Reciprocal Regulation of TRPS1 and miR-221 in Intervertebral Disc Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Tissue Samples
2.2. Isolation of Human IVD Cells
2.3. Cell Transfection
2.4. Luciferase Reporter Gene Assay
2.5. Immunocytochemistry
2.6. RNA Extraction and Quantitative Real-Time (qRT)-PCR
2.7. Histochemical Analysis
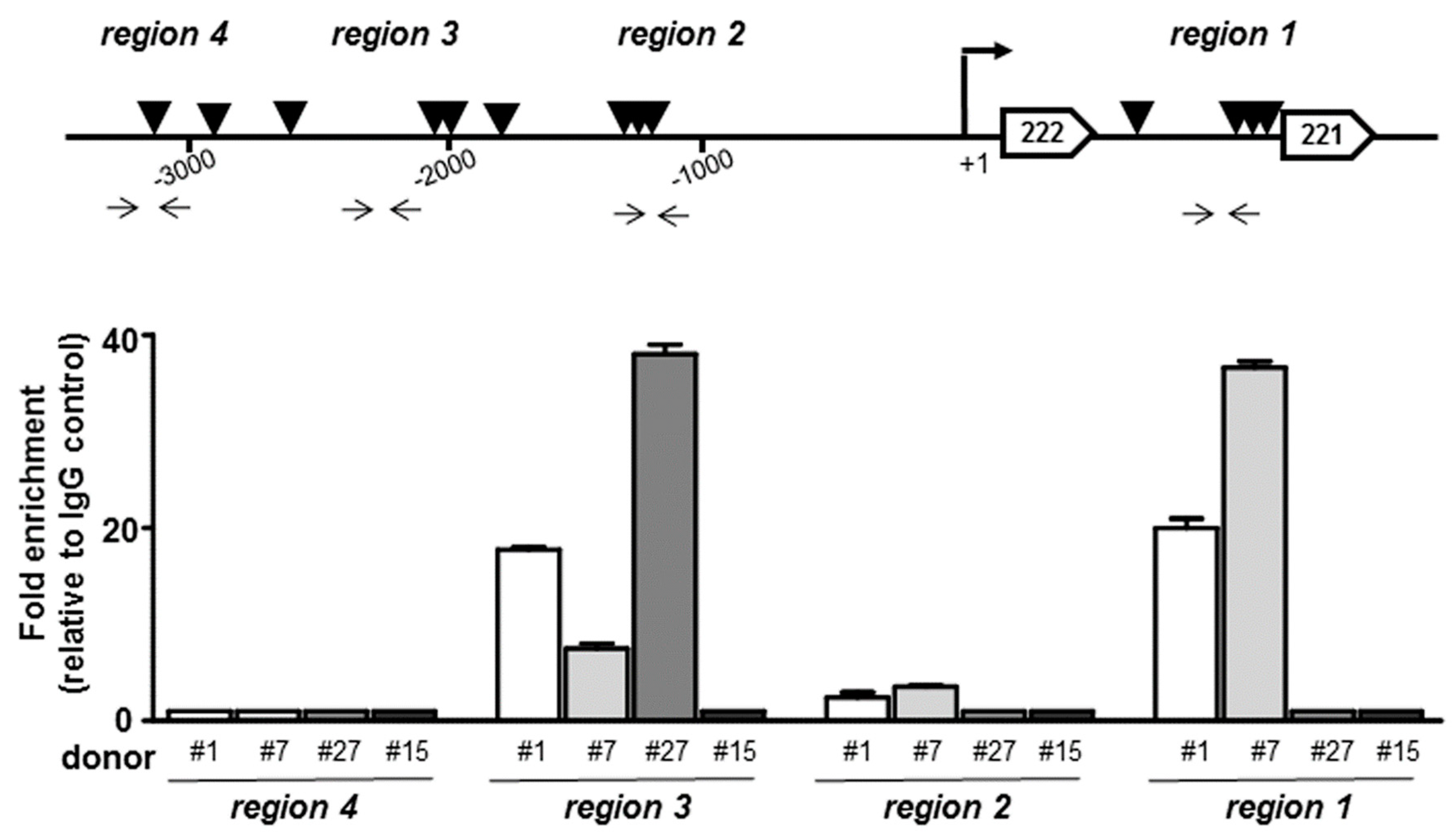
2.8. In Silico Analysis of the miR221/222 Human Promoter
2.9. Chromatin Immunoprecipitation (ChIP) Assay
2.10. Statistical Analysis
3. Results
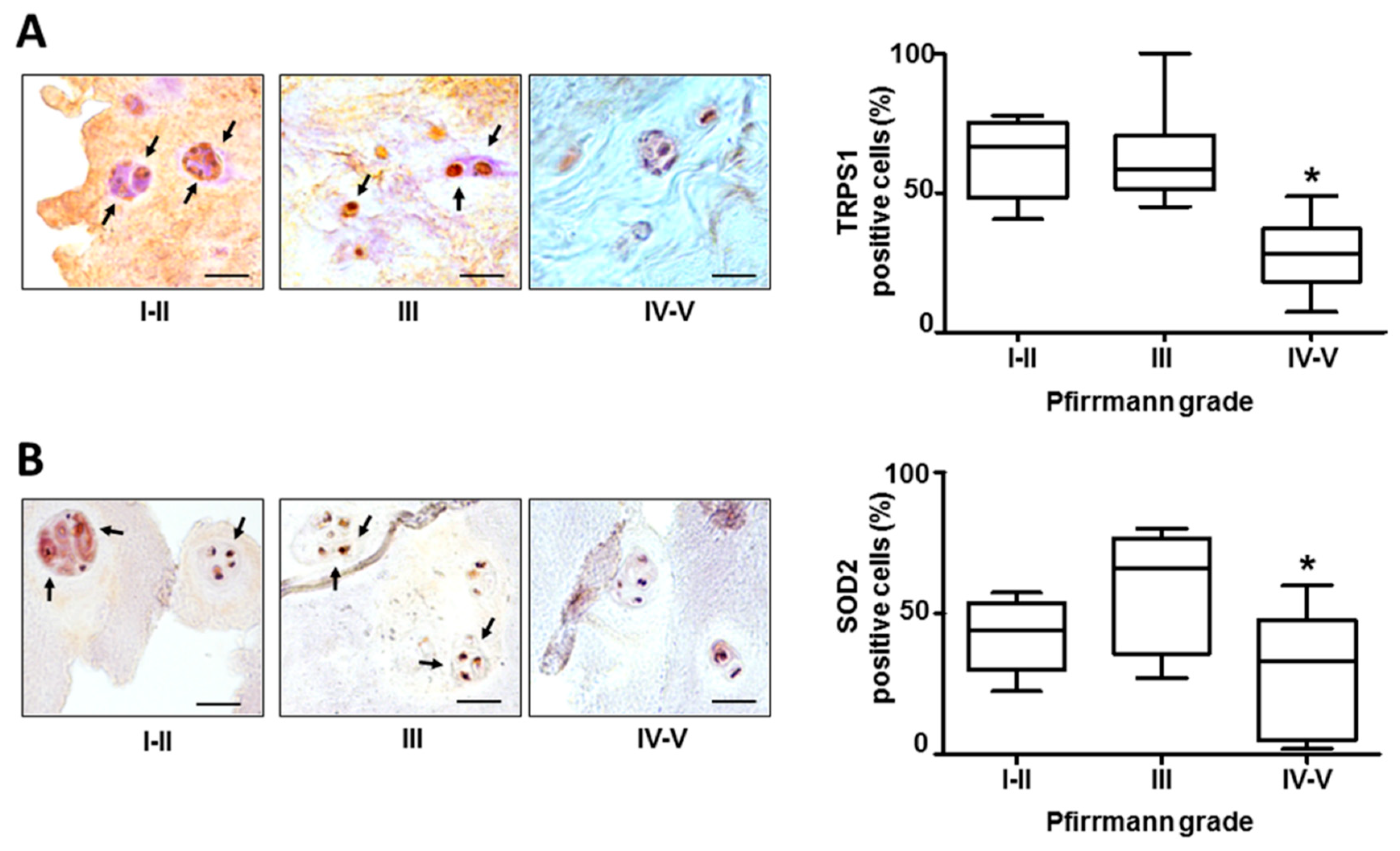
3.1. The Expression of TRPS1 in Lumbar Degenerated IVD Tissues
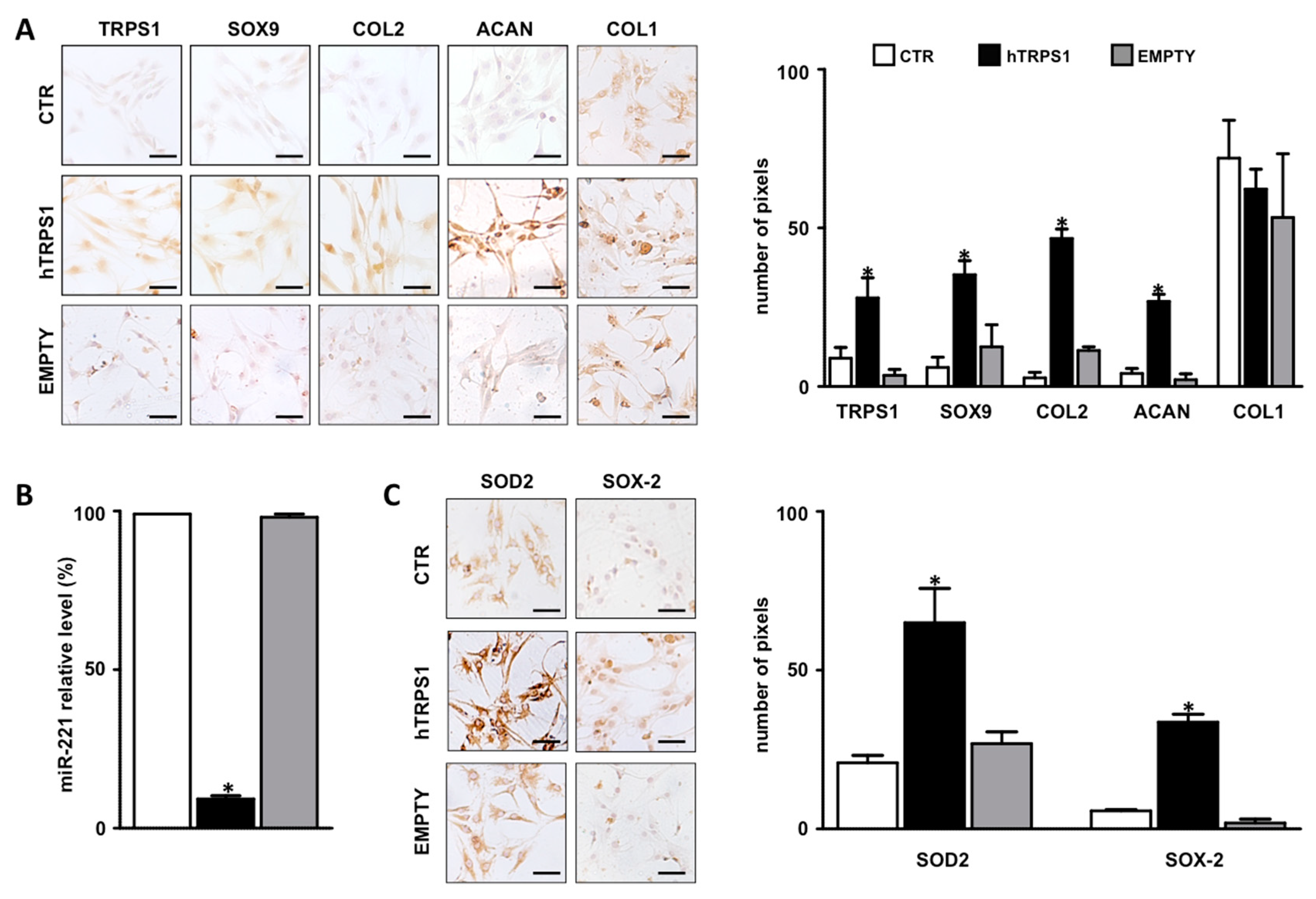
3.2. TRPS1 Signaling and miR-221
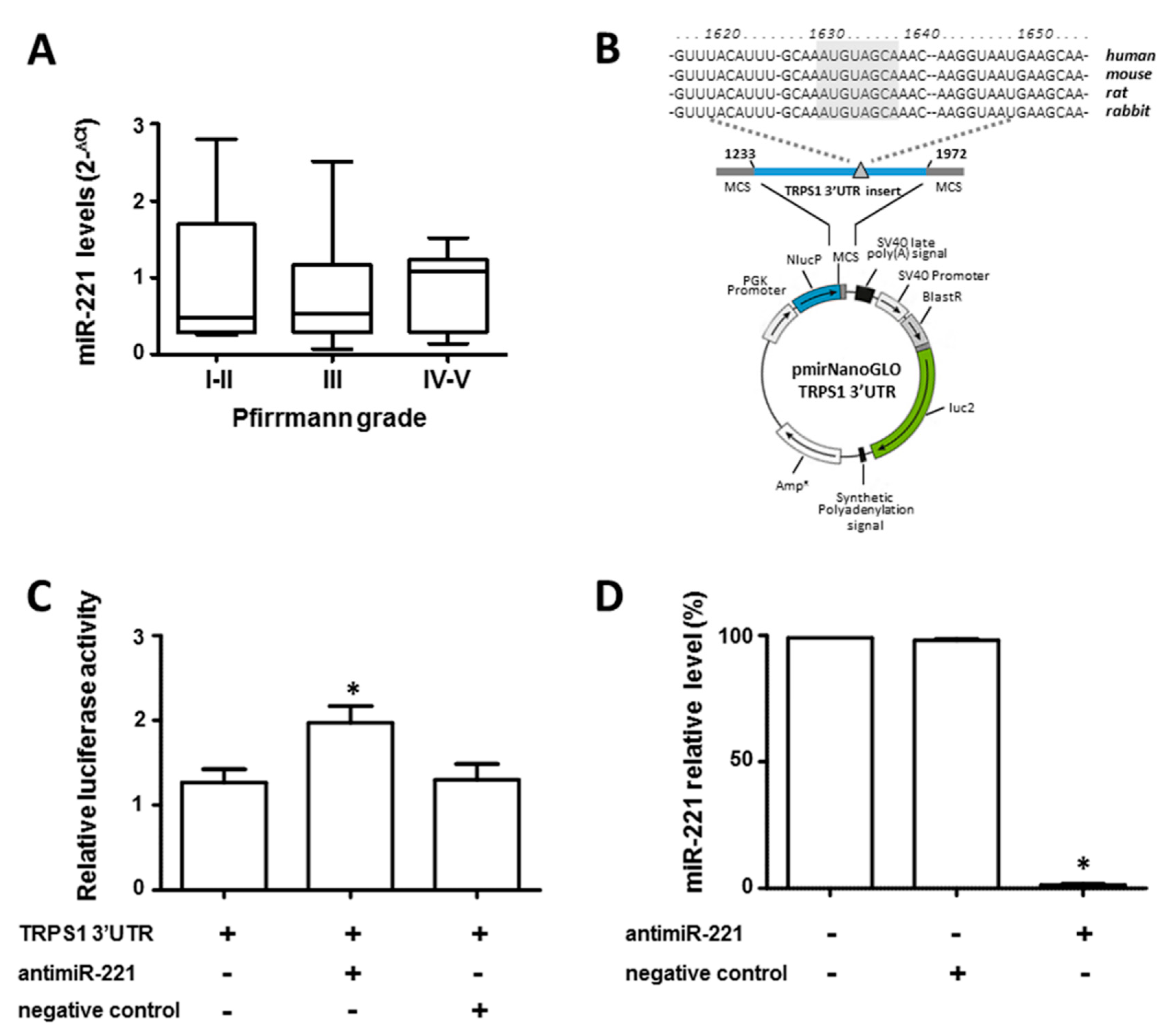
3.3. miR-221 Directly Targets TRPS1
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Richardson, S.M.; Freemont, A.J.; Hoyland, J.A. Pathogenesis of intervertebral disc degeneration. In The Intervertebral Disc-Molecular and Structural Studies of the Disc in Health and Disease; Shapiro, I.M., Risbud, M.V., Eds.; Springer: Vienna, Austria, 2014; pp. 177–200. [Google Scholar]
- Shapiro, I.M.; Risbud, M.V. Introduction to the structure, function, and comparative anatomy of the vertebrae and the intervertebral disc. In The Intervertebral Disc-Molecular and Structural Studies of the Disc in Health and Disease; Shapiro, I.M., Risbud, M.V., Eds.; Springer: Vienna, Austria, 2014; pp. 3–16. [Google Scholar]
- Lyu, F.J.; Cheung, K.M.; Zheng, Z.; Wang, H.; Sakai, D.; Leung, V.Y. IVD progenitor cells: A new horizon for understanding disc homeostasis and repair. Nat. Rev. Rheumatol. 2019, 15, 102–112. [Google Scholar] [CrossRef]
- Dowdell, J.; Erwin, M.; Choma, T.; Vaccaro, A.; Iatridis, J.; Cho, S.K. Intervertebral disk degeneration and repair. Neurosurgery 2017, 80, S46–S54. [Google Scholar] [CrossRef] [PubMed]
- Erwin, W.M. Biologically based therapy for the intervertebral disk: Who is the patient? Glob. Spine J. 2013, 3, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Momeni, P.; Glöckner, G.; Schmidt, O.; von Holtum, D.; Albrecht, B.; Gillessen-Kaesbach, G.; Hennekam, R.; Meinecke, P.; Zabel, B.; Rosenthal, A.; et al. Mutations in a new gene, encoding a zinc-finger protein, cause tricho-rhino-phalangeal syndrome type I. Nat. Genet. 