The Anti-Cancer Effect of Mangifera indica L. Peel Extract is Associated to γH2AX-mediated Apoptosis in Colon Cancer Cells

 , ,
, ,  ,
,  , ,
, ,  and
and 
Abstract
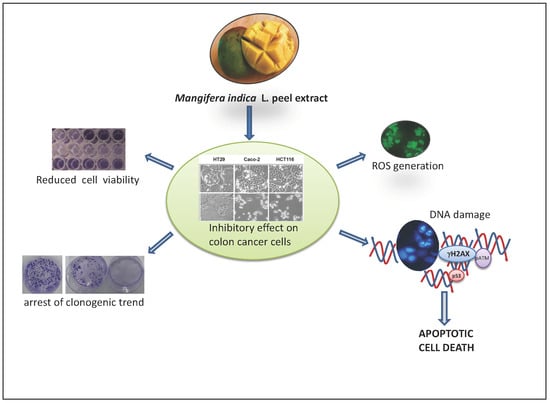
:
1. Introduction
2. Materials and Methods
2.1. Preparation of Mango Peel Extract
2.2. Reversed Phase HPLC/MS Experiments
2.3. Cell Cultures and Compounds
2.4. Cell Viability Assessment
2.5. Clonogenic Assay
2.6. Analysis of DNA Damage and Cell Cycle Distribution
2.7. Acridine Orange and Ethidium Bromide Double Staining for the Detection of Apoptosis
2.8. Assessment of Intracellular Generation of Reactive Oxygen Species
2.9. Western Blotting Analysis
2.10. Statistical Analysis
3. Results
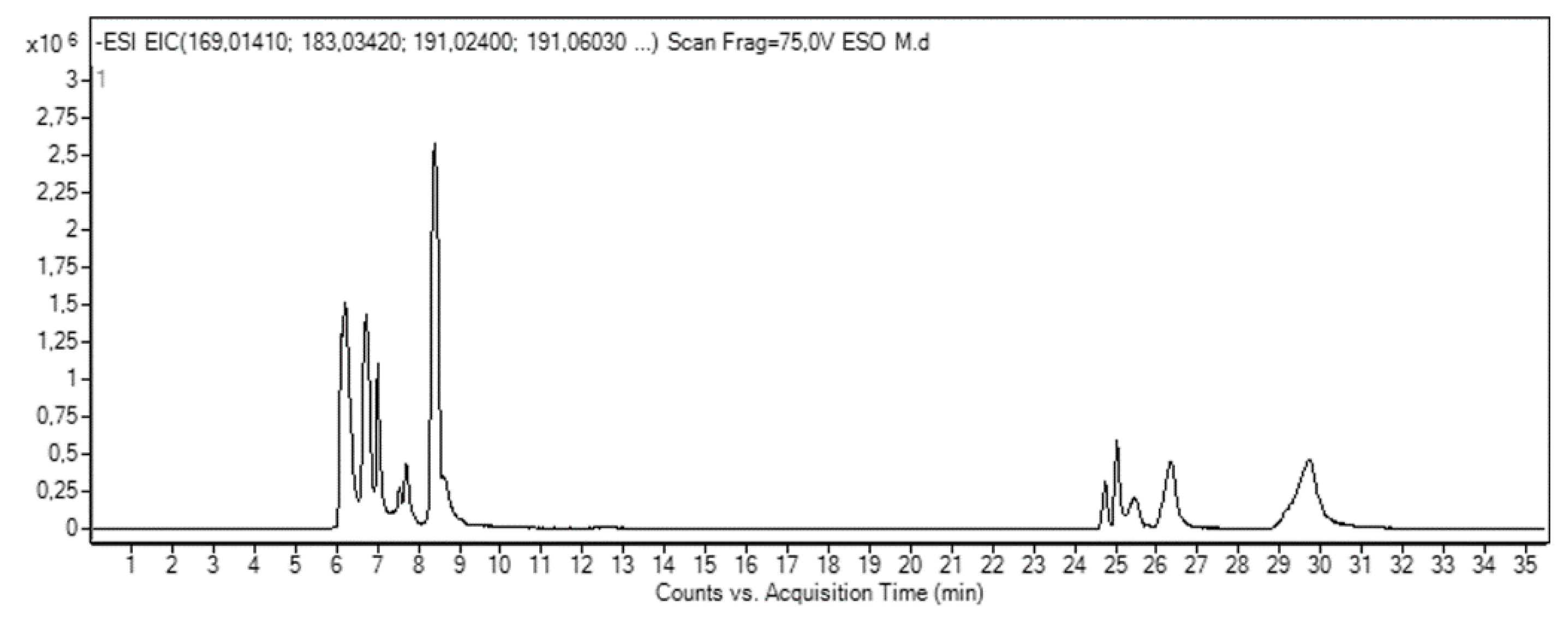
3.1. HPLC-ESI-QTOF-MS Analysis Reveals the Composition of Mango Peel Extract (MPE)
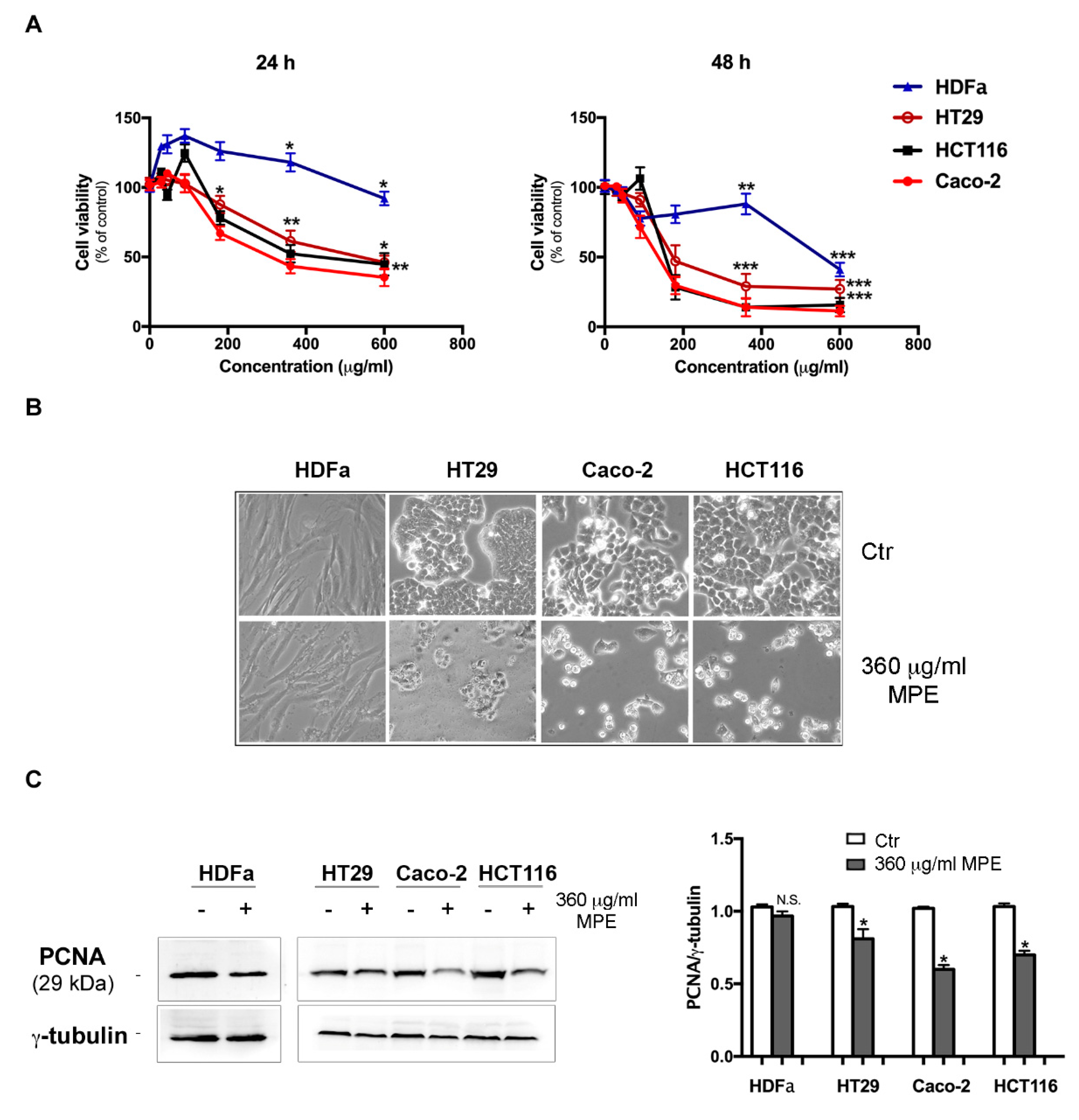
3.2. MPE Induces Cytotoxic Effects and Morphological Changes in Colon Cancer Cells
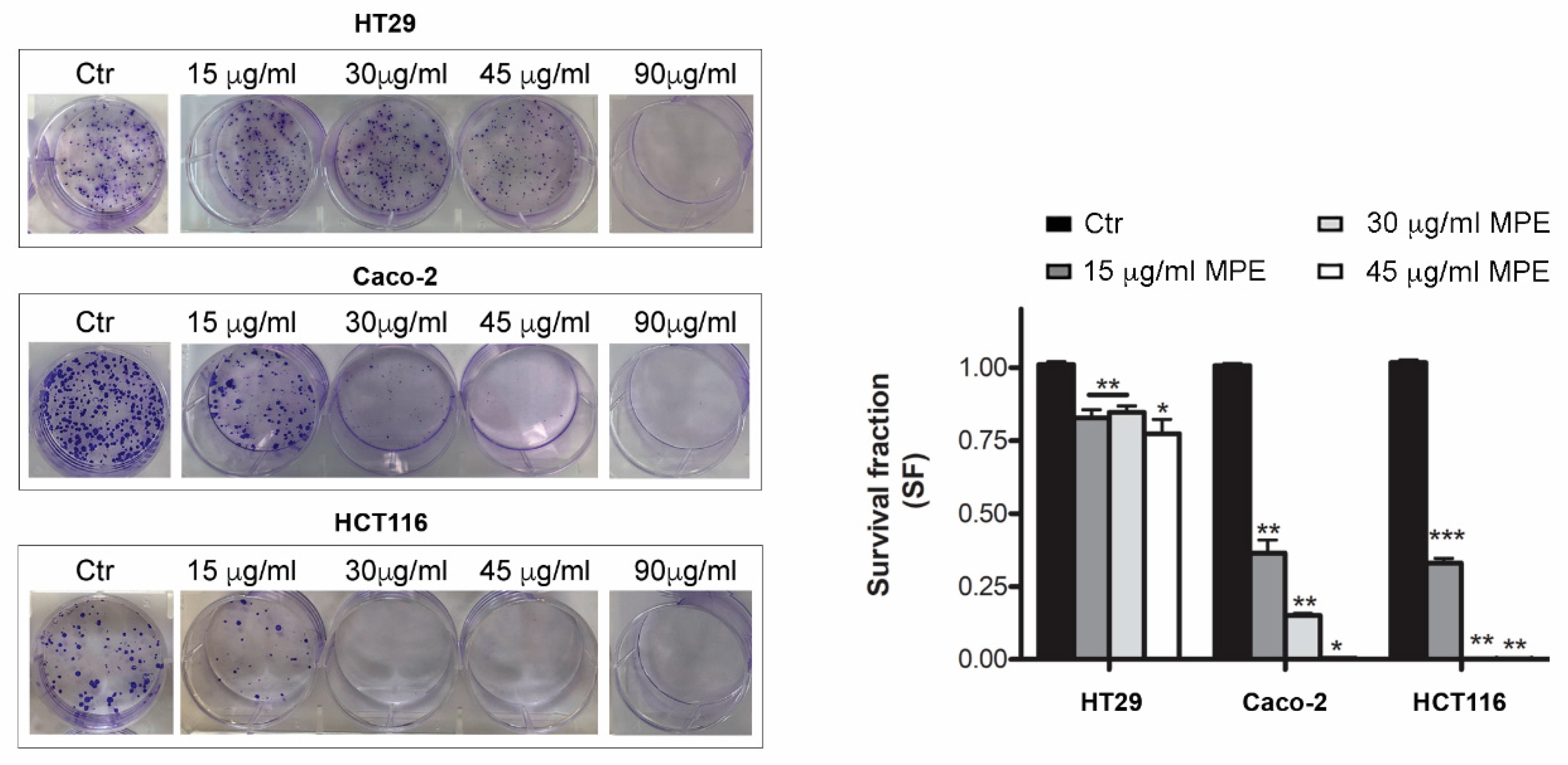
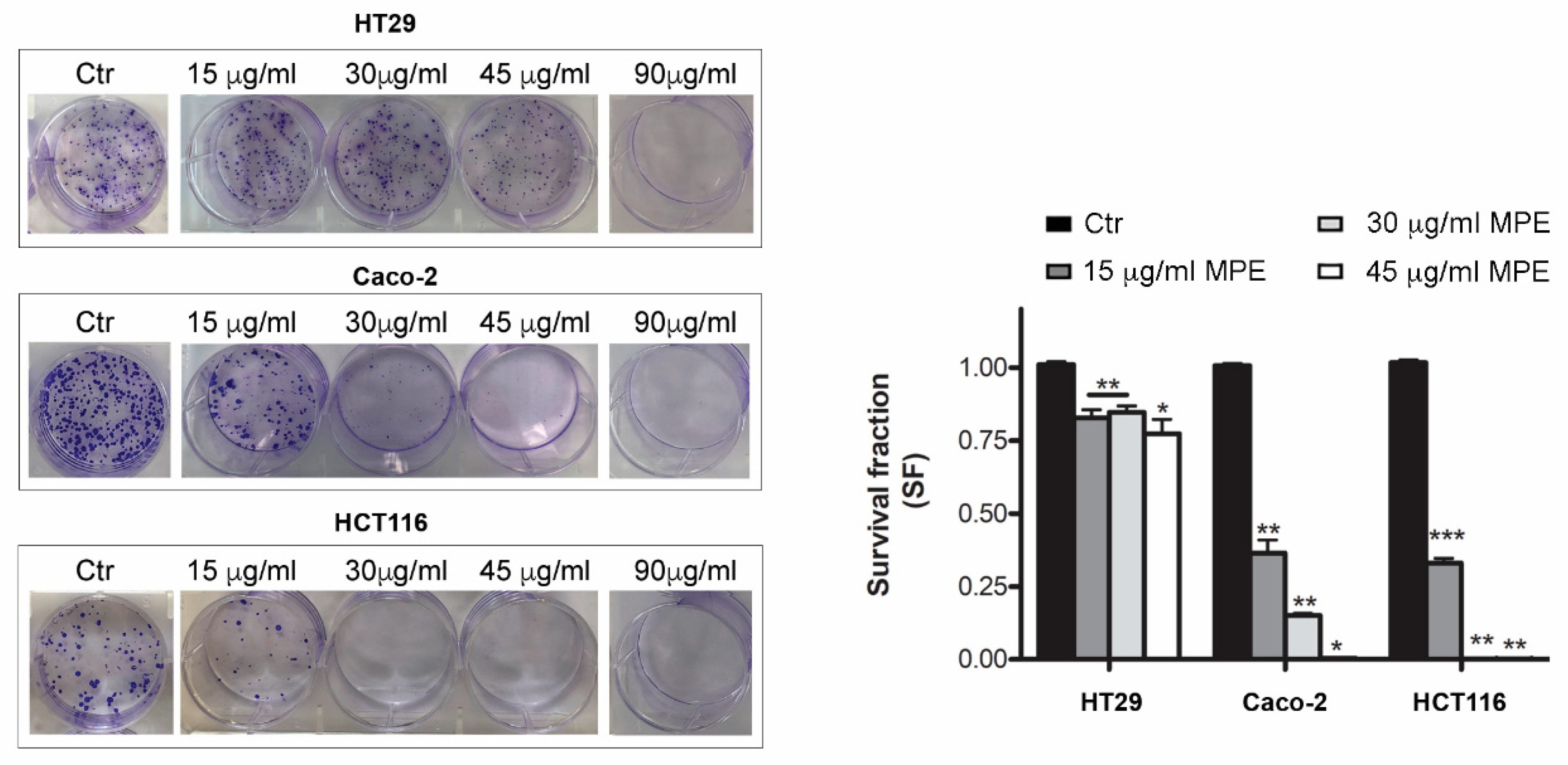
3.3. MPE Treatment Inhibits Clonogenic Ability of Colon Cancer Cells
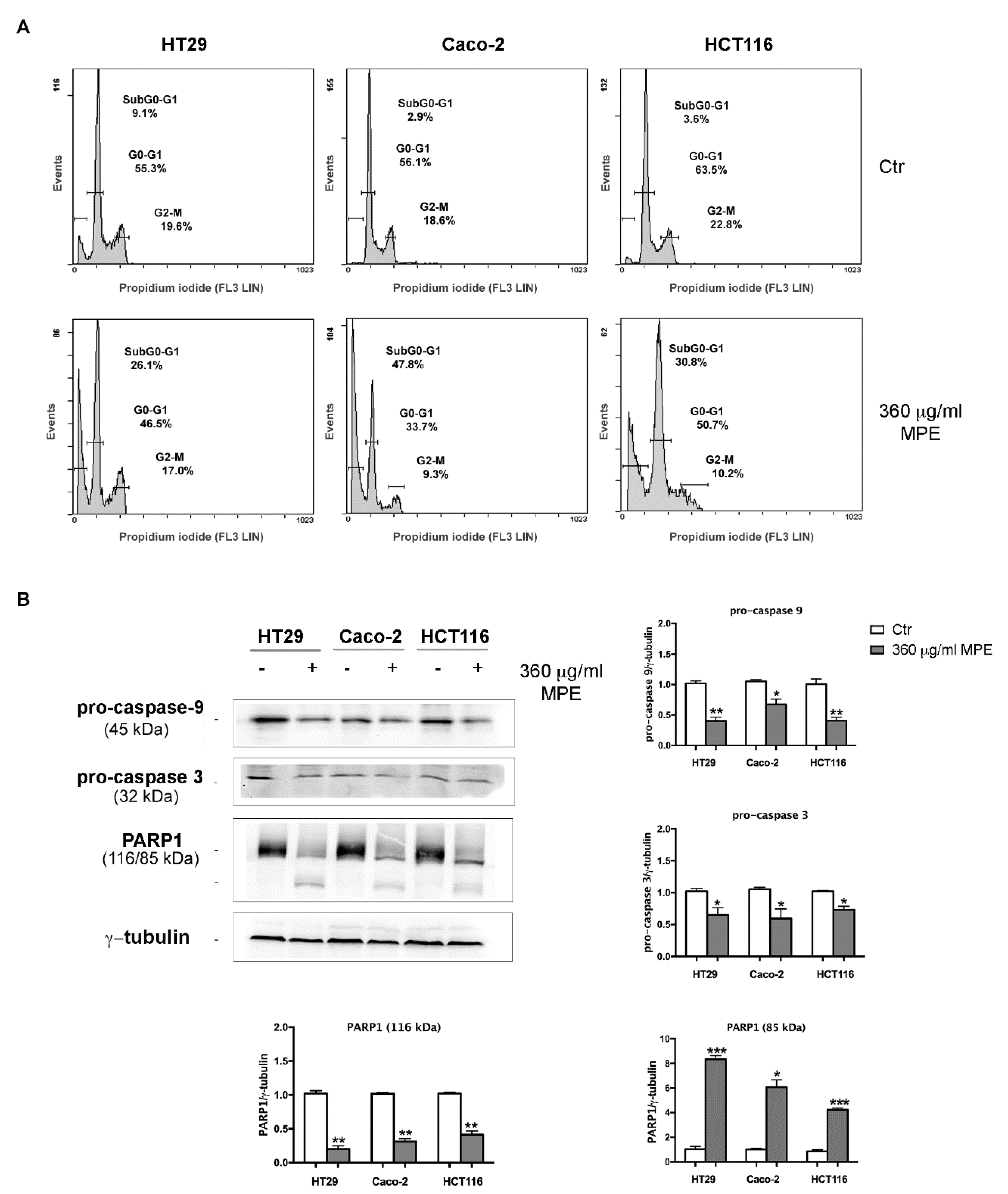
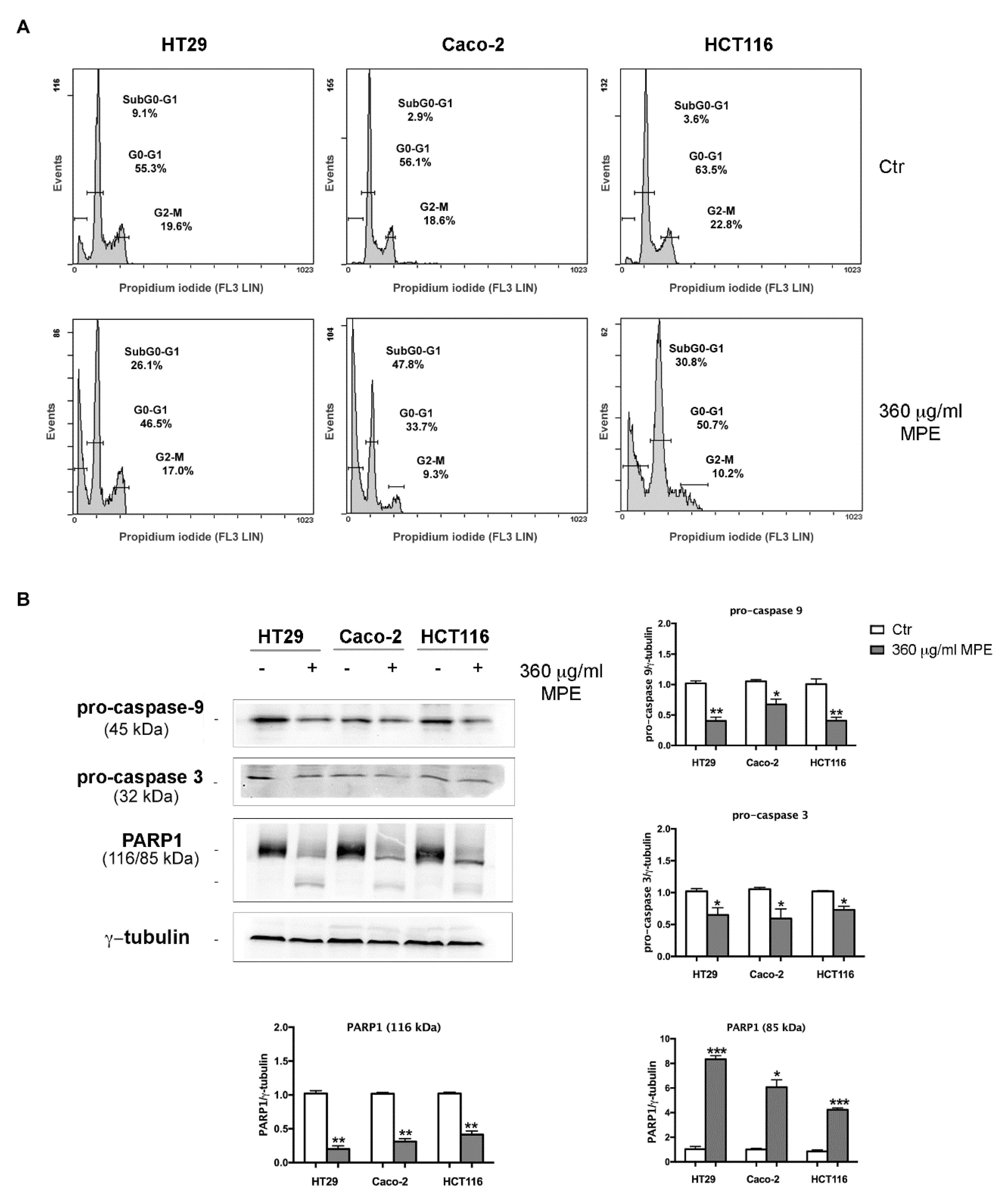
3.4. MPE Triggers DNA Fragmentation and Apoptotic Cell Death in Colon Cancer Cells
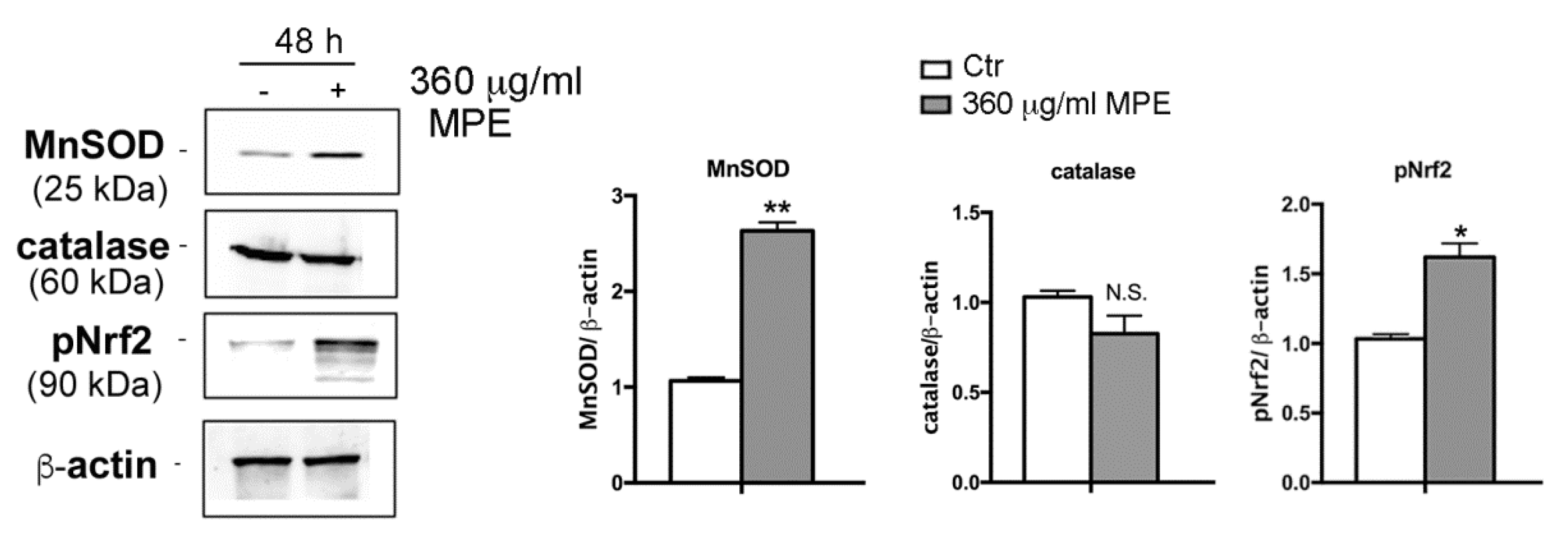
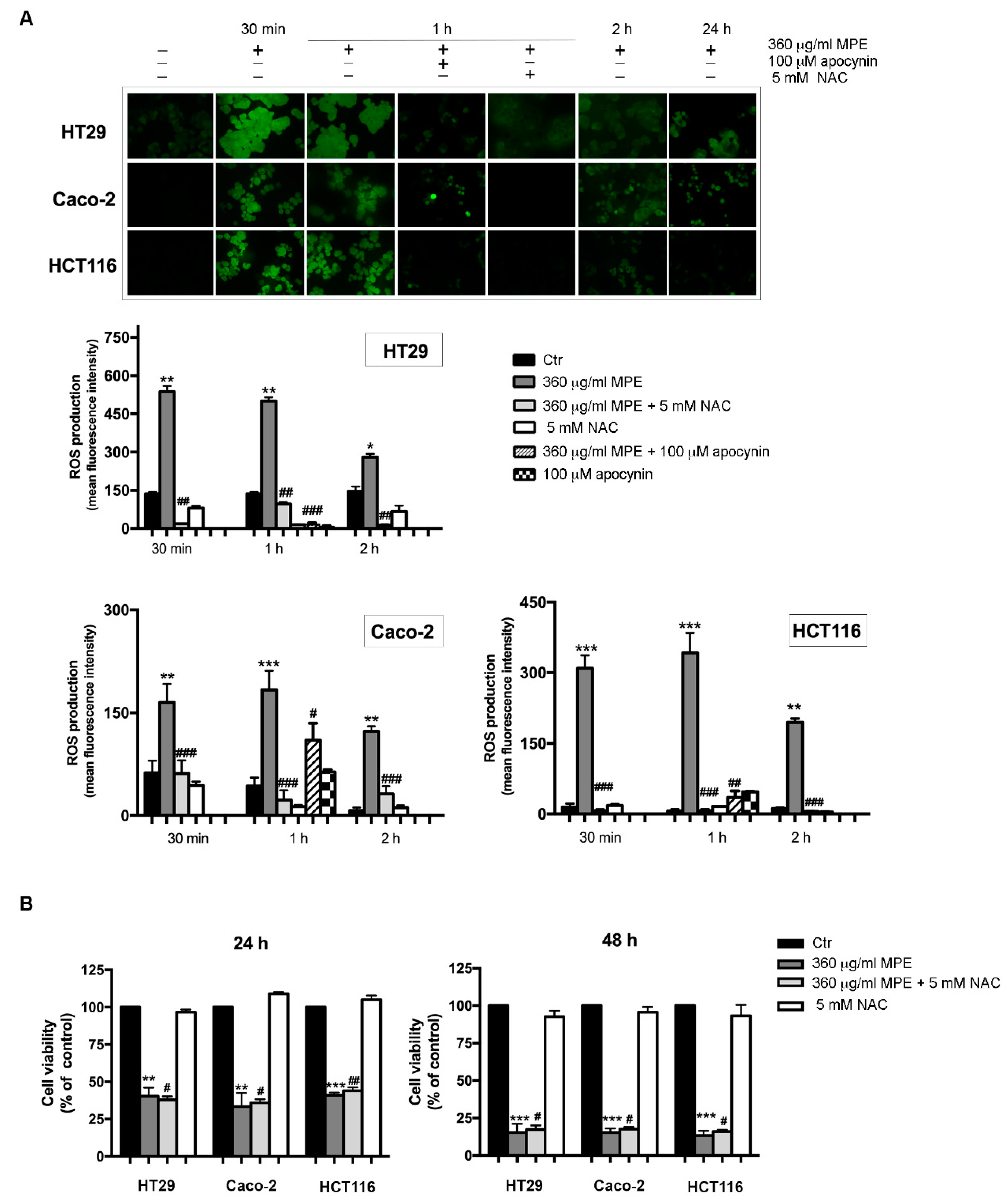
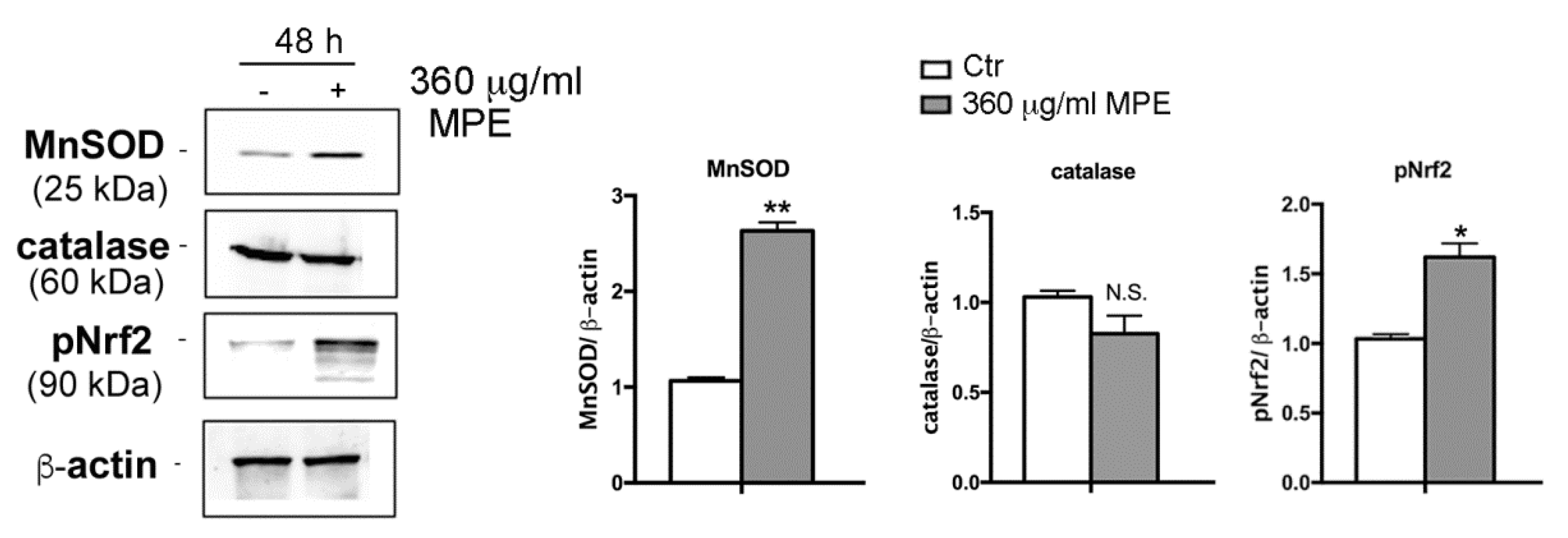
3.5. Assessment of Oxidative Stress Mediated Signaling in MPE-Treated Cells
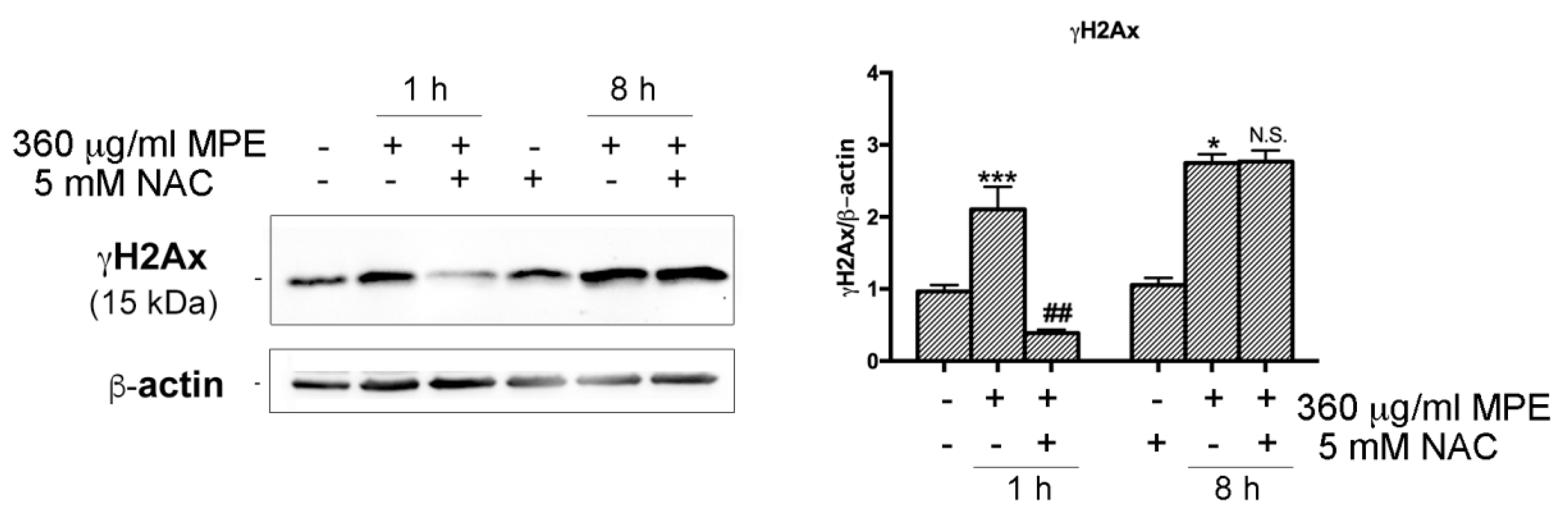
3.6. DNA Fragmentation and Apoptotic Cell Death Induced by MPE is Related to γH2AX-Mediated Genotoxic Stress
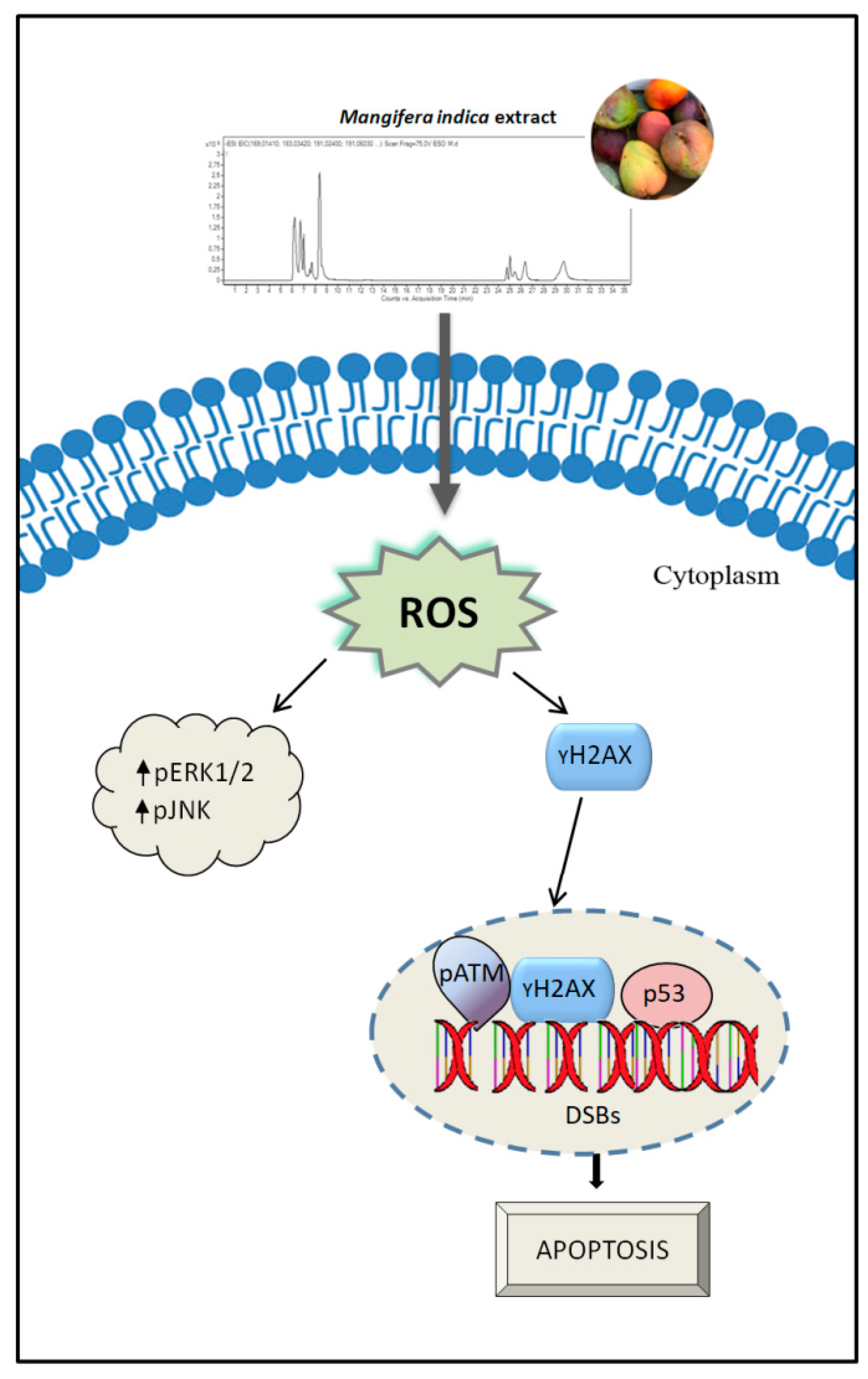
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MPE | Mango peel extract |
| HPLC/MS | High performance liquid chromatography/mass spectrometry |
| ROS | Reactive oxygen species |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| HDFa | human dermal fibroblasts |
| H2DCFDA | 5-(and-6)-carboxy-2′,7′-dichlorodihydrofluorescein diacetate |
| γH2AX | phosphorylated histone 2AX |
| pERK1/2 | phosphorylated extracellular signal-regulated protein kinases 1 and 2 |
References
- Battino, M.; Forbes-Hernández, T.Y.; Gasparrini, M.; Afrin, S.; Cianciosi, D.; Zhang, J.; Manna, P.P.; Reboredo-Rodríguez, P.; Varela Lopez, A.; Quiles, J.L.; et al. Relevance of functional foods in the Mediterranean diet: The role of olive oil, berries and honey in the prevention of cancer and cardiovascular diseases. Crit. Rev. Food Sci. Nutr. 2018, 1–28. [Google Scholar] [CrossRef]
