center at the C-side of the mitochondrial membrane. FEBS Lett. 1985, 185, 311–315. [Google Scholar] [CrossRef]
- Yamashita, T.; Inaoka, D.K.; Shiba, T.; Oohashi, T.; Iwata, S.; Yagi, T.; Kosaka, H.; Miyoshi, H.; Harada, S.; Kita, K.; et al. Ubiquinone binding site of yeast NADH dehydrogenase revealed by structures binding novel competitive- and mixed-type inhibitors. Sci. Rep. 2018, 8, 2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomitsuka, E.; Kita, K.; Esumi, H. Regulation of succinate-ubiquinone reductase and fumarate reductase activities in human complex II by phosphorylation of its flavoprotein subunit. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 258–265. [Google Scholar] [CrossRef]
- Mogi, T.; Kita, K. Diversity in mitochondrial metabolic pathways in parasitic protists Plasmodium and Cryptosporidium. Parasitol. Int. 2010, 59, 305–312. [Google Scholar] [CrossRef]
- Madiraju, A.K.; Erion, D.M.; Rahimi, Y.; Zhang, X.M.; Braddock, D.T.; Albright, R.A.; Prigaro, B.J.; Wood, J.L.; Bhanot, S.; MacDonald, M.J.; et al. Metformin suppresses gluconeogenesis by inhibiting mitochondrial glycerophosphate dehydrogenase. Nature 2014, 510, 542–546. [Google Scholar] [CrossRef]
- Sato, T.; Morita, A.; Mori, N.; Miura, S. The role of glycerol-3-phosphate dehydrogenase 1 in the progression of fatty liver after acute ethanol administration in mice. Biochem. Biophys. Res. Commun. 2014, 444, 525–530. [Google Scholar] [CrossRef]
- Inaoka, D.K.; Iida, M.; Tabuchi, T.; Honma, T.; Lee, N.; Hashimoto, S.; Matsuoka, S.; Kuranaga, T.; Sato, K.; Shiba, T.; et al. The open form inducer approach for structure-based drug design. PLoS ONE 2016, 11, e0167078. [Google Scholar] [CrossRef] [PubMed]
- Inaoka, D.K.; Sakamoto, K.; Shimizu, H.; Shiba, T.; Kurisu, G.; Nara, T.; Aoki, T.; Kita, K.; Harada, S. Structures of Trypanosoma cruzi dihydroorotate dehydrogenase complexed with substrates and products: Atomic resolution insights into mechanisms of dihydroorotate oxidation and fumarate reduction. Biochemistry 2008, 47, 10881–10891. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Inaoka, D.K.; Shiba, T.; Saimoto, H.; Sakura, T.; Amalia, E.; Kido, Y.; Sakai, C.; Nakamura, M.; Moore, A.L.; et al. Selective cytotoxicity of dihydroorotate dehydrogenase inhibitors to human cancer cells under hypoxia and nutrient-deprived conditions. Front. Pharmacol. 2018, 9, 997. [Google Scholar] [CrossRef] [PubMed]
- Unden, G.; Bongaerts, J. Alternative respiratory pathways of Escherichia coli: Energetics and transcriptional regulation in response to electron acceptors. Biochim. Biophys. Acta 1997, 1320, 217–234. [Google Scholar] [CrossRef]
- Morales, J.; Mogi, T.; Mineki, S.; Takashima, E.; Mineki, R.; Hirawake, H.; Sakamoto, K.; Omura, S.; Kita, K. Novel mitochondrial complex II isolated from Trypanosoma cruzi is composed of 12 peptides including a heterodimeric Ip subunit. J. Biol. Chem. 2009, 284, 7255–7263. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, K.; Mogi, T.; Tanaka, T.Q.; Hata, M.; Miyoshi, H.; Kita, K. Mitochondrial dehydrogenases in the aerobic respiratory chain of the rodent malaria parasite Plasmodium yoelii yoelii. J. Biochem. 2009, 145, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Muhleip, A.W.; Joos, F.; Wigge, C.; Frangakis, A.S.; Kuhlbrandt, W.; Davies, K.M. Helical arrays of U-shaped ATP synthase dimers form tubular cristae in ciliate mitochondria. Proc. Natl. Acad. Sci. USA 2016, 113, 8442–8447. [Google Scholar] [CrossRef]