2000, 24, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Fantauzzo, K.A.; Christiano, A.M. Trps1 activates a network of secreted Wnt inhibitors and transcription factors crucial to vibrissa follicle morphogenesis. Development 2012, 139, 203–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witwicki, R.M.; Ekram, M.B.; Qiu, X.; Janiszewska, M.; Shu, S.; Kwon, M.; Trinh, A.; Frias, E.; Ramadan, N.; Hoffman, G.; et al. TRPS1 is a lineage-specific transcriptional dependency in breast cancer. Cell Rep. 2018, 25, 1255–1267. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, Y.; Liu, Y.; Yu, S.; Xie, H.; Shi, X.; Qin, S.; Ma, F.; Tan, T.Z.; Thiery, J.P.; et al. A central role for TRPS1 in the control of cell cycle and cancer development. Oncotarget 2014, 5, 7677–7690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, T.H.; Shoichet, S.A.; Latham, P.; Kroll, T.G.; Peters, L.L.; Shivdasani, R.A. Transcriptional repression and developmental functions of the atypical vertebrate GATA protein TRPS1. EMBO J. 2001, 20, 1715–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gai, Z.; Gui, T.; Muragaki, Y. The function of TRPS1 in the development and differentiation of bone, kidney, and hair follicles. Histol. Histopathol. 2011, 26, 915–921. [Google Scholar]
- Sohn, P.; Cox, M.; Chen, D.; Serra, R. Molecular profiling of the developing mouse axial skeleton: A role for Tgfbr2 in the development of the intervertebral disc. BMC Dev. Biol. 2010, 10, 29. [Google Scholar] [CrossRef]
- Kuzynski, M.; Goss, M.; Bottini, M.; Yadav, M.C.; Mobley, C.; Winters, T.; Poliard, A.; Kellermann, O.; Lee, B.; Millan, J.L.; et al. Dual role of the Trps1 transcription factor in dentin mineralization. J. Biol. Chem. 2014, 289, 27481–27493. [Google Scholar] [CrossRef] [PubMed]
- Wuelling, M.; Kaiser, F.J.; Buelens, L.A.; Braunholz, D.; Shivdasani, R.A.; Depping, R.; Vortkamp, A. Trps1, a regulator of chondrocyte proliferation and differentiation, interacts with the activator form of Gli3. Dev. Biol. 2009, 328, 40–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergroesen, P.P.; Kingma, I.; Emanuel, K.S.; Hoogendoorn, R.J.; Welting, T.J.; van Royen, B.J.; van Dieën, J.H.; Smit, T.H. Mechanics and biology in intervertebral disc degeneration: A vicious circle. Osteoarthr. Cartil. 2015, 23, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Henry, N.; Clouet, J.; Le Bideau, J.; Le Visage, C.; Guicheux, J. Innovative strategies for intervertebral disc regenerative medicine: From cell therapies to multiscale delivery systems. Biotechnol. Advan. 