LC/MS-Based Polar Metabolite Profiling Identified Unique Biomarker Signatures for Cervical Cancer and Cervical Intraepithelial Neoplasia Using Global and Targeted Metabolomics

Abstract
:1. Introduction
2. Results
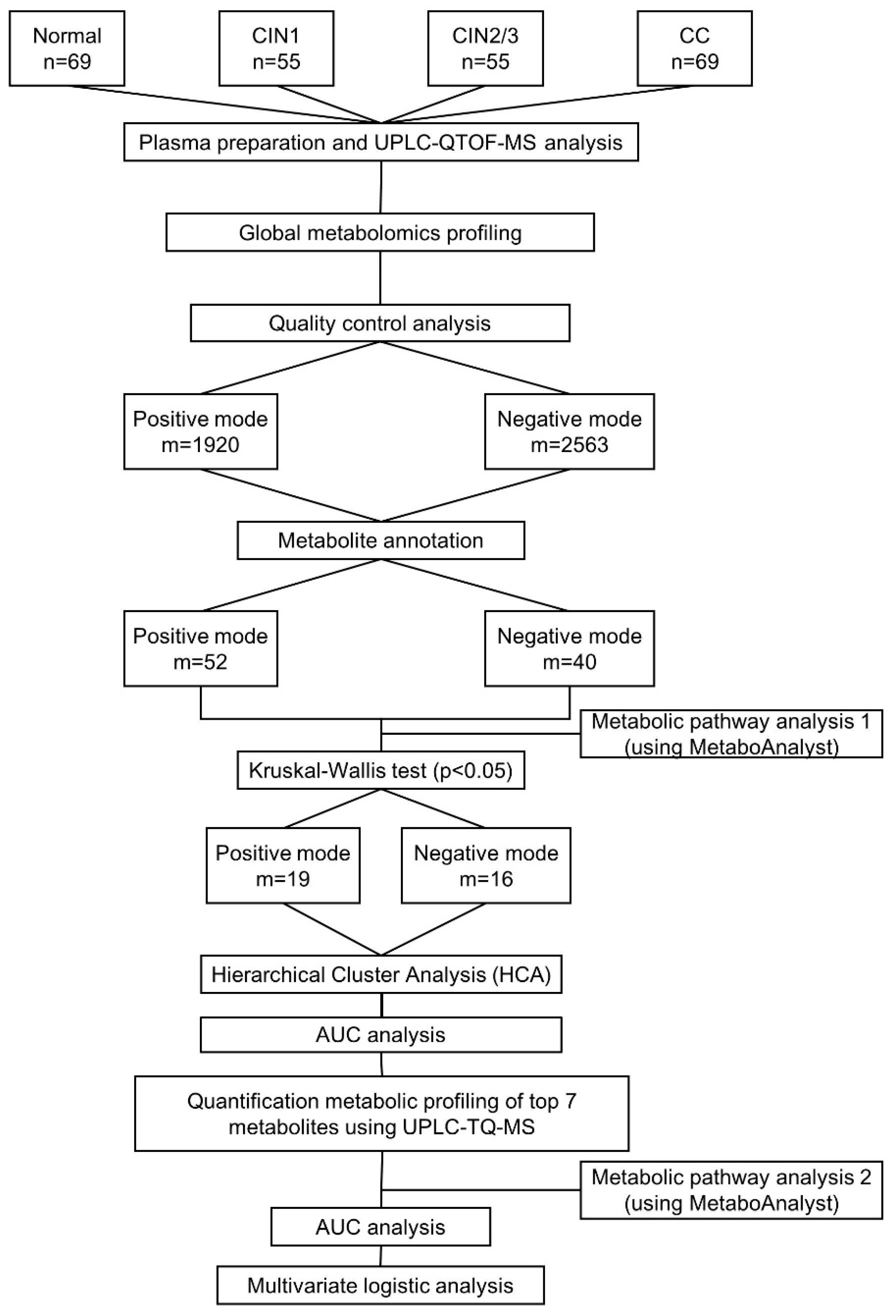
2.1. General Characteristics of Study Participants
2.2. Global Metabolic Profiling of Plasma by UPLC-QTOF-MS
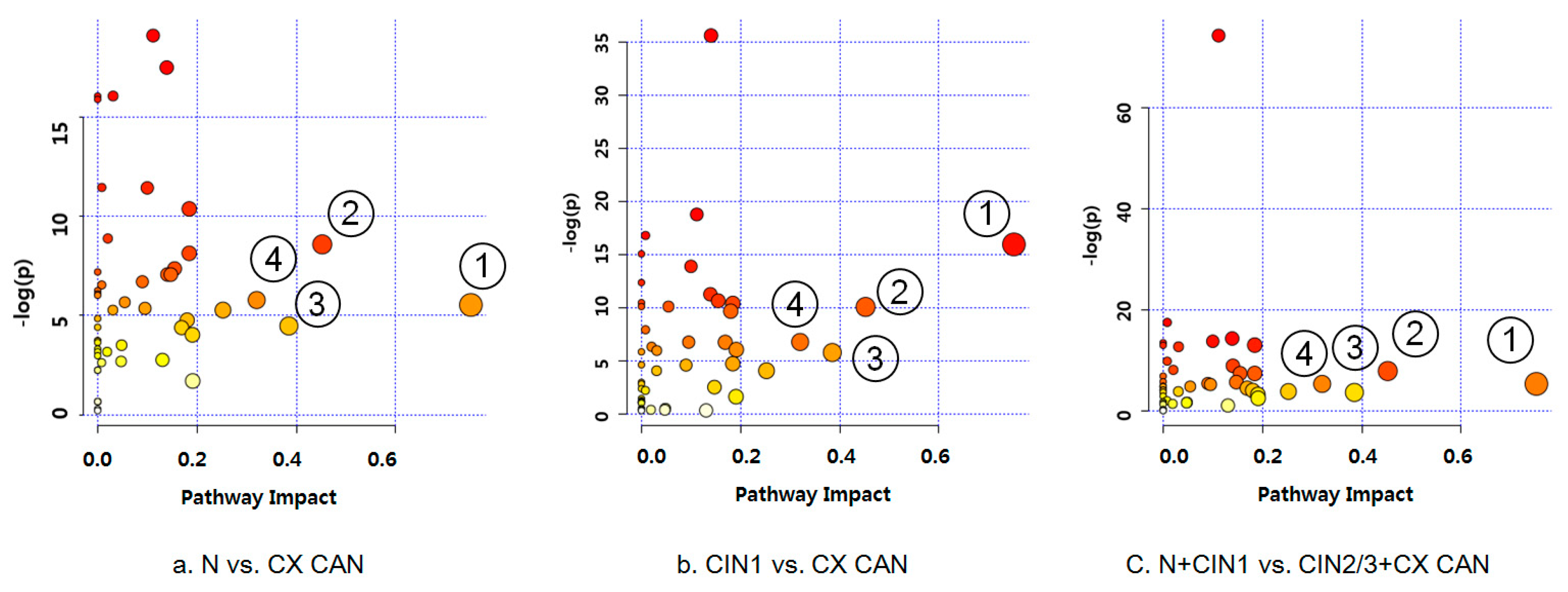
2.3. Differential Mapping of Metabolites in Pathway Analysis
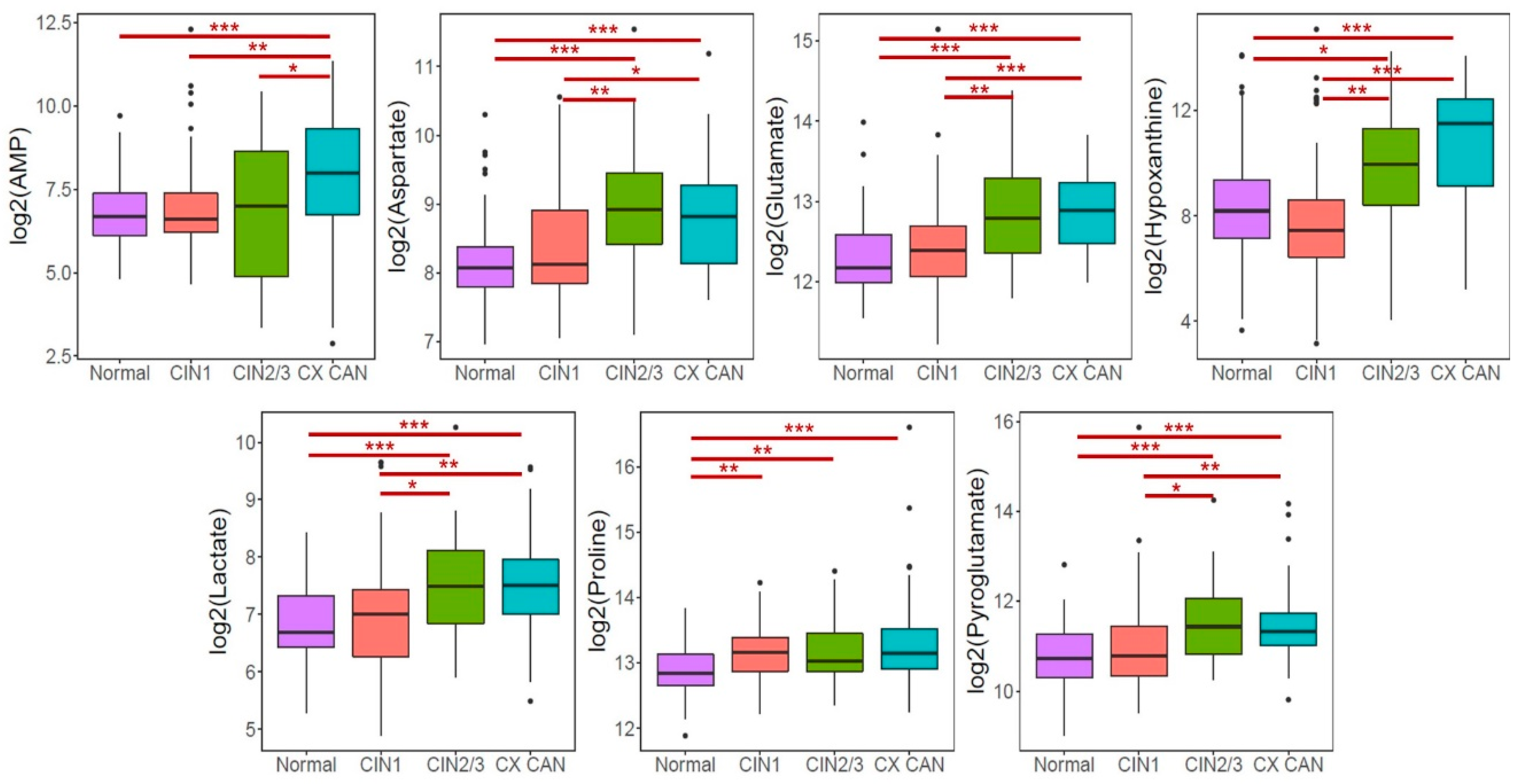
2.4. Altered Metabolites in Patients with CINs and Cervical Cancer
2.5. Validation and Diagnostic Performance of Selected Metabolites
2.6. Association of Metabolites with CINs and Cervical Cancer Risk
2.7. Combined Effects of Targeted Metabolites with HPV Status
2.8. Pathway Analysis for Quantitative Metabolites
3. Discussion
4. Materials and Methods
4.1. Study Population and Sample Collection
4.2. Global Metabolite Profiling Using UPLC-QTOF-MS
4.3. Metabolite Quantification Using UPLC-TQ-MS
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMP | Adenosine monophosphate |
| AUC | Area under the curve |
| BMI | Body mass index |
| CI | Confidence interval |
| CIN | Cervical intraepithelial neoplasia |
| CV | Coefficients of variation |
| FDR | False discovery rate |
| HCA | Hierarchical cluster analysis |
| HPV | Human papillomavirus |
| OR | Odds ratio |
| PCA | Principal component analysis |
| QC | Quality control |
| UPLC-QTOF-MS | Ultra-performance liquid chromatography-quadrupole-time-of-flight mass spectrometry |
| UPLC-TQ-MS | Ultra-performance liquid chromatography-triple-quadrupole mass spectrometry |
References
- Yang, X.; Da, M.; Zhang, W.; Qi, Q.; Zhang, C.; Han, S. Role of Lactobacillus in cervical cancer. Cancer Manag. Res. 2018, 10, 1219. [Google Scholar] [CrossRef]
- Fitzmaurice, C.; Dicker, D.; Pain, A.; Hamavid, H.; Moradi-Lakeh, M.; MacIntyre, M.F.; Allen, C.; Hansen, G.; Woodbrook, R.; Wolfe, C.; et al. The Global Burden of Cancer 2013. JAMA Oncol. 2015, 1, 505–527. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.-S.; Oh, H.Y.; Lee, J.-K.; Kong, J.-S.; Lee, D.O.; Kim, M.K. Combined effect of diet and cervical microbiome on the risk of cervical intraepithelial neoplasia. Clin. Nutr. 2016, 35, 1434–1441. [Google Scholar] [CrossRef]
- Suehiro, T.T.; Malaguti, N.; Damke, E.; Uchimura, N.S.; Gimenes, F.; Souza, R.P.; da Silva, V.R.S.; Consolaro, M.E.L. Association of human papillomavirus and bacterial vaginosis with increased risk of high-grade squamous intraepithelial cervical lesions. Int. J. Gynecol. Cancer 2019, 29, 242–249. [Google Scholar] [CrossRef]
- Yang, J.; Nolte, F.S.; Chajewski, O.S.; Lindsey, K.G.; Houser, P.M.; Pellicier, J.; Wang, Q.; Ehsani, L. Cytology and high risk HPV testing in cervical cancer screening program: Outcome of 3-year follow-up in an academic institute. Diagn. Cytopathol. 2018, 46, 22–27. [Google Scholar] [CrossRef]