- Ullah, R.; Khan, M.; Shah, S.A.; Saeed, K.; Kim, M.O. Natural Antioxidant Anthocyanins—A Hidden Therapeutic Candidate in Metabolic Disorders with Major Focus in Neurodegeneration. Nutrients 2019, 11, 1195. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, M.; Wawrzyniak, D.; Rolle, K.; Chomczyński, P.; Oziewicz, S.; Jurga, S.; Barciszewski, J. Let food be your medicine: Nutraceutical properties of lycopene. Food Funct. 2019, 10, 3090–3102. [Google Scholar] [CrossRef]
- Emanuele, S.; Lauricella, M.; Calvaruso, G.; D’Anneo, A.; Giuliano, M. Litchi chinensis as a Functional Food and a Source of Antitumor Compounds: An Overview and a Description of Biochemical Pathways. Nutrients 2017, 9, 992. [Google Scholar] [CrossRef] [PubMed]
- Jahurul, M.H.A.; Zaidul, I.S.M.; Ghafoor, K.; Al-Juhaimi, F.Y.; Nyam, K.-L.; Norulaini, N.A.N.; Sahena, F.; Mohd Omar, A.K. Mango (Mangifera indica L.) by-products and their valuable components: A review. Food Chem. 2015, 183, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Lauricella, M.; Emanuele, S.; Calvaruso, G.; Giuliano, M.; D’Anneo, A. Multifaceted Health Benefits of Mangifera indica L. (Mango): The Inestimable Value of Orchards Recently Planted in Sicilian Rural Areas. Nutrients 2017, 9, 525. [Google Scholar] [CrossRef] [PubMed]
- Testa, R.; Tudisca, S.; Schifani, G.; Di Trapani, A.; Migliore, G. Tropical Fruits as an Opportunity for Sustainable Development in Rural Areas: The Case of Mango in Small-Sized Sicilian Farms. Sustainability 2018, 10, 1436. [Google Scholar] [CrossRef]
- Emanuele, S.; Notaro, A.; Palumbo Piccionello, A.; Maggio, A.; Lauricella, M.; D’Anneo, A.; Cernigliaro, C.; Calvaruso, G.; Giuliano, M. Sicilian Litchi Fruit Extracts Induce Autophagy versus Apoptosis Switch in Human Colon Cancer Cells. Nutrients 2018, 10, 1490. [Google Scholar] [CrossRef]
- Berardini, N.; Carle, R.; Schieber, A. Characterization of gallotannins and benzophenone derivatives from mango (Mangifera indica L. cv. “Tommy Atkins”) peels, pulp and kernels by high-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 2208–2216. [Google Scholar] [CrossRef]
- Gold-Smith, F.; Fernandez, A.; Bishop, K. Mangiferin and Cancer: Mechanisms of Action. Nutrients 2016, 8, 396. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, B.; Li, S.; Zeng, L.; Chen, Y.; Fang, J. Mangiferin increases Nrf2 protein stability by inhibiting its ubiquitination and degradation in human HL60 myeloid leukemia cells. Int. J. Mol. Med. 2014, 33, 1348–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajeshkumar, S.; Kumar, S.V.; Ramaiah, A.; Agarwal, H.; Lakshmi, T.; Roopan, S.M. Biosynthesis of zinc oxide nanoparticles using Mangifera indica leaves and evaluation of their antioxidant and cytotoxic properties in lung cancer (A549) cells. Enzyme Microb. Technol. 