2018, 36, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Lolli, A.; Lambertini, E.; Penolazzi, L.; Angelozzi, M.; Morganti, C.; Franceschetti, T.; Pelucchi, S.; Gambari, R.; Piva, R. Pro-chondrogenic effect of miR-221 and slug depletion in human MSCs. Stem Cell. Rev. 2014, 10, 841–855. [Google Scholar] [CrossRef] [PubMed]
- Penolazzi, L.; Lambertini, E.; Bergamin, L.S.; Roncada, T.; De Bonis, P.; Cavallo, M.; Piva, R. MicroRNA-221 silencing attenuates the degenerated phenotype of intervertebral disc cells. Aging 2018, 10, 2001–2015. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, X.; Shen, J.; Chan, M.T.; Wu, W.K. MicroRNA in intervertebral disc degeneration. Cell. Prolif. 2015, 48, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, W.J.; Yan, Y.G.; Xiang, Y.X.; Zhang, J.; Tang, Z.H.; Jiang, Z.S. MicroRNAs: New players in intervertebral disc degeneration. Clin. Chim. Acta 2015, 450, 333–341. [Google Scholar] [CrossRef]
- Pfirrmann, C.W.; Metzdorf, A.; Zanetti, M.; Hodler, J.; Boos, N. Magnetic resonance classification of lumbar intervertebral disc degeneration. Spine 2001, 26, 1873–1878. [Google Scholar]
- Lambertini, E.; Lisignoli, G.; Torreggiani, E.; Manferdini, C.; Gabusi, E.; Franceschetti, T.; Penolazzi, L.; Gambari, R.; Facchini, A.; Piva, R. Slug gene expression supports human osteoblast maturation. Cell. Mol. Life Sci. 2009, 66, 3641–3653. [Google Scholar] [CrossRef]
- Lambertini, E.; Penolazzi, L.; Angelozzi, M.; Grassi, F.; Gambari, L.; Lisignoli, G.; De Bonis, P.; Cavallo, M.; Piva, R. The expression of cystathionine gamma-lyase is regulated by estrogen receptor alpha in human osteoblasts. Oncotarget 2017, 8, 101686–101696. [Google Scholar] [CrossRef] [PubMed]
- Candas, D.; Li, J.J. MnSOD in oxidative stress response-potential regulation via mitochondrial protein influx. Antioxid. Redox Signal. 2014, 20, 1599–1617. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Garcia, O.; Matsuzaki, T.; Olmer, M.; Masuda, K.; Lotz, M.K. Age-related reduction in the expression of FOXO transcription factors and correlations with intervertebral disc degeneration. J. Orthop. Res. 2017, 35, 2682–2691. [Google Scholar] [CrossRef] [PubMed]
