BRCA1 and BRCA2 Gene Expression: Diurnal Variability and Influence of Shift Work

 ,
,  ,
, 
Abstract
:
1. Introduction
2. Results
2.1. Research in Healthy Volunteers
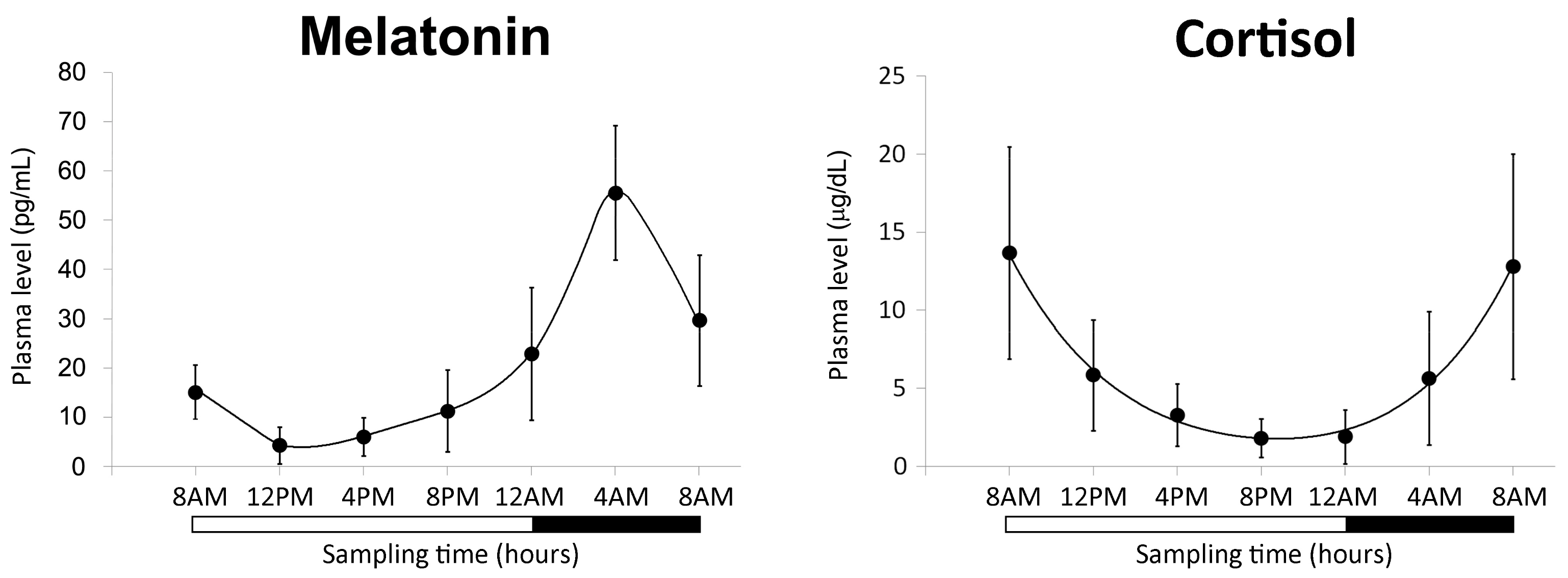
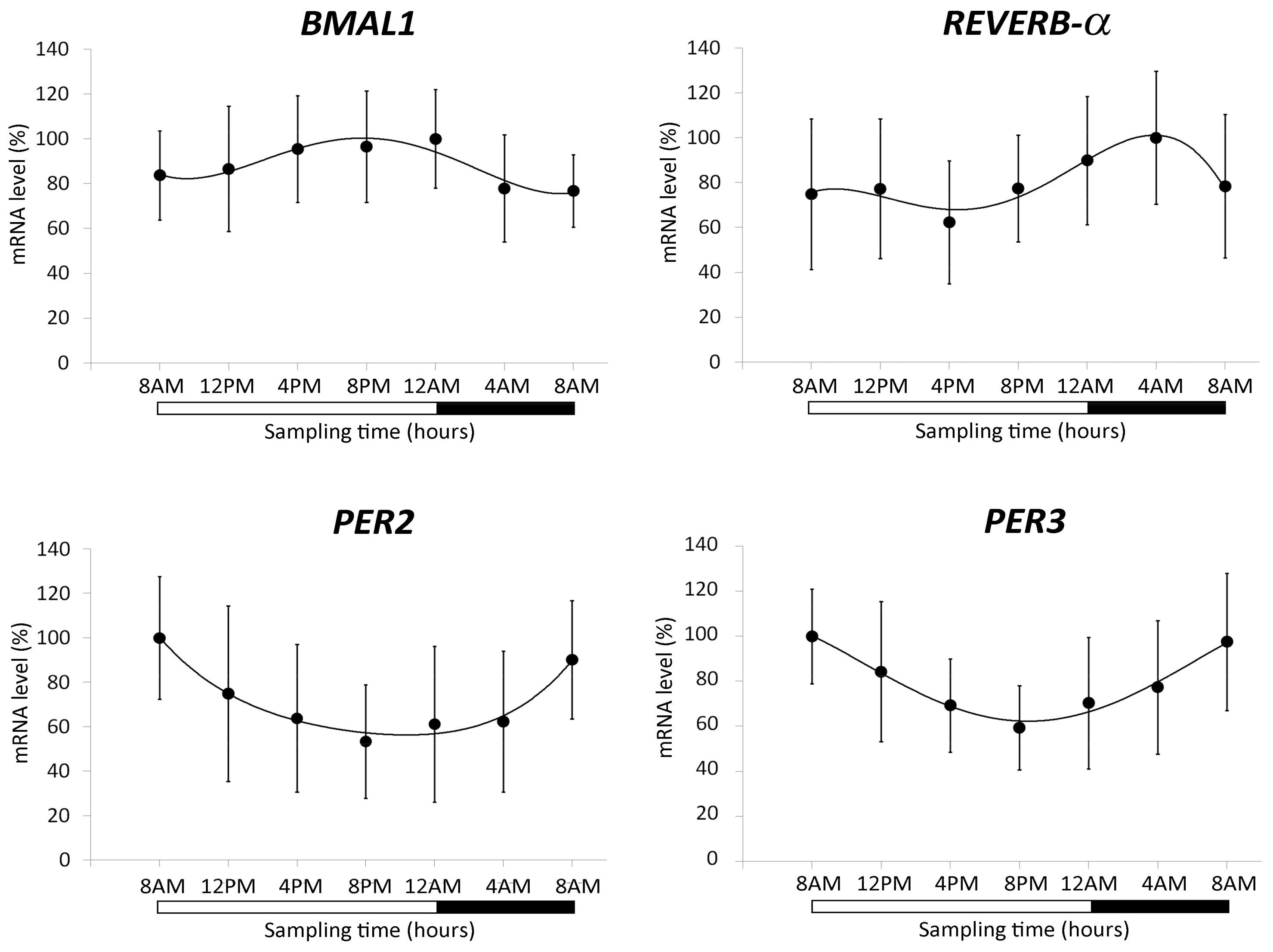
2.1.1. Characterization of the Biological Clock
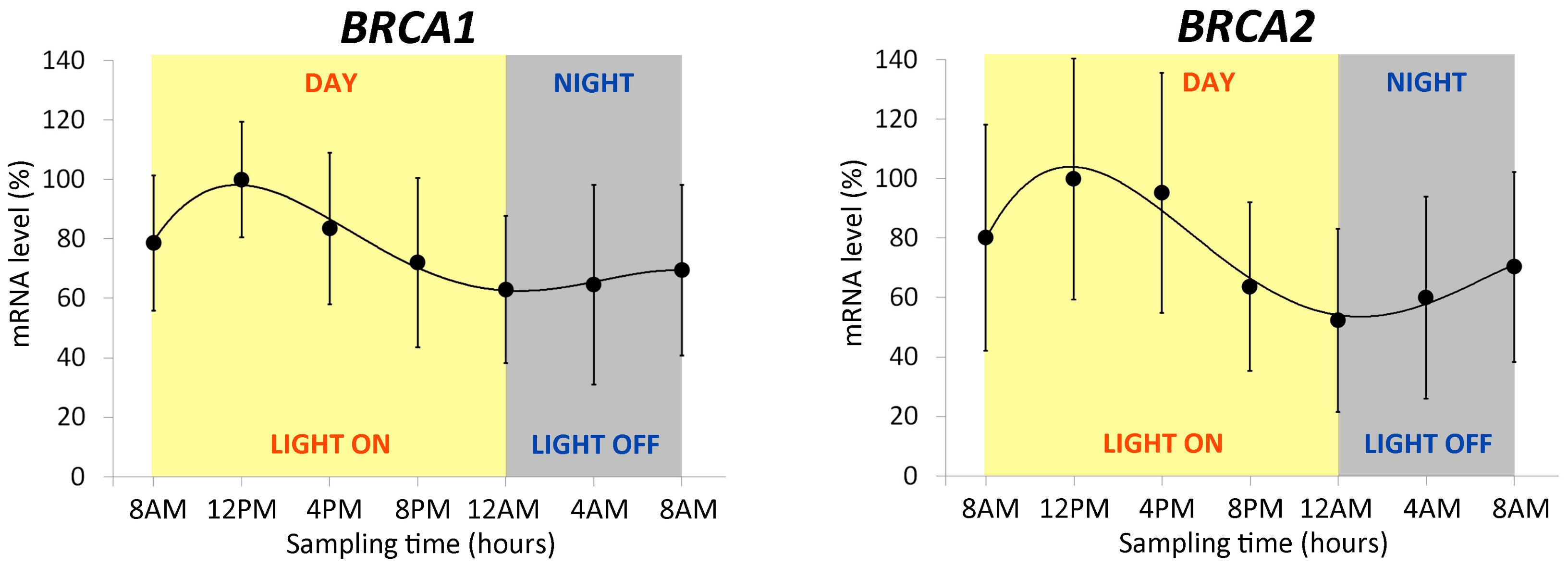
2.1.2. Expression Profile of BRCA1 and BRCA2 Genes
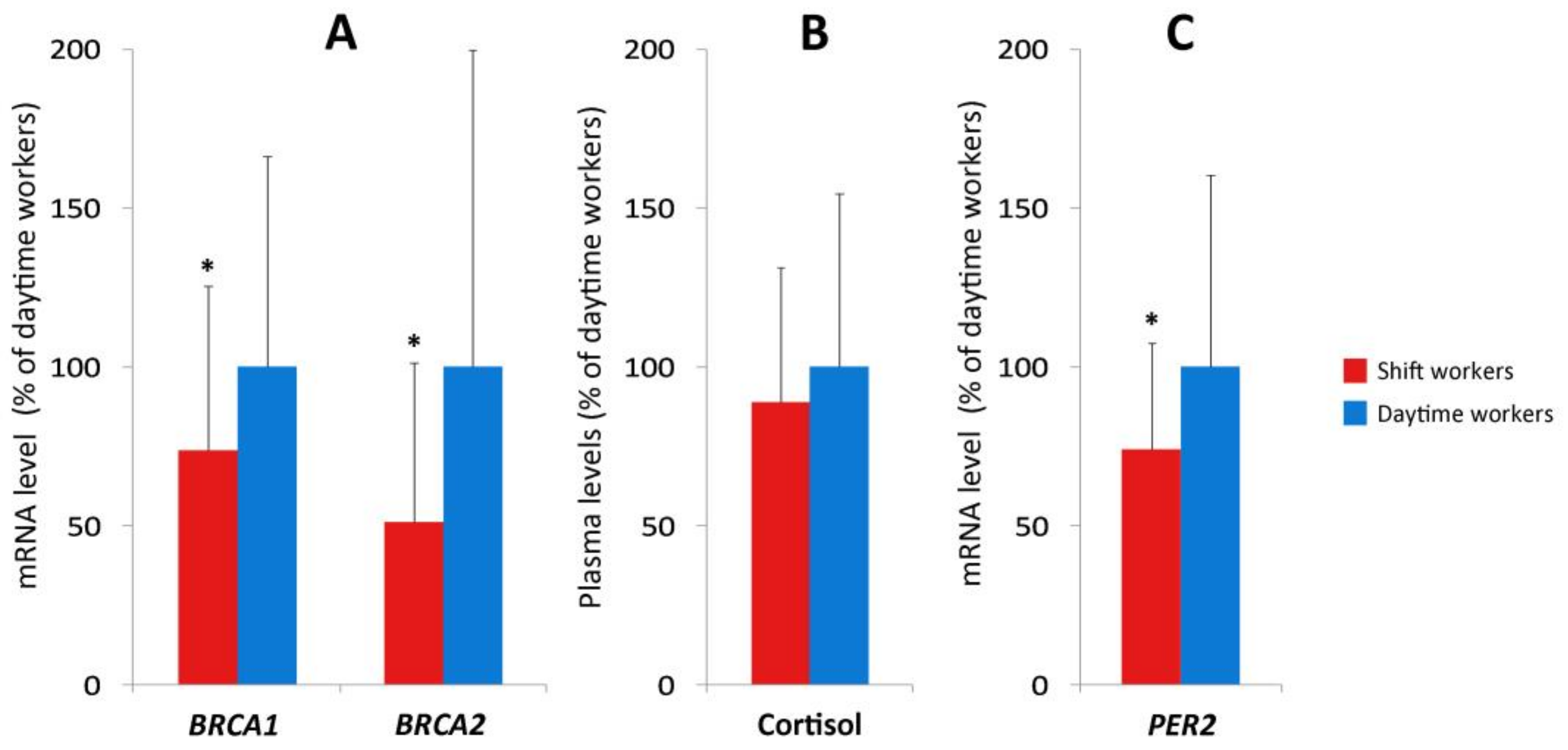
2.2. Research in Shift and Daytime Workers
3. Discussion
4. Materials and Methods
4.1. Participants and Sampling
4.1.1. Healthy Volunteers
4.1.2. Shift and Daytime Workers
4.2. Melatonin and Cortisol Assay
4.3. Gene Expression Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Antoch, M.P.; Kondratov, R.V.; Takahashi, J.S. Circadian clock genes as modulators of sensitivity to genotoxic stress. Cell Cycle 2005, 4, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Collis, S.J.; Boulton, S.J. Emerging links between the biological clock and the DNA damage response. Chromosoma 2007, 116, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Gaddameedhi, S.; Sancar, A. Melanoma and DNA damage from a distance (farstander effect). Pigment Cell Melanoma Res. 2011, 24, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A.; Lindsey-Boltz, L.A.; Kang, T.H.; Reardon, J.T.; Lee, J.H.; Ozturk, N. Circadian clock control of the cellular response to DNA damage. FEBS Lett. 2010, 584, 2618–2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaddameedhi, S.; Selby, C.P.; Kaufmann, W.K.; Smart, R.C.; Sancar, A. Control of skin cancer by the circadian rhythm. Proc. Natl. Acad. Sci. USA 2011, 108, 18790–18795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, T.H.; Reardon, J.T.; Kemp, M.; Sancar, A. Circadian oscillation of nucleotide excision repair in mammalian brain. Proc. Natl. Acad. Sci. USA 2009, 106, 2864–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppe, G.; Rayborn, M.E.; Sears, J.E. Diurnal rhythm of the chromatin protein hmgb1 in rat photoreceptors is under circadian regulation. J. Comp. Neurol. 2007, 501, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Manzella, N.; Bracci, M.; Strafella, E.; Staffolani, S.; Ciarapica, V.; Copertaro, A.; Rapisarda, V.; Ledda, C.; Amati, M.; Valentino, M.; et al. Circadian modulation of 8-oxoguanine DNA damage repair. Sci. Rep. 2015, 5, 13752. [Google Scholar] [CrossRef] [PubMed]