- Krungkrai, J. The multiple roles of the mitochondrion of the malarial parasite. Parasitology 2004, 129, 511–524. [Google Scholar] [CrossRef]
- Sturm, A.; Mollard, V.; Cozijnsen, A.; Goodman, C.D.; McFadden, G.I. Mitochondrial ATP synthase is dispensable in blood-stage Plasmodium berghei rodent malaria but essential in the mosquito phase. Proc. Natl. Acad. Sci. USA 2015, 112, 10216–10223. [Google Scholar] [CrossRef]
- Lemgruber, L.; Lupetti, P. Crystalloid body, refractile body and virus-like particles in Apicomplexa: What is in there? Parasitology 2012, 139, 285–293. [Google Scholar] [CrossRef]
- Wang, C.C.; Weppelman, R.M.; Lopez-Ramos, B. Isolation of amylopectin granules and identification of amylopectin phosphorylase in the oocysts of Eimeria tenella. J. Protozool. 1975, 22, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Komatsuya, K.; Hata, M.; Balogun, E.O.; Hikosaka, K.; Suzuki, S.; Takahashi, K.; Tanaka, T.; Nakajima, M.; Ogura, S.; Sato, S.; et al. Synergy of ferrous ion on 5-aminolevulinic acid-mediated growth inhibition of Plasmodium falciparum. J. Biochem. 2013, 154, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Siregar, J.E.; Kurisu, G.; Kobayashi, T.; Matsuzaki, M.; Sakamoto, K.; Mi-ichi, F.; Watanabe, Y.; Hirai, M.; Matsuoka, H.; Syafruddin, D.; et al. Direct evidence for the atovaquone action on the Plasmodium cytochrome bc1 complex. Parasitol. Int. 2015, 64, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Takashima, E.; Takamiya, S.; Takeo, S.; Mi-ichi, F.; Amino, H.; Kita, K. Isolation of mitochondria from Plasmodium falciparum showing dihydroorotate dependent respiration. Parasitol. Int. 2001, 50, 273–278. [Google Scholar] [CrossRef]
- Hino, A.; Hirai, M.; Tanaka, T.Q.; Watanabe, Y.; Matsuoka, H.; Kita, K. Critical roles of the mitochondrial complex II in oocyst formation of rodent malaria parasite Plasmodium berghei. J. Biochem. 2012, 152, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.Q.; Hirai, M.; Watanabe, Y.; Kita, K. Toward understanding the role of mitochondrial complex II in the intraerythrocytic stages of Plasmodium falciparum: Gene targeting of the Fp subunit. Parasitol. Int. 2012, 61, 726–728. [Google Scholar] [CrossRef]
- Roberts, W.L.; Hammond, D.M. Ultrastructural and cytologic studies of the sporozoites of four Eimeria species. J. Protozool. 1970, 17, 76–86. [Google Scholar] [CrossRef]
- Lane, A.N.; Fan, T.W. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Morcillo, M.; Grande-Garcia, A.; Ruiz-Ramos, A.; Del Cano-Ochoa, F.; Boskovic, J.; Ramon-Maiques, S. Structural insight into the core of CAD, the multifunctional protein leading de novo pyrimidine biosynthesis. Structure 2017, 25, 912–923.e5. [Google Scholar] [CrossRef]
- Ryley, J.F.; Bentley, M.; Manners, D.J.; Stark, J.R. Amylopectin, the storage polysaccharide of the Coccidia Eimeria brunetti and E. tenella. J. Parasitol. 1969, 55, 839–845. [Google Scholar] [CrossRef]
- Schmatz, D.M. The mannitol cycle in Eimeria. Parasitology 1997, 114, S81–S89. [Google Scholar] [PubMed]
- Van Hellemond, J.J.; Tielens, A.G. Expression and functional properties of fumarate reductase. Biochem. J. 1994, 304, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kita, K.; Takamiya, S.; Furushima, R.; Ma, Y.C.; Suzuki, H.; Ozawa, T.; Oya, H. Electron-transfer complexes of Ascaris suum muscle mitochondria. III. Composition and fumarate reductase activity of complex II. Biochim. Biophys. Acta 1988, 935, 130–140. [Google Scholar] [CrossRef]
