Effect of a Collagen-Based Compound on Morpho-Functional Properties of Cultured Human Tenocytes

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Cell Cultures
2.3. Coating with MD-Tissue
2.4. Growth Curves
2.5. Immunofluorescence Analysis
2.6. Real-Time PCR
2.7. Slot Blot
2.8. SDS-Zymography
2.9. Wound Healing Assay
2.10. Statistical Analysis
3. Results
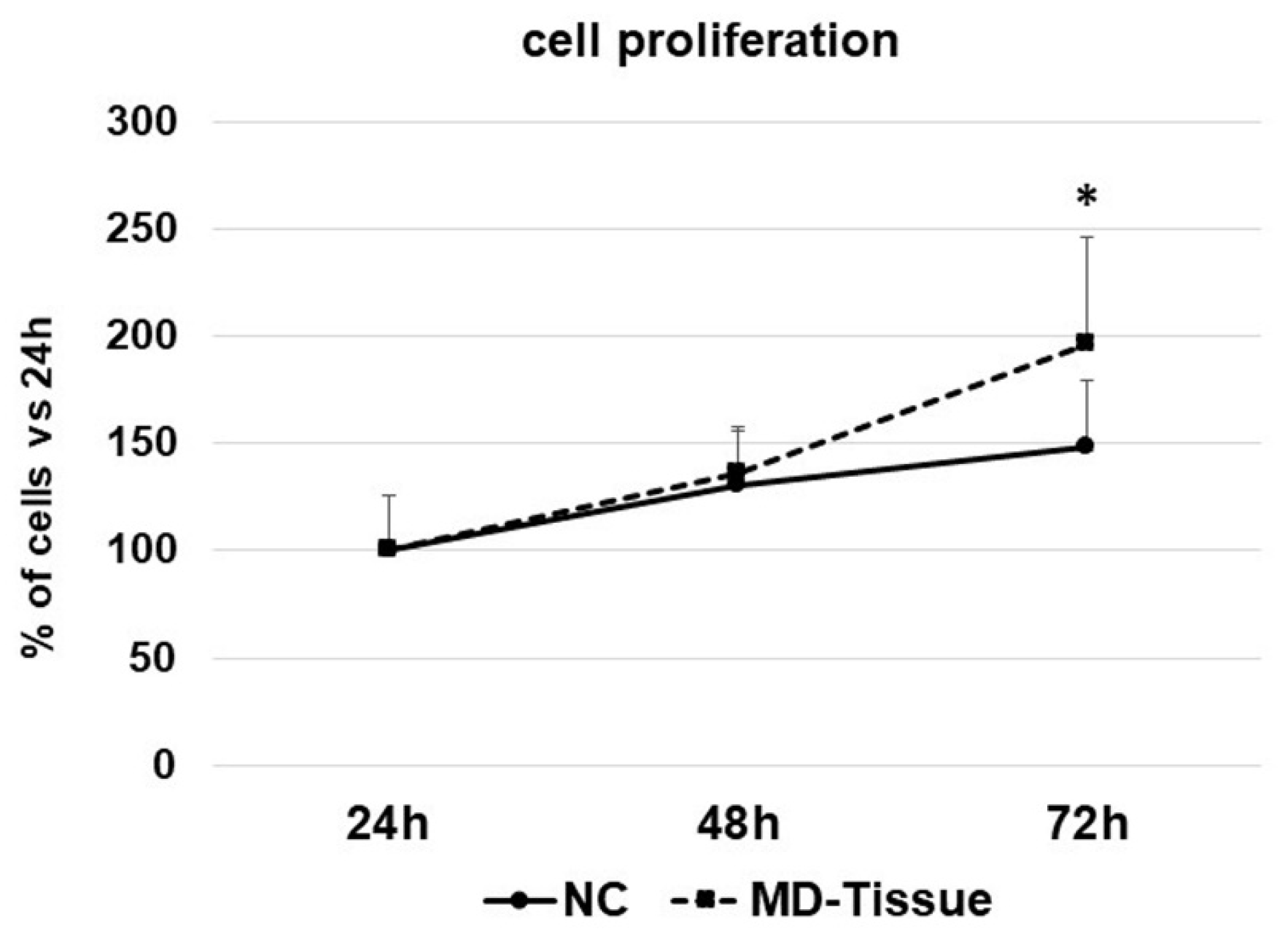
3.1. Cell Growth
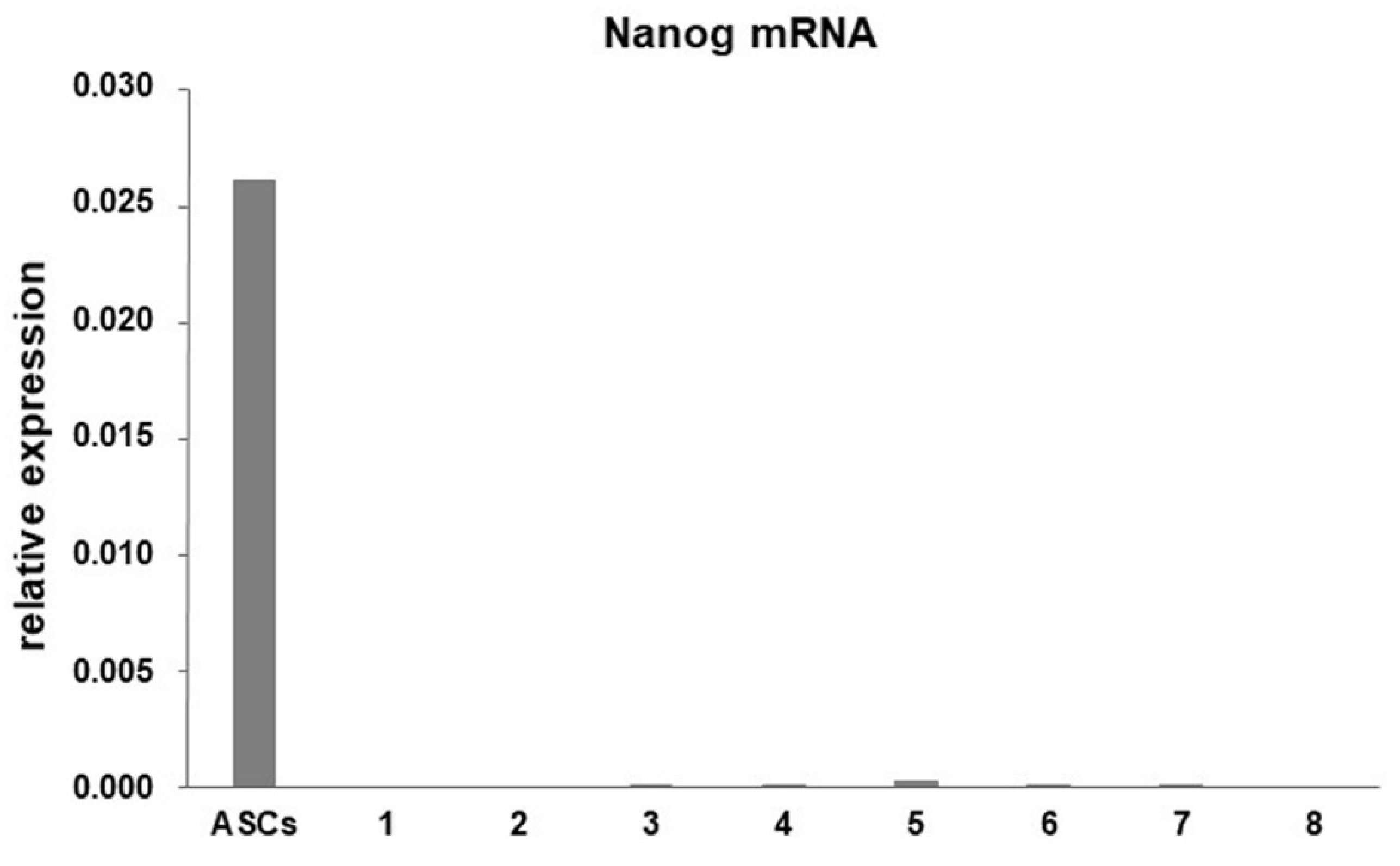
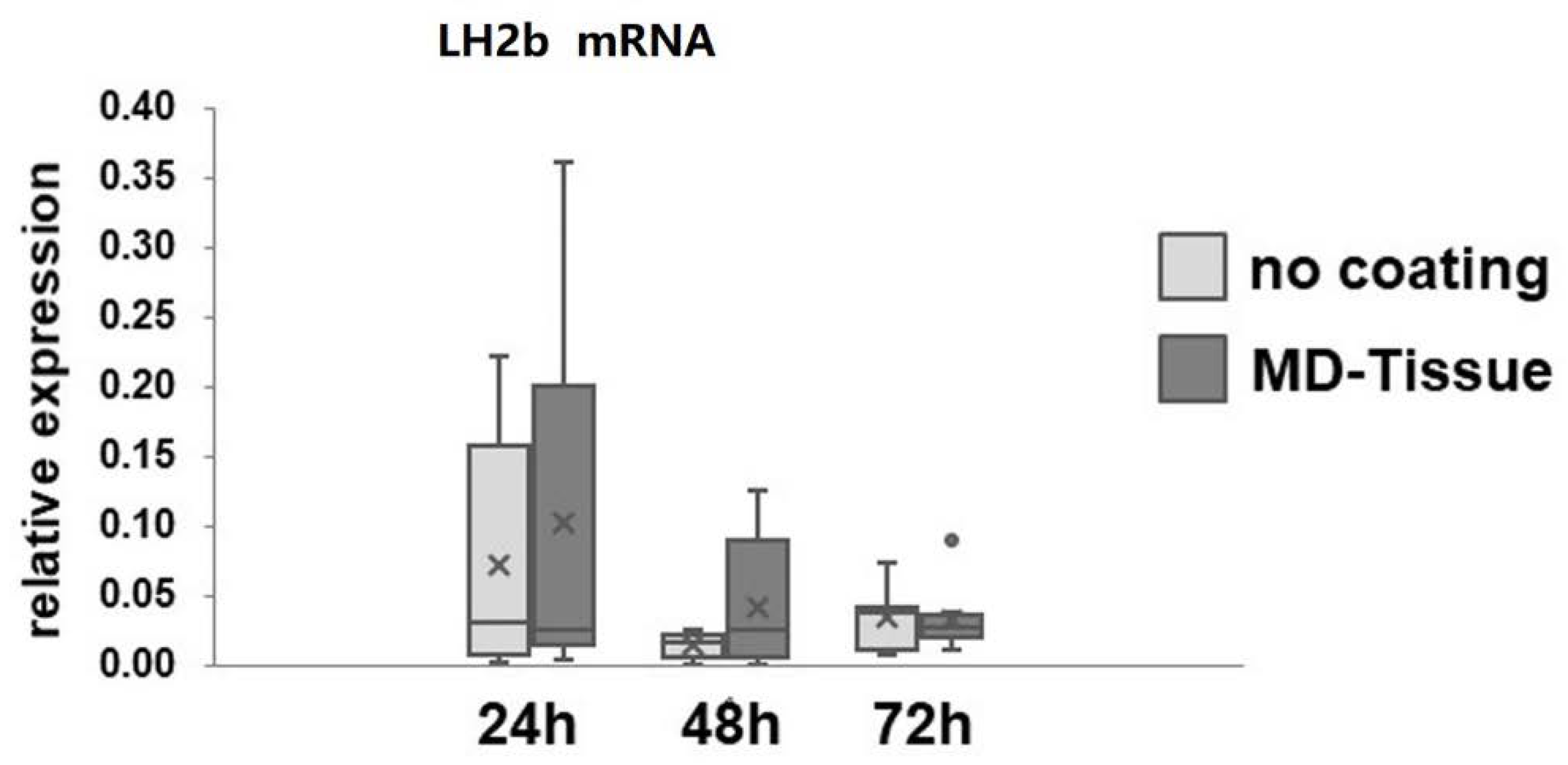
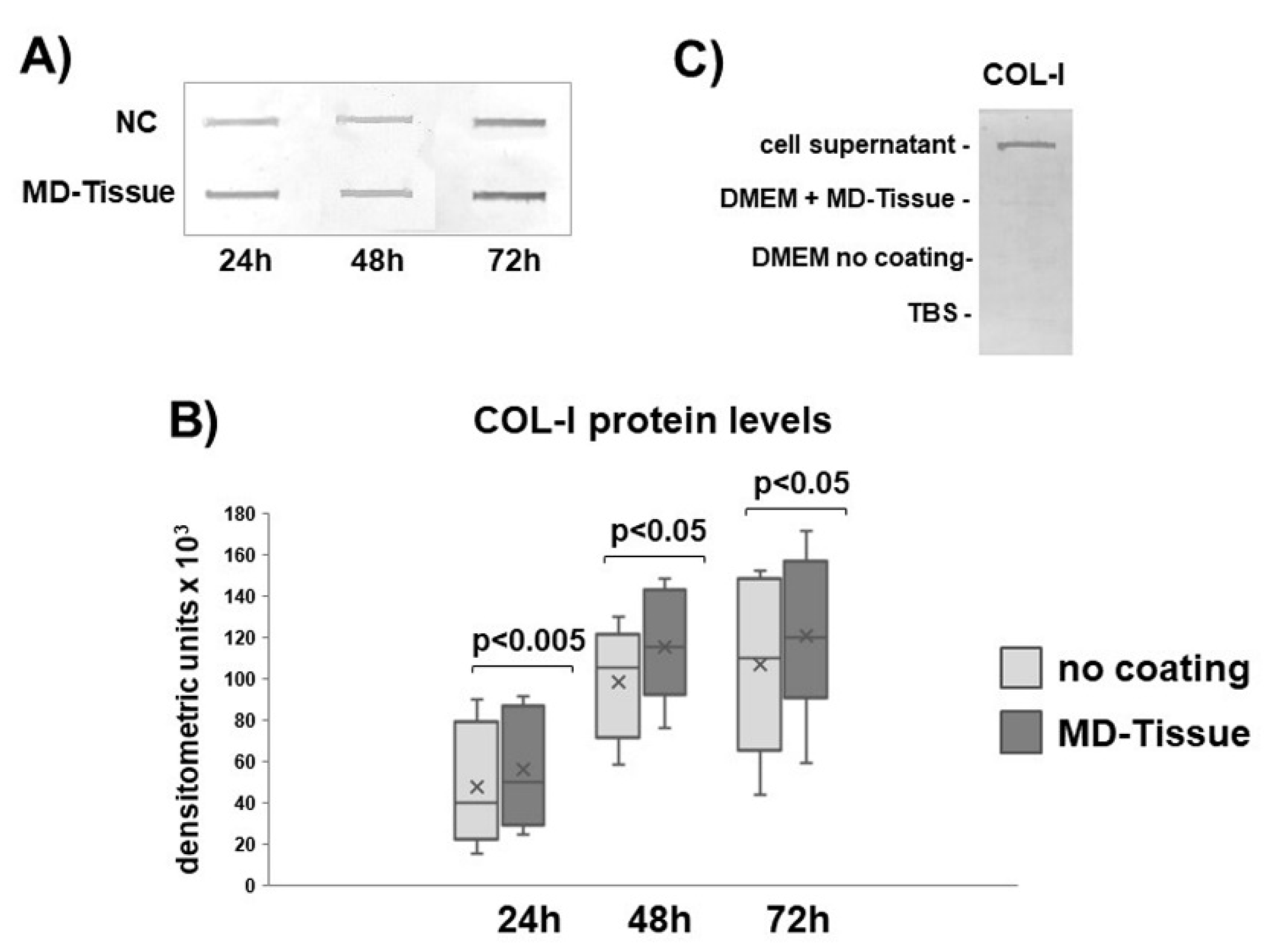
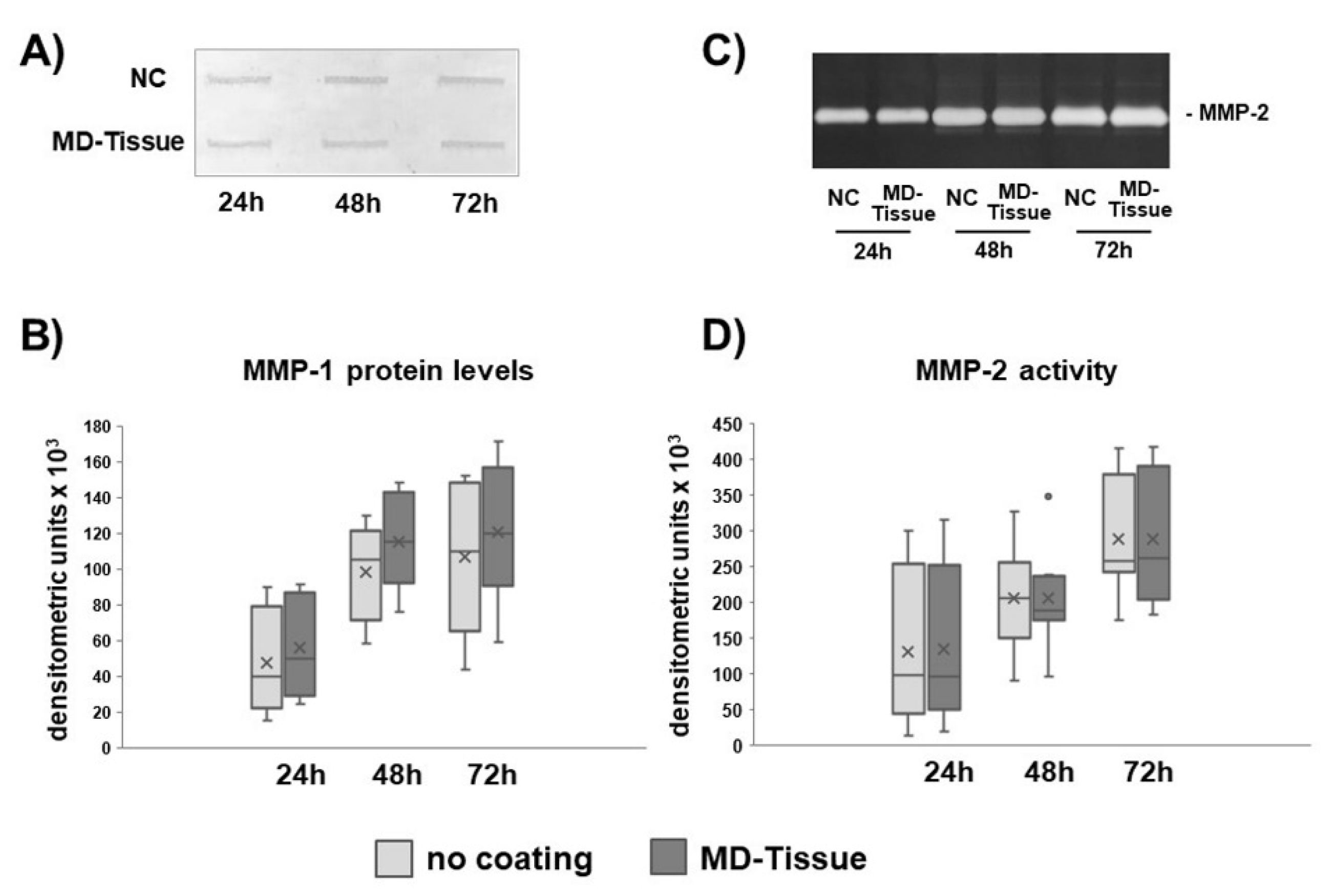
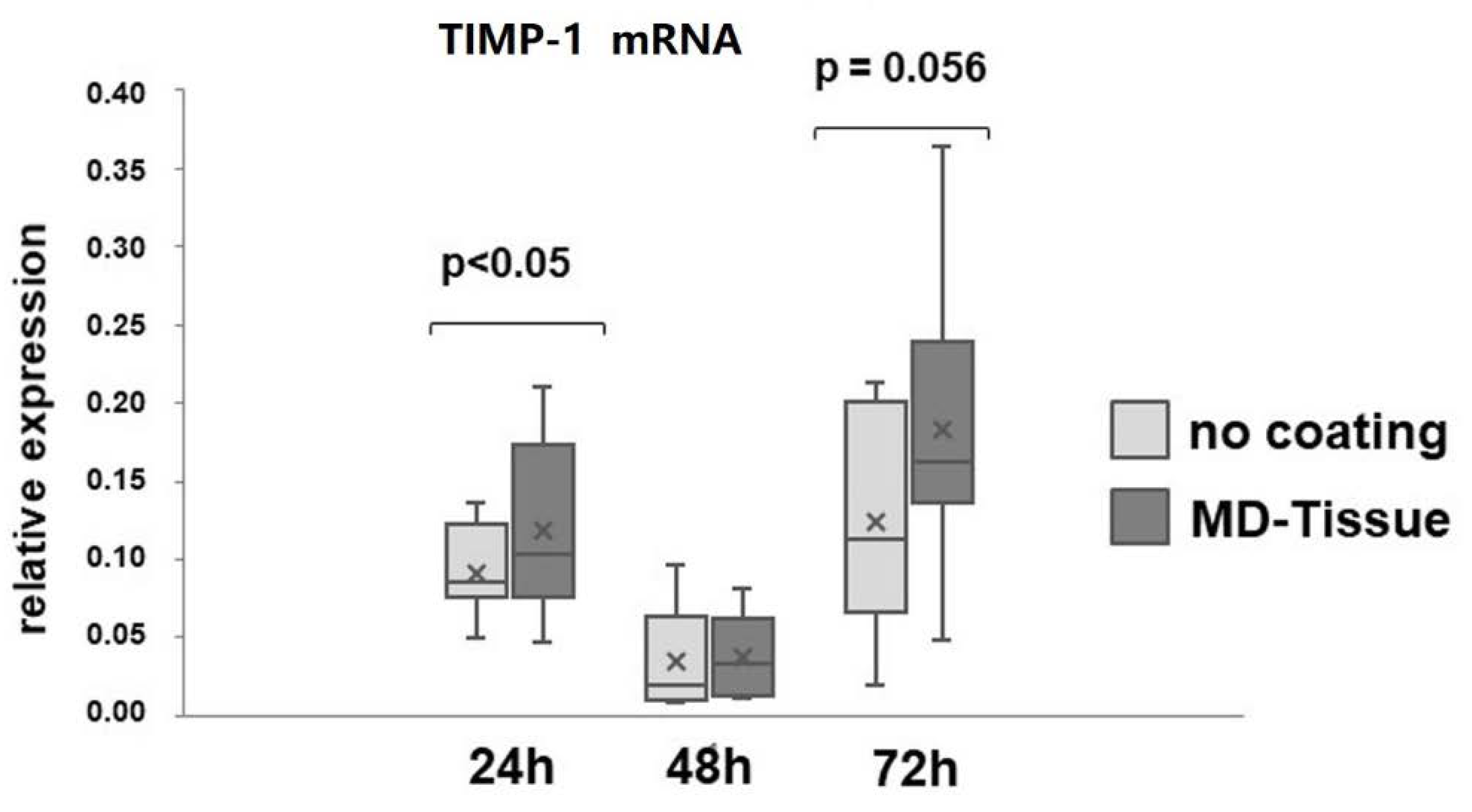
3.2. Expression of Genes and Proteins Related to Collagen Turnover
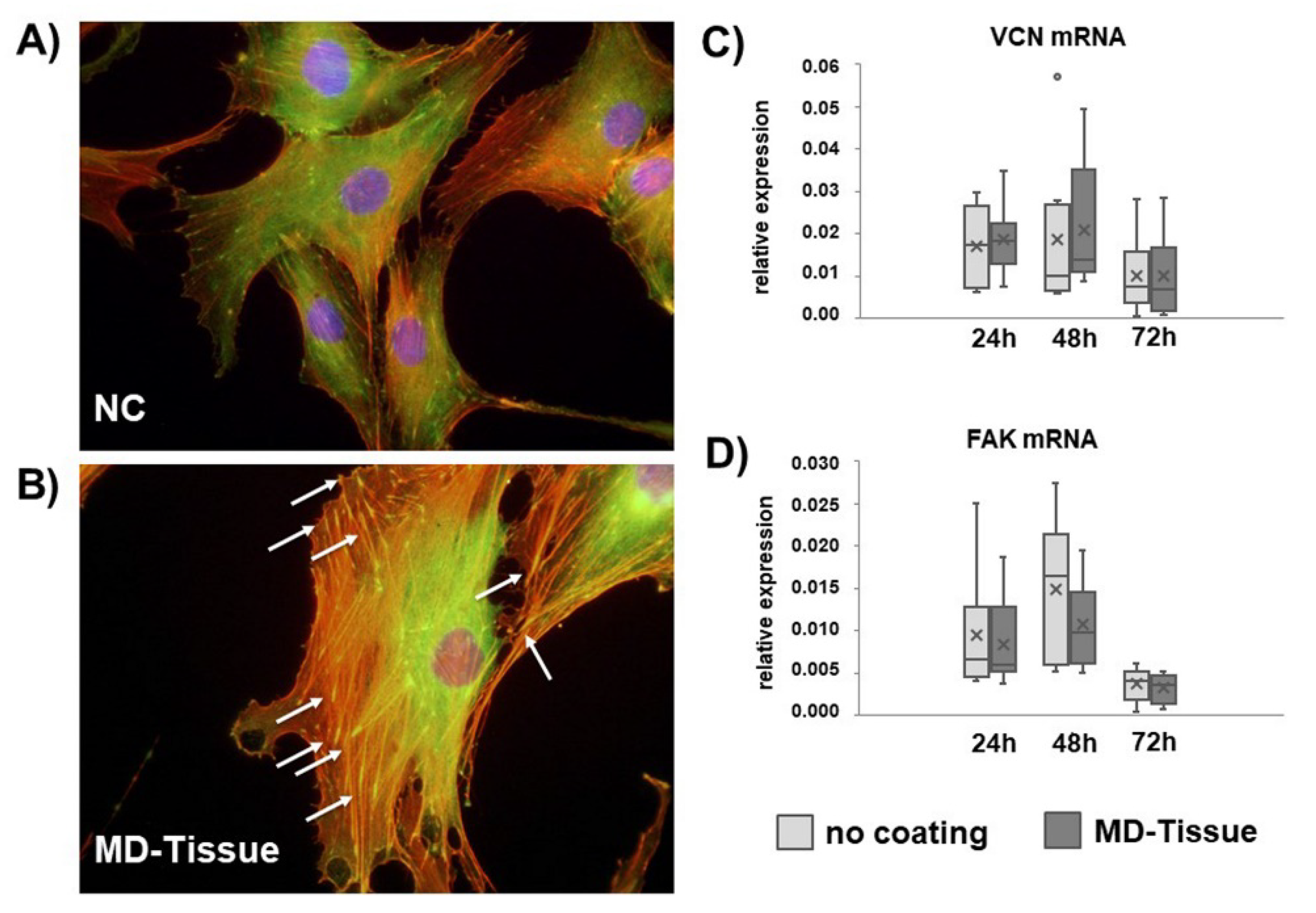
3.3. Cytoskeleton Arrangement and Focal Adhesion
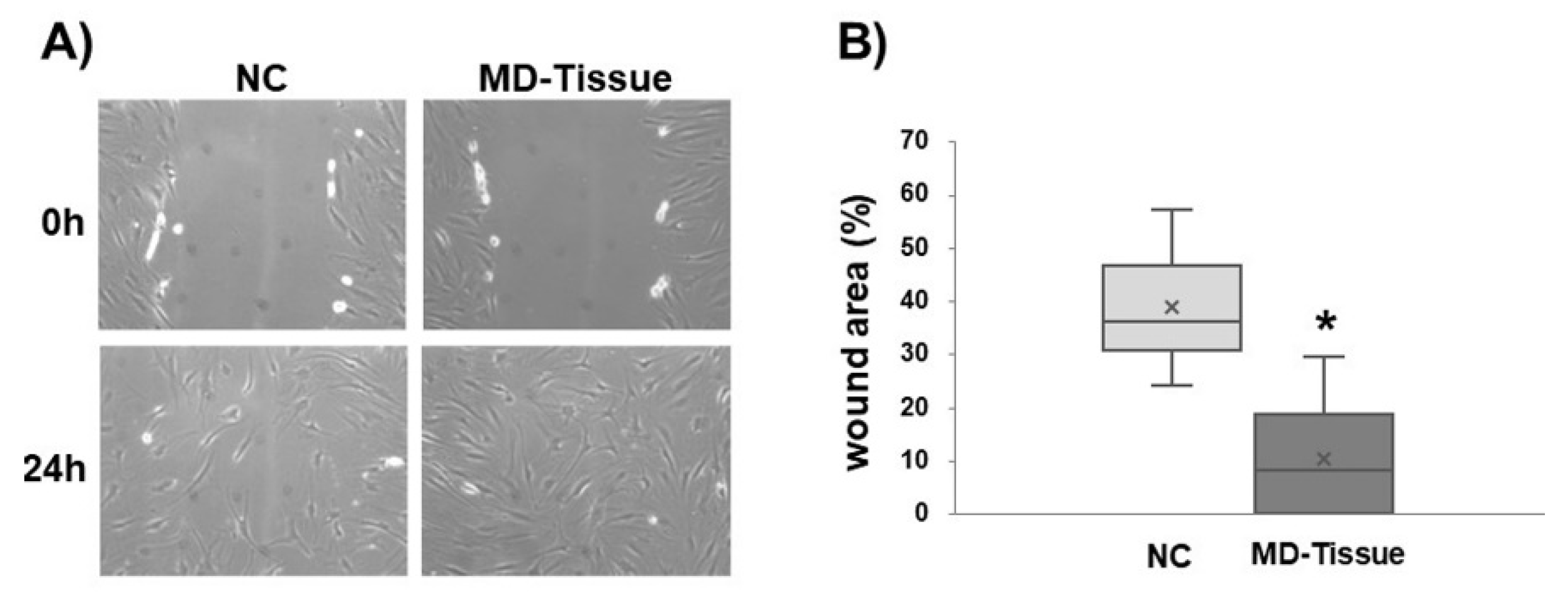
3.4. Wound Healing Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giai Via, A.; Papa, G.; Oliva, F.; Maffulli, N. Tendinopathy. Curr. Phys. Med. Rehabil. Rep. 2016, 4, 50–55. [Google Scholar] [CrossRef]
- Ejnisman, L.; Safran, M.R. Biologics in hip preservation. Ann. Joint 2018, 3, 50. [Google Scholar] [CrossRef]
- Karpinski, M.R.; Piggott, H. Greater trochanteric pain syndrome. A. report of 15 cases. J. Bone Joint Surg. Br. 1985, 67, 762–763. [Google Scholar] [CrossRef] [PubMed]
- Fearon, A.M.; Scarvell, J.M.; Neeman, T.; Cook, J.L.; Cormick, W.; Smith, P.N. Greater trochanteric pain syndrome: Defining the clinical syndrome. Br. J. Sports Med. 2013, 47, 649–653. [Google Scholar] [CrossRef]