- Venkitaraman, A.R. Cancer susceptibility and the functions of BRCA1 and BRCA2. Cell 2002, 108, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.R.; Sossick, A.J.; Boulton, S.J.; Jackson, S.P. BRCA1-associated exclusion of 53bp1 from DNA damage sites underlies temporal control of DNA repair. J. Cell Sci. 2012, 125, 3529–3534. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, E.; Hochegger, H.; Saberi, A.; Taniguchi, Y.; Takeda, S. Differential usage of non-homologous end-joining and homologous recombination in double strand break repair. DNA Repair (Amst) 2006, 5, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Lavin, M.F. Atm and the mre11 complex combine to recognize and signal DNA double-strand breaks. Oncogene 2007, 26, 7749–7758. [Google Scholar] [CrossRef] [PubMed]
- Rupnik, A.; Lowndes, N.F.; Grenon, M. Mrn and the race to the break. Chromosoma 2010, 119, 115–135. [Google Scholar] [CrossRef] [PubMed]
- Bau, D.T.; Fu, Y.P.; Chen, S.T.; Cheng, T.C.; Yu, J.C.; Wu, P.E.; Shen, C.Y. Breast cancer risk and the DNA double-strand break end-joining capacity of nonhomologous end-joining genes are affected by BRCA1. Cancer Res. 2004, 64, 5013–5019. [Google Scholar] [CrossRef] [PubMed]
- Coupier, I.; Baldeyron, C.; Rousseau, A.; Mosseri, V.; Pages-Berhouet, S.; Caux-Moncoutier, V.; Papadopoulo, D.; Stoppa-Lyonnet, D. Fidelity of DNA double-strand break repair in heterozygous cell lines harbouring BRCA1 missense mutations. Oncogene 2004, 23, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Chen, C.F.; Chen, P.L.; Lee, W.H. BRCA1 facilitates microhomology-mediated end joining of DNA double strand breaks. J. Biol. Chem. 2002, 277, 28641–28647. [Google Scholar] [CrossRef] [PubMed]
- Bochar, D.A.; Wang, L.; Beniya, H.; Kinev, A.; Xue, Y.; Lane, W.S.; Wang, W.; Kashanchi, F.; Shiekhattar, R. BRCA1 is associated with a human swi/snf-related complex: Linking chromatin remodeling to breast cancer. Cell 2000, 102, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Cantor, S.B.; Bell, D.W.; Ganesan, S.; Kass, E.M.; Drapkin, R.; Grossman, S.; Wahrer, D.C.; Sgroi, D.C.; Lane, W.S.; Haber, D.A.; et al. BACH1, a novel helicase-like protein, interacts directly with BRCA1 and contributes to its DNA repair function. Cell 2001, 105, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cortez, D.; Yazdi, P.; Neff, N.; Elledge, S.J.; Qin, J. BASC, a super complex of BRCA1-associated proteins involved in the recognition and repair of aberrant DNA structures. Genes Dev. 2000, 14, 927–939. [Google Scholar] [PubMed]
- Yarden, R.I.; Brody, L.C. BRCA1 interacts with components of the histone deacetylase complex. Proc. Natl. Acad. Sci. USA 1999, 96, 4983–4988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, A.A.; Masson, J.Y.; McIlwraith, M.J.; Stasiak, A.Z.; Stasiak, A.; Venkitaraman, A.R.; West, S.C. Role of BRCA2 in control of the RAD51 recombination and DNA repair protein. Mol. Cell 2001, 7, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Chun, J.; Powell, S.N. BRCA1 and BRCA2: Different roles in a common pathway of