2018, 117, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Noratto, G.D.; Bertoldi, M.C.; Krenek, K.; Talcott, S.T.; Stringheta, P.C.; Mertens-Talcott, S.U. Anticarcinogenic Effects of Polyphenolics from Mango (Mangifera indica) Varieties. J. Agric. Food Chem. 2010, 58, 4104–4112. [Google Scholar] [CrossRef] [PubMed]
- Corrales-Bernal, A.; Amparo Urango, L.; Rojano, B.; Maldonado, M.E. In vitro and in vivo effects of mango pulp (Mangifera indica cv. Azucar) in colon carcinogenesis. Arch. Latinoam. Nutr. 2014, 64, 16–23. [Google Scholar] [PubMed]
- Prasad, S.; Kalra, N.; Shukla, Y. Induction of Apoptosis by Lupeol and Mango Extract in Mouse Prostate and LNCaP Cells. Nutr. Cancer 2007, 60, 120–130. [Google Scholar] [CrossRef]
- Banerjee, N.; Kim, H.; Krenek, K.; Talcott, S.T.; Mertens-Talcott, S.U. Mango polyphenolics suppressed tumor growth in breast cancer xenografts in mice: Role of the PI3K/AKT pathway and associated microRNAs. Nutr. Res. 2015, 35, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Helleday, T.; Petermann, E.; Lundin, C.; Hodgson, B.; Sharma, R.A. DNA repair pathways as targets for cancer therapy. Nat. Rev. Cancer 2008, 8, 193–204. [Google Scholar] [CrossRef]
- Raffa, D.; Plescia, F.; Maggio, B.; Raimondi, M.V.; D’Anneo, A.; Lauricella, M.; Daidone, G. Anthranilamide-based 2-phenylcyclopropane-1-carboxamides, 1,1′-biphenyl-4-carboxamides and 1,1′-biphenyl-2-carboxamides: Synthesis biological evaluation and mechanism of action. Eur. J. Med. Chem. 2017, 132, 262–273. [Google Scholar] [CrossRef]
- Nikolova, T.; Dvorak, M.; Jung, F.; Adam, I.; Krämer, E.; Gerhold-Ay, A.; Kaina, B. The γH2AX Assay for Genotoxic and Nongenotoxic Agents: Comparison of H2AX Phosphorylation with Cell Death Response. Toxicol. Sci. 2014, 140, 103–117. [Google Scholar] [CrossRef]
- Bonner, W.M.; Redon, C.E.; Dickey, J.S.; Nakamura, A.J.; Sedelnikova, O.A.; Solier, S.; Pommier, Y. γH2AX and cancer. Nat. Rev. Cancer 2008, 8, 957–967. [Google Scholar] [CrossRef]
- Abdullah, A.-S.H.; Mohammed, A.S.; Abdullah, R.; Mirghani, M.E.S.; Al-Qubaisi, M. Cytotoxic effects of Mangifera indica L. kernel extract on human breast cancer (MCF-7 and MDA-MB-231 cell lines) and bioactive constituents in the crude extract. BMC Complement. Altern. Med. 2014, 14, 199. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.X.; Do, T.N.V.; Le, T.H.; Nguyen, M.T.T.; Nguyen, N.T.; Esumi, H.; Awale, S. Chemical Constituents of Mangifera indica and Their Antiausterity Activity against the PANC-1 Human Pancreatic Cancer Cell Line. J. Nat. Prod. 2016, 79, 2053–2059. [Google Scholar] [CrossRef] [PubMed]