- Chase, D.; Goulder, A.; Zenhausern, F.; Monk, B.; Herbst-Kralovetz, M. The vaginal and gastrointestinal microbiomes in gynecologic cancers: A review of applications in etiology, symptoms and treatment. Gynecol. Oncol. 2015, 138, 190–200. [Google Scholar] [CrossRef]
- Shi, P.; Zhang, L.; Ye, N. Serum metabolomic analysis of cervical cancer patients by gas chromatography-mass spectrometry. Asian J. Chem. 2015, 27, 547–551. [Google Scholar]
- Walker, H.; Burrell, M.; Flatley, J.; Powers, H. A metabolite profiling method for diagnosis of precancerous cervical lesions and HPV persistence. Bioanalysis 2017, 9, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef] [Green Version]
- Khatami, F.; Payab, M.; Sarvari, M.; Gilany, K.; Larijani, B.; Arjmand, B.; Tavangar, S.M. Oncometabolites as biomarkers in thyroid cancer: A systematic review. Cancer Manag. Res. 2019, 11, 1829–1841. [Google Scholar] [CrossRef]
- Tebani, A.; Abily-Donval, L.; Afonso, C.; Marret, S.; Bekri, S. Clinical metabolomics: The new metabolic window for inborn errors of metabolism investigations in the post-genomic era. Int. J. Mol. Sci. 2016, 17, 1167. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Quantitative metabolomics using NMR. Trends Anal. Chem 2008, 27, 228–237. [Google Scholar] [CrossRef]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef] [PubMed]
- López-López, Á.; López-Gonzálvez, Á.; Clive-Baker, T.; Barbas, C. A review of validated biomarkers obtained through metabolomics. Expert Rev. Mol. Diagn. 2018, 18, 557–575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, Y.; Zhao, W.; Deng, K.; Wang, Z.; Yang, C.; Ma, L.; Openkova, M.S.; Hou, Y.; Li, K. Metabolomics for biomarker discovery in the diagnosis, prognosis, survival and recurrence of colorectal cancer: A systematic review. Oncotarget 2017, 8, 35460–35472. [Google Scholar] [CrossRef]
- McCartney, A.; Vignoli, A.; Biganzoli, L.; Love, R.; Tenori, L.; Luchinat, C.; Di Leo, A. Metabolomics in breast cancer: A decade in review. Cancer Treat. Rev. 2018, 67, 88–96. [Google Scholar] [CrossRef]
- Huang, S.; Chong, N.; Lewis, N.E.; Jia, W.; Xie, G.; Garmire, L.X. Novel personalized pathway-based metabolomics models reveal key metabolic pathways for breast cancer diagnosis. Genome Med. 2016, 8, 34. [Google Scholar] [CrossRef]
- Lehtovirta, P.; Viinikka, L.; Ylikorkala, O. Comparison between squamous cell carcinoma-associated antigen and CA-125 in patients with carcinoma of the cervix. Gynecol. Oncol. 1990, 37, 276–278. [Google Scholar] [CrossRef]
- Jin, Y.; Kim, S.C.; Kim, H.J.; Ju, W.; Kim, Y.H.; Kim, H.-J. A lectin-based diagnostic system using circulating antibodies to detect cervical intraepithelial neoplasia and cervical cancer. Glycobiology 2015, 26, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Barrón, E.V.; Roman-Bassaure, E.; Sánchez-Sandoval, A.L.; Espinosa, A.M.; Guardado-Estrada, M.; Medina, I.; Juárez, E.; Alfaro, A.; Bermúdez, M.; Zamora, R. CDKN3 mRNA as a biomarker for survival and therapeutic target in cervical cancer. PLoS ONE 2015, 10, e0137397. [Google Scholar] [CrossRef]