genome protection. Nat. Rev. Cancer 2011, 12, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Miki, Y. Role of BRCA1 and BRCA2 as regulators of DNA repair, transcription, and cell cycle in response to DNA damage. Cancer Sci. 2004, 95, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Shahid, T.; Soroka, J.; Kong, E.; Malivert, L.; McIlwraith, M.J.; Pape, T.; West, S.C.; Zhang, X. Structure and mechanism of action of the BRCA2 breast cancer tumor suppressor. Nat. Struct. Mol. Biol. 2014, 21, 962–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robson, M.E. Clinical considerations in the management of individuals at risk for hereditary breast and ovarian cancer. Cancer Control 2002, 9, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Blakeman, V.; Williams, J.L.; Meng, Q.J.; Streuli, C.H. Circadian clocks and breast cancer. Breast Cancer Res. 2016, 18, 89. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.H.; Farkas, M.E. Altered circadian rhythms and breast cancer: From the human to the molecular level. Front. Endocrinol. (Lausanne) 2018, 9, 219. [Google Scholar] [CrossRef] [PubMed]
- Gaddameedhi, S.; Reardon, J.T.; Ye, R.; Ozturk, N.; Sancar, A. Effect of circadian clock mutations on DNA damage response in mammalian cells. Cell Cycle 2012, 11, 3481–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, S.M.; Belancio, V.P.; Dauchy, R.T.; Xiang, S.; Brimer, S.; Mao, L.; Hauch, A.; Lundberg, P.W.; Summers, W.; Yuan, L.; et al. Melatonin: An inhibitor of breast cancer. Endocr. Relat. Cancer 2015, 22, R183–R204. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A.; Lindsey-Boltz, L.A.; Gaddameedhi, S.; Selby, C.P.; Ye, R.; Chiou, Y.Y.; Kemp, M.G.; Hu, J.; Lee, J.H.; Ozturk, N. Circadian clock, cancer, and chemotherapy. Biochemistry 2015, 54, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.G. Circadian disruption and breast cancer: From melatonin to clock genes. Epidemiology 2005, 16, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between light at night, melatonin secretion, sleep deprivation, and the internal clock: Health impacts and mechanisms of circadian disruption. Life Sci. 2017, 173, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Copertaro, A.; Bracci, M. Working against the biological clock: A review for the occupational physician. Ind. Health 2019, 57. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.G.; Zhu, Y. Electric light, particularly at night, disrupts human circadian rhythmicity: Is that a problem? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, P.; Mirick, D.K.; Randolph, T.W.; Gong, J.; Buchanan, D.T.; Zhang, J.J.; Davis, S. Oxidative DNA damage during night shift work. Occup. Environ. Med. 2017, 74, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, G.C.; Rapisarda, V.; Marconi, A.; Scalisi, A.; Catalano, F.; Proietti, L.; Travali, S.; Libra, M.; Fenga, C. Correlation of the risk of breast cancer and disruption of the circadian rhythm (review). Oncol. Rep. 2012, 28, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Cordina-Duverger, E.; Menegaux, F.; Popa, A.; Rabstein, S.; Harth, V.; Pesch, B.; Bruning, T.; Fritschi, L.; Glass, D.C.; Heyworth, J.S.; et al. Night shift work and breast cancer: A pooled analysis of population-based case-control studies with complete work history. Eur. J. Epidemiol. 2018, 33, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.G.; Brainard, G.C.; Blask, D.E.; Lockley, S.W.; Motta, M.E. Breast cancer and circadian disruption from electric lighting in the modern world. CA Cancer J. Clin. 2014, 64, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Wegrzyn, L.R.; Tamimi, R.M.; Rosner, B.A.; Brown, S.B.; Stevens, R.G.; Eliassen, A.H.; Laden, F.; Willett, W.C.; Hankinson, S.E.; Schernhammer, E.S. Rotating night-shift work and the risk of breast cancer in the nurses’ health studies. Am. J. Epidemiol. 2017, 186, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Zhu, C.; Wang, M.; Mo, F.; Du, W.; Ma, X. Night shift work increases the risks of multiple primary cancers in women: A systematic review and meta-analysis of 61 articles. Cancer Epidemiol. Biomarkers Prev. 2018, 27, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J. Night shift work and risk of breast cancer. Curr. Environ. Health Rep. 2017, 4, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.E.; Lee, J.; Jang, T.W.; Kim, I.A.; Park, J.; Song, J. The relationship between night work and breast cancer. Ann. Occup. Environ. Med. 2018, 30, 11. [Google Scholar] [CrossRef] [PubMed]
- IARC Monographs Volume 124 Working Group. Carcinogenicity of night shift work. Lancet Oncol. 2019, 20, 1058–1059. [Google Scholar] [CrossRef] [PubMed]
- Lunn, R.M.; Blask, D.E.; Coogan, A.N.; Figueiro, M.G.; Gorman, M.R.; Hall, J.E.; Hansen, J.; Nelson, R.J.; Panda, S.; Smolensky, M.H.; et al. Health consequences of electric lighting practices in the modern world: A report on the national toxicology program’s workshop on shift work at night, artificial light at night, and circadian disruption. Sci. Total Environ. 2017, 607–608, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Murthy, P.; Muggia, F. Women’s cancers: How the discovery of BRCA genes is driving current concepts of cancer biology and therapeutics. Ecancermedicalscience 2019, 13, 904. [Google Scholar] [CrossRef] [PubMed]