- Frizziero, A.; Vittadini, F.; Pignataro, A.; Gasparre, G.; Biz, C.; Ruggieri, P.; Masiero, S. Conservative management of tendinopathies around hip. Muscles, Ligaments Tendons J. 2016, 6, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Reid, D. The management of greater trochanteric pain syndrome: A systematic literature review. J. Orthop. 2016, 13, 15–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimaldi, A.; Mellor, R.; Hodges, P.; Bennell, K.; Wajswelner, H.; Vicenzino, B. Gluteal tendinopathy: A review of mechanisms, assessment and management. Sports Med. 2015, 45, 1107–1119. [Google Scholar] [CrossRef]
- Seo, K.H.; Lee, J.Y.; Yoon, K.; Do, J.G.; Park, H.J.; Lee, S.Y.; Park, Y.S.; Lee, Y.T. Long-term outcome of low-energy extracorporeal shockwave therapy on gluteal tendinopathy documented by magnetic resonance imaging. PLoS ONE 2018, 13, e019746. [Google Scholar] [CrossRef]
- Del Buono, A.; Papalia, R.; Khanduja, V.; Denaro, V.; Maffulli, N. Management of the greater trochanteric pain syndrome: A. systematic review. Br. Med. Bull. 2012, 102, 115–131. [Google Scholar] [CrossRef]
- Fitzpatrick, J.; Bulsara, M.K.; O’Donnell, J.; McCrory, P.R.; Zheng, M.H. The effectiveness of platelet-rich plasma injections in gluteal tendinopathy a randomized, double-blind controlled trial comparing a single platelet-rich plasma injection with a single corticosteroid injection. Am. J. Sports Med. 2018, 46, 933–939. [Google Scholar] [CrossRef]