- Hasim, A.; Aili, A.; Maimaiti, A.; Abudula, A.; Upur, H. Plasma-free amino acid profiling of cervical cancer and cervical intraepithelial neoplasia patients and its application for early detection. Mol. Biol. Rep. 2013, 40, 5853–5859. [Google Scholar] [CrossRef]
- Yin, M.-z.; Tan, S.; Li, X.; Hou, Y.; Cao, G.; Li, K.; Kou, J.; Lou, G. Identification of phosphatidylcholine and lysophosphatidylcholine as novel biomarkers for cervical cancers in a prospective cohort study. Tumor Biol. 2016, 37, 5485–5492. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Lucarelli, G.; Galleggiante, V.; Rutigliano, M.; Sanguedolce, F.; Cagiano, S.; Bufo, P.; Lastilla, G.; Maiorano, E.; Ribatti, D.; Giglio, A.; et al. Metabolomic profile of glycolysis and the pentose phosphate pathway identifies the central role of glucose-6-phosphate dehydrogenase in clear cell-renal cell carcinoma. Oncotarget 2015, 6, 13371–13386. [Google Scholar] [CrossRef] [Green Version]
- Dang, C.V. Links between metabolism and cancer. Genes Dev. 2012, 26, 877–890. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.S.; Thompson, C.B. Signaling in control of cell growth and metabolism. Cold Spring Harb. Perspect. Biol. 2012, 4, a006783. [Google Scholar] [CrossRef]
- Liu, W.; Hancock, C.N.; Fischer, J.W.; Harman, M.; Phang, J.M. Proline biosynthesis augments tumor cell growth and aerobic glycolysis: Involvement of pyridine nucleotides. Sci. Rep. 2015, 5, 17206. [Google Scholar] [CrossRef]
- Vissers, Y.L.; Dejong, C.H.; Luiking, Y.C.; Fearon, K.C.; von Meyenfeldt, M.F.; Deutz, N.E. Plasma arginine concentrations are reduced in cancer patients: Evidence for arginine deficiency? Am. J. Clin. Nut. 2005, 81, 1142–1146. [Google Scholar] [CrossRef]
- Cascino, A.; Muscaritoli, M.; Cangiano, C.; Conversano, L.; Laviano, A.; Ariemma, S.; Meguid, M.; Rossi, F.F. Plasma amino acid imbalance in patients with lung and breast cancer. Anticancer Res. 1995, 15, 507–510. [Google Scholar]
- Yang, K.; Xia, B.; Wang, W.; Cheng, J.; Yin, M.; Xie, H.; Li, J.; Ma, L.; Yang, C.; Li, A. A comprehensive analysis of metabolomics and transcriptomics in cervical cancer. Sci. Rep. 2017, 7, 43353. [Google Scholar] [CrossRef]
- Ye, N.; Liu, C.; Shi, P. Metabolomics analysis of cervical cancer, cervical intraepithelial neoplasia and chronic cervicitis by 1H NMR spectroscopy. Eur. J. Gynaecol. Oncol 2015, 36, 174–180. [Google Scholar]
- Zhang, Q.; Takahashi, M.; Noguchi, Y.; Sugimoto, T.; Kimura, T.; Okumura, A.; Ishikawa, T.; Kakumu, S. Plasma amino acid profiles applied for diagnosis of advanced liver fibrosis in patients with chronic hepatitis C infection. Hepatol. Res. 2006, 34, 170–177. [Google Scholar] [CrossRef]
- Maeda, J.; Higashiyama, M.; Imaizumi, A.; Nakayama, T.; Yamamoto, H.; Daimon, T.; Yamakado, M.; Imamura, F.; Kodama, K. Possibility of multivariate function composed of plasma amino acid profiles as a novel screening index for non-small cell lung cancer: A case control study. BMC Cancer 2010, 10, 690. [Google Scholar] [CrossRef]