- Boivin, D.B.; James, F.O.; Wu, A.; Cho-Park, P.F.; Xiong, H.; Sun, Z.S. Circadian clock genes oscillate in human peripheral blood mononuclear cells. Blood 2003, 102, 4143–4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusanagi, H.; Hida, A.; Satoh, K.; Echizenya, M.; Shimizu, T.; Pendergast, J.S.; Yamazaki, S.; Mishima, K. Expression profiles of 10 circadian clock genes in human peripheral blood mononuclear cells. Neurosci. Res. 2008, 61, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Kusanagi, H.; Mishima, K.; Satoh, K.; Echizenya, M.; Katoh, T.; Shimizu, T. Similar profiles in human period1 gene expression in peripheral mononuclear and polymorphonuclear cells. Neurosci. Lett. 2004, 365, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, M.; Hamada, A.; Tomoda, A.; Ohdo, S.; Ohmura, T.; Sakato, H.; Kawatani, J.; Jodoi, T.; Nakagawa, H.; Terazono, H.; et al. Daily expression of clock genes in whole blood cells in healthy subjects and a patient with circadian rhythm sleep disorder. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1273–R1279. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, F.; Matsubara, C.; Myung, J.; Yoritaka, T.; Kamimura, N.; Tsutsumi, S.; Kanai, A.; Suzuki, Y.; Sassone-Corsi, P.; Aburatani, H.; et al. Genome-wide profiling of the core clock protein bmal1 targets reveals a strict relationship with metabolism. Mol. Cell. Biol. 2010, 30, 5636–5648. [Google Scholar] [CrossRef] [PubMed]
- Rey, G.; Cesbron, F.; Rougemont, J.; Reinke, H.; Brunner, M.; Naef, F. Genome-wide and phase-specific DNA-binding rhythms of bmal1 control circadian output functions in mouse liver. PLoS Biol. 2011, 9, e1000595. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, Y.; Li, L.; Su, X.D. Intermolecular recognition revealed by the complex structure of human clock-bmal1 basic helix-loop-helix domains with e-box DNA. Cell Res. 2013, 23, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Yoshitane, H.; Ozaki, H.; Terajima, H.; Du, N.H.; Suzuki, Y.; Fujimori, T.; Kosaka, N.; Shimba, S.; Sugano, S.; Takagi, T.; et al. Clock-controlled polyphonic regulation of circadian rhythms through canonical and noncanonical e-boxes. Mol. Cell. Biol. 2014, 34, 1776–1787. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, P.; Mirick, D.K.; Davis, S. The impact of chronotype on melatonin levels among shift workers. Occup. Environ. Med. 2014, 71, 195–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshizaki, T.; Komatsu, T.; Tada, Y.; Hida, A.; Kawano, Y.; Togo, F. Association of habitual dietary intake with morningness-eveningness and rotating shift work in japanese female nurses. Chronobiol. Int. 2018, 35, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Kantermann, T.; Duboutay, F.; Haubruge, D.; Hampton, S.; Darling, A.L.; Berry, J.L.; Kerkhofs, M.; Boudjeltia, K.Z.; Skene, D.J. The direction of shift-work