- Thomas, A.; Bucher, T.A.; Ebert, J.R.; Smith, A.; Breidahl, W.; Fallon, M.; Wang, T.; Zheng, M.H.; Janes, G.C. Autologous tenocyte injection for the treatment of chronic recalcitrant gluteal tendinopathy a prospective pilot study. Orthopaedic J. Sports Med. 2017, 5, 1–10. [Google Scholar] [CrossRef]
- Andia, I.; Maffulli, N. How far have biological therapies come in regenerative sports medicine? Expert Opin. Biol. Ther. 2018, 18, 785–793. [Google Scholar] [CrossRef]
- Kannus, P. Structure of the tendon connective tissue. Scand J. Med. Sci Sports 2000, 10, 312–320. [Google Scholar] [CrossRef]
- Kjaer, M. Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol. Rev. 2004, 84, 649–698. [Google Scholar] [CrossRef] [PubMed]
- Riley, G.P.; Harrall, R.L.; Constant, C.R.; Chard, M.D.; Cawston, T.E.; Hazleman, B.L. Glycosaminoglycans of human rotator cuff tendons: Changes with age and in chronic rotator cuff tendinitis. Ann. Rheum. Dis. 1994, 53, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Banes, A.J.; Horesovsky, G.; Larson, C.; Tsuzaki, M.; Judex, S.; Archambault, J.; Zernicke, R.; Herzog, W.; Kelley, S.; Miller, L. Mechanical load stimulates expression of novel genes in vivo and in vitro in avian flexor tendon cells. Osteoarthritis Cartilage 1999, 7, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, M.; Ralphs, J.R. The cell and developmental biology of tendons and ligaments. Int. Rev. Cytol. 2000, 196, 85–130. [Google Scholar]