- Mustafa, A.; Gupta, S.; Hudes, G.R.; Egleston, B.L.; Uzzo, R.G.; Kruger, W.D. Serum amino acid levels as a biomarker for renal cell carcinoma. J. Urol. 2011, 186, 1206–1212. [Google Scholar] [CrossRef]
- Mayers, J.R.; Wu, C.; Clish, C.B.; Kraft, P.; Torrence, M.E.; Fiske, B.P.; Yuan, C.; Bao, Y.; Townsend, M.K.; Tworoger, S.S. Elevation of circulating branched-chain amino acids is an early event in human pancreatic adenocarcinoma development. Nat. Med. 2014, 20, 1193–1198. [Google Scholar] [CrossRef] [Green Version]
- Kratochvilova, M.; Raudenska, M.; Heger, Z.; Richtera, L.; Cernei, N.; Adam, V.; Babula, P.; Novakova, M.; Masarik, M.; Gumulec, J. Amino acid profiling of zinc resistant prostate cancer cell lines: Associations with cancer progression. Prostate 2017, 77, 604–616. [Google Scholar] [CrossRef]
- Fan, J.; Hong, J.; Hu, J.-D.; Chen, J.-L. Ion chromatography based urine amino acid profiling applied for diagnosis of gastric cancer. Gastroenterol. Res. Pract. 2012, 2012, 474907. [Google Scholar] [CrossRef]
- Morvan, D.; Demidem, A. Metabolomics by proton nuclear magnetic resonance spectroscopy of the response to chloroethylnitrosourea reveals drug efficacy and tumor adaptive metabolic pathways. Cancer Res. 2007, 67, 2150–2159. [Google Scholar] [CrossRef]
- Odunsi, K.; Wollman, R.M.; Ambrosone, C.B.; Hutson, A.; McCann, S.E.; Tammela, J.; Geisler, J.P.; Miller, G.; Sellers, T.; Cliby, W.; et al. Detection of epithelial ovarian cancer using 1H-NMR-based metabonomics. Int. J. Cancer 2005, 113, 782–788. [Google Scholar] [CrossRef]
- Fan, T.W.-M. Metabolite profiling by one-and two-dimensional NMR analysis of complex mixtures. Prog. Nucl. Magn. Reson. Spectrosc. 1996, 28, 161–219. [Google Scholar] [CrossRef]
- Hasim, A.; Ali, M.; Mamtimin, B.; Ma, J.Q.; Li, Q.-Z.; Abudula, A. Metabonomic signature analysis of cervical carcinoma and precancerous lesions in women by 1H NMR spectroscopy. Exp. Ther. Med. 2012, 3, 945–951. [Google Scholar] [CrossRef]
- Obajimi, O.; Melera, P.W. The depletion of cellular ATP by AG2034 mediates cell death or cytostasis in a hypoxanthine-dependent manner in human prostate cancer cells. Cancer Chem. Pharmacol. 2008, 62, 215–226. [Google Scholar] [CrossRef]
- Hille, R. Molybdenum-containing hydroxylases. Arch. Biochem. Biophys. 2005, 433, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Fini, M.A.; Elias, A.; Johnson, R.J.; Wright, R.M. Contribution of uric acid to cancer risk, recurrence, and mortality. Clin. Trans. Med. 2012, 1, 16. [Google Scholar] [CrossRef]
- Spychala, J. Tumor-promoting functions of adenosine. Pharmacol. Ther. 2000, 87, 161–173. [Google Scholar] [CrossRef]
- Satoh, K.; Yachida, S.; Sugimoto, M.; Oshima, M.; Nakagawa, T.; Akamoto, S.; Tabata, S.; Saitoh, K.; Kato, K.; Sato, S.; et al. Global metabolic reprogramming of colorectal cancer occurs at adenoma stage and is induced by MYC. Proc. Natl. Acad. Sci. USA 2017, 114, E7697–E7706. [Google Scholar] [CrossRef] [Green Version]