rotation impacts metabolic risk independent of chronotype and social jetlag—An exploratory pilot study. Chronobiol. Int. 2014, 31, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Yong, M.; Fischer, D.; Germann, C.; Lang, S.; Vetter, C.; Oberlinner, C. Are chronotype, social jetlag and sleep duration associated with health measured by work ability index? Chronobiol. Int. 2016, 33, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Bracci, M.; Ciarapica, V.; Copertaro, A.; Barbaresi, M.; Manzella, N.; Tomasetti, M.; Gaetani, S.; Monaco, F.; Amati, M.; Valentino, M.; et al. Peripheral skin temperature and circadian biological clock in shift nurses after a day off. Int. J. Mol. Sci. 2016, 17, 623. [Google Scholar] [CrossRef] [PubMed]
- Bracci, M.; Manzella, N.; Copertaro, A.; Staffolani, S.; Strafella, E.; Barbaresi, M.; Copertaro, B.; Rapisarda, V.; Valentino, M.; Santarelli, L. Rotating-shift nurses after a day off: Peripheral clock gene expression, urinary melatonin, and serum 17-beta-estradiol levels. Scand. J. Work. Environ. Health 2014, 40, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Stevens, R.G.; Hoffman, A.E.; Tjonneland, A.; Vogel, U.B.; Zheng, T.; Hansen, J. Epigenetic impact of long-term shiftwork: Pilot evidence from circadian genes and whole-genome methylation analysis. Chronobiol. Int. 2011, 28, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Forbes, C.; Fayter, D.; de Kock, S.; Quek, R.G. A systematic review of international guidelines and recommendations for the genetic screening, diagnosis, genetic counseling, and treatment of BRCA-mutated breast cancer. Cancer Manag. Res. 2019, 11, 2321–2337. [Google Scholar] [CrossRef] [PubMed]
- Kotsopoulos, J. BRCA mutations and breast cancer prevention. Cancers 2018, 10, 524. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.; Tutt, A.; Ashworth, A. Targeting the DNA repair defect of BRCA tumours. Curr. Opin. Pharmacol. 2005, 5, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Papantoniou, K.; Vetter, C.; Schernhammer, E.S. Shift work practices and opportunities for intervention. Occup. Environ. Med. 2017, 74, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Razavi, P.; Devore, E.E.; Bajaj, A.; Lockley, S.W.; Figueiro, M.G.; Ricchiuti, V.; Gauderman, W.J.; Hankinson, S.E.; Willett, W.C.; Schernhammer, E.S. Shift work, chronotype, and melatonin rhythm in nurses. Cancer Epidemiol. Biomarkers Prev. 2019, 28, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, S.A.R.; Mortazavi, S.M.J. Women with hereditary breast cancer predispositions should avoid using their smartphones, tablets, and laptops at night. Iran. J. Basic Med. Sci. 2018, 21, 112–115. [Google Scholar] [PubMed]