- Cernigliaro, C.; D’Anneo, A.; Carlisi, D.; Giuliano, M.; Marino Gammazza, A.; Barone, R.; Longhitano, L.; Cappello, F.; Emanuele, S.; Distefano, A.; et al. Ethanol-Mediated Stress Promotes Autophagic Survival and Aggressiveness of Colon Cancer Cells via Activation of Nrf2/HO-1 Pathway. Cancers 2019, 11, 505. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-C.; Liang, X.; Liu, Z.-Y.; Cui, J.; Liu, Y.; Jing, L.; Jiang, L.-L.; Ma, J.-Q.; Han, L.-L.; Guo, Q.-Q.; et al. In vitro synergistic antitumor efficacy of sequentially combined chemotherapy/icotinib in non-small cell lung cancer cell lines. Oncol. Rep. 2015, 33, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Kelly, K.J.; Sandoval, R.M.; Dunn, K.W.; Molitoris, B.A.; Dagher, P.C. A novel method to determine specificity and sensitivity of the TUNEL reaction in the quantitation of apoptosis. Am. J. Physiol.-Cell Physiol. 2003, 284, C1309–C1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribble, D.; Goldstein, N.B.; Norris, D.A.; Shellman, Y.G. A simple technique for quantifying apoptosis in 96-well plates. BMC Biotechnol. 2005, 5, 12. [Google Scholar] [CrossRef]
- Carlisi, D.; D’Anneo, A.; Martinez, R.; Emanuele, S.; Buttitta, G.; Di Fiore, R.; Vento, R.; Tesoriere, G.; Lauricella, M. The oxygen radicals involved in the toxicity induced by parthenolide in MDA-MB-231 cells. Oncol. Rep. 2014, 32, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Anai, T.; Kosemura, S.; Yamamura, S.; Hasegawa, K. Structure-activity relationship of lepidimoide and its analogues. Phytochemistry 1996, 41, 671–673. [Google Scholar] [CrossRef]
- Gómez-Caravaca, A.M.; López-Cobo, A.; Verardo, V.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC-DAD-q-TOF-MS as a powerful platform for the determination of phenolic and other polar compounds in the edible part of mango and its by-products (peel, seed, and seed husk): Liquid phase separations. ELECTROPHORESIS 2016, 37, 1072–1084. [Google Scholar] [CrossRef]
- Lu, Z.; Nie, G.; Belton, P.S.; Tang, H.; Zhao, B. Structure–activity relationship analysis of antioxidant ability and neuroprotective effect of gallic acid derivatives. Neurochem. Int. 2006, 48, 263–274. [Google Scholar] [CrossRef]
