FAM83A and FAM83B as Prognostic Biomarkers and Potential New Therapeutic Targets in NSCLC

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
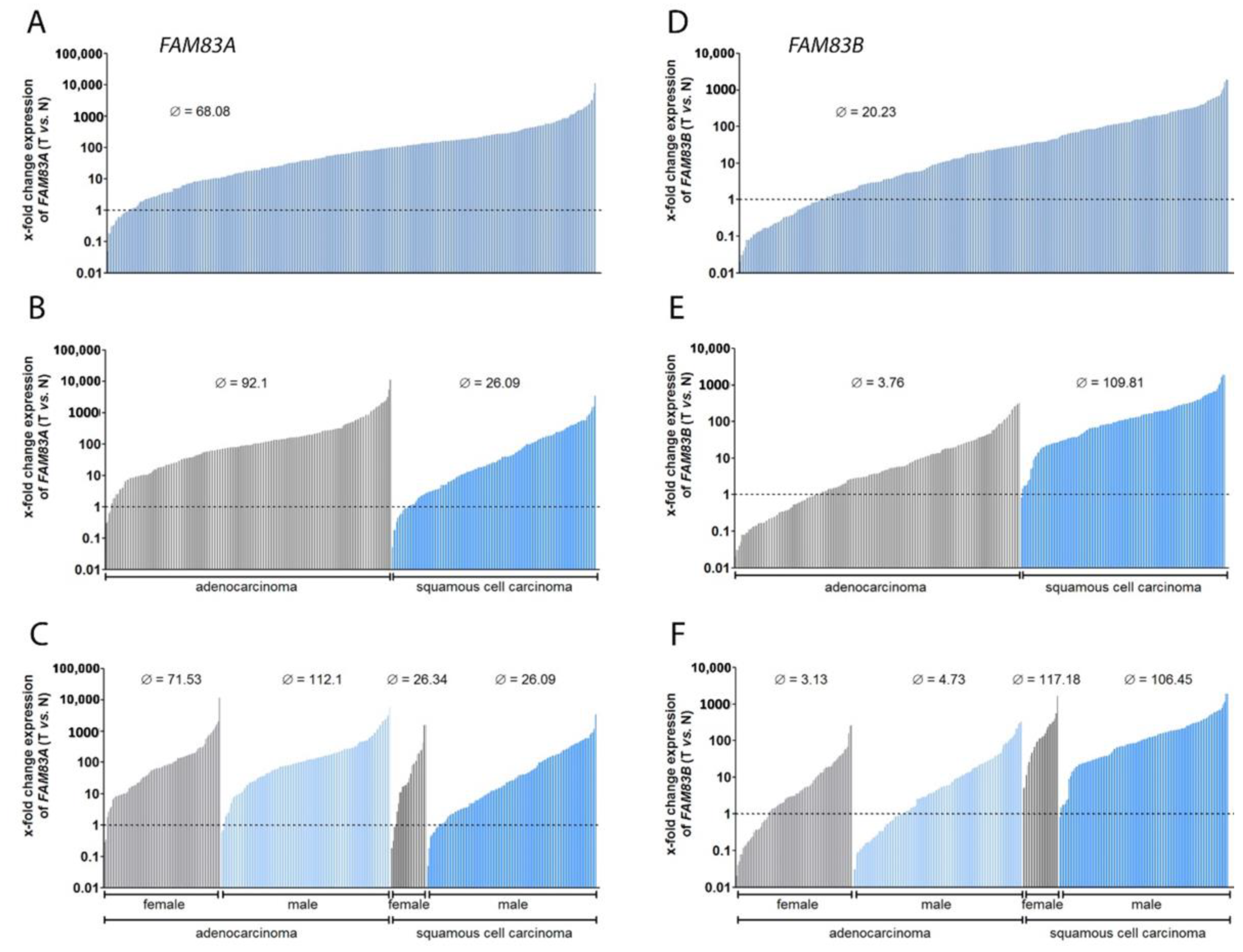
2.1. FAM83A and B Are Highly Overexpressed in NSCLC
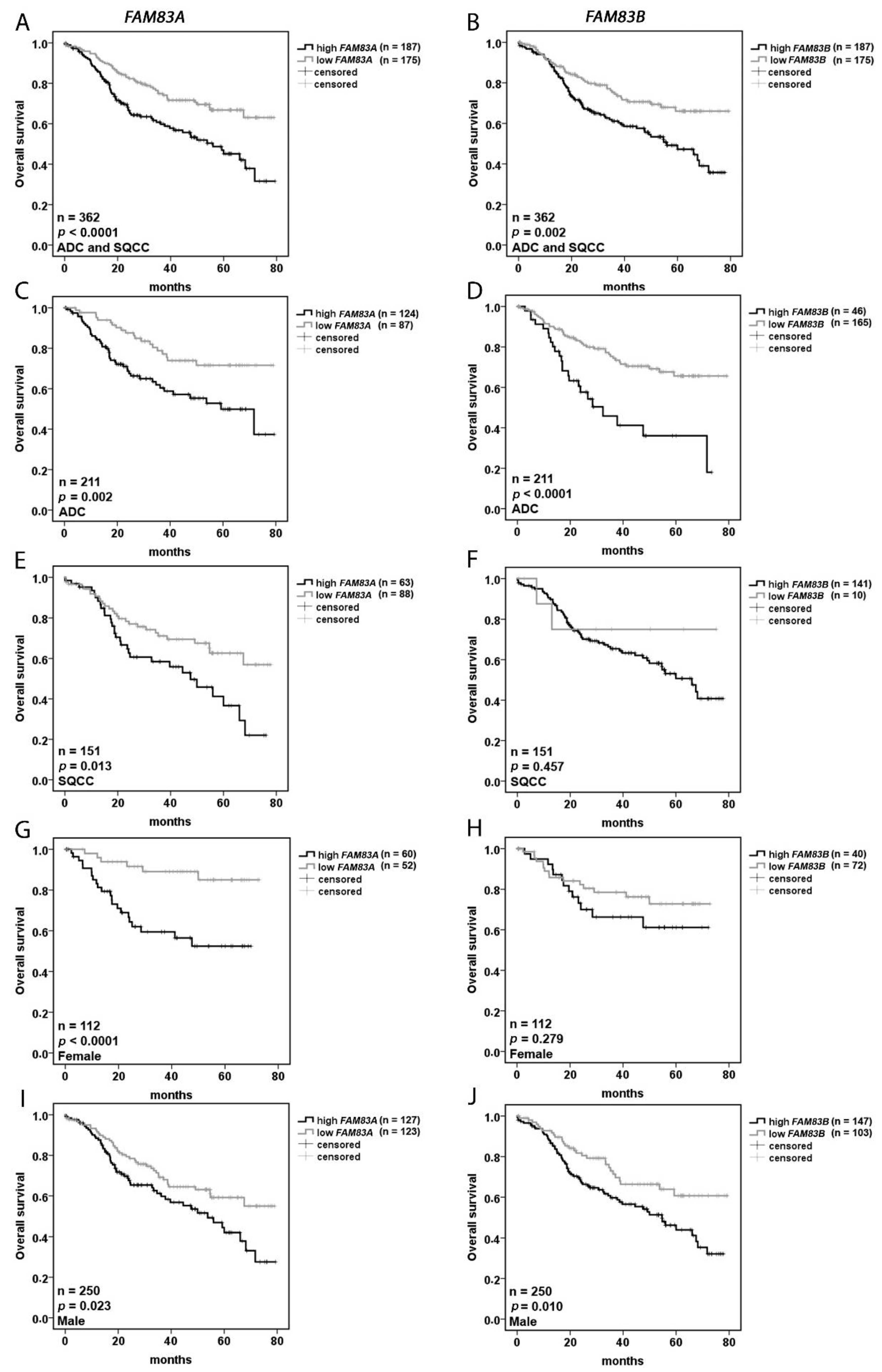
2.2. FAM83A and FAM83B Are Prognostic Markers for NSCLC
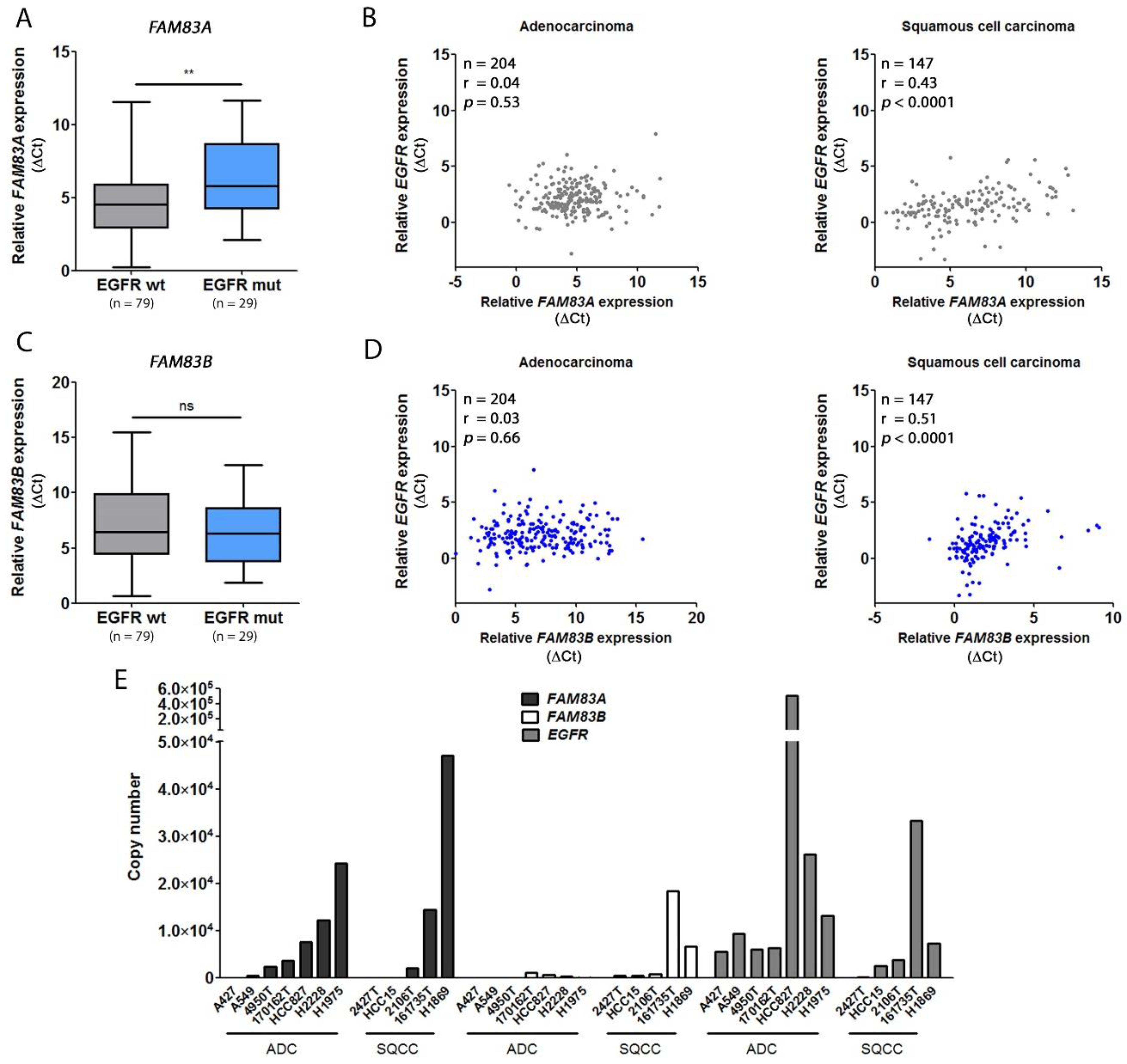
2.3. The Expression Levels of FAM83A and B Are Related to EGFR Expression In Vitro and In Vivo
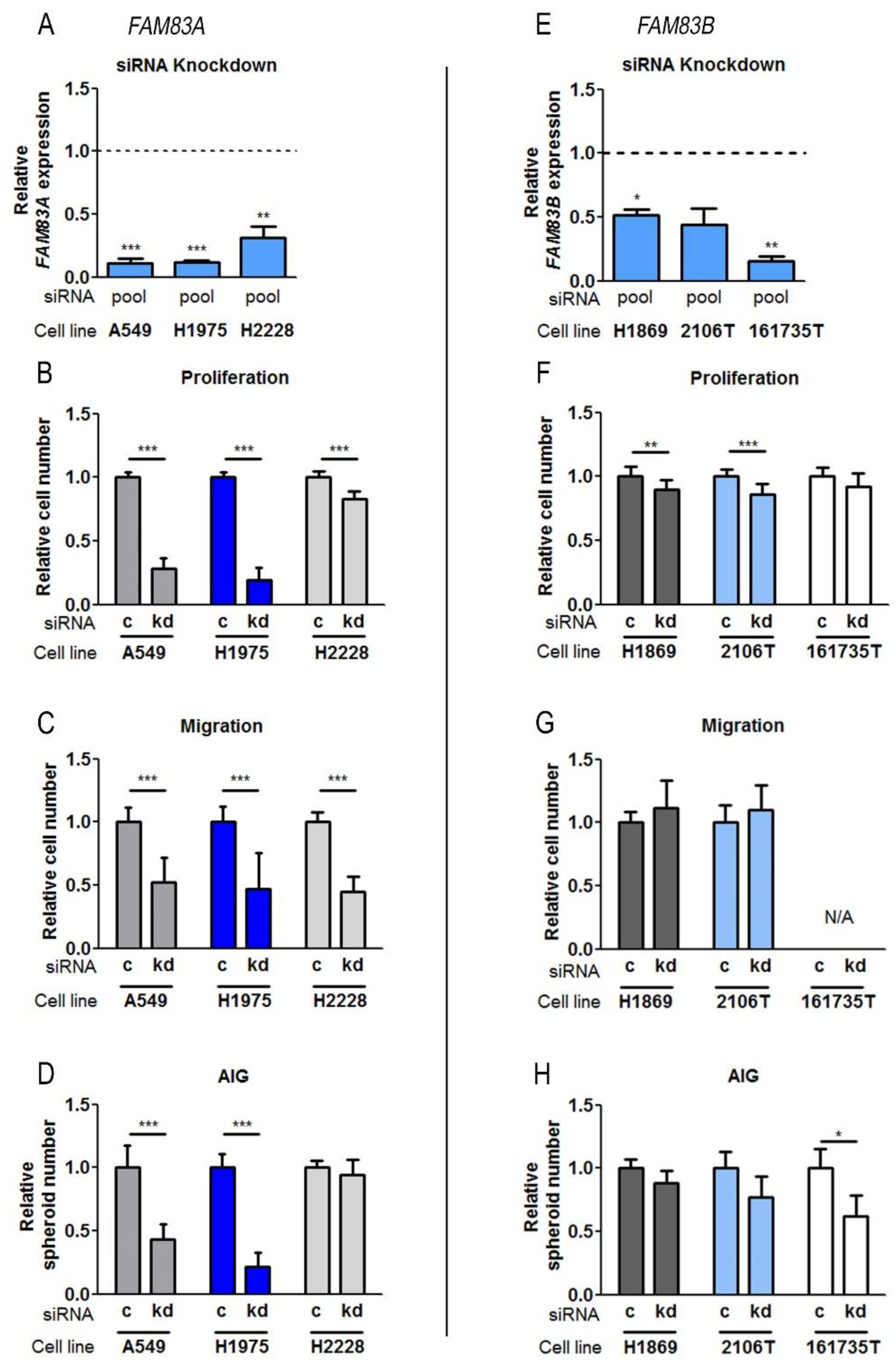
2.4. Influence of FAM83A and FAM83B on Biological Functions in NSCLC Cell Lines
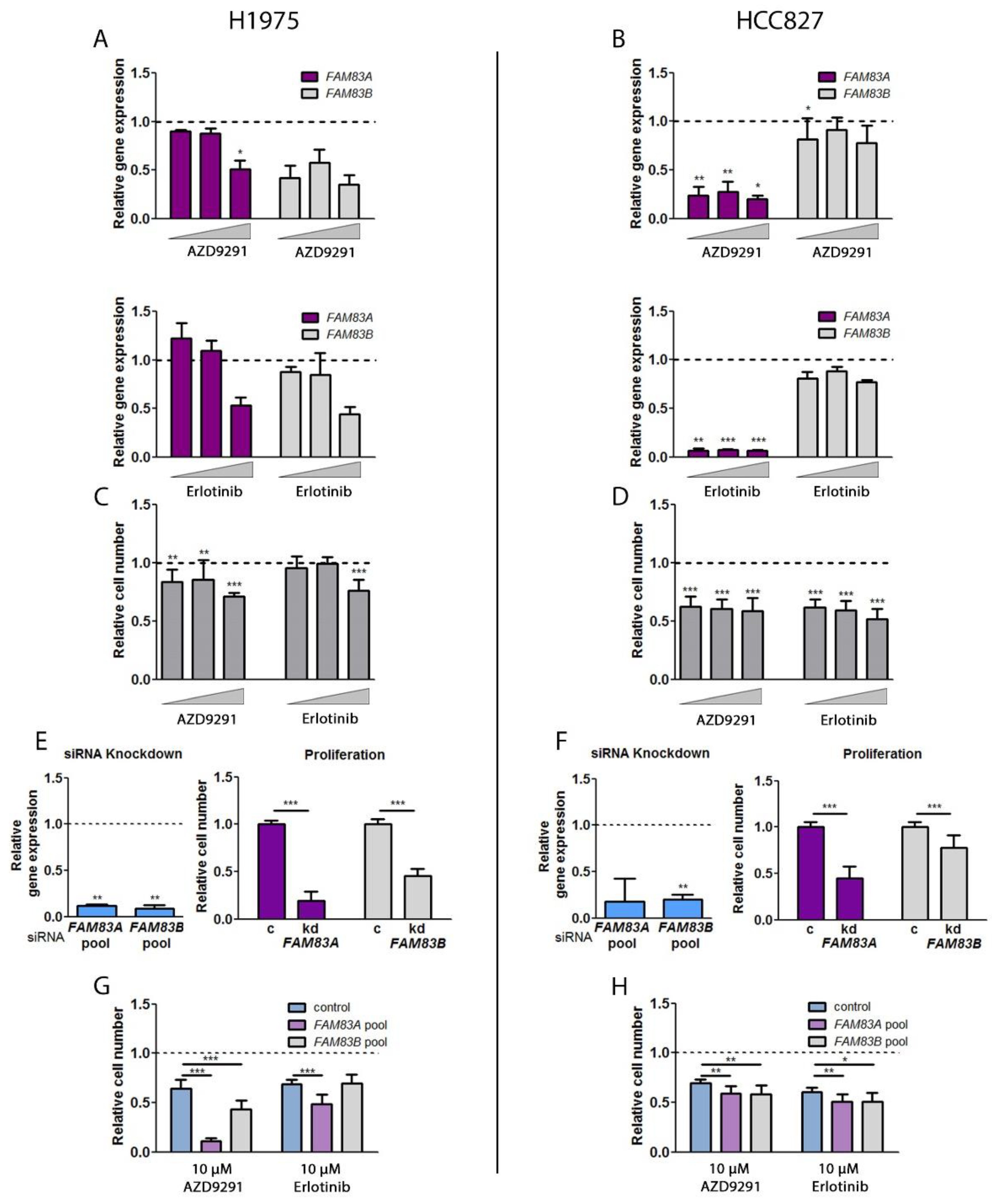
2.5. Impact of TKI Treatment and EGFR Inhibition on FAM83A and B Gene Expression and Cell Viability
3. Discussion
4. Materials and Methods
4.1. Tissue Sample Collection, Characterization and Preparation