- Horne, J.A.; Ostberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar] [PubMed]
- Bracci, M.; Copertaro, A.; Manzella, N.; Staffolani, S.; Strafella, E.; Nocchi, L.; Barbaresi, M.; Copertaro, B.; Rapisarda, V.; Valentino, M.; et al. Influence of night-shift and napping at work on urinary melatonin, 17-beta-estradiol and clock gene expression in pre-menopausal nurses. J. Biol. Regul. Homeost. Agents 2013, 27, 267–274. [Google Scholar] [PubMed]
- Randler, C.; Engelke, J. Gender differences in chronotype diminish with age: A meta-analysis based on morningness/chronotype questionnaires. Chronobiol. Int. 2019, 36, 888–905. [Google Scholar] [CrossRef] [PubMed]
- Taillard, J.; Philip, P.; Chastang, J.F.; Bioulac, B. Validation of horne and ostberg morningness-eveningness questionnaire in a middle-aged population of french workers. J. Biol. Rhythms 2004, 19, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Tonetti, L.; Natale, V. Discrimination between extreme chronotypes using the full and reduced version of the morningness-eveningness questionnaire. Chronobiol. Int. 2019, 36, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, M.; Dinich, J.; Merrow, M.; Roenneberg, T. Social jetlag: Misalignment of biological and social time. Chronobiol. Int. 2006, 23, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Pick, R.; He, W.; Chen, C.S.; Scheiermann, C. Time-of-day-dependent trafficking and function of leukocyte subsets. Trends Immunol. 2019, 40, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Teboul, M.; Barrat-Petit, M.A.; Li, X.M.; Claustrat, B.; Formento, J.L.; Delaunay, F.; Levi, F.; Milano, G. Atypical patterns of circadian clock gene expression in human peripheral blood mononuclear cells. J. Mol. Med. (Berl.) 2005, 83, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time pcr data by the comparative c(t) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Refinetti, R. Circadian software. Available online: https://www.circadian.org (accessed on 30 July 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Shift Workers (n = 44) | % | Daytime Workers (n = 45) | % | p-Value | ||
|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||||
| Age (years) | 40.2 | 9.5 | 43.0 | 10.7 | 0.196 | ||
| Job Seniority (years) | 15.9 | 5.4 | 17.1 | 4.7 | 0.266 | ||
| Shift Work Seniority (years) | 15.9 | 5.4 | |||||
| Night Shift Work (nights per month) | 5.6 | 1.3 | |||||
| Smokers (%) | 20.5 | 17.8 | 0.748 | ||||
| BMI | 23.6 | 3.4 | 23.5 | 7.5 | 0.936 | ||
| Physical Activity (hours/week) | 3.1 | 3.2 | 2.7 | 2.0 | 0.480 | ||
| Chronotype (MEQ score) | 53.7 | 7.1 | 58.3 | 7.9 | 0.005 | ||
| Wake-up Time on Blood Sampling Day | 6:22 | 0:20 | 6:18 | 0:42 | 0.569 | ||
| Social Jet Lag (minutes) | 52.4 | 31.1 | 36.7 | 20.7 | 0.006 | ||
