Cord-Blood Lipidome in Progression to Islet Autoimmunity and Type 1 Diabetes


 , ,
, , 
Abstract
:1. Introduction
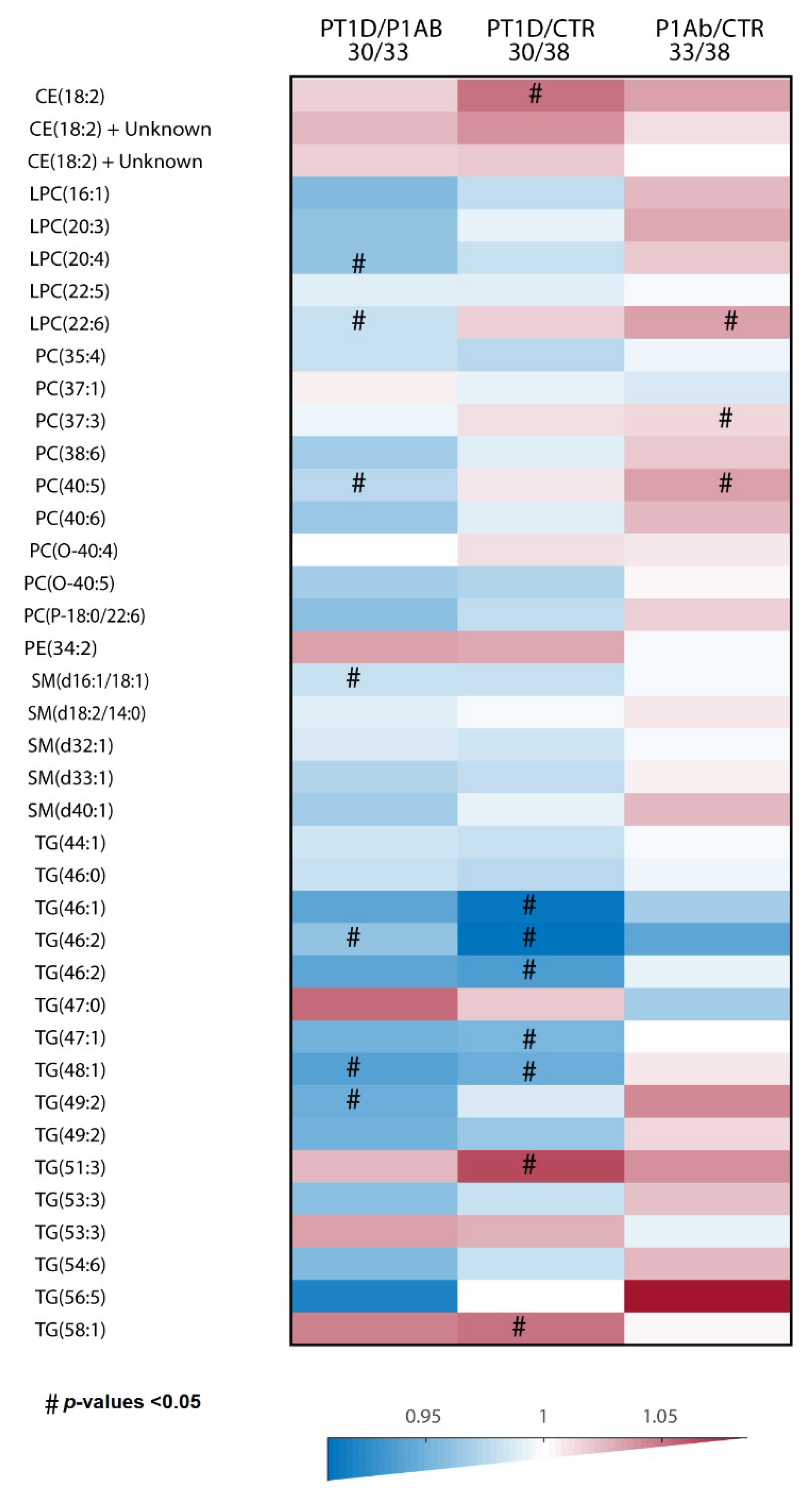
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Design and Sampling
4.2. Islet-Cell Autoimmunity Determination
4.3. Sample Preparation and Ultra High Performance Liquid Chromatography-Mass Spectrometer Analysis
4.4. Data Pre-Processing
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Katsarou, A.; Gudbjörnsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, Å. Type 1 diabetes mellitus. Nat. Rev. Dis. Primers 2017, 3, 17016. [Google Scholar] [CrossRef] [PubMed]
- Patterson, C.C.; Dahlquist, G.G.; Gyürüs, E.; Green, A.; Soltész, G.; EURODIAB Study Group. Incidence trends for childhood type 1 diabetes in Europe during 1989-2003 and predicted new cases 2005-20: A multicentre prospective registration study. Lancet 2009, 373, 2027–2033. [Google Scholar] [CrossRef]
- Achenbach, P.; Bonifacio, E.; Koczwara, K.; Ziegler, A.G. Natural history of type 1 diabetes. Diabetes 2005, 54 (Suppl. 2), S25–S31. [Google Scholar] [CrossRef] [PubMed]
- Knip, M.; Siljander, H. The role of the intestinal microbiota in type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2016, 12, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Knip, M.; Virtanen, S.M.; Akerblom, H.K. Infant feeding and the risk of type 1 diabetes. Am. J. Clin. Nutr. 2010, 91, 1506s–1513s. [Google Scholar] [CrossRef] [PubMed]
- Siljander, H.T.; Simell, S.; Hekkala, A.; Lähde, J.; Simell, T.; Vähäsalo, P.; Veijola, R.; Ilonen, J.; Simell, O.; Knip, M. Predictive characteristics of diabetes-associated autoantibodies among children with HLA-conferred disease susceptibility in the general population. Diabetes 2009, 58, 2835–2842. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, S.; Ahonen, L.; Dyrlund, T.S.; Kemppainen, E.; Siljander, H.; Hyoty, H.; Ilonen, J.; Toppari, J.; Veijola, R.; Hyotylainen, T.; et al. Dynamics of Plasma Lipidome in Progression to Islet Autoimmunity and Type 1 Diabetes—Type 1 Diabetes Prediction and Prevention Study (DIPP). Sci. Rep. 2018, 8, 10635. [Google Scholar] [CrossRef]