- Da Silva, S.L.; Chaar, J. da S.; Yano, T. Chemotherapeutic potential of two gallic acid derivative compounds from leaves of Casearia sylvestris Sw (Flacourtiaceae). Eur. J. Pharmacol. 2009, 608, 76–83. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, L.C.; Bauermeister, A.; Rezende-Teixeira, P.; dos Santos, E.A.; de Moraes, L.A.B.; Machado-Neto, J.A.; Costa-Lotufo, L.V. Pradimicin-IRD exhibits antineoplastic effects by inducing DNA damage in colon cancer cells. Biochem. Pharmacol. 2019, 168, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Liu, P.; Liu, R.; Wu, X. Dual AO/EB Staining to Detect Apoptosis in Osteosarcoma Cells Compared with Flow Cytometry. Med. Sci. Monit. Basic Res. 2015, 21, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maluf, S.W.; Marroni, N.P.; Heuser, V.D.; Prá, D. DNA Damage and Oxidative Stress in Human Disease. BioMed Res. Int. 2013, 2013, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.-G.; Zhang, K.-J.; Qu, J.-H.; Ren, Y.-D.; Luan, Q. Propofol induces apoptosis of non-small cell lung cancer cells via ERK1/2-dependent upregulation of PUMA. Eur. Rev. Med. Pharmacol. Sci. 2018, 4341–4349. [Google Scholar] [CrossRef]
- Emanuele, S.; D’Anneo, A.; Calvaruso, G.; Cernigliaro, C.; Giuliano, M.; Lauricella, M. The Double-Edged Sword Profile of Redox Signaling: Oxidative Events as Molecular Switches in the Balance between Cell Physiology and Cancer. Chem. Res. Toxicol. 2018, 31, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, W.; Su, Z.; Kong, A.-N.T. The complexity of the Nrf2 pathway: Beyond the antioxidant response. J. Nutr. Biochem. 2015, 26, 1401–1413. [Google Scholar] [CrossRef]
- Kuo, L.J.; Yang, L.-X. Gamma-H2AX—A novel biomarker for DNA double-strand breaks. Vivo Athens Greece 2008, 22, 305–309. [Google Scholar]
- Sluss, H.K.; Davis, R.J. H2AX Is a Target of the JNK Signaling Pathway that Is Required for Apoptotic DNA Fragmentation. Mol. Cell 2006, 23, 152–153. [Google Scholar] [CrossRef]
- Lu, C.; Shi, Y.; Wang, Z.; Song, Z.; Zhu, M.; Cai, Q.; Chen, T. Serum starvation induces H2AX phosphorylation to regulate apoptosis via p38 MAPK pathway. FEBS Lett. 2008, 582, 2703–2708. [Google Scholar] [CrossRef] [Green Version]
- Min, S.; Kim, K.; Kim, S.-G.; Cho, H.; Lee, Y. Chromatin-remodeling factor, RSF1, controls p53-mediated transcription in apoptosis upon DNA strand breaks. Cell Death Dis. 2018, 9, 1079. [Google Scholar] [CrossRef] [PubMed]
- Djelloul, S.; Forgue-Lafitte, M.-E.; Hermelin, B.; Mareel, M.; Bruyneel, E.; Baldi, A.; Giordano, A.; Chastre, E.; Gespach, C. Enterocyte differentiation is compatible with SV40 large T expression and loss of p53 function in human colonic Caco-2 cells: Status of the pRb1 and pRb2 tumor suppressor gene products. FEBS Lett. 1997, 406, 234–242. [Google Scholar] [CrossRef]