- Lolli, A.; Narcisi, R.; Lambertini, E.; Penolazzi, L.; Angelozzi, M.; Kops, N.; Gasparini, S.; van Osch, G.J.; Piva, R. Silencing of antichondrogenic microRNA-221 in human mesenchymal stem cells promotes cartilage repair in vivo. Stem Cells 2016, 34, 1801–1811. [Google Scholar] [CrossRef]
- Sarkar, A.; Hochedlinger, K. The sox family of transcription factors: Versatile regulators of stem and progenitor cell fate. Cell Stem Cell 2013, 12, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, H.B.; Svala, E.; Skioldebrand, E.; Lindahl, A.; Brisby, H. Support of concept that migrating progenitor cells from stem cell niches contribute to normal regeneration of the adult mammal intervertebral disc: A descriptive study in the New Zealand white rabbit. Spine 2012, 37, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Stinson, S.; Lackner, M.R.; Adai, A.T.; Yu, N.; Kim, H.J.; O’Brien, C.; Spoerke, J.; Jhunjhunwala, S.; Boyd, Z.; Januario, T.; et al. TRPS1 targeting by miR-221/222 promotes the epithelial-to-mesenchymal transition in breast cancer. Sci. Signal. 2011, 4, ra41. [Google Scholar]
- Shamji, M.F.; Setton, L.A.; Jarvis, W.; So, S.; Chen, J.; Jing, L.; Bullock, R.; Isaacs, R.E.; Brown, C.; Richardson, W.J. Proinflammatory cytokine expression profile in degenerated and herniated human intervertebral disc tissues. Arthr. Rheum. 2010, 62, 1974–1982. [Google Scholar]
- Moriguchi, Y.; Alimi, M.; Khair, T.; Manolarakis, G.; Berlin, C.; Bonassar, L.J.; Härtl, R. Biological treatment approaches for degenerative disk disease: A literature review of in vivo animal and clinical data. Glob. Spine 2016, 6, 497–518. [Google Scholar] [CrossRef]
- Lan, P.H.; Liu, Z.H.; Pei, Y.J.; Wu, Z.G.; Yu, Y.; Yang, Y.F.; Liu, X.; Che, L.; Ma, C.J.; Xie, Y.K.; et al. Landscape of RNAs in human lumbar disc degeneration. Oncotarget 2016, 7, 63166–63176. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Quan, Z.; Zhang, Q.; Wang, Z.; Song, Q.; Zhuang, X.; Fu, C.; Xu, F.; Liu, Y.; Wang, Y.; et al. Comprehensive evaluation of differential lncRNA and gene expression in patients with intervertebral disc degeneration. Mol. Med. Rep. 2018, 18, 1504–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantenbein-Ritter, B.; Benneker, L.M.; Alini, M.; Grad, S. Differential response of human bone marrow stromal cells to either TGF-β(1) or rhGDF-5. Eur. Spine J. 2011, 20, 962–971. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, J.A.; Collins, J.A.; Loeser, R.F. Reactive oxygen species, aging and articular cartilage homeostasis. Free Radic. Biol. Med. 2019, 132, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.E.; McConnell, J.C.; Sherratt, M.J.; Derby, B.; Richardson, S.M.; Hoyland, J.A. Growth differentiation factor 6 and transforming growth factor-beta differentially mediate mesenchymal stem cell differentiation, composition, and micromechanical properties of nucleus pulposus constructs. Arthr. Res. Ther. 2014, 16, R67. [Google Scholar] [CrossRef]
- Naqvi, S.M.; Gansau, J.; Gibbons, D.; Buckley, C.T. In vitro co-culture and ex vivo organ culture assessment of primed and cryopreserved stromal cell microcapsules for intervertebral disc regeneration. Eur. Cell. Mater. 2019, 37, 134–152. [Google Scholar] [CrossRef]
- Risbud, M.V.; Schoepflin, Z.R.; Mwale, F.; Kandel, R.A.; Grad, S.; Iatridis, J.C.; Sakai, D.; Hoyland, J.A. Defining the phenotype of young healthy nucleus pulposus cells: Recommendations of the Spine Research Interest Group at the 2014 annual ORS meeting. J. Orthop. Res. 2015, 33, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, D.M.; Johansen, E.B.; Derynck, R. Identification of the GATA factor TRPS1 as a repressor of the osteocalcin promoter. J. Biol. Chem. 2009, 284, 31690–31703. [Google Scholar] [CrossRef]
- Di Leva, G.; Gasparini, P.; Piovan, C.; Ngankeu, A.; Garofalo, M.; Taccioli, C.; Iorio, M.V.; Li, M.; Volinia, S.; Alder, H.; et al. MicroRNA cluster 221-222 and estrogen receptor alpha interactions in breast cancer. J. Natl. Cancer Inst. 2010, 102, 706–721. [Google Scholar] [CrossRef]
- Hu, P.; Feng, B.; Wang, G.; Ning, B.; Jia, T. Microarray based analysis of gene regulation by microRNA in intervertebral disc degeneration. Mol. Med. Rep. 2015, 12, 4925–4930. [Google Scholar] [CrossRef]
- Liu, J.; Yu, J.; Jiang, W.; He, M.; Zhao, J. Targeting of CDKN1B by miR-222-3p may contribute to the development of intervertebral disc degeneration. FEBS Open Bio. 2019, 9, 728–735. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.; Zhou, X.; Wang, N.; Li, Z.; Zhou, Y.; Feng, J.; Shen, D.; Zhao, W. Knockdown of miR-222 inhibits inflammation and the apoptosis of LPS-stimulated human intervertebral disc nucleus pulposus cells. Int. J. Mol. Med. 2019, 44, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Smolders, L.A.; Meij, B.P.; Onis, D.; Riemers, F.M.; Bergknut, N.; Wubbolts, R.; Grinwis, G.C.; Houweling, M.; Groot Koerkamp, M.J.; van Leenen, D.; et al. Gene expression profiling of early intervertebral disc degeneration reveals a down-regulation of canonical Wnt signaling and caveolin-1 expression: Implications for development of regenerative strategies. Arthr. Res. Ther. 2013, 15, R23. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tian, Y.; Wang, J.; Phillips, K.L.; Binch, A.L.; Dunn, S.; Cross, A.; Chiverton, N.; Zheng, Z.; Shapiro, I.M.; et al. Inflammatory cytokines induce NOTCH signaling in nucleus pulposus cells: Implications in intervertebral disc degeneration. J. Biol. Chem. 2013, 288, 16761–16774. [Google Scholar] [CrossRef] [PubMed]
- Kazezian, Z.; Gawri, R.; Haglund, L.; Ouellet, J.; Mwale, F.; Tarrant, F.; O’Gaora, P.; Pandit, A.; Alini, M.; Grad, S. Gene expression profiling identifies interferon signalling molecules and IGFBp3 in human degenerative annulus fibrosus. Sci. Rep. 2015, 5, 15662. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, J.V.; Gantenbein-Ritter, B.; Bertolo, A.; Aebli, N.; Baur, M.; Alini, M.; Grad, S. Role of hypoxia and growth and differentiation factor-5 on differentiation of human mesenchymal stem cells towards intervertebral nucleus pulposus-like cells. Eur. Cell. Mater. 2011, 21, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Nisar, M.; Huang, C.; Pan, X.; Lin, D.; Zheng, G.; Jin, H.; Chen, D.; Tian, N.; Huang, Q.; et al. Small molecule natural compound agonist of SIRT3 as a therapeutic target for the treatment of intervertebral disc degeneration. Exp. Mol. Med. 2018, 50, 146. [Google Scholar] [CrossRef] [PubMed]
- Wuertz, K.; Vo, N.; Kletsas, D.; Boos, N. Inflammatory and catabolic signalling in intervertebral discs: The roles of NF-κB and MAP kinases. Eur. Cell. Mater. 2012, 23, 103–119. [Google Scholar] [CrossRef]
- Vo, N.V.; Hartman, R.A.; Patil, P.R.; Risbud, M.V.; Kletsas, D.; Iatridis, J.C.; Hoyland, J.A.; Le Maitre, C.L.; Sowa, G.A.; Kang, J.D. Molecular mechanisms of biological aging in intervertebral discs. J. Orthop. Res. 2016, 34, 1289–1306. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Liu, H.; Yang, M.; Zhang, Y.; Huang, B.; Zhou, Y. Disc cell senescence in intervertebral disc degeneration: Causes and molecular pathways. Cell Cycle 2016, 15, 1674–1684. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhou, Y.; Chu, T.W.; Li, C.Q.; Wang, J.; Zhang, Z.F.; Huang, B. Distinguishing characteristics of stem cells derived from different anatomical regions of human degenerated intervertebral discs. Eur. Spine J. 2016, 25, 2691–2704. [Google Scholar] [CrossRef]