- Sahu, D.; Lotan, Y.; Wittmann, B.; Neri, B.; Hansel, D.E. Metabolomics analysis reveals distinct profiles of nonmuscle-invasive and muscle-invasive bladder cancer. Cancer Med. 2017, 6, 2106–2120. [Google Scholar] [CrossRef]
- Ulmer, H.; Borena, W.; Rapp, K.; Klenk, J.; Strasak, A.; Diem, G.; Concin, H.; Nagel, G. Serum triglyceride concentrations and cancer risk in a large cohort study in Austria. Br. J. Cancer 2009, 101, 1202–1206. [Google Scholar] [CrossRef] [Green Version]
- Parazzini, F.; La Vecchia, C.; Negri, E.; Fasoli, M.; Cecchetti, G. Risk factors for adenocarcinoma of the cervix: A case-control study. Br. J. Cancer 1988, 57, 201–204. [Google Scholar] [CrossRef]
- Ursin, G.; Pike, M.C.; Preston-Martin, S.; d’Ablaing, G.; Peters, R.K. Sexual, reproductive, and other risk factors for adenocarcinoma of the cervix: Results from a population-based case-control study (California, United States). Cancer Causes Control 1996, 7, 391–401. [Google Scholar] [CrossRef]
- Ulmer, H.; Bjørge, T.; Concin, H.; Lukanova, A.; Manjer, J.; Hallmans, G.; Borena, W.; Häggström, C.; Engeland, A.; Almquist, M. Metabolic risk factors and cervical cancer in the metabolic syndrome and cancer project (Me–Can). Gynecol. Oncol. 2012, 125, 330–335. [Google Scholar] [CrossRef] [Green Version]
- WHO (World Health Organization) Human Papillomavirus (HPV) and Cervical Cancer. 2018. Available online: www.who.int/ (accessed on 25 January 2019).
- Lei, J.; Ploner, A.; Lagheden, C.; Eklund, C.; Kleppe, S.N.; Andrae, B.; Elfström, K.M.; Dillner, J.; Sparén, P.; Sundström, K. High-risk human papillomavirus status and prognosis in invasive cervical cancer: A nationwide cohort study. PLoS Med. 2018, 15, e1002666. [Google Scholar] [CrossRef]
- Hooi, D.J.; Lissenberg-Witte, B.I.; de Koning, M.N.; Pinedo, H.M.; Kenter, G.G.; Meijer, C.J.; Quint, W.G. High prevalence of high-risk HPV genotypes other than 16 and 18 in cervical cancers of Curaçao: Implications for choice of prophylactic HPV vaccine. Sex Transm. Infect. 2018, 94, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Han, L.; Li, X.; Gao, Q.; Pan, L.; Wu, L.; Luo, Y.; Wang, W.; Zheng, Z.; Guo, X. Prevalence and risk factors for cervical neoplasia: A cervical cancer screening program in Beijing. BMC Public Health 2014, 14, 1185. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Lee, J.K.; Kim, T.J.; Kim, M.K. The association between fruit and vegetable consumption and HPV viral load in high-risk HPV-positive women with cervical intraepithelial neoplasia. Cancer Causes Control 2010, 21, 51–59. [Google Scholar] [CrossRef]
- Carstensen, B.; Plummer, M.; Laara, E.; Hills, M. Epi: A Package for Statistical Analysis in Epidemiology, R Package Version 2.35; 2019. Available online: https://CRAN.R-project.org/package=Epi (accessed on 2 March 2019).
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Australia, 2015. [Google Scholar]
- Pohlert, T. The Pairwise Multiple Comparison of Mean Ranks Package (PMCMR). R Package. 2014. Available online: http://CRAN.R-project.org/package=PMCMR (accessed on 3 February 2019).