- Oresic, M.; Simell, S.; Sysi-Aho, M.; Näntö-Salonen, K.; Seppänen-Laakso, T.; Parikka, V.; Katajamaa, M.; Hekkala, A.; Mattila, I.; Keskinen, P.; et al. Dysregulation of lipid and amino acid metabolism precedes islet autoimmunity in children who later progress to type 1 diabetes. J. Exp. Med. 2008, 205, 2975–2984. [Google Scholar] [CrossRef] [Green Version]
- Oresic, M.; Gopalacharyulu, P.; Mykkänen, J.; Lietzen, N.; Mäkinen, M.; Nygren, H.; Simell, S.; Simell, V.; Hyöty, H.; Veijola, R.; et al. Cord serum lipidome in prediction of islet autoimmunity and type 1 diabetes. Diabetes 2013, 62, 3268–3274. [Google Scholar] [CrossRef]
- La Torre, D.; Seppänen-Laakso, T.; Larsson, H.E.; Hyötyläinen, T.; Ivarsson, S.A.; Lernmark, Å.; Orešič, M.; DiPiS Study Group. Decreased cord-blood phospholipids in young age-at-onset type 1 diabetes. Diabetes 2013, 62, 3951–3956. [Google Scholar] [CrossRef]
- Lecerf, J.; Fouilland, L.; Gagniarre, J. Evidence for a high activity of sphingomyelin biosynthesis by phosphocholine transfer from phosphatidylcholine to ceramides in lung lamellar bodies. Biochim. Biophys. Acta 1987, 918, 48–59. [Google Scholar] [CrossRef]
- Eppler, C.M.; Malewicz, B.; Jenkin, H.M.; Baumann, W.J. Phosphatidylcholine as the choline donor in sphingomyelin synthesis. Lipids 1987, 22, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. Dietary choline deficiency causes DNA strand breaks and alters epigenetic marks on DNA and histones. Mutat. Res. 2012, 733, 34–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, E. Lipid metabolism in pregnancy and its consequences in the fetus and newborn. Endocrine 2002, 19, 43–55. [Google Scholar] [CrossRef]
- Garner, S.C.; Mar, M.H.; Zeisel, S.H. Choline distribution and metabolism in pregnant rats and fetuses are influenced by the choline content of the maternal diet. J. Nutr. 1995, 125, 2851–2858. [Google Scholar] [PubMed]