4.2. Cultivation of Cells
4.3. Transient Gene Knockdown by siRNA Transfection
4.4. Tyrosine Kinase Treatment
4.5. Total RNA Isolation and cDNA Synthesis
4.6. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.7. Proliferation Assay and Quantification of Cells by Lactate Dehydrogenase (LDH) Activity Measurement
4.8. Soft Agar Assay
4.9. Migration Assay
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Forman, D.; Mathers, C.; Soerjomataram, I.; Bray, F.; Eser, S.; Rebelo, M.; Dikshit, R.; Parkin, D.M.; Ferlay, J. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2014, 136, E359–E386. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA. Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Devesa, S.S.; Bray, F.; Vizcaino, A.P.; Parkin, D.M. International lung cancer trends by histologic type: Male:female differences diminishing and adenocarcinoma rates rising. Int. J. Cancer 2005, 117, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288–300. [Google Scholar] [CrossRef]
- Lynch, T.J.; Bell, D.W.; Sordella, R.; Gurubhagavatula, S.; Okimoto, R.A.; Brannigan, B.W.; Harris, P.L.; Haserlat, S.M.; Supko, J.G.; Haluska, F.G.; et al. Activating Mutations in the Epidermal Growth Factor Receptor Underlying Responsiveness of Non–Small-Cell Lung Cancer to Gefitinib. N. Engl. J. Med. 2004, 350, 2129–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Groenendijk, F.H.; Bernards, R. Drug resistance to targeted therapies: Déjà vu all over again. Mol. Oncol. 2014, 8, 1067–1083. [Google Scholar] [CrossRef] [PubMed]