- Ray, R.M.; McCormack, S.A.; Johnson, L.R. Polyamine depletion arrests growth of IEC-6 and Caco-2 cells by different mechanisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G37–G43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trainer, D.L.; Kline, T.; McCabe, F.L.; Faucette, L.F.; Feild, J.; Chaikin, M.; Anzano, M.; Rieman, D.; Hoffstein, S.; Li, D.J. Biological characterization and oncogene expression in human colorectal carcinoma cell lines. Int. J. Cancer 1988, 41, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xiong, M.; Xu, C.; Duan, L.; Dong, Y.; Luo, Y.; Niu, T.; Lu, C. Resveratrol induces apoptosis of human chronic myelogenous leukemia cells in vitro through p38 and JNK-regulated H2AX phosphorylation. Acta Pharmacol. Sin. 2015, 36, 353–361. [Google Scholar] [CrossRef] [PubMed]
- D’Anneo, A.; Carlisi, D.; Lauricella, M.; Puleio, R.; Martinez, R.; Di Bella, S.; Di Marco, P.; Emanuele, S.; Di Fiore, R.; Guercio, A.; et al. Parthenolide generates reactive oxygen species and autophagy in MDA-MB231 cells. A soluble parthenolide analogue inhibits tumour growth and metastasis in a xenograft model of breast cancer. Cell Death Dis. 2013, 4, e891. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lu, H.; Bai, Y. Nrf2 in cancers: A double-edged sword. Cancer Med. 2019, 8, 2252–2267. [Google Scholar] [CrossRef]
- Woo, Y.; Oh, J.; Kim, J.-S. Suppression of Nrf2 Activity by Chestnut Leaf Extract Increases Chemosensitivity of Breast Cancer Stem Cells to Paclitaxel. Nutrients 2017, 9, 760. [Google Scholar] [CrossRef]
- Pall, M.L.; Levine, S. Nrf2, a master regulator of detoxification and also antioxidant, anti-inflammatory and other cytoprotective mechanisms, is raised by health promoting factors. Sheng Li Xue Bao 2015, 67, 1–18. [Google Scholar] [CrossRef]
- Xu, Y.; Ren, H.; Liu, J.; Wang, Y.; Meng, Z.; He, Z.; Miao, W.; Chen, G.; Li, X. A switchable NO-releasing nanomedicine for enhanced cancer therapy and inhibition of metastasis. Nanoscale 2019, 11, 5474–5488. [Google Scholar] [CrossRef]