- Morishita, Y.; Buser, Z.; D’Oro, A.; Shiba, K.; Wang, J.C. Clinical relationship of degenerative changes between the cervical and lumbar spine. Asian Spine J. 2018, 12, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Weiler, C.; Schietzsch, M.; Kirchner, T.; Nerlich, A.G.; Boos, N.; Wuertz, K. Age-related changes in human cervical, thoracal and lumbar intervertebral disc exhibit a strong intra-individual correlation. Eur. Spine J. 2012, 21, S810–S818. [Google Scholar] [CrossRef] [PubMed]
- Okada, E.; Matsumoto, M.; Fujiwara, H.; Toyama, Y. Disc degeneration of cervical spine on MRI in patients with lumbar disc herniation: Comparison study with asymptomatic volunteers. Eur. Spine J. 2011, 20, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Sakai, D.; Grad, S. Advancing the cellular and molecular therapy for intervertebral disc disease. Adv. Drug Deliv. Rev. 2015, 84, 159–171. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor | IVD Level | Age | Sex | Symptoms | Duration of Symptoms Prior to Surgery | Pfirrmann Grade |
|---|---|---|---|---|---|---|
| #1 | L4L5 | 51 | female | radiculopathy: pain and palsy, neurogenic claudication | 5 months | III |
| #2 | L5S1 | 57 | male | radiculopathy: palsy | 5 months | IV |
| #3 | L2L3 | 56 | male | radiculopathy: pain and palsy | 2 months | II |
| #4 | L4L5 | 49 | female | radiculopathy: pain | 2 months | II |
| #5 | L4L5 | 52 | male | radiculopathy: pain and palsy | 1 month | III |
| #6 | L4L5 | 79 | male | radiculopathy: pain and palsy | 5 months | IV |
| #7 | L4L5 | 47 | female | radiculopathy: pain | 7 months | II |
| #8 | L4L5 | 54 | female | back pain and claudicatio | 5 months | IV |
| #9 | L5S1 | 44 | male | radiculopathy: pain | 2 months | II |
| #10 | L5S1 | 63 | female | back pain and radiculopathy: pain and palsy | 12 months | V |
| #11 | L4L5 | 57 | female | claudicatio | 24 months | V |
| #12 | L4L5 | 70 | male | radiculopathy: pain and palsy | 1 month | IV |
| #13 | L4L5 | 40 | female | radiculopathy: pain | 5 months | II |
| #14 | L5S1 | 38 | male | back pain and radiculopathy: pain and palsy | 12 months | IV |
| #15 | L4L5 | 74 | female | radiculopathy: pain and neurogenic claudication | 12 months | V |