- Martin, L.S.; Massafra, U.; Bizzi, E.; Migliore, A. A double blind randomized active-controlled clinical trial on the intra-articular use of Md-Knee versus sodium hyaluronate in patients with knee osteoarthritis (“Joint”). BMC Musculoskelet Disord 2016, 17, 94. [Google Scholar] [CrossRef]
- Pavelka, K.; Jarosova, H.; Sleglova, O.; Svobodova, R.; Votavova, M.; Milani, L.; Prochazka, Z.; Kotlarova, L.; Kostiuk, P.; Sliva, J.; et al. Chronic Low Back Pain: Current Pharmacotherapeutic Therapies and a New Biological Approach. Curr. Med. Chem. 2018, 25, 1–8. [Google Scholar] [CrossRef]
- Ritty, T.M.; Herzog, J. Tendon cells produce gelatinases in response to type I collagen attachment. J. Orthop. Res. 2003, 21, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Menon, A.; Pettinari, L.; Martinelli, C.; Colombo, G.; Portinaro, N.; Dalle-Donne, I.; d’Agostino, M.C.; Gagliano, N. New insights in extracellular matrix remodeling and collagen turnover related pathways in cultured human tenocytes after ciprofloxacin administration. Muscles Ligaments Tendons J. 2013, 113, 122–131. [Google Scholar]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Viganò, M.; Perucca Orfei, C.; de Girolamo, L.; Pearson, J.R.; Ragni, E.; De Luca, P.; Colombini, A. Housekeeping Gene Stability in Human Mesenchymal Stem and Tendon Cells Exposed to Tenogenic Factors. Tissue Eng. Part. C Methods 2018, 24, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Calloni, R.; Cordero, E.A.; Henriques, J.A.; Bonatto, D. Reviewing and updating the major molecular markers for stem cells. Stem Cells Dev. 2013, 22, 1455–1476. [Google Scholar] [CrossRef] [PubMed]