- Cipriano, R.; Graham, J.; Miskimen, K.L.S.; Bryson, B.L.; Bruntz, R.C.; Scott, S.A.; Brown, H.A.; Stark, G.R.; Jackson, M.W. FAM83B mediates EGFR- and RAS-driven oncogenic transformation. J. Clin. Investig. 2012, 122, 3197–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Meier, R.; Furuta, S.; Lenburg, M.E.; Kenny, P.A.; Xu, R.; Bissell, M.J. FAM83A confers EGFR-TKI resistance in breast cancer cells and in mice. J. Clin. Investig. 2012, 122, 3211–3220. [Google Scholar] [CrossRef] [PubMed]
- Bartel, C.A.; Parameswaran, N.; Cipriano, R.; Jackson, M.W. FAM83 proteins: Fostering new interactions to drive oncogenic signaling and therapeutic resistance. Oncotarget 2016, 7. [Google Scholar] [CrossRef]
- Bufton, J.C.; Dunbar, K.; Fulcher, L.J.; Pinkas, D.M.; Weidlich, S.; Smith, J.C.; Macartney, T.J.; Tachie-Menson, T.; Wood, N.T.; Sapkota, G.P.; et al. The DUF1669 domain of FAM83 family proteins anchor casein kinase 1 isoforms. Sci. Signal. 2018, 11, eaao2341. [Google Scholar]