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 2, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Normal, n = 69 | CIN1, n = 55 | CIN2/3, n = 42 | CX CAN, n = 60 | pa | |
|---|---|---|---|---|---|---|
| Age (years) | 48 (43, 51) | 35 (31, 40) | 39.5 (33, 49) | 50 (42, 51) | <0.0001 | |
| BMI (Kg/m2) | 21.64 (20.5, 23.2) | 20.6 (19.4, 21.9) | 20.8 (19.8, 23.4) | 23.2 (20.6, 25.7) | 0.0003 | |
| HPV | Positive | 30 (43.5) | 30 (54.6) | 30 (71.4) | 47 (78.3) | 0.0002 |
| Marital status | Single | 4 (5.8) | 21 (38.2) | 7 (20) | 4 (6.67) | <0.0001 |
| Married | 59 (85.5) | 29 (52.7) | 24 (68.6) | 47 (78.3) | ||
| Divorce, bereavement | 6 (8.7) | 5 (9.1) | 4 (11.4) | 9 (15) | ||
| Education | ≤9 y | 12 (17.4) | 2 (3.6) | 6 (17.1) | 30 (50) | <0.0001 |
| 9–12 y | 24 (34.8) | 19 (34.6) | 13 (37.1) | 21 (35) | ||
| ≥12 y | 33 (47.8) | 34 (61.8) | 16 (45.7) | 9 (15.0) | ||
| Postmenopausal | Yes | 28 (40.6) | 4 (7.3) | 8 (22.9) | 34 (56.7) | <0.0001 |
| Pregnancy | No | 4 (5.8) | 21 (38.2) | 9 (25.7) | 5 (8.3) | <0.0001 |
| Yes | 65 (94.2) | 34 (61.8) | 26 (74.3) | 55 (91.7) | ||
| Oral contraceptive | Yes | 11 (15.9) | 8 (14.8) | 12 (34.3) | 9 (17.7) | 0.0949 |
| Smoking status | Yes, include past | 8 (11.8) | 18 (32.7) | 7 (20) | 7 (11.7) | 0.0097 |
| Metabolite | N vs. CIN2/3 | N vs. CX CAN | CIN1 vs. CIN2/3 | CIN1 vs. CX CAN | N+CIN1 vs. CIN2/3 + CX CAN |
|---|---|---|---|---|---|
| AMP | 0.53 | 0.71 | <0.50 | 0.68 | 0.62 |
| Aspartate | 0.80 | 0.76 | 0.71 | 0.67 | 0.74 |
| Glutamate | 0.76 | 0.81 | 0.69 | 0.73 | 0.76 |
| Hypoxanthine | 0.68 | 0.77 | 0.72 | 0.80 | 0.74 |
| Lactate | 0.74 | 0.74 | 0.67 | 0.69 | 0.71 |
| Proline | 0.68 | 0.71 | 0.51 | 0.54 | 0.62 |
| Pyroglutamate | 0.72 | 0.74 | 0.68 | 0.69 | 0.71 |
| Aspartate,Glutamate | 0.80 | 0.81 | 0.70 | 0.71 | 0.76 |
| Aspartate,Hypoxanthine | 0.82 | 0.76 | 0.73 | 0.73 | 0.73 |
| Glutamate,Hypoxanthine | 0.76 | 0.81 | 0.67 | 0.76 | 0.76 |
| Aspartate,Glutamate,Hypoxanthine | 0.81 | 0.81 | 0.73 | 0.74 | 0.75 |
| AMP,Aspartate,Glutamate,Hypoxanthine | 0.82 | 0.82 | 0.71 | 0.73 | 0.75 |
| Aspartate,Glutamate,Hypoxanthine,Lactate | 0.80 | 0.80 | 0.73 | 0.73 | 0.76 |
| Aspartate,Glutamate,Hypoxanthine,Proline | 0.82 | 0.81 | 0.72 | 0.72 | 0.75 |
| Aspartate,Glutamate,Hypoxanthine,Pyroglutamate | 0.80 | 0.81 | 0.74 | 0.76 | 0.76 |
| AMP,Aspartate,Glutamate,Hypoxanthine,Lactate | 0.81 | 0.82 | 0.71 | 0.73 | 0.75 |
| AMP,Aspartate,Glutamate,Hypoxanthine,Proline | 0.82 | 0.83 | 0.72 | 0.72 | 0.75 |
| AMP,Aspartate,Glutamate,Hypoxanthine,Pyroglutamate | 0.81 | 0.82 | 0.73 | 0.77 | 0.78 |
| Aspartate,Glutamate,Hypoxanthine,Lactate,Proline | 0.82 | 0.81 | 0.72 | 0.72 | 0.75 |
| Aspartate,Glutamate,Hypoxanthine,Lactate,Pyroglutamate | 0.81 | 0.80 | 0.74 | 0.76 | 0.77 |
| AMP,Aspartate,Glutamate,Hypoxanthine,Lactate,Proline | 0.82 | 0.83 | 0.71 | 0.72 | 0.75 |
| AMP,Aspartate,Glutamate,Hypoxanthine,Lactate,Pyroglutamate | 0.81 | 0.82 | 0.72 | 0.78 | 0.78 |
| Aspartate,Glutamate,Hypoxanthine,Lactate,Proline,Pyroglutamate | 0.82 | 0.82 | 0.73 | 0.77 | 0.78 |
| AMP,Aspartate,Glutamate,Hypoxanthine,Lactate,Proline,Pyroglutamate | 0.82 | 0.83 | 0.72 | 0.78 | 0.78 |
| Metabolite | N | CIN2/3 | N vs. CIN2/3 | CX CAN | N vs. CX CAN | CIN1 | CIN23 | CIN1 vs. CIN2/3 | CX CAN | CIN1 vs. CX CAN | N + CIN1 | CIN23 + CX CAN | N + CIN1 vs. CIN23 + CX CAN |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n = 69 | n = 42 | mOR (95% CI) | n = 60 | mOR (95% CI) | n = 55 | n = 42 | mOR (95% CI) | n = 60 | mOR (95% CI) | n = 124 | n = 102 | mOR (95% CI) | |