- Yang, H.; Bai, W.; Gao, L.; Jiang, J.; Tang, Y.; Niu, Y.; Lin, H.; Li, L. Mangiferin alleviates hypertension induced by hyperuricemia via increasing nitric oxide releases. J. Pharmacol. Sci. 2018, 137, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Paskas, S.; Mazzon, E.; Basile, M.S.; Cavalli, E.; Al-Abed, Y.; He, M.; Rakocevic, S.; Nicoletti, F.; Mijatovic, S.; Maksimovic-Ivanic, D. Lopinavir-NO, a nitric oxide-releasing HIV protease inhibitor, suppresses the growth of melanoma cells in vitro and in vivo. Invest. New Drugs 2019, 37, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Basile, M.; Mazzon, E.; Krajnovic, T.; Draca, D.; Cavalli, E.; Al-Abed, Y.; Bramanti, P.; Nicoletti, F.; Mijatovic, S.; Maksimovic-Ivanic, D. Anticancer and Differentiation Properties of the Nitric Oxide Derivative of Lopinavir in Human Glioblastoma Cells. Molecules 2018, 23, 2463. [Google Scholar] [CrossRef] [PubMed]
- Maksimovic-Ivanic, D.; Mojic, M.; Bulatovic, M.; Radojkovic, M.; Kuzmanovic, M.; Ristic, S.; Stosic-Grujicic, S.; Miljkovic, D.; Cavalli, E.; Libra, M.; et al. The NO-modified HIV protease inhibitor as a valuable drug for hematological malignancies: Role of p70S6K. Leuk. Res. 2015, 39, 1088–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seabra, A.B.; Durán, N. Nitric oxide donors for prostate and bladder cancers: Current state and challenges. Eur. J. Pharmacol. 2018, 826, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Paull, T.T.; Rogakou, E.P.; Yamazaki, V.; Kirchgessner, C.U.; Gellert, M.; Bonner, W.M. A critical role for histone H2AX in recruitment of repair factors to nuclear foci after DNA damage. Curr. Biol. CB 2000, 10, 886–895. [Google Scholar] [CrossRef] [Green Version]
- Rogakou, E.P.; Nieves-Neira, W.; Boon, C.; Pommier, Y.; Bonner, W.M. Initiation of DNA fragmentation during apoptosis induces phosphorylation of H2AX histone at serine 139. J. Biol. Chem. 2000, 275, 9390–9395. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.-G.; Shimizu, T.; Pommier, Y. 7-Hydroxystaurosporine (UCN-01) Induces Apoptosis in Human Colon Carcinoma and Leukemia Cells Independently of p53. Exp. Cell Res. 1997, 234, 388–397. [Google Scholar] [CrossRef]
- Solier, S.; Pommier, Y. The nuclear γ-H2AX apoptotic ring: Implications for cancers and autoimmune diseases. Cell. Mol. Life Sci. 2014, 71, 2289–2297. [Google Scholar] [CrossRef]
- Sekhar, S.; Venkatesh, J.; Cheriyan, V.; Muthu, M.; Levi, E.; Assad, H.; Meister, P.; Undyala, V.; Gauld, J.; Rishi, A. A H2AX–CARP-1 Interaction Regulates Apoptosis Signaling Following DNA Damage. Cancers 2019, 11, 221. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | RT (min) | ESI− [M − H]− (m/z) | Molecular Formula | mg/100 g | ||
|---|---|---|---|---|---|---|
| Teor. | Exp. | |||||
| 1 | Disaccaride | 6.22 | 341.1089 [M − H]− 377.0856 [M + Cl]− 387.1144 [M + FA]− | 341.1090 [M − H]− 377.0857 [M + Cl]− 387.1145 [M + FA]− | C12H22O11 | ND a |