- Yamaguchi, K.; Yang, L.; McCall, S.; Huang, J.; Yu, X.X.; Pandey, S.K.; Bhanot, S.; Monia, B.P.; Li, Y.X.; Diehl, A.M. Inhibiting triglyceride synthesis improves hepatic steatosis but exacerbates liver damage and fibrosis in obese mice with nonalcoholic steatohepatitis. Hepatology 2007, 45, 1366–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivera, A.; Rivera, J. Sphingolipids and the balancing of immune cell function: Lessons from the mast cell. J. Immunol. 2005, 174, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Iannello, S.; Cavaleri, A.; Milazzo, P.; Cantarella, S.; Belfiore, F. Low fasting serum triglyceride level as a precocious marker of autoimmune disorders. MedGenMed 2003, 5, 20. [Google Scholar] [PubMed]
- Haller, M.J.; Schatz, D.A. The DIPP project: 20 years of discovery in type 1 diabetes. Pediatr. Diabetes 2016, 17 (Suppl. 22), 5–7. [Google Scholar] [CrossRef]
- Knip, M.; Virtanen, S.M.; Seppä, K.; Ilonen, J.; Savilahti, E.; Vaarala, O.; Reunanen, A.; Teramo, K.; Hämäläinen, A.M.; Paronen, J.; et al. Dietary intervention in infancy and later signs of beta-cell autoimmunity. N. Engl. J. Med. 2010, 363, 1900–1908. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| PT1D (n = 30) | P1Ab (n = 33) | CTR (n = 38) | |
|---|---|---|---|
| Gender (girls, boys) | (12, 18) | (12, 21) | (14, 24) |
| Age at time of diagnosis (years) (mean ± SE) | 4.94 ± 0.56 | NA | NA |
| Age at time of first seroconversion (years) (mean ± SE) | 1.28 ± 0.11 | 3.03 ± 0.45 | NA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamichhane, S.; Ahonen, L.; Sparholt Dyrlund, T.; Dickens, A.M.; Siljander, H.; Hyöty, H.; Ilonen, J.; Toppari, J.; Veijola, R.; Hyötyläinen, T.; et al. Cord-Blood Lipidome in Progression to Islet Autoimmunity and Type 1 Diabetes. Biomolecules 2019, 9, 33. https://doi.org/10.3390/biom9010033
Lamichhane S, Ahonen L, Sparholt Dyrlund T, Dickens AM, Siljander H, Hyöty H, Ilonen J, Toppari J, Veijola R, Hyötyläinen T, et al. Cord-Blood Lipidome in Progression to Islet Autoimmunity and Type 1 Diabetes. Biomolecules. 2019; 9(1):33. https://doi.org/10.3390/biom9010033
Chicago/Turabian StyleLamichhane, Santosh, Linda Ahonen, Thomas Sparholt Dyrlund, Alex M. Dickens, Heli Siljander, Heikki Hyöty, Jorma Ilonen, Jorma Toppari, Riitta Veijola, Tuulia Hyötyläinen, and et al. 2019. "Cord-Blood Lipidome in Progression to Islet Autoimmunity and Type 1 Diabetes" Biomolecules 9, no. 1: 33. https://doi.org/10.3390/biom9010033