- Bartel, C.A.; Jackson, M.W. HER2-positive breast cancer cells expressing elevated FAM83A are sensitive to FAM83A loss. PLoS ONE 2017, 12, 1–18. [Google Scholar] [CrossRef]
- Cipriano, R.; Miskimen, K.L.S.; Bryson, B.L.; Foy, C.R.; Bartel, C.A.; Jackson, M.W. FAM83B-mediated activation of PI3K/AKT and MAPK signaling cooperates to promote epithelial cell transformation and resistance to targeted therapies. Oncotarget 2013, 4, 729–738. [Google Scholar] [CrossRef] [Green Version]
- Grant, S. FAM83A and FAM83B: Candidate oncogenes and TKI resistance mediators. J. Clin. Investig. 2012, 122, 3048–3051. [Google Scholar] [CrossRef]
- Alamgeer, M.; Ganju, V.; Watkins, D.N. Novel therapeutic targets in non-small cell lung cancer. Curr. Opin. Pharmacol. 2013, 13, 394–401. [Google Scholar] [CrossRef]
- Chu, P.G.; Lyda, M.H.; Weiss, L.M. Cytokeratin 14 expression in epithelial neoplasms: A survey of 435 cases with emphasis on its value in differentiating squamous cell carcinomas from other epithelial tumours. Histopathology 2001, 39, 9–16. [Google Scholar] [CrossRef]
- Szirmai, J. The least dense hyperball covering of regular prism tilings in hyperbolic n-space. Ann. di Mat. Pura ed Appl. 2016, 195, 235–248. [Google Scholar] [CrossRef]
- Schneider, M.A.; Translational Research Unit, Thoraxklinik at Heidelberg University Hospital, Heidelberg, Germany; Meister, M.; Translational Research Unit, Thoraxklinik at Heidelberg University Hospital, Heidelberg, Germany. Personal communication, 2018.