- Viidik, A. The effect of training on the tensile strength of isolated rabbit tendons. Scand J. Plast Reconstr. Surg. 1967, 1, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Ritter, M.A.; Amiel, D.; Sanders, T.M.; Gomez, M.A.; Kuei, S.C.; Garfin, S.R.; Akeson, W.H. The biomechanical and biochemical properties of swine tendons--long term effects of exercise on the digital extensors. Connect. Tissue Res. 1980, 7, 177–183. [Google Scholar] [CrossRef]
- Woo, S.L.; Gomez, M.A.; Amiel, D.; Ritter, M.A.; Gelberman, R.H.; Akeson, W.H. The effects of exercise on the biomechanical and biochemical properties of swine digital flexor tendons. J. Biomech. Eng. 1981, 103, 51–56. [Google Scholar] [CrossRef]
- Langberg, H.; Rosendal, L.; Kjaer, M. Training-induced changes in peritendinous type I collagen turnover determined by microdialysis in humans. J. Physiol. 2001, 534, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Silver, F.H.; Christiansen, D.; Snowhill, P.B.; Chen, Y.; Landis, W.J. The role of mineral in the storage of elastic energy in turkey tendons. Biomacromolecules 2000, 1, 180–185. [Google Scholar] [CrossRef]
- Walker, L.C.; Overstreet, M.A.; Yeowell, H.N. Tissue-specific expression and regulation of the alternatively-spliced forms of lysyl hydroxylase 2 (LH2) in human kidney cells and skin fibroblasts. Matrix. Biol. 2005, 23, 515–523. [Google Scholar] [CrossRef]