| AMP | |||||||||||||
| Low | 49.3 | 38.1 | ref | 25 | ref | 40.1 | 35.7 | ref | 25 | ref | 50 | 30.4 | ref |
| High | 50.7 | 61.9 | 2.11 (0.79–5.63) | 75 | 1.52 (0.54–4.28) | 50.9 | 64.3 | 4.34 (1.29–14.6) | 75 | 0.62 (0.12–3.28) | 50 | 69.6 | 2.34 (1.13–4.83) |
| Aspartate | |||||||||||||
| Low | 50.7 | 11.9 | ref | 21.7 | ref | 49.1 | 14.3 | ref | 25 | ref | 50 | 20.6 | ref |
| High | 49.3 | 88.1 | 4.31 (1.38–13.5) | 78.3 | 1.64 (0.59–4.58) | 50.9 | 85.7 | 8.92 (2.38–33.4) | 75 | 0.81 (0.15–4.24) | 50 | 79.4 | 2.80 (1.36–5.78) |
| Glutamate | |||||||||||||
| Low | 50.7 | 11.9 | ref | 8.3 | ref | 50.9 | 28.6 | ref | 20 | ref | 50.8 | 14.7 | ref |
| High | 49.3 | 88.1 | 7.99 (2.31–27.6) | 91.7 | 6.31 (1.74–22.9) | 49.1 | 71.4 | 2.68 (0.87–8.20) | 80 | 0.60 (0.10–3.71) | 49.2 | 85.3 | 4.49 (2.05–9.82) |
| Hypoxanthine | |||||||||||||
| Low | 49.3 | 23.8 | ref | 15 | ref | 50.9 | 14.3 | ref | 8.3 | ref | 50.8 | 17.7 | ref |
| High | 50.7 | 76.2 | 5.30 (1.64–17.1) | 85 | 4.33 (1.30–14.5) | 49.1 | 85.7 | 7.90 (2.00–31.3) | 91.7 | 2.36 (0.42–13.3) | 49.2 | 32.3 | 4.26 (1.95–9.28) |
| Lactate | |||||||||||||
| Low | 50.7 | 14.3 | ref | 13.3 | ref | 50.9 | 33.3 | ref | 25 | ref | 50 | 20.6 | ref |
| High | 49.3 | 85.7 | 5.18 (1.74–15.4) | 86.7 | 3.93 (1.24–12.5) | 49.1 | 66.7 | 1.98 (0.68–5.78) | 75 | 0.96 (0.17–5.50) | 50 | 79.4 | 2.30 (1.13–4.69) |
| Proline | |||||||||||||
| Low | 50.7 | 23.8 | ref | 20 | ref | 50.9 | 54.8 | ref | 53.3 | ref | 50 | 30.4 | ref |
| High | 49.3 | 76.2 | 5.13 (1.71–15.4) | 80 | 5.23 (1.50–18.3) | 49.1 | 45.2 | 1.04 (0.35–3.09) | 46.7 | 0.69 (0.12–3.94) | 50 | 69.6 | 2.12 (1.02–4.43) |
| Pyroglutamate | |||||||||||||
| Low | 50.7 | 23.8 | ref | 10 | ref | 49.1 | 23.8 | ref | 11.7 | ref | 50.8 | 16.7 | ref |
| High | 49.3 | 76.2 | 2.94 (1.10–7.84) | 90 | 8.02 (2.13–30.3) | 50.9 | 76.2 | 5.46 (1.63–18.4) | 88.3 | 4.54 (0.65–32.0) | 49.2 | 83.3 | 4.14 (1.92–8.92) |
| Metabolite | HPV | N vs. CIN2/3 | N vs. CX CAN | CIN1 vs. CIN2/3 | CIN1 vs. CX CAN | N+CIN1 vs. CIN2/3 + CX CAN |
|---|---|---|---|---|---|---|
| mOR (95% CI) | mOR (95% CI) | mOR (95% CI) | mOR (95% CI) | mOR (95% CI) | ||
| AMP | ||||||
| Low | Neg | ref | ref | ref | ref | ref |
| Low | Pos | 6.56 (0.99–43.2) | 5.36 (0.99–29.0) | 5.32 (0.79–35.9) | 5.31 (0.98–28.8) | 3.36 (1.02–11.1) |
| High | Neg | 1.90 (0.27–13.3) | 0.87 (0.12–6.06) | 1.82 (0.26–12.8) | 0.85 (0.12–5.94) | 1.24 (0.32–4.82) |
| High | Pos | 12.19 (2.21–67.1) | 8.54 (1.63–44.8) | 12.78 (2.31–70.7) | 8.46 (1.61–44.6) | 9.33 (2.94–29.6) |
| p interaction 1 | 0.0022 | 0.0184 | 0.0008 | 0.0184 | <0.0001 | |
| p trend 2 | 0.0065 | 0.0489 | 0.0034 | 0.0513 | 0.0002 | |
| Aspartate | ||||||
| Low | Neg | ref | ref | ref | ref | ref |
| Low | Pos | 2.52 (0.31–20.5) | 3.58 (0.67–19.2) | 3.45 (0.47–25.3) | 4.67 (0.93–23.4) | 2.47 (0.73–8.34) |
| High | Neg | 1.64 (0.24–11.2) | 0.46 (0.06–3.46) | 1.74 (0.25–12.0) | 0.50 (0.07–3.76) | 1.08 (0.28–4.13) |
| High | Pos | 13.93 (2.62–74.0) | 7.71 (1.60–37.1) | 13.52 (2.55–71.7) | 6.97 (1.45–33.4) | 9.15 (2.96–28.3) |
| p interaction | <0.0001 | 0.0067 | 0.0002 | 0.0236 | <0.0001 | |
| p trend | 0.0009 | 0.041 | 0.0013 | 0.0802 | <0.0001 | |
| Glutamate | ||||||
| Low | Neg | ref | ref | ref | ref | ref |
| Low | Pos | 0.98 (0.11–9.18) | 1.14 (0.11–12.3) | 3.62 (0.69–18.9) | 3.28 (0.60–18.0) | 2.76 (0.68–11.3) |