| Exposure to Sunlight (minutes) | 100.9 | 69.7 | 105.3 | 71.4 | 0.769 | ||
| Use of Video Display Devices after Dinner (minutes) | 104.1 | 59.7 | 99.0 | 60.1 | 0.689 | ||
| Home Light Intensity (low/medium/high) | 15.9/79.5/4.6 | 22.2/51.1/26.7 | 0.006 | ||||
| Parameters | BRCA1 | BRCA2 | Cortisol | PER2 | Night Shift Work (nights per month) |
|---|---|---|---|---|---|
| BRCA1 | 1 | 0.830 * | −0.005 | 0.496 * | −0.284 * |
| BRCA2 | 0.830 * | 1 | 0.107 | 0.477 * | −0.309 * |
| Cortisol | −0.005 | 0.107 | 1 | 0.067 | −0.127 |
| PER2 | 0.496 * | 0.477 * | 0.067 | 1 | −0.267 * |
| Night-shift work (nights per month) | −0.284 * | −0.309 * | −0.127 | −0.267 * | 1 |
| Parameters | BRCA1 Gene Expression | BRCA2 Gene Expression | Cortisol | PER2 Gene Expression | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | p-Value | β | p-Value | β | p-Value | β | p-Value | β | p-Value | β | p-Value | β | p-Value | β | p-Value | |
| Shift Work | −0.258 | 0.016 | −0.298 | 0.012 | −0.282 | 0.008 | −0.296 | 0.012 | −0.115 | 0.310 | −0.177 | 0.170 | −0.269 | 0.011 | −0.262 | 0.031 |
| Age | — | — | −0.057 | 0.633 | — | — | −0.093 | 0.492 | — | — | −0.179 | 0.173 | — | — | −0.004 | 0.976 |
| Chronotype | — | — | −0.231 | 0.072 | — | — | −0.085 | 0.500 | — | — | −0.196 | 0.151 | — | — | −0.118 | 0.356 |
| Wake Up Time on Blood Sampling Day | — | — | −0.257 | 0.039 | — | — | −0.270 | 0.028 | — | — | −0.139 | 0.288 | — | — | −0.133 | 0.297 |
| Social Jet Lag | — | — | 0.035 | 0.775 | — | — | 0.083 | 0.487 | — | — | −0.205 | 0.120 | — | — | 0.011 | 0.927 |
| Home Light Intensity | — | — | 0.092 | 0.428 | — | — | 0.131 | 0.253 | — | — | −0.061 | 0.622 | — | — | 0.034 | 0.778 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bracci, M.; Ciarapica, V.; Eléxpuru Zabaleta, M.; Tartaglione, M.F.; Pirozzi, S.; Giuliani, L.; Piva, F.; Valentino, M.; Ledda, C.; Rapisarda, V.; et al. BRCA1 and BRCA2 Gene Expression: Diurnal Variability and Influence of Shift Work. Cancers 2019, 11, 1146. https://doi.org/10.3390/cancers11081146
Bracci M, Ciarapica V, Eléxpuru Zabaleta M, Tartaglione MF, Pirozzi S, Giuliani L, Piva F, Valentino M, Ledda C, Rapisarda V, et al. BRCA1 and BRCA2 Gene Expression: Diurnal Variability and Influence of Shift Work. Cancers. 2019; 11(8):1146. https://doi.org/10.3390/cancers11081146
Chicago/Turabian StyleBracci, Massimo, Veronica Ciarapica, Maria Eléxpuru Zabaleta, Maria Fiorella Tartaglione, Silvia Pirozzi, Letizia Giuliani, Francesco Piva, Matteo Valentino, Caterina Ledda, Venerando Rapisarda, and et al. 2019. "BRCA1 and BRCA2 Gene Expression: Diurnal Variability and Influence of Shift Work" Cancers 11, no. 8: 1146. https://doi.org/10.3390/cancers11081146