- Sakai, T.; Gross, J. Some properties of the products of reaction of tadpole collagenase with collagen. Biochemistry 1967, 6, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Woessner, F.J. Matrix metalloproteinases and their inhibitors in connective tissue remodelling. FASEB J. 1991, 5, 2145–2154. [Google Scholar] [CrossRef] [PubMed]
- Arnoczky, S.P.; Tian, T.; Lavagnino, M.; Gardner, K. Ex vivo static tensile loading inhibits MMP-1 expression in rat tail tendon cells through a cytoskeletally based mechanotransduction mechanism. J. Orthop. Res. 2004, 22, 328–333. [Google Scholar] [CrossRef] [Green Version]
- Brew, K.; Dinakarpandian, D.; Nagase, H. Tissue inhibitors of metalloproteinases: Evolution, structure and function. Biochim. Biophys. Acta 2001, 1477, 267–283. [Google Scholar] [CrossRef]
- Murphy, G.; Willenbrock, F.; Crabbe, T.; O’Shea, M.; Ward, R.; Atkinson, S.; O’Connell, J.; Docherty, A. Regulation of matrix metalloproteinase activity. Ann. N Y Acad. Sci. 1994, 732, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Phipps, R.P.; Borrello, M.A.; Blieden, T.M. Fibroblast heterogeneity in the periodontium and other tissues. J. Periodontal Res. 1997, 32, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Archana, A.; Srikanth, V.; Sasireka; Kurien, B.; Ebenezer. Fibroblast Heterogeneity in Periodontium. Int. J. Dental Sci. Res. 2014, 2, 50–54. [Google Scholar] [CrossRef]