| High | Neg | 1.49 (0.20–11.3) | 1.33 (0.16–10.7) | 1.51 (0.21–10.6) | 0.92 (0.12–7.06) | 2.29 (0.56–9.40) |
| High | Pos | 27.18 (4.23–175) | 24.52 (3.61–167) | 17.01 (3.38–85.6) | 14.33 (2.64–77.8) | 14.85 (4.27–51.7) |
| p interaction | <0.0001 | <0.0001 | 0.0002 | 0.001 | <0.0001 | |
| p trend | <0.0001 | 0.0005 | 0.0006 | 0.0047 | <0.0001 | |
| Hypoxanthine | ||||||
| Low | Neg | ref | ref | ref | ref | ref |
| Low | Pos | 7.59 (0.93–61.9) | 3.35 (0.45–24.9) | 46.45 (2.11–999) | 10.83 (0.73–161) | 2.11 (0.56–8.04) |
| High | Neg | 5.25 (0.61–45.3) | 1.82 (0.24–13.6) | 9.76 (0.70–135) | 1.27 (0.18–9.21) | 1.58 (0.42–6.05) |
| High | Pos | 21.89 (3.36–142) | 14.88 (2.57–86.2) | 33.48 (2.83–396) | 8.19 (1.44–46.6) | 11.18 (3.55–35.2) |
| p interaction | 0.0005 | 0.0009 | 0.003 | 0.0076 | <0.0001 | |
| p trend | 0.0006 | 0.0023 | 0.0024 | 0.0216 | <0.0001 | |
| Lactate | ||||||
| Low | Neg | ref | ref | ref | ref | ref |
| Low | Pos | 2.33 (0.31–17.8) | 5.35 (0.69–41.7) | 10.40 (1.80–60.0) | 3.30 (0.71–15.4) | 4.24 (1.23–14.6) |
| High | Neg | 1.86 (0.27–12.8) | 2.79 (0.36–21.4) | 3.57 (0.50–25.3) | 0.34 (0.03–3.8) | 1.63 (0.42–6.36) |
| High | Pos | 17.99 (3.31–97.8) | 26.84 (3.69–196) | 16.54 (2.86–95.7) | 12.01 (2.35–61.3) | 9.93 (3.05–32.4) |
| p interaction | <0.0001 | 0.0006 | 0.0105 | 0.0022 | <0.0001 | |
| p trend | 0.0002 | 0.0013 | 0.0058 | 0.0139 | 0.0003 | |
| Proline 3 | ||||||
| Low | Neg | ref | ref | ref | ref | ref |
| Low | Pos | – | 4.16 (0.52–33.5) | 6.61 (1.53–28.6) | 4.83 (1.07–21.8) | 7.65 (1.95–29.9) |
| High | Neg | – | 2.79 (0.35–22.2) | 4.29 (0.63–29.2) | 1.38 (0.18–10.7) | 3.10 (0.73–13.2) |
| High | Pos | - | 34.22 (3.96–296) | 66.90 (8.82–507) | 22.04 (3.60–135) | 13.46 (3.51–51.7) |
| p interaction | - | 0.0003 | 0.0004 | 0.0028 | <.0001 | |
| p trend | - | 0.0009 | 0.0001 | 0.0029 | <.0001 | |
| Pyroglutamate | ||||||
| Low | Neg | ref | ref | ref | ref | ref |
| Low | Pos | 6.76 (0.92–49.9) | 3.81 (0.39–37.7) | 6.90 (0.94–50.5) | 6.71 (0.75–60.2) | 4.23 (1.07–16.8) |
| High | Neg | 2.68 (0.34–21.0) | 4.59 (0.56–37.4) | 2.83 (0.36–22.0) | 4.76 (0.60–37.7) | 3.15 (0.77–12.9) |
| High | Pos | 15.50 (2.79–86.0) | 27.93 (4.03–194) | 15.79 (2.86–87.1) | 22.76 (3.47–149) | 16.17 (4.55–57.5) |
| p interaction | 0.001 | 0.0004 | 0.001 | 0.0013 | <0.0001 | |
| p trend | 0.0019 | 0.0005 | 0.0017 | 0.0011 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, I.; Nam, M.; Kwon, M.; Seo, S.-s.; Jung, S.; Han, J.S.; Hwang, G.-S.; Kim, M.K. LC/MS-Based Polar Metabolite Profiling Identified Unique Biomarker Signatures for Cervical Cancer and Cervical Intraepithelial Neoplasia Using Global and Targeted Metabolomics. Cancers 2019, 11, 511. https://doi.org/10.3390/cancers11040511
Khan I, Nam M, Kwon M, Seo S-s, Jung S, Han JS, Hwang G-S, Kim MK. LC/MS-Based Polar Metabolite Profiling Identified Unique Biomarker Signatures for Cervical Cancer and Cervical Intraepithelial Neoplasia Using Global and Targeted Metabolomics. Cancers. 2019; 11(4):511. https://doi.org/10.3390/cancers11040511
Chicago/Turabian StyleKhan, Imran, Miso Nam, Minji Kwon, Sang-soo Seo, Sunhee Jung, Ji Soo Han, Geum-Sook Hwang, and Mi Kyung Kim. 2019. "LC/MS-Based Polar Metabolite Profiling Identified Unique Biomarker Signatures for Cervical Cancer and Cervical Intraepithelial Neoplasia Using Global and Targeted Metabolomics" Cancers 11, no. 4: 511. https://doi.org/10.3390/cancers11040511