| 2 | Quinic acid | 6.70 | 191.0561 [M − H]− | 191.0562 [M − H]− | C7H12O6 | ND a |
| 3 | Glucosyl gallate | 7.00 | 331.0671 [M − H]− | 331.0673 [M − H]− | C13H16O10 | 108.14 |
| 4 | Gluconolactone | 7.53 | 223.0459 [M + FA]− | 223.0460 [M + FA]− | C6H10O6 | ND a |
| 5 | Lepidimoic Acid | 7.70 | 965.2627 [3M − H]− | 965.2614 [3M − H]− | C36H54O30 | ND a |
| 6 | Citric acid | 8.39 | 191.0197 [M − H]− | 191.0198 [M − H]− | C6H8O7 | ND a |
| 7 | Gallic acid | 8.63 | 169.0142 [M − H]− | 169.0141 [M − H]− | C7H6O5 | 118.57 |
| 8 | Maclurin mono-O-galloyl-glucoside | 24.74 | 575.1042 [M − H]− | 575.1041 [M − H]− | C26H24O15 | 72.03 |
| 9 | Mangiferin | 25.01 | 421.0776 [M − H]− | 421.0776 [M − H]− | C19H18O11 | 2.81 |
| 10 | Maclurin di-O-galloyl glucoside | 25.34 | 727.1152 [M − H]− | 727.1145 [M − H]− | C33H28O19 | 20.21 |
| 11 | Digallic acid | 25.46 | 321.0252 [M − H]− | 321.0255 [M − H]− | C14H10O9 | Trace |
| 12 | Maclurin tri-O-galloyl-glucoside | 26.21 | 879.1262 [M − H]− | 879.1253 [M − H]− | C40H32O23 | 6.05 |
| 13 | Tetragalloyl glucose | 26.31 | 787.0999 [M − H]− | 787.0998 [M − H]− | C34H28O22 | 4.88 |
| 14 | Methylgallate | 26.36 | 183.0299 [M − H]− | 183.0301 [M − H]− | C8H8O5 | 225.87 |
| 15 | Pentagalloyl glucose | 26.73 | 939.1109 [M − H]− | 939.1100 [M − H]− | C41H32O26 | 17.89 |
| 16 | Methyl-digallate ester | 29.68 | 335.0409 [M − H]− | 335.0411 [M − H]− | C15H12O9 | 487.15 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauricella, M.; Lo Galbo, V.; Cernigliaro, C.; Maggio, A.; Palumbo Piccionello, A.; Calvaruso, G.; Carlisi, D.; Emanuele, S.; Giuliano, M.; D’Anneo, A. The Anti-Cancer Effect of Mangifera indica L. Peel Extract is Associated to γH2AX-mediated Apoptosis in Colon Cancer Cells. Antioxidants 2019, 8, 422. https://doi.org/10.3390/antiox8100422
Lauricella M, Lo Galbo V, Cernigliaro C, Maggio A, Palumbo Piccionello A, Calvaruso G, Carlisi D, Emanuele S, Giuliano M, D’Anneo A. The Anti-Cancer Effect of Mangifera indica L. Peel Extract is Associated to γH2AX-mediated Apoptosis in Colon Cancer Cells. Antioxidants. 2019; 8(10):422. https://doi.org/10.3390/antiox8100422
Chicago/Turabian StyleLauricella, Marianna, Valentina Lo Galbo, Cesare Cernigliaro, Antonella Maggio, Antonio Palumbo Piccionello, Giuseppe Calvaruso, Daniela Carlisi, Sonia Emanuele, Michela Giuliano, and Antonella D’Anneo. 2019. "The Anti-Cancer Effect of Mangifera indica L. Peel Extract is Associated to γH2AX-mediated Apoptosis in Colon Cancer Cells" Antioxidants 8, no. 10: 422. https://doi.org/10.3390/antiox8100422