- Burridge, K.; Guilluy, C. Focal adhesions, stress fibers and mechanical tension. Exp. Cell. Res. 2016, 343, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.H.; Guo, Q.; Li, B. Tendon Biomechanics and Mechanobiology-A Minireview of Basic Concepts and Recent Advancements. J. Hand Ther. 2012, 25, 133–140. [Google Scholar] [CrossRef]
- Docking, S.; Samiric, T.; Scase, E.; Purdam, C.; Cook, J. Relationship between compressive loading and ECM changes in tendons. Muscle Ligaments Tendons J. 2013, 3, 7–11. [Google Scholar] [CrossRef]
- Heinemeier, K.M.; Olesen, J.L.; Haddad, F.; Schjerling, P.; Baldwin, K.M.; Kjaer, M. Effect of unloading followed by reloading on expression of collagen and related growth factors in rat tendon and muscle. J. Appl. Physiol. 2009, 106, 178–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, M.J.; Ferara, N.S.; Vernon, R.B. Impaired migration, integrin function, and actin cytoskeletal organization in dermal fibroblasts from a subset of aged human donors. Mech. Ageing Dev. 2001, 122, 1203–1220. [Google Scholar] [CrossRef]
- Jones, M.E.; Mudera, V.; Brown, R.A.; Cambrey, A.D.; Grobbelaar, A.O.; McGrouther, D.A. The early surface cell response to flexor tendon injury. J. Hand Surg. Am. 2003, 28, 221–230. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
| Age ranging 18–70 years. | Patients diagnosed of great trochanter tendinopathy |
| Indication for total hip arthroplasty. | Patients affected by genetic collagen disorders. |