- Tomitsuka, E.; Kita, K.; Esumi, H. The NADH-fumarate reductase system, a novel mitochondrial energy metabolism, is a new target for anticancer therapy in tumor microenvironments. Ann. N. Y. Acad. Sci. 2010, 1201, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Tomitsuka, E.; Kita, K.; Esumi, H. An anticancer agent, pyrvinium pamoate inhibits the NADH-fumarate reductase system—A unique mitochondrial energy metabolism in tumour microenvironments. J. Biochem. 2012, 152, 171–183. [Google Scholar] [CrossRef]
- Fry, M.; Beesley, J.E. Mitochondria of mammalian Plasmodium spp. Parasitology 1991, 102, 17–26. [Google Scholar] [CrossRef]
- Millar, A.H.; Eubel, H.; Jansch, L.; Kruft, V.; Heazlewood, J.L.; Braun, H.P. Mitochondrial cytochrome c oxidase and succinate dehydrogenase complexes contain plant specific subunits. Plant Mol. Biol. 2004, 56, 77–90. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| NADH-cyt c | Succinate-cyt c | |
|---|---|---|
| Day 0-Polytron | 0.011 (100%) | 0.004 (100%) |
| Day 0-N2 Cavitation | 0.050 (100%) | 0.010 (100%) |
| Day 3–4 °C-Polytron | 0.006 (57%) | 0.003 (63%) |
| Day 3–4 °C-N2 Cavitation | 0.008 (16%) | 0.007 (69%) |
| Day 3–80 °C-Polytron | 0.011 (100%) | 0.026 (65%) |
| Day 3–80 °C-N2 Cavitation | 0.037 (74%) | 0.009 (87%) |
| Specific Activity (nmol/min/mg) | ||||
|---|---|---|---|---|
| A | B | C | D | |
| NADH-cyt c 1 | 341 ± 18 | 228 ± 16 | 167 ± 8 | 189 ± 28 |
| G3P-cyt c 1 | 137 ± 3 | 94 ± 1 | 101 ± 6 | 119 ± 5 |
| Malate-cyt c 1 | 108 ± 9 | 75 ± 9 | 95 ± 5 | 77 ± 9 |
| Succinate-cyt c 1 | 50 ± 7 | 50 ± 7 | 71 ± 2 | 54 ± 2 |
| Dihydroorotate-cyt c 1 | 0.0 | 0.0 | 1.7 | 1.7 |
| Specific Activity 1 | |
|---|---|
| NADH oxidase | 5.9 ± 0.9 |
| + 2 mM KCN | 0.0 ± 0.3 |
| + 2 mM fumarate | 1.3 ± 0.3 |
| + 5 mM malonate | −0.5 ± 0.3 |
| NADH-cyt c | 167 ± 8 |
| NADH-dUQ | 224 ± 19 |
| G3P-cyt c | 101 ± 6 |
| G3P-dUQ | 61 ± 2 |
| Malate-cyt c | 95 ± 5 |
| Malate-dUQ | 67 ± 3 |
| Succinate-cyt c | 71 ± 2 |
| Succinate-dUQ | 25 ± 2 |
| Dihydroorotate-cyt c | 1.7 |
| IC50 of Succinate-cyt c Activity (μM) | ||||
|---|---|---|---|---|
| E. tenella | P. y. yoelii [39] | Rat | Porcine | |
| Atpenin A5 | >50 | 4.2 ± 0.2 [39] | 0.007 ± 0.0003 [39] | N.D. |
| Siccanin | 4.0 ± 0.6 (>50) 1 | N.D. [39] | 9.3 ± 1.0 [25] | 861 ± 822 [25] |
| Carboxin | >50 | 3.6 ± 1.0 [39] | 3.8 ± 0.1 [39] | N.D. |
| Flutolanil | >50 | >100 [39] | >100 [39] | >50 [7] |
| Stigmatellin | 78.4 ± 13.1% 2 | N.D. | N.D. | N.D. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsubayashi, M.; Inaoka, D.K.; Komatsuya, K.; Hatta, T.; Kawahara, F.; Sakamoto, K.; Hikosaka, K.; Yamagishi, J.; Sasai, K.; Shiba, T.; et al. Novel Characteristics of Mitochondrial Electron Transport Chain from Eimeria tenella. Genes 2019, 10, 29. https://doi.org/10.3390/genes10010029
Matsubayashi M, Inaoka DK, Komatsuya K, Hatta T, Kawahara F, Sakamoto K, Hikosaka K, Yamagishi J, Sasai K, Shiba T, et al. Novel Characteristics of Mitochondrial Electron Transport Chain from Eimeria tenella. Genes. 2019; 10(1):29. https://doi.org/10.3390/genes10010029
Chicago/Turabian StyleMatsubayashi, Makoto, Daniel Ken Inaoka, Keisuke Komatsuya, Takeshi Hatta, Fumiya Kawahara, Kimitoshi Sakamoto, Kenji Hikosaka, Junya Yamagishi, Kazumi Sasai, Tomoo Shiba, and et al. 2019. "Novel Characteristics of Mitochondrial Electron Transport Chain from Eimeria tenella" Genes 10, no. 1: 29. https://doi.org/10.3390/genes10010029