| #16 | L4L5 | 70 | male | radiculopathy: pain and palsy | 2 months | IV |
| #17 | L3L4 | 56 | male | radiculopathy: pain | 3 months | I |
| #18 | L4L5 | 46 | male | radiculopathy: pain and palsy | 12 months | III |
| #19 | L5S1 | 47 | female | radiculopathy: pain | 2 months | IV |
| #20 | L5S1 | 51 | male | radiculopathy: pain | 6 months | IV |
| #21 | L4L5 | 51 | male | radiculopathy: pain | 3 months | III |
| #22 | L5S1 | 63 | female | radiculopathy: pain and palsy | 1 month | III |
| #23 | L4L5 | 37 | male | radiculopathy: pain and palsy | 2 months | II |
| #24 | L5S1 | 49 | male | radiculopathy: pain and palsy | 9 months | III |
| #25 | L5S1 | 44 | male | radiculopathy: pain | 4 months | III |
| #26 | L4L5 | 54 | male | radiculopathy: pain | 2 months | III |
| #27 | L4L5 | 64 | male | radiculopathy: pain and palsy | 3 months | IV |
| #28 | L4L5 | 56 | female | radiculopathy: pain and neurogenic claudication | 3 months | IV |
| #29 | L4L5 | 77 | male | radiculopathy: pain and palsy, neurogenic claudication | 10 months | III |
| #30 | L4L5 | 72 | female | radiculopathy: pain and palsy | 12 months | II |
| Region | Location | Primer Sequences |
|---|---|---|
| Region 1 | +572/+727 | F-GGTATCATTTGGATAGATCAAT R-TGGATGGAAGGAAGGTCGGATAGATAA |
| Region 2 | −1217/−1058 | F-AGCCACTTTTCTCTTGGTGAT R-CGTCTTAGAATCCTTTGCTGTG |
| Region 3 | −2419/−2234 | F-CCTGTTCTAACCGTGTGGAGT R-CCATACATTCTGGCTAAAGACC |
| Region 4 | −3429/−3140 | F-GGAATCCAAGTTCATAAGAACA R-ATGGTGATGGTATCACAGGTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penolazzi, L.; Lambertini, E.; Scussel Bergamin, L.; Gandini, C.; Musio, A.; De Bonis, P.; Cavallo, M.; Piva, R. Reciprocal Regulation of TRPS1 and miR-221 in Intervertebral Disc Cells. Cells 2019, 8, 1170. https://doi.org/10.3390/cells8101170
Penolazzi L, Lambertini E, Scussel Bergamin L, Gandini C, Musio A, De Bonis P, Cavallo M, Piva R. Reciprocal Regulation of TRPS1 and miR-221 in Intervertebral Disc Cells. Cells. 2019; 8(10):1170. https://doi.org/10.3390/cells8101170
Chicago/Turabian StylePenolazzi, Letizia, Elisabetta Lambertini, Leticia Scussel Bergamin, Carlotta Gandini, Antonio Musio, Pasquale De Bonis, Michele Cavallo, and Roberta Piva. 2019. "Reciprocal Regulation of TRPS1 and miR-221 in Intervertebral Disc Cells" Cells 8, no. 10: 1170. https://doi.org/10.3390/cells8101170