| Patients who signed written informed consent for the surgery. | Patients diagnosed of spondyloarthritis with involvement of the affected hip. |
| Patients able to understand the study conditions and willing to participate for its entire duration. | Patients affected by psoriatic arthritis. |
| Drug-addicted Alcohol-addicted Psychiatric disorders Clinical conditions which could compromise the results of the surgical procedure or of the follow-up. | |
| Informed consent not accepted. | |
| Pregnant or breastfeeding women. | |
| Patients affected by diabetes mellitus. | |
| Patients who had taken fluoroquinolones within 30 days before the surgery. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Randelli, F.; Menon, A.; Giai Via, A.; Mazzoleni, M.G.; Sciancalepore, F.; Brioschi, M.; Gagliano, N. Effect of a Collagen-Based Compound on Morpho-Functional Properties of Cultured Human Tenocytes. Cells 2018, 7, 246. https://doi.org/10.3390/cells7120246
Randelli F, Menon A, Giai Via A, Mazzoleni MG, Sciancalepore F, Brioschi M, Gagliano N. Effect of a Collagen-Based Compound on Morpho-Functional Properties of Cultured Human Tenocytes. Cells. 2018; 7(12):246. https://doi.org/10.3390/cells7120246
Chicago/Turabian StyleRandelli, Filippo, Alessandra Menon, Alessio Giai Via, Manuel Giovanni Mazzoleni, Fabio Sciancalepore, Marco Brioschi, and Nicoletta Gagliano. 2018. "Effect of a Collagen-Based Compound on Morpho-Functional Properties of Cultured Human Tenocytes" Cells 7, no. 12: 246. https://doi.org/10.3390/cells7120246