- Moyer, J.D.; Barbacci, E.G.; Iwata, K.K.; Arnold, L.; Boman, B.; Cnnningham, A.; Diorio, C.; Doty, J.; Morin, M.J.; Moyer, M.P.; et al. Induction of apoptosis and cell cycle arrest by CP-358,774, an inhibitor of epidermal growth factor receptor tyrosine kinase. Cancer Res. 1997, 57, 4838–4848. [Google Scholar]
- Pao, W.; Mardis, E.; Rusch, V.; Wilson, R.; Kupfer, D.; Zakowski, M.; Doherty, J.; Politi, K.; Singh, B.; Heelan, R.; et al. EGF receptor gene mutations are common in lung cancers from “never smokers” and are associated with sensitivity of tumors to gefitinib and erlotinib. Proc. Natl. Acad. Sci. USA 2004, 101, 13306–13311. [Google Scholar] [CrossRef] [Green Version]
- Cross, D.A.E.; Ashton, S.E.; Ghiorghiu, S.; Eberlein, C.; Nebhan, C.A.; Spitzler, P.J.; Orme, J.P.; Finlay, M.R.V.; Ward, R.A.; Mellor, M.J.; et al. AZD9291, an irreversible EGFR TKI, overcomes T790M-mediated resistance to EGFR inhibitors in lung cancer. Cancer Discov. 2014, 4, 1046–1061. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Dong, X.; Yin, Y.; Su, Y.; Xu, Q.; Zhang, Y.; Pang, X.; Chen, W. BJ-TSA-9, a novel human tumor-specific gene, has potential as a biomarker of lung cancer. Neoplasia 2005, 7, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Okabe, N.; Ezaki, J.; Yamaura, T.; Muto, S.; Osugi, J.; Tamura, H.; Imai, J.I.; Ito, E.; Yanagisawa, Y.; Honma, R.; et al. FAM83B is a novel biomarker for diagnosis and prognosis of lung squamous cell carcinoma. Int. J. Oncol. 2015, 46, 999–1006. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Christiani, D.C. East meets west: Ethnic differences in epidemiology and clinical behaviors of lung cancer between east Asians and Caucasians. Chin. J. Cancer 2011, 30, 287–292. [Google Scholar] [CrossRef]
- Boyer, A.P.; Collier, T.S.; Vidavsky, I.; Bose, R. Quantitative Proteomics with siRNA Screening Identifies Novel Mechanisms of Trastuzumab Resistance in HER2 Amplified Breast Cancers. Mol. Cell. Proteom. 2012, 12, 180–193. [Google Scholar] [CrossRef]
- Ullrich, A.; Wong, S.; Slamon, D.; Levin, W.; Clark, G.; McGuire, W. Human breast cancer: Correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 2006, 235, 177–182. [Google Scholar]
- Yamaura, T.; Ezaki, J.; Okabe, N.; Takagi, H.; Ozaki, Y.; Inoue, T.; Watanabe, Y.; Fukuhara, M.; Muto, S.; Matsumura, Y.; et al. Family with sequence similarity 83, member B is a predictor of poor prognosis and a potential therapeutic target for lung adenocarcinoma expressing wild-type epidermal growth factor receptor. Oncol. Lett. 2018, 15, 1549–1558. [Google Scholar] [PubMed]
- Beasley, M.B.; Brambilla, E.; Travis, W.D. The 2004 World Health Organization Classification of Lung Tumors. Semin. Roentgenol. 2005, 40, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Wittekind, C.; Bertolini, J. TNM system 2010. Der Onkol. 2010, 16, 175–180. [Google Scholar] [CrossRef]
- Gottschling, S.; Jauch, A.; Kuner, R.; Herpel, E.; Mueller-Decker, K.; Schnabel, P.A.; Xu, E.C.; Muley, T.; Sültmann, H.; Bender, C.; et al. Establishment and comparative characterization of novel squamous cell non-small cell lung cancer cell lines and their corresponding tumor tissue. Lung Cancer 2012, 75, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.A.; Granzow, M.; Warth, A.; Schnabel, P.A.; Thomas, M.; Herth, F.J.F.; Dienemann, H.; Muley, T.; Meister, M. Glycodelin: A new biomarker with immunomodulatory functions in non-small cell lung cancer. Clin. Cancer Res. 2015, 21, 3529–3540. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum Information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Liu, F. Soft–Agar colony Formation Assay. Bio-protocol 2012, 2, e220. [Google Scholar] [CrossRef]
- Schneider, M.A. In vitro Tumor Cell Migration Assay Using ThinCertsTM (Transwells). Bio-protocol 2016, 6, e1830. [Google Scholar] [CrossRef]
- McShane, L.M.; Altman, D.G.; Sauerbrei, W.; Taube, S.E.; Gion, M.; Clark, G.M. REporting recommendations for tumor MARKer prognostic studies (REMARK). Breast Cancer Res. Treat. 2006, 100, 229–235. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene expression Analyses | |||||
|---|---|---|---|---|---|
| Parameter | n | (%) | Parameter | n | (%) |
| Median Age | 65 (38–88) | P Stage | |||
| Gender | 362 | IA | 37 | 10 | |
| Male | 250 | 69 | IB | 90 | 25 |
| Female | 112 | 31 | IIA | 70 | 19 |
| Histology | IIB | 51 | 14 | ||
| Adeno | 211 | 58 | IIIA | 105 | 29 |
| Squamous | 151 | 42 | IIIB | 9 | 2 |
| Therapy | ECOG | ||||
| OP | 212 | 59 | 0 | 320 | 88 |
| OP/RT | 13 | 4 | 1 | 32 | 9 |
| OP/CT | 100 | 28 | 2 | 4 | 1 |
| OP/RT/CT | 37 | 10 | n.d. | 8 | 2 |
| FAM | FAM83B | Histology | p-Value |
|---|---|---|---|
| FAM83A | r = 0.21 | ADC | 0.0025 |
| FAM83A | r = 0.35 | SQCC | <0.0001 |
| Univariate Analysis | ||
| Variable (high vs. low) | Significance | Hazard Ratio (95% CI) |
| FAM83A | <0.0001 | 1.965 (1.366–2.817) |
| FAM83B | 0.002 | 1.808 (1.252–2.612) |
| Multivariate Analysis | ||
| Variable | Significance | Hazard Ratio (95% CI) |
| FAM83A (high vs. low) | <0.0001 | 1.980 (1.370–2.857) |
| Stage | <0.0001 | 1.076 (1.050–1.103) |
| Age | 0.018 | 1.024 (1.004–1.045) |
| Sex (female vs. male) | 0.062 | 0.664 (0.431–1.022) |
| Histology (ADC vs. SQCC) | 0.391 | 0.853 (0.593–1.227) |
| FAM83B (high vs. low) | 0.001 | 2.283 (1.393–3.745) |
| Stage | <0.0001 | 1.073 (1.047–1.100) |
| Age | 0.018 | 1.024 (1.004–1.044) |
| Sex (female vs. male) | 0.084 | 0.682 (0.442–1.052) |
| Histology (ADC vs. SQCC) | 0.036 | 1.663 (1.034–2.672) |
| Gene Expression Analyses | |||||
|---|---|---|---|---|---|
| Parameter | n | (%) | Parameter | n | (%) |
| Median Age | 62 (40–81) | P Stage | |||
| Gender | 108 | IA | 12 | 11 | |
| Male | 57 | 53 | IB | 34 | 31 |
| Female | 51 | 47 | IIA | 4 | 4 |
| Histology | IIB | 14 | 13 | ||
| Adeno | 108 | 100 | IIIA | 35 | 32 |
| Therapy | IIIB | 4 | 4 | ||
| OP | 52 | 48 | IV | 5 | 5 |
| OP/RT | 8 | 7 | ECOG | ||
| OP/CT | 30 | 28 | 0 | 88 | 81 |
| OP/RT/CT | 18 | 17 | 1 | 17 | 16 |
| EGFR Driver Mutation | 29 | 27 | 2 | 2 | 2 |
| Exon 19 | 14 | 13 | n.d. | 1 | 1 |
| Exon 20 | 1 | 1 | |||
| Exon 21 | 14 | 13 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richtmann, S.; Wilkens, D.; Warth, A.; Lasitschka, F.; Winter, H.; Christopoulos, P.; Herth, F.J.F.; Muley, T.; Meister, M.; Schneider, M.A. FAM83A and FAM83B as Prognostic Biomarkers and Potential New Therapeutic Targets in NSCLC. Cancers 2019, 11, 652. https://doi.org/10.3390/cancers11050652
Richtmann S, Wilkens D, Warth A, Lasitschka F, Winter H, Christopoulos P, Herth FJF, Muley T, Meister M, Schneider MA. FAM83A and FAM83B as Prognostic Biomarkers and Potential New Therapeutic Targets in NSCLC. Cancers. 2019; 11(5):652. https://doi.org/10.3390/cancers11050652
Chicago/Turabian StyleRichtmann, Sarah, Dennis Wilkens, Arne Warth, Felix Lasitschka, Hauke Winter, Petros Christopoulos, Felix J. F. Herth, Thomas Muley, Michael Meister, and Marc A. Schneider. 2019. "FAM83A and FAM83B as Prognostic Biomarkers and Potential New Therapeutic Targets in NSCLC" Cancers 11, no. 5: 652. https://doi.org/10.3390/cancers11050652